
Cracking the Kinase Code: Urinary Biomarkers as Early Alarms for AAA Rupture—A Pilot Study


Abstract
1. Introduction
2. Materials and Methods
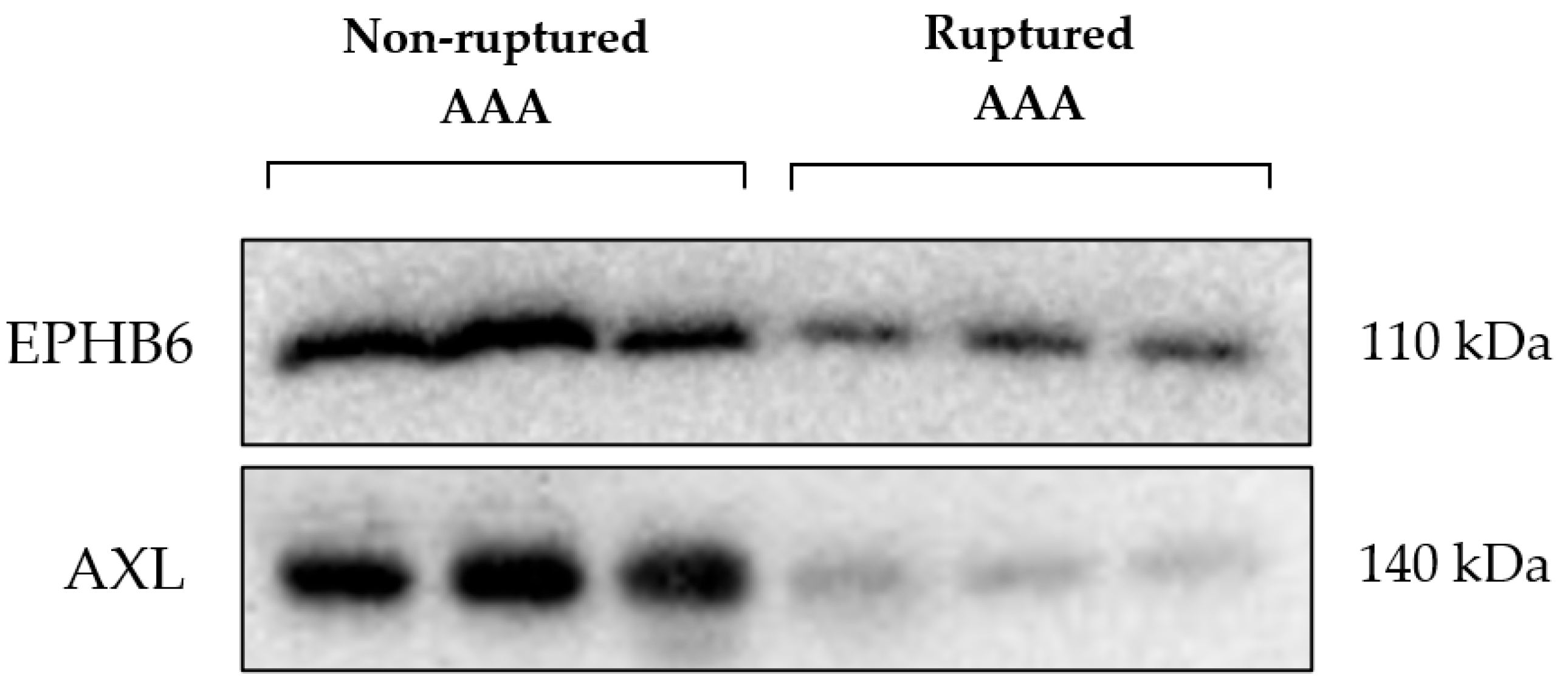
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AAA | Abdominal aortic aneurysms |
| RAAA | Ruptured abdominal aortic aneurysm |
| MIBs | Multiplexed kinase inhibitor-conjugated beads |
| MIB/MS | Quantitative mass spectrometry |
| NSAID | nonsteroidal anti-inflammatory drugs |
| LFQ | label-free quantification |
| TK | Tyrosine kinase |
| NME1 | Nucleoside diphosphate kinase A |
| MAP2K1 | Dual specificity mitogen-activated protein kinase 1 |
| CSNK1A1 | Casein kinase I isoform alpha |
| LYN | Tyrosine-protein kinase Lyn |
| ILK | Integrin-linked protein kinase |
| CDC42BPB | Serine/threonine-protein kinase MRCK beta |
| SLK | STE20-like serine/threonine-protein kinase |
| EPHB6 | Ephrin type-B receptor 6 |
| AXL | Tyrosine-protein kinase receptor UFO |
| EPHB4 | Ephrin type-B receptor 4 |
| DDR1 | Epithelial discoidin domain-containing receptor 1 |
| EPHA2 | Ephrin type-A receptor 2 |
| EPHB3 | Ephrin type-B receptor 3 |
| LYN | Tyrosine-protein kinase Lyn |
| VSMC | vascular smooth muscle cell |
| AAAD | Aortic aneurysms and dissections |
| PDGFRβ | platelet-derived growth factor receptor β |
| MerTK | MER proto-oncogene tyrosine kinase |
References
- Kent, K.C. Clinical practice. Abdominal aortic aneurysms. N. Engl. J. Med. 2014, 371, 2101–2108. [Google Scholar] [CrossRef] [PubMed]
- Chaikof, E.L.; Dalman, R.L.; Eskandari, M.K.; Jackson, B.M.; Lee, W.A.; Mansour, M.A.; Mastracci, T.M.; Mell, M.; Murad, M.H.; Nguyen, L.L.; et al. The Society for Vascular Surgery practice guidelines on the care of patients with an abdominal aortic aneurysm. J. Vasc. Surg. 2018, 67, 2–77.e72. [Google Scholar] [CrossRef]
- Spanos, K.; Eckstein, H.H.; Giannoukas, A.D. Small Abdominal Aortic Aneurysms Are Not All the Same. Angiology 2020, 71, 205–207. [Google Scholar] [CrossRef]
- Olson, S.L.; Wijesinha, M.A.; Panthofer, A.M.; Blackwelder, W.C.; Upchurch, G.R., Jr.; Terrin, M.L.; Curci, J.A.; Baxter, B.T.; Matsumura, J.S. Evaluating Growth Patterns of Abdominal Aortic Aneurysm Diameter With Serial Computed Tomography Surveillance. JAMA Surg. 2021, 156, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.; Abu-Raisi, M.; Feasson, M.; Shaikh, F.; Saposnik, G.; Mamdani, M.; Qadura, M. Current Prognostic Biomarkers for Abdominal Aortic Aneurysm: A Comprehensive Scoping Review of the Literature. Biomolecules 2024, 14, 661. [Google Scholar] [CrossRef]
- Xue, C.; Yang, B.; Fu, L.; Hou, H.; Qiang, J.; Zhou, C.; Gao, Y.; Mao, Z. Urine biomarkers can outperform serum biomarkers in certain diseases. Urine 2023, 5, 57–64. [Google Scholar] [CrossRef]
- Mateos-Cáceres, P.J.; García-Méndez, A.; López Farré, A.; Macaya, C.; Núñez, A.; Gómez, J.; Alonso-Orgaz, S.; Carrasco, C.; Burgos, M.E.; de Andrés, R.; et al. Proteomic analysis of plasma from patients during an acute coronary syndrome. J. Am. Coll. Cardiol. 2004, 44, 1578–1583. [Google Scholar] [CrossRef]
- Loscalzo, J. Proteomics in cardiovascular biology and medicine. Circulation 2003, 108, 380–383. [Google Scholar] [CrossRef]
- Zhou, Y.W.; Xu, P.C.; Cheng, Y. Basic pathogenesis of asthenia of healthy energy and blood stasis in liver cirrhosis studied by serum proteomics. Zhongguo Zhong Xi Yi Jie He Za Zhi 2011, 31, 595–602. [Google Scholar]
- Arrell, D.K.; Neverova, I.; Van Eyk, J.E. Cardiovascular proteomics: Evolution and potential. Circ. Res. 2001, 88, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.J.; Cox, N.J.; Zimmerman, E.I.; Dewar, B.J.; Duncan, J.S.; Whittle, M.C.; Nguyen, T.A.; Jones, L.S.; Ghose Roy, S.; Smalley, D.M.; et al. Application of multiplexed kinase inhibitor beads to study kinome adaptations in drug-resistant leukemia. PLoS ONE 2013, 8, e66755. [Google Scholar] [CrossRef] [PubMed]
- Unwin, R.D.; Griffiths, J.R.; Whetton, A.D. Simultaneous analysis of relative protein expression levels across multiple samples using iTRAQ isobaric tags with 2D nano LC-MS/MS. Nat. Protoc. 2010, 5, 1574–1582. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Luo, W.; Brouwer, C. Pathview: An R/Bioconductor package for pathway-based data integration and visualization. Bioinformatics 2013, 29, 1830–1831. [Google Scholar] [CrossRef] [PubMed]
- Carbon, S.; Ireland, A.; Mungall, C.J.; Shu, S.; Marshall, B.; Lewis, S.; AmiGO Hub; Web Presence Working Group. AmiGO: Online access to ontology and annotation data. Bioinformatics 2008, 25, 288–289. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- The Gene Ontology Consortium; Aleksander, S.A.; Balhoff, J.; Carbon, S.; Cherry, J.M.; Drabkin, H.J.; Ebert, D.; Feuermann, M.; Gaudet, P.; Harris, N.L.; et al. The Gene Ontology knowledgebase in 2023. Genetics 2023, 224, iyad031. [Google Scholar] [CrossRef]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef]
- Chiang, M.-T.; Chen, I.-M.; Hsu, F.-F.; Chen, Y.-H.; Tsai, M.-S.; Hsu, Y.-W.; Leu, H.-B.; Huang, P.-H.; Chen, J.-W.; Liu, F.-T.; et al. Gal-1 (Galectin-1) Upregulation Contributes to Abdominal Aortic Aneurysm Progression by Enhancing Vascular Inflammation. Arterioscler. Thromb. Vasc. Biol. 2021, 41, 331–345. [Google Scholar] [CrossRef]
- Raffort, J.; Lareyre, F.; Clément, M.; Hassen-Khodja, R.; Chinetti, G.; Mallat, Z. Monocytes and macrophages in abdominal aortic aneurysm. Nat. Rev. Cardiol. 2017, 14, 457–471. [Google Scholar] [CrossRef]
- Choke, E.; Cockerill, G.W.; Dawson, J.; Wilson, R.W.; Jones, A.; Loftus, I.M.; Thompson, M.M. Increased angiogenesis at the site of abdominal aortic aneurysm rupture. Ann. N. Y. Acad. Sci. 2006, 1085, 315–319. [Google Scholar] [CrossRef]
- Kugo, H.; Zaima, N.; Tanaka, H.; Mouri, Y.; Yanagimoto, K.; Hayamizu, K.; Hashimoto, K.; Sasaki, T.; Sano, M.; Yata, T.; et al. Adipocyte in vascular wall can induce the rupture of abdominal aortic aneurysm. Sci. Rep. 2016, 6, 31268. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.M.; Jeong, K.; Lim, S.S. FAK Family Kinases in Vascular Diseases. Int. J. Mol. Sci. 2020, 21, 3630. [Google Scholar] [CrossRef]
- Wang, J.; Zhuang, S. Src family kinases in chronic kidney disease. Am. J. Physiol. Ren. Physiol. 2017, 313, F721–F728. [Google Scholar] [CrossRef]
- Yin, Z.; Zou, Y.; Wang, D.; Huang, X.; Xiong, S.; Cao, L.; Zhang, Y.; Sun, Y.; Zhang, N. Regulation of the Tec family of non-receptor tyrosine kinases in cardiovascular disease. Cell Death Discov. 2022, 8, 119. [Google Scholar] [CrossRef] [PubMed]
- Rombouts, K.B.; van Merrienboer, T.A.R.; Henneman, A.A.; Knol, J.C.; Pham, T.V.; Piersma, S.R.; Jimenez, C.R.; Bogunovic, N.; van der Velden, J.; Yeung, K.K. Insight in the (Phospho)proteome of Vascular Smooth Muscle Cells Derived From Patients With Abdominal Aortic Aneurysm Reveals Novel Disease Mechanisms. Arterioscler. Thromb. Vasc. Biol. 2024, 44, 2226–2243. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Kong, W. Cellular signaling in Abdominal Aortic Aneurysm. Cell. Signal. 2020, 70, 109575. [Google Scholar] [CrossRef]
- Li, S.; Liu, G.; Cheng, S.; Li, X.; Weng, X.; Yang, J. Pharmacological and genetic inhibition of BTK ameliorates vascular degeneration, dissection, and rupture. Life Sci. 2025, 369, 123533. [Google Scholar] [CrossRef]
- Liu, S.; Wu, J.; Banerjee, O.; Xue, B.; Shi, H.; Ding, Z. Big data analytics and scRNA-seq in human aortic aneurysms and dissections: Role of endothelial MerTK. Theranostics 2025, 15, 202–215. [Google Scholar] [CrossRef]
- Nakayama, A.; Nakayama, M.; Turner, C.J.; Höing, S.; Lepore, J.J.; Adams, R.H. Ephrin-B2 controls PDGFRβ internalization and signaling. Genes Dev. 2013, 27, 2576–2589. [Google Scholar] [CrossRef]
- Oike, Y.; Ito, Y.; Hamada, K.; Zhang, X.Q.; Miyata, K.; Arai, F.; Inada, T.; Araki, K.; Nakagata, N.; Takeya, M.; et al. Regulation of vasculogenesis and angiogenesis by EphB/ephrin-B2 signaling between endothelial cells and surrounding mesenchymal cells. Blood 2002, 100, 1326–1333. [Google Scholar] [CrossRef]
- Liu, H.; Devraj, K.; Möller, K.; Liebner, S.; Hecker, M.; Korff, T. EphrinB-mediated reverse signalling controls junctional integrity and pro-inflammatory differentiation of endothelial cells. Thromb. Haemost. 2014, 112, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Zalewski, D.; Chmiel, P.; Kołodziej, P.; Borowski, G.; Feldo, M.; Kocki, J.; Bogucka-Kocka, A. Dysregulations of Key Regulators of Angiogenesis and Inflammation in Abdominal Aortic Aneurysm. Int. J. Mol. Sci. 2023, 24, 12087. [Google Scholar] [CrossRef] [PubMed]
- Coulthard, M.G.; Morgan, M.; Woodruff, T.M.; Arumugam, T.V.; Taylor, S.M.; Carpenter, T.C.; Lackmann, M.; Boyd, A.W. Eph/Ephrin signaling in injury and inflammation. Am. J. Pathol. 2012, 181, 1493–1503. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Hu, T.; Lu, W.; Yu, Y.; Xue, S.; Wu, K.; Liu, Y.; Lin, J.; Bai, H.; Yun, Z.; et al. Morphology and biomechanical index predict the rupture location and rupture risk of abdominal aortic aneurysm. Sci. Rep. 2025, 15, 9604. [Google Scholar] [CrossRef]
- Polzer, S.; Gasser, T.C.; Vlachovský, R.; Kubíček, L.; Lambert, L.; Man, V.; Novák, K.; Slažanský, M.; Burša, J.; Staffa, R. Biomechanical indices are more sensitive than diameter in predicting rupture of asymptomatic abdominal aortic aneurysms. J. Vasc. Surg. 2020, 71, 617–626.e616. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Non-Ruptured AAA | Ruptured AAA | Total 1 | ||
|---|---|---|---|---|
| Number of patients | 5 | 5 | 10 | |
| Gender: males/females | 2/3 | 2/3 | 4/6 | |
| Age (years) | Median | 71.6 (68–75) | 75.4 (68–83) | 73.5 (68–83) |
| (Range) | ||||
| Aneurysm diameter (mm) | Median | 56.2 (50–72) | 57 (49–90) | 56.6 (50–90) |
| (Range) | ||||
| Body mass index | 28.2 | 29.1 | 28.5 | |
| Smoking status (smokers/nonsmokers) | 3/2 | 3/2 | 6/4 | |
| NSAID (Yes/No) | 4/1 | 2/3 | 6/4 | |
| Statins (Yes/No) | 5/0 | 5/0 | 10/0 | |
| Outcome (Alive/Death) | 5/0 | 4/1 | 9/1 | |
| Gene | Family | Ruptured AAA FFQ Mean (Range) | Non-Ruptured AAA LFQ Mean (Range) | p-Value | Majority Protein IDs |
|---|---|---|---|---|---|
| EPHB6 1 | TK | 21.73 | 26.44 | 0.0001 | O15197 |
| (20.99–22.47) | (25.42–27.46) | ||||
| AXL 2 | TK | 22.73 | 31.1 | 0.0001 | P30530 |
| (21.88–23.58) | (29.33–32.87) | ||||
| EPHB4 3 | TK | 24.16 | 28.84 | 0.018 | P54760 |
| (23.31–25.02) | (28.55–29.14) | ||||
| DDR1 4 | TK | 24.16 | 27.73 | 0.031 | Q08345 |
| (22.35–25.98) | (27.11–28.35) | ||||
| EPHA2 5 | TK | 23.53 | 28.02 | 0.013 | P29317 |
| (21.67–25.39) | (26.99–29.06) | ||||
| EPHB3 6 | TK | 22.81 | 26.73 | 0.040 | P54753 |
| (21.87–23.76) | (26.14–27.32) |
| Gene | Family | Ruptured AAA FFQ Mean (Range) | Non-Ruptured AAA LFQ Mean (Range) | p-Value | Majority Protein IDs |
|---|---|---|---|---|---|
| NME1 1 | Metabolic | 23.02 | 25.37 | 0.055 | P15531 |
| (22.69–23.36) | (24.64–26.11) | ||||
| MAP2K1 2 | STE | 24.24 | 25.99 | 0.060 | Q02750 |
| (23.48–25.01 | (25.56–26.43) | ||||
| CSNK1A1 3 | CK1 | 21.90 | 24.01 | 0.066 | P48729 |
| (21.51–22.30) | (22.45–25.57) | ||||
| LYN 4 | TK | 22.04 | 24.54 | 0.071 | P07948 |
| (21.75–22.34) | (22.84–26.25) | ||||
| ILK 5 | TKL | 22.31 | 23.87 | 0.082 | Q13418 |
| (21.72–22.90) | (22.52–25.22) | ||||
| CDC42BPB 6 | AGC | 22.82 | 25.87 | 0.086 | Q9Y5S2 |
| (22.44–23.20) | (25.48–26.27) | ||||
| SLK 7 | STE | 26.79 | 29.46 | 0.059 | Q9H2G2 |
| (25.72–27.86) | (28.59–30.34) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Östling, E.M.; Baltrunas, T.; Grootenboer, N.; Urbonavicius, S. Cracking the Kinase Code: Urinary Biomarkers as Early Alarms for AAA Rupture—A Pilot Study. J. Clin. Med. 2025, 14, 3845. https://doi.org/10.3390/jcm14113845
Östling EM, Baltrunas T, Grootenboer N, Urbonavicius S. Cracking the Kinase Code: Urinary Biomarkers as Early Alarms for AAA Rupture—A Pilot Study. Journal of Clinical Medicine. 2025; 14(11):3845. https://doi.org/10.3390/jcm14113845
Chicago/Turabian StyleÖstling, Emma Maria, Tomas Baltrunas, Nathalie Grootenboer, and Sigitas Urbonavicius. 2025. "Cracking the Kinase Code: Urinary Biomarkers as Early Alarms for AAA Rupture—A Pilot Study" Journal of Clinical Medicine 14, no. 11: 3845. https://doi.org/10.3390/jcm14113845
APA StyleÖstling, E. M., Baltrunas, T., Grootenboer, N., & Urbonavicius, S. (2025). Cracking the Kinase Code: Urinary Biomarkers as Early Alarms for AAA Rupture—A Pilot Study. Journal of Clinical Medicine, 14(11), 3845. https://doi.org/10.3390/jcm14113845