
The Effects of a High Carbohydrate Diet Combined with High Molecular Weight Carbohydrate Supplementation on Anaerobic Performance and Oxidative Stress in Elite Swimmers


Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
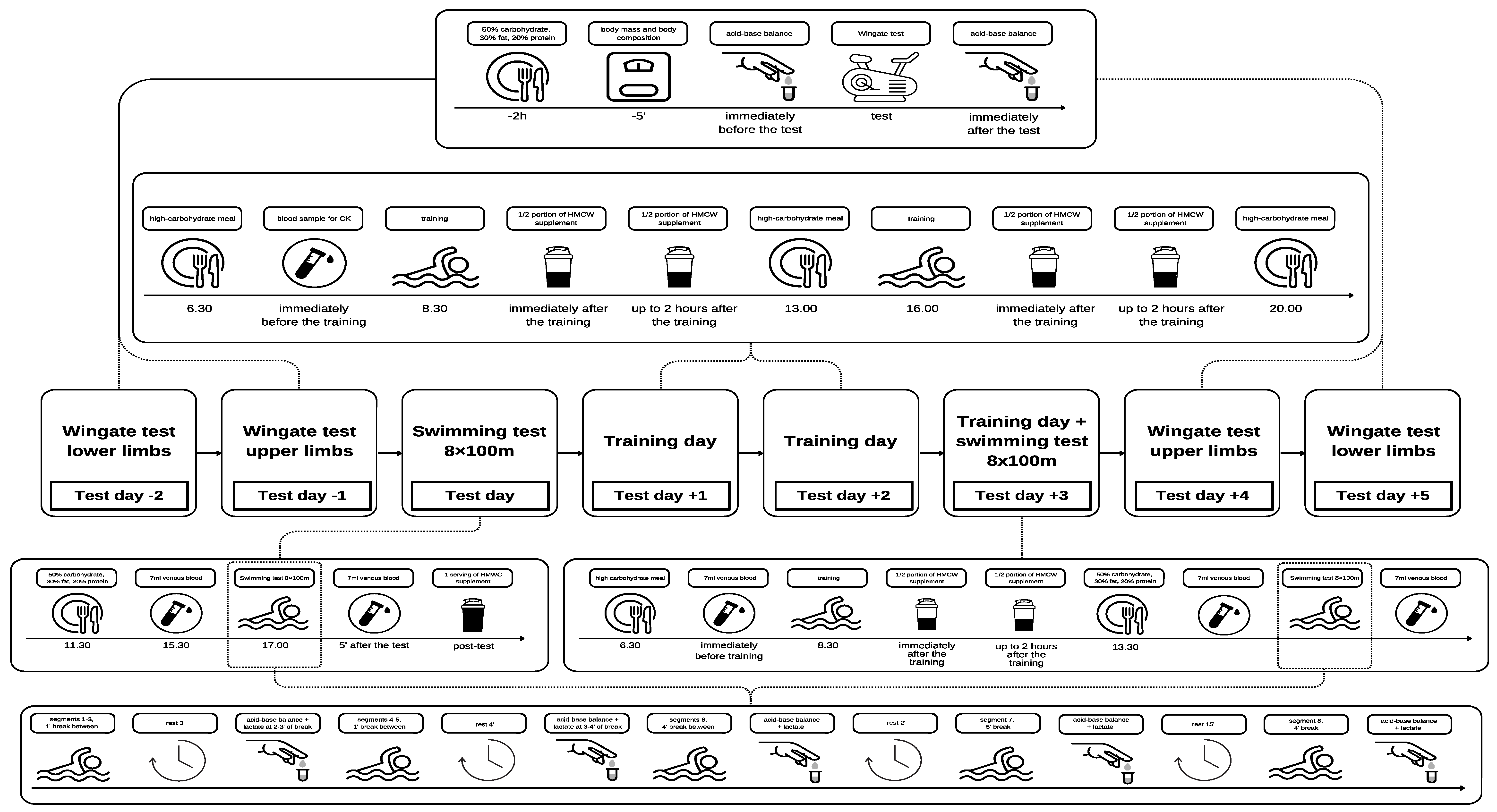
2.2. Experimental Design
2.3. Experimental Protocol
2.3.1. Wingate Test
2.3.2. Swimming Test
- 3 × 100 m at 75% max (1 min, then 3 min rest interval; blood sampling at 2–3 min);
- 2 × 100 m at 85% max (1 min and 4 min rest interval; blood sampling at 3–4 min);
- 1 × 100 m at 90% max (6 min rest interval; blood sampling at 4–5 min post-exercise);
- 1 × 100 m at 95% max (20 min rest interval; blood sampling at 5–6 min post-exercise);
- 1 × 100 m at 100% max (blood samples drawn at 4 min post-exercise).
2.4. Nutrition and Supplementation
2.5. Biochemical Analysis
2.6. Statistical Analysis
2.7. Limitations
3. Results
4. Discussion
5. Conclusions
6. Practical Implications
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bompa, T.O.; Buzzichelli, C. Periodization—Theory and Methodology of Training; Human Kinetics: Champaign, IL, USA, 2018. [Google Scholar]
- Hultman, E.; Greenhaff, P.L. Skeletal muscle energy metabolism and fatigue during intense exercise in man. Sci. Prog. 1991, 75, 361–370. [Google Scholar] [PubMed]
- Vigh-Larsen, J.F.; Ørtenblad, N.; Spriet, L.L.; Overgaard, K.; Mohr, M. Muscle glycogen metabolism and high-intensity exercise performance: A narrative review. Sports Med. 2021, 51, 1855–1874. [Google Scholar] [CrossRef]
- Issurin, V.B. Benefits and limitations of block periodized training approaches to athletes’ preparation: A review. Sports Med. 2016, 46, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Marquet, L.-A.; Brisswalter, J.; Louis, J.; Tiollier, E.; Burke, L.; Hawley, J.; Hausswirth, C. Enhanced Endurance Performance by Periodization of CHO Intake:” sleep low” strategy. Med. Sci. Sports Exerc. 2016, 48, 663–672. [Google Scholar] [CrossRef]
- Mujika, I.; Halson, S.; Burke, L.M.; Balagué, G.; Farrow, D. An integrated, multifactorial approach to periodization for optimal performance in individual and team sports. Int. J. Sports Physiol. Perform. 2018, 13, 538–561. [Google Scholar] [CrossRef]
- Burke, L.M.; Van Loon, L.J.C.; Hawley, J.A. Postexercise muscle glycogen resynthesis in humans. J. Appl. Physiol. 2017, 122, 1055–1067. [Google Scholar] [CrossRef]
- Hingst, J.R.; Bruhn, L.; Hansen, M.B.; Rosschou, M.F.; Birk, J.B.; Fentz, J.; Foretz, M.; Viollet, B.; Sakamoto, K.; Færgeman, N.J. Exercise-induced molecular mechanisms promoting glycogen supercompensation in human skeletal muscle. Mol. Metab. 2018, 16, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Peeling, P.; Binnie, M.J.; Goods, P.S.R.; Sim, M.; Burke, L.M. Evidence-based supplements for the enhancement of athletic performance. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 178–187. [Google Scholar] [CrossRef]
- Rothschild, J.A.; Bishop, D.J. Effects of dietary supplements on adaptations to endurance training. Sports Med. 2020, 50, 25–53. [Google Scholar] [CrossRef]
- Vigh-Larsen, J.F.; Ørtenblad, N.; Nielsen, J.; Emil Andersen, O.L.E.; Overgaard, K.; Mohr, M. The Role of Muscle Glycogen Content and Localization in High-Intensity Exercise Performance: A Placebo-Controlled Trial. Med. Sci. Sports Exerc. 2022, 54, 2073–2086. [Google Scholar] [CrossRef]
- Areta, J.L.; Hopkins, W.G. Skeletal Muscle Glycogen Content at Rest and During Endurance Exercise in Humans: A Meta-Analysis. Sports Med. 2018, 48, 2091–2102. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.L.; Heigenhauser, G.J.F.; Hultman, E.; Spriet, L.L. Dietary carbohydrate, muscle glycogen content, and endurance performance in well-trained women. J. Appl. Physiol. 2000, 88, 2151–2158. [Google Scholar] [CrossRef]
- Hargreaves, M.; Spriet, L.L. Skeletal muscle energy metabolism during exercise. Nat. Metab. 2020, 2, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.; Figueiredo, P.; Sousa, A.; Monteiro, J.; Pelarigo, J.; Vilas-Boas, J.P.; Toussaint, H.M.; Fernandes, R.F. VO2 kinetics and metabolic contributions during full and upper body extreme swimming intensity. Eur. J. Appl. Physiol. 2015, 115, 1117–1124. [Google Scholar] [CrossRef]
- Zamparo, P.; Capelli, C.; Cautero, M.; Di Nino, A. Energy cost of front-crawl swimming at supra-maximal speeds and underwater torque in young swimmers. Eur. J. Appl. Physiol. 2000, 83, 487–491. [Google Scholar] [CrossRef]
- Bestard, M.A.; Rothschild, J.A.; Crocker, G.H. Effect of low- and high-carbohydrate diets on swimming economy: A crossover study. J. Int. Soc. Sports Nutr. 2020, 17, 64. [Google Scholar] [CrossRef]
- Crowley, E.; Harrison, A.J.; Lyons, M. Dry-Land Resistance Training Practices of Elite Swimming Strength and Conditioning Coaches. J. Strength Cond. Res. 2018, 32, 2592–2600. [Google Scholar] [CrossRef]
- Muniz-Pardos, B.; Gomez-Bruton, A.; Matute-Llorente, A.; Gonzalez-Aguero, A.; Gomez-Cabello, A.; Gonzalo-Skok, O.; Casajus, J.A.; Vicente-Rodriguez, G. Swim-Specific Resistance Training: A Systematic Review. J. Strength Cond. Res. 2019, 33, 2875–2881. [Google Scholar] [CrossRef] [PubMed]
- Gorostiaga, E.M.; Navarro-Amézqueta, I.; Cusso, R.; Hellsten, Y.; Calbet, J.A.L.; Guerrero, M.; Granados, C.; González-Izal, M.; Ibáñez, J.; Izquierdo, M. Anaerobic energy expenditure and mechanical efficiency during exhaustive leg press exercise. PLoS ONE 2010, 5, e13486. [Google Scholar] [CrossRef]
- Piehl Aulin, K.; Söderlund, K.; Hultman, E. Muscle glycogen resynthesis rate in humans after supplementation of drinks containing carbohydrates with low and high molecular masses. Eur. J. Appl. Physiol. 2000, 81, 346–351. [Google Scholar] [CrossRef]
- Leiper, J.B.; Aulin, K.P.; Söderlund, K. Improved gastric emptying rate in humans of a unique glucose polymer with gel-forming properties. Scand. J. Gastroenterol. 2000, 35, 1143–1149. [Google Scholar] [PubMed]
- Cabrera Gomez, C.-M.; Viña, J.; Ji, L.L. Interplay of oxidants and antioxidants during exercise: Implications for muscle health. Physician Sportsmed. 2009, 37, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.J.; Yamada, T.; Rassier, D.E.; Andersson, D.C.; Westerblad, H.; Lanner, J.T. Reactive oxygen/nitrogen species and contractile function in skeletal muscle during fatigue and recovery. J. Physiol. 2016, 594, 5149–5160. [Google Scholar] [CrossRef]
- Pereira, B.; Curi, R.; Kokubun, E.; Bechara, E.J. 5-Aminolevulinic acid-induced alterations of oxidative metabolism in sedentary and exercise-trained rats. J. Appl. Physiol. 1992, 72, 226–230. [Google Scholar] [CrossRef]
- Kaczor, J.J.; Robertshaw, H.A.; Tarnopolsky, M.A. Higher oxidative stress in skeletal muscle of McArdle disease patients. Mol. Genet. Metab. Rep. 2017, 12, 69–75. [Google Scholar] [CrossRef]
- de Mello, A.S.; Frusciante, M.R.; Kneib, L.G.; Bortolato, G.; Cé, J. A-glucosidase deficiency promotes increasing protein oxidative damage in Pompe disease patients. J. Alzheimer’s Dis. Park. 2017, 7, 2161–2460. [Google Scholar]
- Bloomer, R.J.; Goldfarb, A.H. Anaerobic exercise and oxidative stress: A review. Can. J. Appl. Physiol. 2004, 29, 245–263. [Google Scholar] [CrossRef] [PubMed]
- Thirupathi, A.; Wang, M.; Lin, J.K.; Fekete, G.; István, B.; Baker, J.S.; Gu, Y. Effect of different exercise modalities on oxidative stress: A systematic review. BioMed Res. Int. 2021, 2021, 1947928. [Google Scholar] [CrossRef]
- Maglischo, E.W. Swimming Fastest; Human Kinetics: Champaign, IL, USA, 2003. [Google Scholar]
- Beutler, E.; Duron, O.; Kelly, B.M. Improved method for the determination of blood glutathione. J. Lab. Clin. Med. 1963, 61, 882–888. [Google Scholar]
- Aebi, H. Catalase in vitro. Methods Enzimol. 1984, 105, 121–126. [Google Scholar]
- Glatzle, D.; Körner, W.F.; Christeller, S.; Wiss, O. Method for the detection of a biochemical riboflavin deficiency. Stimulation of NADPH2-dependent glutathione reductase from human erythrocytes by FAD in vitro. Investigations on the vitamin B2 status in healthly people and geriatric patients. Int. J. Vitam. Res. 1970, 40, 166–183. [Google Scholar]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- ISO 9001:2015; Quality Management Systems—Requirements. (Certificate No. PW-19912-18B); Polish Committee for Standardization: Warsaw, Poland, 2015.
- Burke, L.M.; Kiens, B.; Ivy, J.L. Carbohydrates and fat for training and recovery. In Food, Nutrition and Sports Performance II; Routledge: London, UK, 2004; pp. 24–49. [Google Scholar]
- Jensen, T.E.; Richter, E.A. Regulation of glucose and glycogen metabolism during and after exercise. J. Physiol. 2012, 590, 1069–1076. [Google Scholar] [CrossRef]
- Issurin, V. Block periodization versus traditional training theory: A review. J. Sports Med. Phys. Fit. 2008, 48, 65–75. [Google Scholar]
- Dingley, A.A.; Pyne, D.B.; Youngson, J.; Burkett, B. Effectiveness of a dry-land resistance training program on strength, power, and swimming performance in paralympic swimmers. J. Strength Cond. Res. 2015, 29, 619–626. [Google Scholar] [CrossRef]
- Mata, F.; Valenzuela, P.L.; Gimenez, J.; Tur, C.; Ferreria, D.; Domínguez, R.; Sanchez-Oliver, A.J.; Sanz, J.M.M. Carbohydrate availability and physical performance: Physiological overview and practical recommendations. Nutrients 2019, 11, 1084. [Google Scholar] [CrossRef]
- McGlory, C.; Morton, J.P. The effects of postexercise consumption of high-molecular-weight versus low-molecular-weight carbohydrate solutions on subsequent high-intensity interval-running capacity. Int. J. Sport Nutr. Exerc. Metab. 2010, 20, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.M.; Almada, A.L.; Van Eck, L.E.; Shah, M.; Mitchell, J.B.; Jones, M.T.; Jagim, A.R.; Rowlands, D.S. Ingestion of high molecular weight carbohydrate enhances subsequent repeated maximal power: A randomized controlled trial. PLoS ONE 2016, 11, e0163009. [Google Scholar] [CrossRef]
- Stephens, F.B.; Roig, M.; Armstrong, G.; Greenhaff, P.L. Post-exercise ingestion of a unique, high molecular weight glucose polymer solution improves performance during a subsequent bout of cycling exercise. J. Sports Sci. 2008, 26, 149–154. [Google Scholar] [CrossRef]
- Toussaint, H.M.; Beek, P.J. Biomechanics of competitive front crawl swimming. Sports Med. 1992, 13, 8–24. [Google Scholar] [CrossRef]
- Grijota, F.J.; Toro-Román, V.; Bartolomé, I.; Cordero-Román, E.; López, C.S.; Jiménez, J.M.; Martínez-Guardado, I. Acute Effects of 30 g Cyclodextrin Intake during CrossFit® Training on Performance and Fatigue. J. Funct. Morphol. Kinesiol. 2024, 9, 27. [Google Scholar] [CrossRef]
- Stager, J.M.; Tanner, D.A. Handbook of Sports Medicine and Science; Blackwell Science: Chichester, UK, 2005. [Google Scholar]
- Demarie, S.; Chirico, E.; Gianfelici, A.; Vannozzi, G. Anaerobic capacity assessment in elite swimmers through inertial sensors. Physiol. Meas. 2019, 40, 064003. [Google Scholar] [CrossRef] [PubMed]
- Hawley, J.A.; Williams, M.M.; Vickovic, M.M.; Handcock, P.J. Muscle power predicts freestyle swimming performance. Br. J. Sports Med. 1992, 26, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Hawley, J.A.; Williams, M.M. Relationship between upper body anaerobic power and freestyle swimming performance. Int. J. Sports Med. 1991, 12, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Kachaunov, M.; Petrov, L. Upper body anaerobic power and freestyle swimming performance. J. Phys. Educ. Sport 2020, 20, 1957–1963. [Google Scholar]
- Yapıcı, A.; Cengiz, C. The relationship between lower extremity Wingate anaerobic test (want) and 50m freestyle swimming performance. Int. J. Sport Cult. Sci. 2015, 3, 44–54. [Google Scholar] [CrossRef]
- García-Ramos, A.; Tomazin, K.; Feriche, B.; Strojnik, V.; de la Fuente, B.; Argüelles-Cienfuegos, J.; Strumbelj, B.; Štirn, I. The relationship between the lower-body muscular profile and swimming start performance. J. Hum. Kinet. 2016, 50, 157. [Google Scholar] [CrossRef]
- Keiner, M.; Wirth, K.; Fuhrmann, S.; Kunz, M.; Hartmann, H.; Haff, G.G. The influence of upper-and lower-body maximum strength on swim block start, turn, and overall swim performance in sprint swimming. J. Strength Cond. Res. 2021, 35, 2839–2845. [Google Scholar] [CrossRef]
- Kwok, W.Y.; So, B.C.L.; Tse, D.H.T.; Ng, S.S.M. A systematic review and meta-analysis: Biomechanical evaluation of the effectiveness of strength and conditioning training programs on front crawl swimming performance. J. Sports Sci. Med. 2021, 20, 564–585. [Google Scholar] [CrossRef]
- Thng, S.; Pearson, S.; Mitchell, L.J.G.; Meulenbroek, C.; Keogh, J.W.L. On-block mechanistic determinants of start performance in high performance swimmers. Sports Biomech. 2024, 23, 682–694. [Google Scholar] [CrossRef]
- West, D.J.; Owen, N.J.; Cunningham, D.J.; Cook, C.J.; Kilduff, L.P. Strength and power predictors of swimming starts in international sprint swimmers. J. Strength Cond. Res. 2011, 25, 950–955. [Google Scholar] [CrossRef] [PubMed]
- Qaisar, R.; Bhaskaran, S.; Van Remmen, H. Muscle fiber type diversification during exercise and regeneration. Free Radic. Biol. Med. 2016, 98, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Thirupathi, A.; De Souza, C.T. Multi-regulatory network of ROS: The interconnection of ROS, PGC-1 alpha, and AMPK-SIRT1 during exercise. J. Physiol. Biochem. 2017, 73, 487–494. [Google Scholar] [CrossRef]
- Beck, K.L.; von Hurst, P.R.; O’Brien, W.J.; Badenhorst, C.E. Micronutrients and athletic performance: A review. Food Chem. Toxicol. 2021, 158, 112618. [Google Scholar] [CrossRef]
- Rogers, D.R.; Lawlor, D.J.; Moeller, J.L. Vitamin C Supplementation and Athletic Performance: A Review. Curr. Sports Med. Rep. 2023, 22, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef]
- Berry, C.E.; Hare, J.M. Xanthine oxidoreductase and cardiovascular disease: Molecular mechanisms and pathophysiological implications. J. Physiol. 2004, 555, 589–606. [Google Scholar] [CrossRef]
- Bessa, A.L.; Oliveira, V.N.; Agostini, G.G.; Oliveira, R.J.S.; Oliveira, A.C.S.; White, G.E.; Wells, G.D.; Teixeira, D.N.S.; Espindola, F.S. Exercise intensity and recovery: Biomarkers of injury, inflammation, and oxidative stress. J. Strength Cond. Res. 2016, 30, 311–319. [Google Scholar] [CrossRef]
- Bloomer, R.J. Effect of exercise on oxidative stress biomarkers. Adv. Clin. Chem. 2008, 46, 1–50. [Google Scholar]
- Park, S.-Y.; Kwak, Y.-S. Impact of aerobic and anaerobic exercise training on oxidative stress and antioxidant defense in athletes. J. Exerc. Rehabil. 2016, 12, 113–117. [Google Scholar] [CrossRef]
- Shi, M.; Wang, X.; Yamanaka, T.; Ogita, F.; Nakatani, K.; Takeuchi, T. Effects of anaerobic exercise and aerobic exercise on biomarkers of oxidative stress. Environ. Health Prev. Med. 2007, 12, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Poprzecki, S.; Czuba, M.; Zajac, A.; Karpinski, J.; Wilk, R.; Bril, G.; Maszczyk, A.; Toborek, M. The blood antioxidant defence capacity during intermittent hypoxic training in elite swimmers. Biol. Sport 2016, 33, 353–360. [Google Scholar] [CrossRef]
- Škrgat, S.; Marčun, R.; Kern, I.; Šilar, M.; Šelb, J.; Fležar, M.; Korošec, P. Systemic and airway oxidative stress in competitive swimmers. Respir. Med. 2018, 137, 129–133. [Google Scholar] [CrossRef]
- Michalczyk, M.; Poprzęcki, S.; Czuba, M.; Zydek, G.; Jagsz, S.; Sadowska-Krępa, E.; Zając, A. Blood antioxidant status in road cyclists during progressive (VO2max) and constant cyclist intensity test (MLSS). J. Sports Med. Phys. Fit. 2015, 55, 855–864. [Google Scholar]
- Arazi, H.; Eghbali, E.; Suzuki, K. Creatine Supplementation, Physical Exercise and Oxidative Stress Markers: A Review of the Mechanisms and Effectiveness. Nutrients 2021, 13, 869. [Google Scholar] [CrossRef] [PubMed]
- Souza-Junior, T.P.; Lorenço-Lima, L.; Ganini, D.; Vardaris, C.V.; Polotow, T.G.; Barros, M.P. Delayed uric acid accumulation in plasma provides additional anti-oxidant protection against iron-triggered oxidative stress after a Wingate test. Biol. Sport 2014, 31, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Gaskill, S.E.; Serfass, R.C.; Bacharach, D.W.; Kelly, J.M. Responses to training in cross-country skiers. Med. Sci. Sports Exerc. 1999, 31, 1211–1217. [Google Scholar] [CrossRef]
- Sylta, Ø.; Tønnessen, E.; Hammarström, D.; Danielsen, J.; Skovereng, K.; Ravn, T.; Rønnestad, B.R.; Sandbakk, Ø.; Seiler, S. The Effect of Different High-Intensity Periodization Models on Endurance Adaptations. Med. Sci. Sports Exerc. 2016, 48, 2165–2174. [Google Scholar] [CrossRef]

{kind=link}
| Test I (T1) | Test II (T2) | |||||||
|---|---|---|---|---|---|---|---|---|
| SD | SE | SD | SE | d | ||||
| PP, W | UL | 456.6 | 81.5 | 28.81 | 465.1 | 54.8 | 19.37 | 0.12 |
| LL | 791.3 | 106.3 | 37.58 | 839.8 * | 120.1 | 42.46 | 0.43 | |
| PP relative, W/UL | UL | 5.73 | 1.07 | 0.38 | 5.78 | 0.62 | 0.22 | 0.06 |
| LL | 9.9 | 1.29 | 0.46 | 10.44 * | 1.38 | 0.49 | 0.40 | |
| Time to PP, s | UL | 11.02 | 5.68 | 2.01 | 9.95 * | 5.55 | 1.96 | 0.19 |
| LL | 9.87 | 4.52 | 1.60 | 9.4 | 6.63 | 2.34 | 0.08 | |
| Rate of fatigue, % | UL | 12.4 | 5.21 | 1.84 | 12 | 6.3 | 2.23 | 0.07 |
| LL | 13.8 | 7.0 | 2.47 | 19.3 # | 8.1 | 2.86 | 0.73 | |
| TW, J | UL | 3221.2 | 575.7 | 203.54 | 3601 | 1977.7 | 699.22 | 0.26 |
| LL | 6025 | 2531.5 | 895.02 | 5849.3 | 3060 | 1081.87 | 0.06 | |
| Work to PP relative, J/UL | UL | 40.4 | 7.8 | 2.76 | 44.8 | 24.3 | 8.59 | 0.24 |
| LL | 75 | 29.8 | 10.54 | 72.5 | 37.1 | 13.12 | 0.07 | |
| Work to PP po Pmax, J | UL | 8459.7 | 1559.7 | 551.44 | 8108.3 | 2265.7 | 801.05 | 0.18 |
| LL | 13,714 | 3591.6 | 1269.82 | 13,974.3 | 4749.3 | 1679.13 | 0.06 | |
| W from PP relative, J/UL | UL | 106.3 | 20.8 | 7.35 | 100.9 | 28.4 | 10.04 | 0.22 |
| LL | 172 | 45.3 | 16.02 | 173.6 | 57.9 | 20.47 | 0.03 | |
| TW, J | UL | 11,664.3 | 1607.1 | 568.20 | 11,709.9 | 934.7 | 330.47 | 0.04 |
| LL | 19,739.1 | 1655 | 585.13 | 19,823.6 | 2311.3 | 817.17 | 0.04 | |
| TW relative, J/UL | UL | 146.1 | 22.8 | 8.06 | 145.6 | 12.6 | 4.45 | 0.03 |
| LL | 246.9 | 17.8 | 6.29 | 246 | 24.1 | 8.52 | 0.04 | |
| Mean Power, W | UL | 388.8 | 53.7 | 18.99 | 390.4 | 31.1 | 11.00 | 0.04 |
| LL | 658 | 55.1 | 19.48 | 660.6 | 77 | 27.22 | 0.04 | |
| Mean Power relative, W/UL | UL | 4.88 | 0.75 | 0.27 | 4.86 | 0.42 | 0.15 | 0.03 |
| LL | 8.24 | 0.59 | 0.21 | 8.21 | 0.82 | 0.29 | 0.04 | |
| Men (M) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Test I (T1) | Test II (T2) | T1 vs. T2 | ||||||||
| Time | SD | SE | SD | SE | η2 | |||||
| LA, mmol/L | Rest | UL | 1.53 | 0.30 | 0.11 | 1.59 | 0.32 | 0.11 | NS | |
| LL | 1.53 | 0.30 | 0.11 | 1.59 | 0.32 | 0.11 | NS | |||
| P max | UL | 7.51 * | 1.71 | 0.60 | 8.04 * | 1.20 | 0.42 | NS | ||
| LL | 8.89 # | 1.38 | 0.49 | 9.38 # | 1.80 | 0.64 | NS | |||
| pH | Rest | UL | 7.42 | 0.05 | 0.02 | 7.41 | 0.02 | 0.01 | NS | |
| LL | 7.42 | 0.05 | 0.02 | 7.41 | 0.02 | 0.01 | NS | |||
| P max | UL | 7.30 * | 0.05 | 0.02 | 7.30 * | 0.05 | 0.02 | NS | ||
| LL | 7.28 # | 0.05 | 0.02 | 7.27 # | 0.05 | 0.02 | NS | |||
| SB, mmol/L | Rest | UL | 25.10 | 0.70 | 0.25 | 23.90 | 1.20 | 0.42 | p ≤ 0.05 | 0.05 |
| LL | 25.10 | 0.70 | 0.25 | 23.90 | 1.20 | 0.42 | p ≤ 0.05 | 0.05 | ||
| P max | UL | 17.10 * | 2.70 | 0.95 | 16.30 * | 1.50 | 0.53 | NS | ||
| LL | 16.30 # | 1.80 | 0.64 | 15.60 # | 1.80 | 0.64 | NS | |||
| Speed [m/s] | T1 | T2 | |||||
|---|---|---|---|---|---|---|---|
| SD | SE | SD | SE | d | |||
| Vmean | 1.483 | 0.08 | 0.03 | 1.481 | 0.07 | 0.02 | 0.027 |
| V7 | 1.598 | 0.10 | 0.04 | 1.625 * | 0.10 | 0.04 | 0.27 |
| V8 | 1.670 | 0.11 | 0.04 | 1.709 * | 0.13 | 0.05 | 0.32 |
| Time | Men (M) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Test I (T1) | Test II (T2) | T1 vs. T2 | |||||||
| SD | SE | SD | SE | η2 | |||||
| Glucose, mg/dL | Rest | 80.00 | 10.30 | 3.64 | 80.00 | 11.50 | 4.07 | NS | |
| After | 97.80 * | 10.10 | 3.57 | 102.00 * | 9.10 | 3.22 | NS | ||
| LA mmol/L | Rest | 1.53 | 0.27 | 0.10 | 1.28 | 0.20 | 0.07 | NS | |
| After | 10.46 * | 2.06 | 0.73 | 10.38 * | 1.63 | 0.58 | NS | ||
| UA, mg/dL | Rest | 4.99 | 0.68 | 0.24 | 5.34 | 0.39 | 0.14 | NS | |
| After | 6.69 * | 0.97 | 0.34 | 5.01 | 1.03 | 0.36 | p ≤ 0.05 | 0.10 | |
| pH | Rest | 7.39 | 0.03 | 0.01 | 7.40 | 0.02 | 0.01 | NS | |
| After | 7.17 * | 0.07 | 0.02 | 7.18 * | 0.06 | 0.02 | NS | ||
| SB, mmol/L | Rest | 23.90 | 1.70 | 0.60 | 24.50 | 0.80 | 0.28 | NS | |
| After | 12.00 * | 2.50 | 0.88 | 11.90 * | 2.00 | 0.71 | NS | ||
| BE, mmol/L | Rest | −0.60 | 1.90 | 0.67 | 0.10 | 1.00 | 0.35 | NS | |
| After | −17.20 * | 4.00 | 1.41 | −17.20 * | 3.20 | 1.13 | NS | ||
| GPx, U/g Hb | Rest | 46.20 | 9.60 | 3.39 | 34.80 | 5.60 | 1.98 | NS | |
| After | 44.70 | 7.10 | 2.51 | 36.60 | 7.40 | 2.62 | NS | ||
| SOD, U/g Hb | Rest | 1500.20 | 243.50 | 86.09 | 1112.90 | 481.70 | 170.31 | NS | |
| After | 1560.90 | 239.50 | 84.68 | 1113.60 | 492.30 | 174.05 | NS | ||
| GR, U/g Hb | Rest | 22.30 | 6.10 | 2.16 | 20.90 | 4.90 | 1.73 | NS | |
| After | 22.70 | 7.70 | 2.72 | 20.70 | 8.80 | 3.11 | NS | ||
| CAT, k/g Hb | Rest | 167.90 | 46.20 | 16.33 | 185.40 | 45.60 | 16.12 | NS | |
| After | 205.30 | 52.60 | 18.60 | 187.50 | 53.90 | 19.06 | NS | ||
| GSH, μg/mg Hb | Rest | 2.4 | 0.8 | 0.28 | 2.5 | 0.5 | 0.18 | NS | |
| After | 2 | 0.6 | 0.21 | 2.7 | 0.4 | 0.14 | NS | ||
| CK, U/L | Rest | 96.8 | 39.1 | 13.82 | 211.7 | 93.1 | 32.92 | p ≤ 0.01 | 0.14 |
| After | 164.2 * | 101.7 | 35.96 | 236.4 * | 100.7 | 35.60 | NS | ||
| LDH total, U/L | Rest | 240.5 | 54.1 | 19.13 | 247.2 | 79.6 | 28.14 | NS | |
| After | 299.5 | 47.6 | 16.83 | 272.9 | 47.1 | 16.65 | NS | ||
| FRAP, μM Troloxu | Rest | 867.3 | 118.2 | 41.79 | 814 | 51.7 | 18.28 | NS | |
| After | 988.5 | 139.2 | 49.21 | 816.5 | 69.5 | 24.57 | p ≤ 0.05 | 0.13 | |
| MDA, nmol/mL | Rest | 2.41 | 0.9 | 0.32 | 4.97 | 2.1 | 0.74 | p ≤ 0.05 | 0.12 |
| After | 2.72 | 0.91 | 0.32 | 5.3 | 2.37 | 0.84 | p ≤ 0.05 | 0.06 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gawełczyk, M.; Jagsz, S.; Zając, A.; Langfort, J. The Effects of a High Carbohydrate Diet Combined with High Molecular Weight Carbohydrate Supplementation on Anaerobic Performance and Oxidative Stress in Elite Swimmers. J. Clin. Med. 2025, 14, 3846. https://doi.org/10.3390/jcm14113846
Gawełczyk M, Jagsz S, Zając A, Langfort J. The Effects of a High Carbohydrate Diet Combined with High Molecular Weight Carbohydrate Supplementation on Anaerobic Performance and Oxidative Stress in Elite Swimmers. Journal of Clinical Medicine. 2025; 14(11):3846. https://doi.org/10.3390/jcm14113846
Chicago/Turabian StyleGawełczyk, Mateusz, Sławomir Jagsz, Adam Zając, and Józef Langfort. 2025. "The Effects of a High Carbohydrate Diet Combined with High Molecular Weight Carbohydrate Supplementation on Anaerobic Performance and Oxidative Stress in Elite Swimmers" Journal of Clinical Medicine 14, no. 11: 3846. https://doi.org/10.3390/jcm14113846
APA StyleGawełczyk, M., Jagsz, S., Zając, A., & Langfort, J. (2025). The Effects of a High Carbohydrate Diet Combined with High Molecular Weight Carbohydrate Supplementation on Anaerobic Performance and Oxidative Stress in Elite Swimmers. Journal of Clinical Medicine, 14(11), 3846. https://doi.org/10.3390/jcm14113846