
Loop-Mediated Isothermal Amplification in Schistosomiasis

 ,
,  ,
,
Abstract
1. Introduction
2. Loop-Mediated Isothermal Amplification (LAMP)
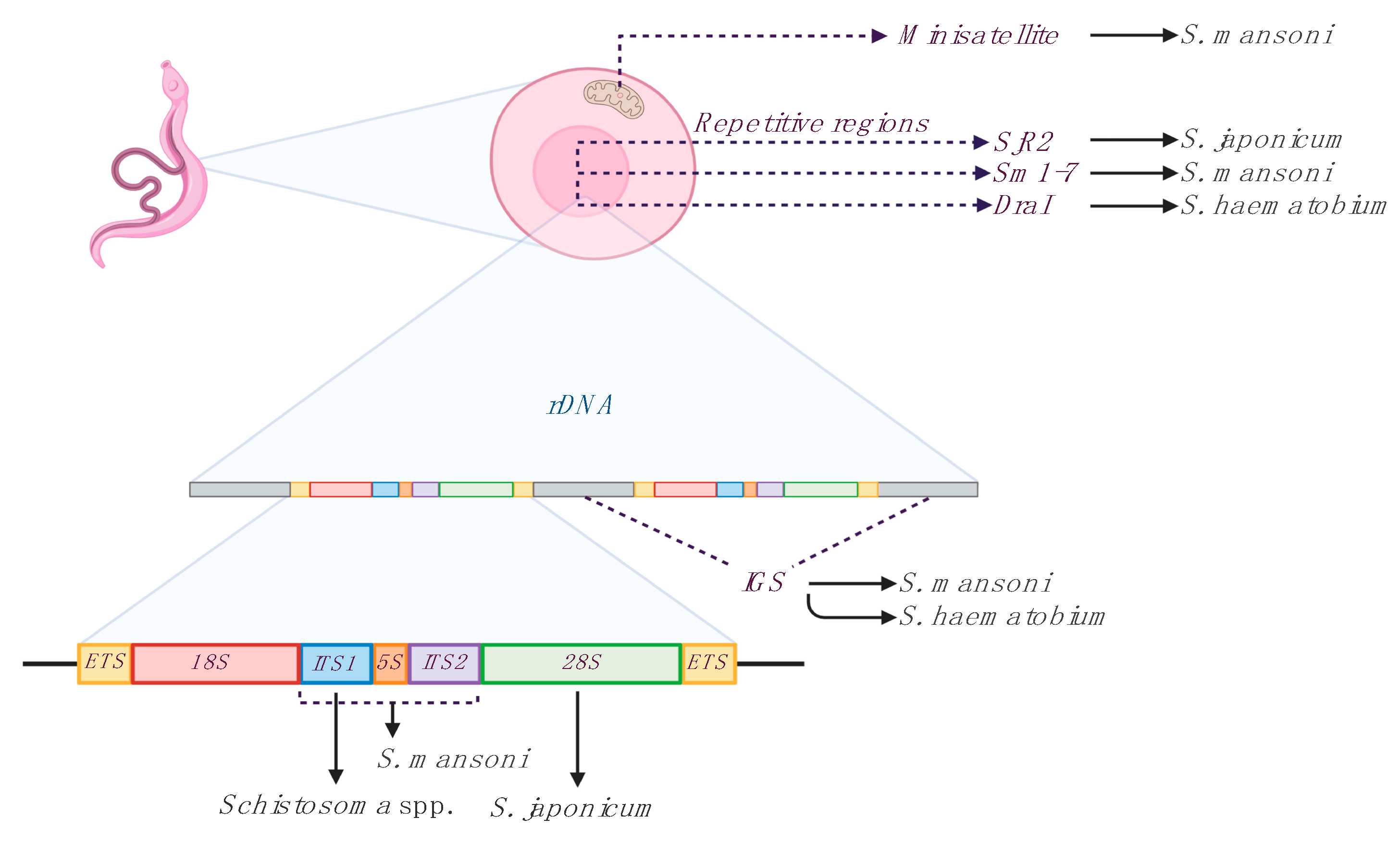
3. LAMP and Schistosomiasis
3.1. LAMP in Schistosomes Infected Snails
3.2. LAMP in Schistosomes Experimental Infections

{kind=link}
{kind=link}
| Assay Features | Clinical Application | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Species 1 | Target 2 | Detection 3 | Sensitivity 4 | Specimen 5 | n6 | Sensitivity (LAMP+/Ref+) | Specificity (LAMP−/Ref−) | Ref. Diag 7 | Ref. |
| S spp. | ITS-1 | SGI/Gel | 0.1 pg Sh 1 pg Sm/Si 10 pg Sb | gDNA | N/A | N/A | N/A | N/A | Fernández-Soto et al. [31] |
| Sh | DraI | SGI/Gel | 0.1 fg | gDNA, snails | N/A | N/A | N/A | N/A | Abbasi et al. [34] |
| DraI | SGI/Gel | N/D | Snails | N/A | N/A | N/A | N/A | Hamburguer et al. [37] | |
| IGS | SGI/Gel | 1 fg | gDNA, Urine | 94 | 100% (18/18) | 86.7% (68/76) | Micro | Gandasegui et al. [48] | |
| DraI | SGI/Gel | N/D | Urine | 86 | 100% (72/72) | 100% (14/14) | PCR | Lodh et al. [49] | |
| IGS | T/SGI/Gel | N/D | Urine | 172 | 86.2% (75/87) | N/D | Micro | Gandasegui et al. [50] | |
| Sm | Sm1–7 | SGI/Gel | 0.1 fg | gDNA, Snails | N/A | N/A | N/A | N/A | Abbasi et al. [34] |
| Sm1–7 | SGI/Gel | N/D | Snails | N/A | N/A | N/A | N/A | Hamburguer et al. [37] | |
| Mito | SGI/Gel | 1 fg | gDNA, mice stool | N/A | N/A | N/A | N/A | Fernández-Soto et al. [42] | |
| Sm1–7 | RT-EG | 0.5 fg | gDNA, mice blood, serum | N/A | N/A | N/A | N/A | Song et al. [41] | |
| 28S-18S | SGI/Gel | 0.1 fg | gDNA, snails | N/A | N/A | N/A | N/A | Gandasegui et al. [35] | |
| Sm1–7 | SGI/Gel | N/D | Urine | 86 | 100% (81/81) | 100% (5/5) | KK, PCR | Lodh et al. [49] | |
| ITS | SGI/Gel | 70 fg | gDNA, snails | N/A | N/A | N/A | N/A | Caldeira et al. [38] | |
| Mito | SGI/Gel | N/D | Snails, stool | 162 | 92.86% (12/13) | 80.11% (112/149) | KK | Gandasegui et al. [39] | |
| Sm1–7 | SGI/Gel | 32 fg | gDNA, stool | 383 | 97% (166/171) | 100% (207/207) | KK | Mwangi et al. [43] | |
| Sm1–7 | SGI/Gel | Urine | 111 | 100% (97/111) | 100% (14/14) | KK, PCR | Price et al. [51] | ||
| Mito | RT-EG/SGI/Gel | 0.01 fg/µL | Urine | 28 | 71.4% (5/7) | 71.4% (15/21) | KK | Fernández-Soto et al. [44] | |
| Mito | RT-EG/SGI/Gel | N/D | gDNA, hepatic, skin, appendix | N/A | N/A | N/A | N/A | García-Bernalt Diego et al. [52] | |
| Sj | SjR2 | SGI/Gel | 0.08 fg | gDNA, serum | 50 | 96.7% (29/30) | 100% (20/20) | Micro, PCR | Xu et al. [45] |
| 28S | T/Gel | 100 fg | gDNA, snails | N/A | N/A | N/A | N/A | Kumagi et al. [33] | |
| SjR2 | Gel | 0.1 fg | gDNA, rabbits blood | N/A | N/A | N/A | N/A | Wang et al. [47] | |
| 28S | Calcein | 100 fg | gDNA, snails | N/A | N/A | N/A | N/A | Tong et al. [36] | |
| SjR2 | SG I | N/D | Human, rabbits serum | 170 | 95.5% (105/110) | 100% (60/60) | KK | Xu et al. [46] | |
| 28S | Calcein | N/D | Snails | N/A | N/A | N/A | N/A | Qin et al. [40] | |
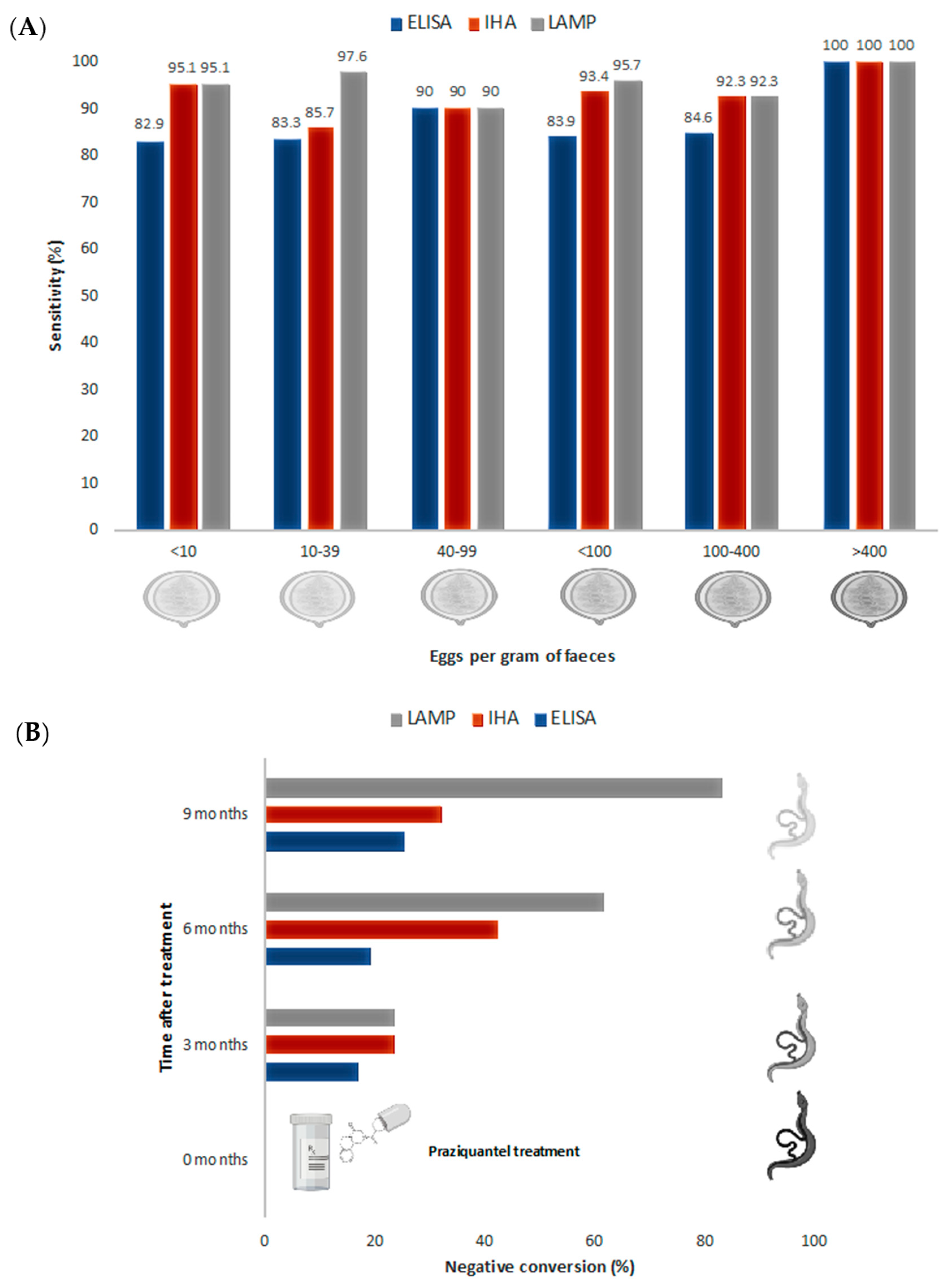
3.3. LAMP in Human Schistosomiasis Diagnosis
3.4. LAMP to Evaluate Treatment Success
3.5. Towards a True Point-of-Care Schistosomiasis Diagnostic?
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Colley, D.G.; Bustinduy, A.L.; Secor, W.E.; King, C.H. Human schistosomiasis. Lancet 2014, 383, 2253–2264. [Google Scholar] [CrossRef]
- LoVerde, P.T. Schistosomiasis. In Digenetic Trematodes, Advances in Experimental Medicine and Biology; Toledo, R., Fried, B., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 45–70. ISBN 978-3-030-18616-6. [Google Scholar]
- Chuah, C.; Gobert, G.N.; Heo, C.C.; Heo, C.C.; Leow, C.Y. Schistosomiasis in Malaysia: A review. Acta Trop. 2019, 190, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Leger, E.; Webster, J.P. Hybridizations within the GenusSchistosoma: Implications for evolution, epidemiology and control. Parasitology 2016, 144, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Berry, A.; Fillaux, J.; Martin-Blondel, G.; Boissier, J.; Iriart, X.; Marchou, B.; Magnaval, J.F.; Delobel, P. Evidence for a permanent presence of schistosomiasis in Corsica, France, 2015. Eurosurveillance 2016, 21, 5–8. [Google Scholar] [CrossRef]
- Kincaid-Smith, J.; Rey, O.; Toulza, E.; Berry, A.; Boissier, J. Emerging Schistosomiasis in Europe: A Need to Quantify the Risks. Trends Parasitol. 2017, 33, 600–609. [Google Scholar] [CrossRef]
- Meltzer, E.; Artom, G.; Marva, E.; Assous, M.V.; Rahav, G.; Schwartz, E. Schistosomiasis among Travelers: New Aspects of an Old Disease. Emerg. Infect. Dis. 2006, 12, 1696–1700. [Google Scholar] [CrossRef]
- Ross, A.G.; Vickers, D.; Olds, G.R.; Shah, S.M.; McManus, D.P. Katayama syndrome. Lancet Infect. Dis. 2007, 7, 218–224. [Google Scholar] [CrossRef]
- Bustinduy, A.L.; King, C.H. Schistosomiasis. In Manson’s Tropical Diseases; Farrar, J., White, N., Hotez, P.J., Junghanss, T., Lalloo, D., Kang, G., Eds.; Saunders Elsevier: Amsterdam, The Netherlands, 2014; pp. 698–725. [Google Scholar]
- Bhagwandeen, S.B. Schistosomiasis and carcinoma of the bladder in Zambia. S. Afr. Med. J. 1976, 50, 1616–1620. [Google Scholar]
- Barsoum, R.S.; Esmat, G.; El-Baz, T. Human Schistosomiasis: Clinical Perspective: Review. J. Adv. Res. 2013, 4, 433–444. [Google Scholar] [CrossRef]
- Fischer, P.; Leshem, E. Schistosomiasis. Encycl. Neurol. Sci. 2014, 4, 96–98. [Google Scholar] [CrossRef]
- Hinz, R.; Schwarz, N.G.; Hahn, A.; Frickmann, H. Serological approaches for the diagnosis of schistosomiasis—A review. Mol. Cell. Probes 2017, 31, 2–21. [Google Scholar] [CrossRef] [PubMed]
- Weerakoon, K.G.; Gordon, C.A.; McManus, D.P. DNA Diagnostics for Schistosomiasis Control. Trop. Med. Infect. Dis. 2018, 3, 81. [Google Scholar] [CrossRef] [PubMed]
- Schols, R.; Carolus, H.; Hammoud, C.; Mulero, S.; Mudavanhu, A.; Huyse, T. A rapid diagnostic multiplex PCR approach for xenomonitoring of human and animal schistosomiasis in a ‘One Health’ context. Trans. R. Soc. Trop. Med. Hyg. 2019, 113, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Notomi, T.; Okayama, H.; Masubuchai, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, E63. [Google Scholar] [CrossRef]
- Nagamine, K.; Hase, T.; Notomi, T. Accelerated reaction by loop-mediated isothermal amplification using loop primers. Mol. Cell. Probes 2002, 16, 223–229. [Google Scholar] [CrossRef]
- Mori, Y.; Nagamine, K.; Tomita, N.; Notomi, T. Detection of Loop-Mediated Isothermal Amplification Reaction by Turbidity Derived from Magnesium Pyrophosphate Formation. Biochem. Biophys. Res. Commun. 2001, 289, 150–154. [Google Scholar] [CrossRef]
- Mori, Y.; Kitao, M.; Tomita, N.; Notomi, T. Real-time turbidimetry of LAMP reaction for quantifying template DNA. J. Biochem. Biophys. Methods 2004, 59, 145–157. [Google Scholar] [CrossRef]
- Iwamoto, T.; Sonobe, T.; Hayashi, K. Loop-Mediated Isothermal Amplification for Direct Detection of Mycobacterium tuberculosis Complex, M. avium, and M. intracellulare in Sputum Samples. J. Clin. Microbiol. 2003, 41, 2616–2622. [Google Scholar] [CrossRef]
- Tomita, N.; Mori, Y.; Kanda, H.; Notomi, T. Loop-mediated isothermal amplification (LAMP) of gene sequences and simple visual detection of products. Nat. Protoc. 2008, 3, 877–882. [Google Scholar] [CrossRef]
- Goto, M.; Honda, E.; Ogura, A.; Nomoto, A.; Hanaki, K.-I. Colorimetric detection of loop-mediated isothermal amplification reaction by using hydroxy naphthol blue. Biotechnology 2009, 46, 167–172. [Google Scholar] [CrossRef]
- Nzelu, C.O.; Gomez, E.A.; Cáceres, A.G.; Sakurai, T.; Martini-Robles, L.; Uezato, H.; Mimori, T.; Katakura, K.; Hashiguchi, Y.; Kato, H. Development of a loop-mediated isothermal amplification method for rapid mass-screening of sand flies for Leishmania infection. Acta Trop. 2014, 132, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Poole, C.B.; Li, Z.; Alhassan, A.; Guelig, D.; Diesburg, S.; Tanner, N.A.; Zhang, Y.; Evans, T.C., Jr.; Labarre, P.; Wanji, S.; et al. Colorimetric tests for diagnosis of filarial infection and vector surveillance using non-instrumented nucleic acid loop-mediated isothermal amplification (NINA-LAMP). PLoS ONE 2017, 12, e0169011. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lowe, S.B.; Gooding, J. Brief review of monitoring methods for loop-mediated isothermal amplification (LAMP). Biosens. Bioelectron. 2014, 61, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Xu, Y.; Fohlerova, Z.; Chang, H.; Iliescu, C.; Neuzil, P. LAMP-on-a-chip: Revising microfluidic platforms for loop-mediated DNA amplification. TrAC Trends Anal. Chem. 2019, 113, 44–53. [Google Scholar] [CrossRef]
- Shang, Y.; Sun, J.; Ye, Y.; Zhang, J.; Zhang, Y.; Sun, X. Loop-mediated isothermal amplification-based microfluidic chip for pathogen detection. Crit. Rev. Food Sci. Nutr. 2020, 60, 201–224. [Google Scholar] [CrossRef]
- Ye, X.; Fang, X.; Li, X.; Kong, J. Gold nanoparticle-mediated nucleic acid isothermal amplification with enhanced specificity. Anal. Chim. Acta 2018, 1043, 150–157. [Google Scholar] [CrossRef]
- Ong, D.S.Y.; Poljak, M. Smartphones as mobile microbiological laboratories. Clin. Microbiol. Infect. 2020, 26, 421–424. [Google Scholar] [CrossRef]
- Hashimoto, M.; Sakamoto, H.; Ido, Y.; Tanaka, M.; Yatsushiro, S.; Kajimoto, K.; Kataoka, M. In situ loop-mediated isothermal amplification (LAMP) for identification of Plasmodium species in wide-range thin blood smears. Malar. J. 2018, 17, 235. [Google Scholar] [CrossRef]
- Fernández-Soto, P.; Avendaño, C.; Sala-Vizcaíno, A.; Crego-Vicente, B.; Febrer-Sendra, B.; Diego, J.G.-B.; Oleaga, A.; López-Abán, J.; Vicente, B.; Patarroyo, M.A.; et al. Molecular Markers for Detecting Schistosoma Species by Loop-Mediated Isothermal Amplification. Dis. Markers 2020, 2020, 1–11. [Google Scholar] [CrossRef]
- Cnops, L.; Soentjens, P.; Clerinx, J.; Van Esbroeck, M. A Schistosoma haematobium-Specific Real-Time PCR for Diagnosis of Urogenital Schistosomiasis in Serum Samples of International Travelers and Migrants. PLoS Negl. Trop. Dis. 2013, 7, e2413. [Google Scholar] [CrossRef]
- Kumagai, T.; Lu, S.; Wang, T.-P.; Furushima-Shimogawara, R.; Wen, L.; Ohta, N.; Chen, R.; Ohmae, H. Detection of Early and Single Infections of Schistosoma japonicum in the Intermediate Host Snail, Oncomelania hupensis, by PCR and Loop-Mediated Isothermal Amplification (LAMP) Assay. Am. J. Trop. Med. Hyg. 2010, 83, 542–548. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, I.; Muchiri, E.M.; Hamburger, J.; King, C.H. Detection of Schistosoma mansoni and Schistosoma haematobium DNA by Loop-Mediated Isothermal Amplification: Identification of Infected Snails from Early Prepatency. Am. J. Trop. Med. Hyg. 2010, 83, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Gandasegui, J.; Fernández-Soto, P.; Hernández-Goenaga, J.; López-Abán, J.; Vicente, B.; Muro, A. Biompha-LAMP: A New Rapid Loop-Mediated Isothermal Amplification Assay for Detecting Schistosoma mansoni in Biomphalaria glabrata Snail Host. PLoS Negl. Trop. Dis. 2016, 10, e0005225. [Google Scholar] [CrossRef] [PubMed]
- Tong, Q.-B.; Chen, R.; Zhang, Y.; Yang, G.-J.; Kumagai, T.; Furushima-Shimogawara, R.; Lou, D.; Yang, K.; Wen, L.-Y.; Lu, S.; et al. A new surveillance and response tool: Risk map of infected Oncomelania hupensis detected by Loop-mediated isothermal amplification (LAMP) from pooled samples. Acta Trop. 2015, 141, 170–177. [Google Scholar] [CrossRef]
- Hamburger, J.; Mzungu, E.; Abbasi, I.; Mungai, P.; Wanjala, A.; King, C.H.; Kariuki, C.; Muchiri, E. Evaluation of Loop-Mediated Isothermal Amplification Suitable for Molecular Monitoring of Schistosome-Infected Snails in Field Laboratories. Am. J. Trop. Med. Hyg. 2013, 88, 344–351. [Google Scholar] [CrossRef]
- Caldeira, R.L.; Jannotti-Passos, L.K.; Carvalho, O.D.S. Use of Molecular Methods for the Rapid Mass Detection ofSchistosoma mansoni(Platyhelminthes: Trematoda) inBiomphalariaspp. (Gastropoda: Planorbidae). J. Trop. Med. 2017, 2017, 1–6. [Google Scholar] [CrossRef]
- Gandasegui, J.; Fernández-Soto, P.; Muro, A.; Barbosa, C.S.; De Melo, F.L.; Loyo, R.; Gomes, E.C.D.S. A field survey using LAMP assay for detection of Schistosoma mansoni in a low-transmission area of schistosomiasis in Umbuzeiro, Brazil: Assessment in human and snail samples. PLoS Negl. Trop. Dis. 2018, 12, e0006314. [Google Scholar] [CrossRef]
- Qin, Z.-Q.; Xu, J.; Feng, T.; Lv, S.; Qian, Y.-J.; Zhang, L.-J.; Li, Y.-L.; Lv, C.; Bergquist, R.; Li, S.-Z.; et al. Field Evaluation of a Loop-Mediated Isothermal Amplification (LAMP) Platform for the Detection of Schistosoma japonicum Infection in Oncomelania hupensis Snails. Trop. Med. Infect. Dis. 2018, 3, 124. [Google Scholar] [CrossRef]
- Song, J.; Liu, C.; Bais, S.; Mauk, M.G.; Bau, H.H.; Greenberg, R.M. Molecular Detection of Schistosome Infections with a Disposable Microfluidic Cassette. PLoS Negl. Trop. Dis. 2015, 9, e0004318. [Google Scholar] [CrossRef]
- Fernández-Soto, P.; Arahuetes, J.G.; Hernández, A.S.; Abán, J.L.; Santiago, B.V.; Muro, A. A Loop-Mediated Isothermal Amplification (LAMP) Assay for Early Detection of Schistosoma mansoni in Stool Samples: A Diagnostic Approach in a Murine Model. PLoS Negl. Trop. Dis. 2014, 8, e3126. [Google Scholar] [CrossRef]
- Mwangi, I.N.; Agola, E.L.; Mugambi, R.M.; Shiraho, E.A.; Mkoji, G.M. Development and Evaluation of a Loop-Mediated Isothermal Amplification Assay for Diagnosis of Schistosoma mansoni Infection in Faecal Samples. J. Parasitol. Res. 2018, 2018, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Soto, P.; Gandasegui, J.; Rodríguez, C.C.; Pérez-Arellano, J.L.; Crego-Vicente, B.; Diego, J.G.-B.; López-Abán, J.; Vicente, B.; Muro, A. Detection of Schistosoma mansoni-derived DNA in human urine samples by loop-mediated isothermal amplification (LAMP). PLoS ONE 2019, 14, e0214125. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Rong, R.; Zhang, H.; Shi, C.; Zhu, X.; Xia, C. Sensitive and rapid detection of Schistosoma japonicum DNA by loop-mediated isothermal amplification (LAMP). Int. J. Parasitol. 2010, 40, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Guan, Z.-X.; Zhao, B.; Wang, Y.-Y.; Cao, Y.; Zhang, H.-Q.; Zhu, X.-Q.; He, Y.-K.; Xia, C.-M. DNA Detection of Schistosoma japonicum: Diagnostic Validity of a LAMP Assay for Low-Intensity Infection and Effects of Chemotherapy in Humans. PLoS Negl. Trop. Dis. 2015, 9, e0003668. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, L.; Yin, X.-R.; Hua, W.; Hou, M.; Ji, M.; Yu, C.-X.; Wu, G. Application of DNA-based diagnostics in detection of schistosomal DNA in early infection and after drug treatment. Parasites Vectors 2011, 4, 164. [Google Scholar] [CrossRef]
- Gandasegui, J.; Fernández-Soto, P.; Carranza-Rodríguez, C.; Pérez-Arellano, J.L.; Vicente, B.; López-Abán, J.; Muro, A. The Rapid-Heat LAMPellet Method: A Potential Diagnostic Method for Human Urogenital Schistosomiasis. PLoS Negl. Trop. Dis. 2015, 9, e0003963. [Google Scholar] [CrossRef]
- Lodh, N.; Mikita, K.; Bosompem, K.M.; Anyan, W.K.; Quartey, J.K.; Otchere, J.; Shiff, C.J. Point of care diagnosis of multiple schistosome parasites: Species-specific DNA detection in urine by loop-mediated isothermal amplification (LAMP). Acta Trop. 2017, 173, 125–129. [Google Scholar] [CrossRef]
- Gandasegui, J.; Fernández-Soto, P.; DaCal, E.; Rodriguez, E.; Saugar, J.M.; Yepes, E.; Aznar-Ruiz-De-Alegría, M.L.; Espasa, M.; Ninda, A.; Bocanegra, C.; et al. Field and laboratory comparative evaluation of a LAMP assay for the diagnosis of urogenital schistosomiasis in Cubal, Central Angola. Trop. Med. Int. Health 2018, 23, 992–1001. [Google Scholar] [CrossRef]
- Price, M.; Cyrs, A.; Sikasunge, C.S.; Mwansa, J.; Lodh, N. Testing the Infection Prevalence of Schistosoma mansoni after Mass Drug Administration by Comparing Sensitivity and Specificity of Species-Specific Repeat Fragment Amplification by PCR and Loop-Mediated Isothermal Amplification. Am. J. Trop. Med. Hyg. 2019, 101, 78–83. [Google Scholar] [CrossRef]
- Diego, J.G.-B.; Fernández-Soto, P.; Crego-Vicente, B.; Alonso-Castrillejo, S.; Febrer-Sendra, B.; Gómez-Sánchez, A.; Vicente, B.; López-Abán, J.; Muro, A. Progress in loop-mediated isothermal amplification assay for detection of Schistosoma mansoni DNA: Towards a ready-to-use test. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Bayoumi, A.; Al-Refai, S.; Badr, M.; Abd El-Aal, A.; El Akkad, D.; Saad, N.; KM, E.; Abdel Aziz, I. Loop-mediated isother-mal amplification (LAMP): Sensitive and rapid detection of Schistosoma haematobium DNA in urine samples from Egyptian suspected cases. J. Egypt. Soc. Parasitol. 2016, 46, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Camprubí-Ferrer, D.; Requena-Méndez, A.; Rodríguez, N.; Valls, M.E.; García-Herrera, A.; Estrach, T.; Fustà, X.; Diego, J.G.-B.; Fernández-Soto, P.; Muñoz, J. A 38-year-old woman with zosteriform skin lesions. PLoS Negl. Trop. Dis. 2017, 11, e0005906. [Google Scholar] [CrossRef]
- Guegan, H.; Fillaux, J.; Charpentier, E.; Robert-Gangneux, F.; Chauvin, P.; Guemas, E.; Boissier, J.; Valentin, A.; Cassaing, S.; Gangneux, J.-P.; et al. Real-time PCR for diagnosis of imported schistosomiasis. PLoS Negl. Trop. Dis. 2019, 13, e0007711. [Google Scholar] [CrossRef] [PubMed]
- Papaiakovou, M.; Gasser, R.B.; Littlewood, D.T.J. Quantitative PCR-Based Diagnosis of Soil-Transmitted Helminth Infections: Faecal or Fickle? Trends Parasitol. 2019, 35, 491–500. [Google Scholar] [CrossRef]
- Mabey, D.; Peeling, R.W.; Ustianowski, A.; Perkins, M.D. Diagnostics for the developing world. Nat. Rev. Genet. 2004, 2, 231–240. [Google Scholar] [CrossRef]
- Peeling, R.W.; Holmes, K.K.; Mabey, D.; Ronald, A. Rapid tests for sexually transmitted infections (STIs): The way forward. Sex. Transm. Infect. 2006, 82, v1–v6. [Google Scholar] [CrossRef]
- Land, K.; Boeras, D.I.; Chen, X.-S.; Ramsay, A.R.; Peeling, R.W. REASSURED diagnostics to inform disease control strategies, strengthen health systems and improve patient outcomes. Nat. Microbiol. 2019, 4, 46–54. [Google Scholar] [CrossRef]
- Zou, Y.; Mason, M.G.; Wang, Y.; Wee, E.; Turni, C.; Blackall, P.J.; Trau, M.; Botella, J.R. Nucleic acid purification from plants, animals and microbes in under 30 seconds. PLoS Biol. 2017, 15, e2003916. [Google Scholar] [CrossRef]
- Chotiwan, N.; Brewster, C.D.; Magalhaes, T.; Weger-Lucarelli, J.; Duggal, N.K.; Rückert, C.; Nguyen, C.; Luna, S.M.G.; Fauver, J.R.; Andre, B.; et al. Rapid and specific detection of Asian- and African-lineage Zika viruses. Sci. Transl. Med. 2017, 9, eaag0538. [Google Scholar] [CrossRef]
- Fernández-Soto, P.; Celis-Giraldo, C.; Collar-Fernández, C.; Gorgojo, Ó.; Camargo, M.; Muñoz, J.; Salas-Coronas, J.; Patarroyo, M.A.; Muro, A. Strong-LAMP Assay Based on a Strongyloides spp.-Derived Partial Sequence in the 18S rRNA as Potential Biomarker for Strongyloidiasis Diagnosis in Human Urine Samples. Dis. Markers 2020, 2020, 1–10. [Google Scholar] [CrossRef]
- Ajibola, O.; Gulumbe, B.H.; A Eze, A.; Obishakin, E. Tools for Detection of Schistosomiasis in Resource Limited Settings. Med. Sci. 2018, 6, 39. [Google Scholar] [CrossRef] [PubMed]
- Francois, P.; Tangomo, M.; Hibbs, J.; Bonetti, E.-J.; Boehme, C.C.; Notomi, T.; Perkins, M.D.; Schrenzel, J. Robustness of a loop-mediated isothermal amplification reaction for diagnostic applications. FEMS Immunol. Med. Microbiol. 2011, 62, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Liu, C.; Mauk, M.G.; Rankin, S.C.; Lok, J.B.; Greenberg, R.M.; Bau, H.H. Two-Stage Isothermal Enzymatic Amplification for Concurrent Multiplex Molecular Detection. Clin. Chem. 2017, 63, 714–722. [Google Scholar] [CrossRef] [PubMed]
- Wood, C.S.; Thomas, M.R.; Budd, J.; Mashamba-Thompson, T.P.; Herbst, K.; Pillay, D.; Peeling, R.W.; Johnson, A.M.; McKendry, R.A.; Stevens, M.M. Taking connected mobile-health diagnostics of infectious diseases to the field. Nat. Cell Biol. 2019, 566, 467–474. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Bernalt Diego, J.; Fernández-Soto, P.; Febrer-Sendra, B.; Crego-Vicente, B.; Muro, A. Loop-Mediated Isothermal Amplification in Schistosomiasis. J. Clin. Med. 2021, 10, 511. https://doi.org/10.3390/jcm10030511
García-Bernalt Diego J, Fernández-Soto P, Febrer-Sendra B, Crego-Vicente B, Muro A. Loop-Mediated Isothermal Amplification in Schistosomiasis. Journal of Clinical Medicine. 2021; 10(3):511. https://doi.org/10.3390/jcm10030511
Chicago/Turabian StyleGarcía-Bernalt Diego, Juan, Pedro Fernández-Soto, Begoña Febrer-Sendra, Beatriz Crego-Vicente, and Antonio Muro. 2021. "Loop-Mediated Isothermal Amplification in Schistosomiasis" Journal of Clinical Medicine 10, no. 3: 511. https://doi.org/10.3390/jcm10030511
APA StyleGarcía-Bernalt Diego, J., Fernández-Soto, P., Febrer-Sendra, B., Crego-Vicente, B., & Muro, A. (2021). Loop-Mediated Isothermal Amplification in Schistosomiasis. Journal of Clinical Medicine, 10(3), 511. https://doi.org/10.3390/jcm10030511