Polyamine Transport Protein PotD Protects Mice against Haemophilus parasuis and Elevates the Secretion of Pro-Inflammatory Cytokines of Macrophage via JNK–MAPK and NF–κB Signal Pathways through TLR4

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Ethics Statement
2.2. Strains, Plasmids, Primers and Bacterial Growth Conditions
2.3. Expression/Purification of Recombinant PotD Protein, and Removal of Endotoxin Contamination
2.4. Immunization
2.5. Analysis of Cellular Immune Response of Mice Using Immunofluorescent (IF) Assay and Flow Cytometry
2.6. Analysis of Lymphocyte Proliferation and Detection of Cytokines Using ELISA
2.7. Mouse Challenge
2.8. Bacterial Clearance
2.9. Cell Line and Culture Conditions
2.10. Detected of Pro-Inflammatory Cytokines Induced by PotD Using ELISA and Quantitative Real-Time PCR
2.11. Toll-Like Receptors 2 (TLR2) and 4 (TLR4) Blocking Assay
2.12. Western Blotting Assay for Induction of Pro-Inflammatory Cytokines by PotD
2.13. Determination of Working Concentrations of Inhibitors Using MTT Assay
2.14. Effect of Inhibitor on the MAPK and NF-κB Signal Pathways
2.15. Statistical Analysis
3. Results
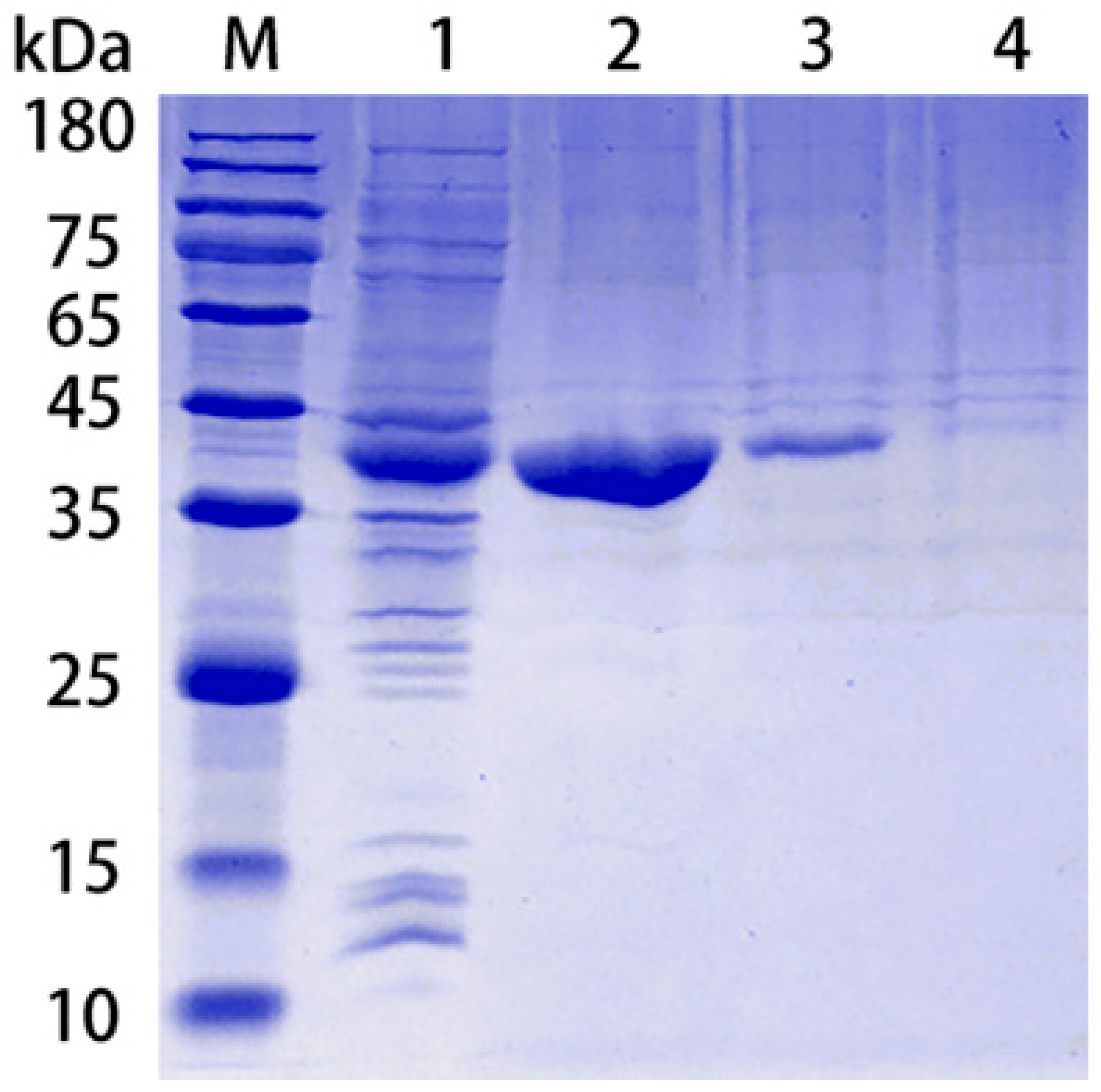
3.1. Purification of rPotD Protein, Endotoxin Elimination, and Polyclonal Antibody Preparation
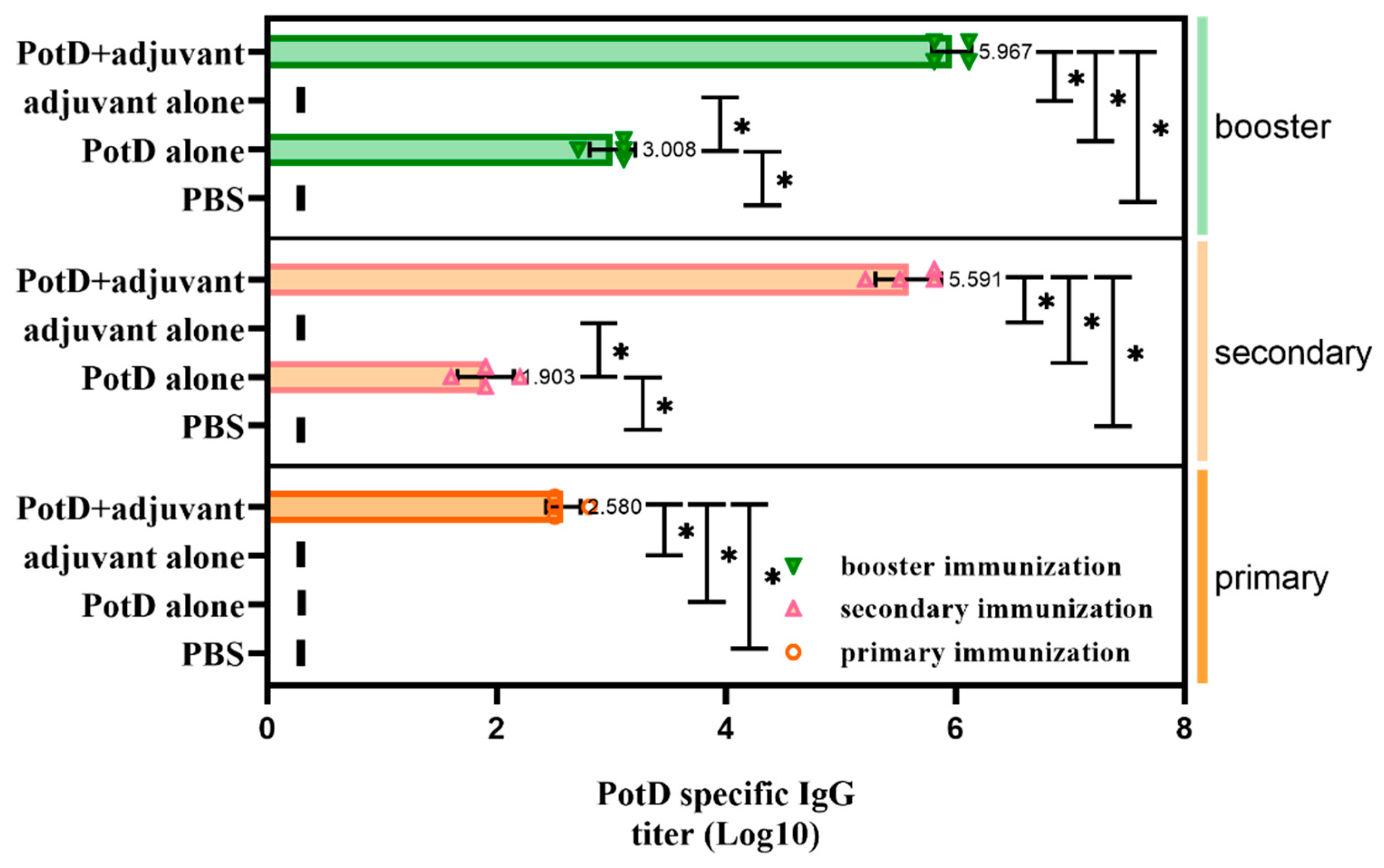
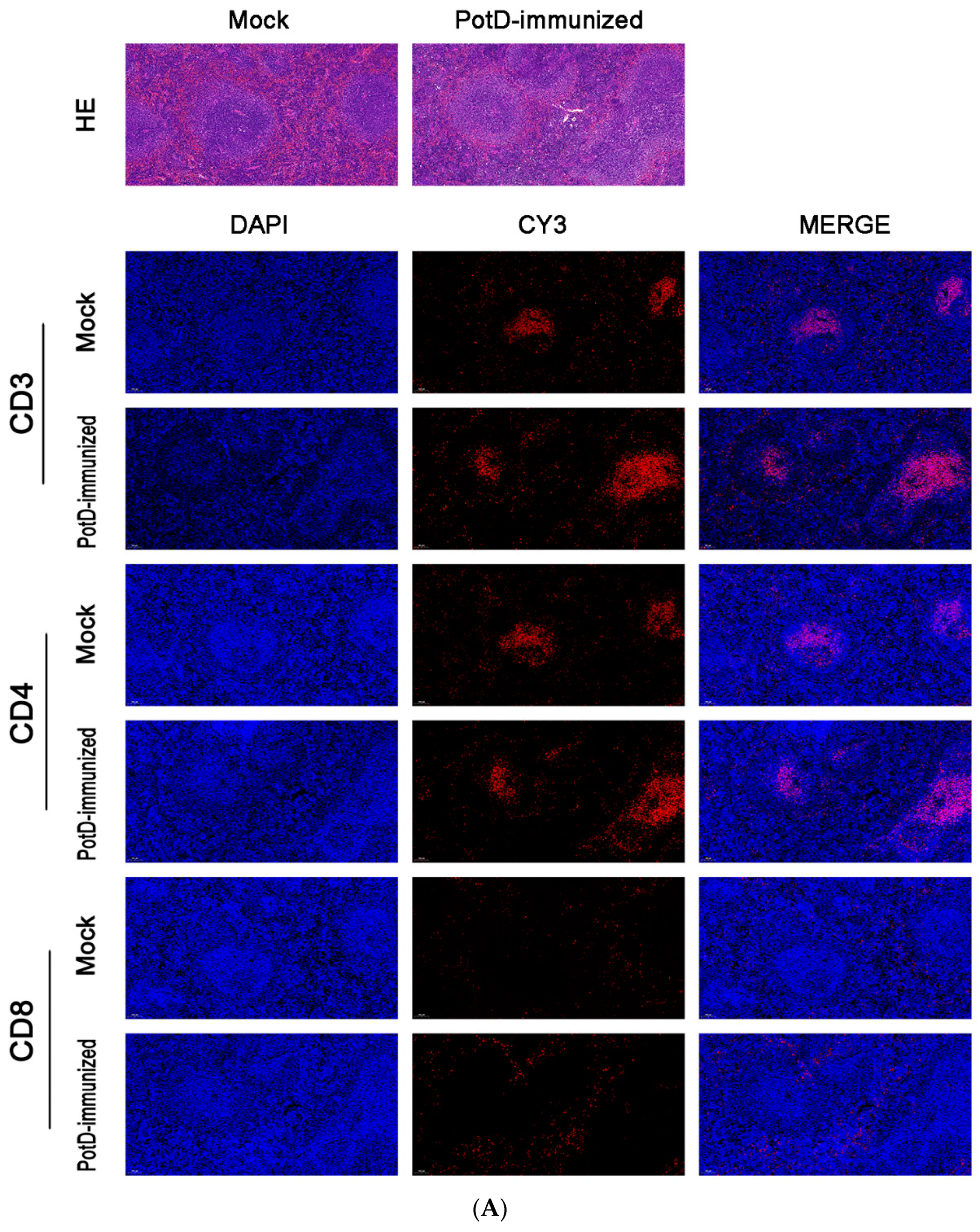
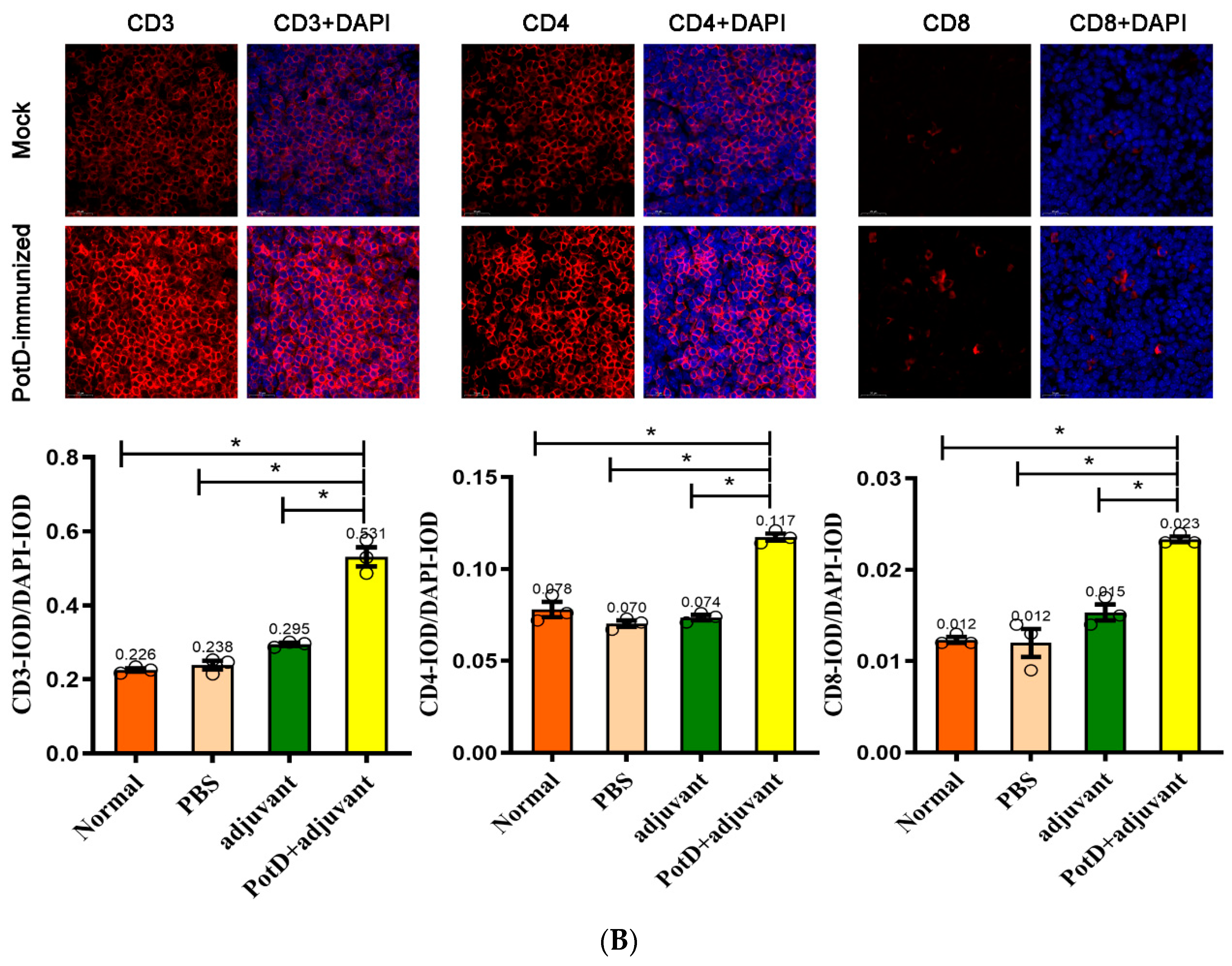
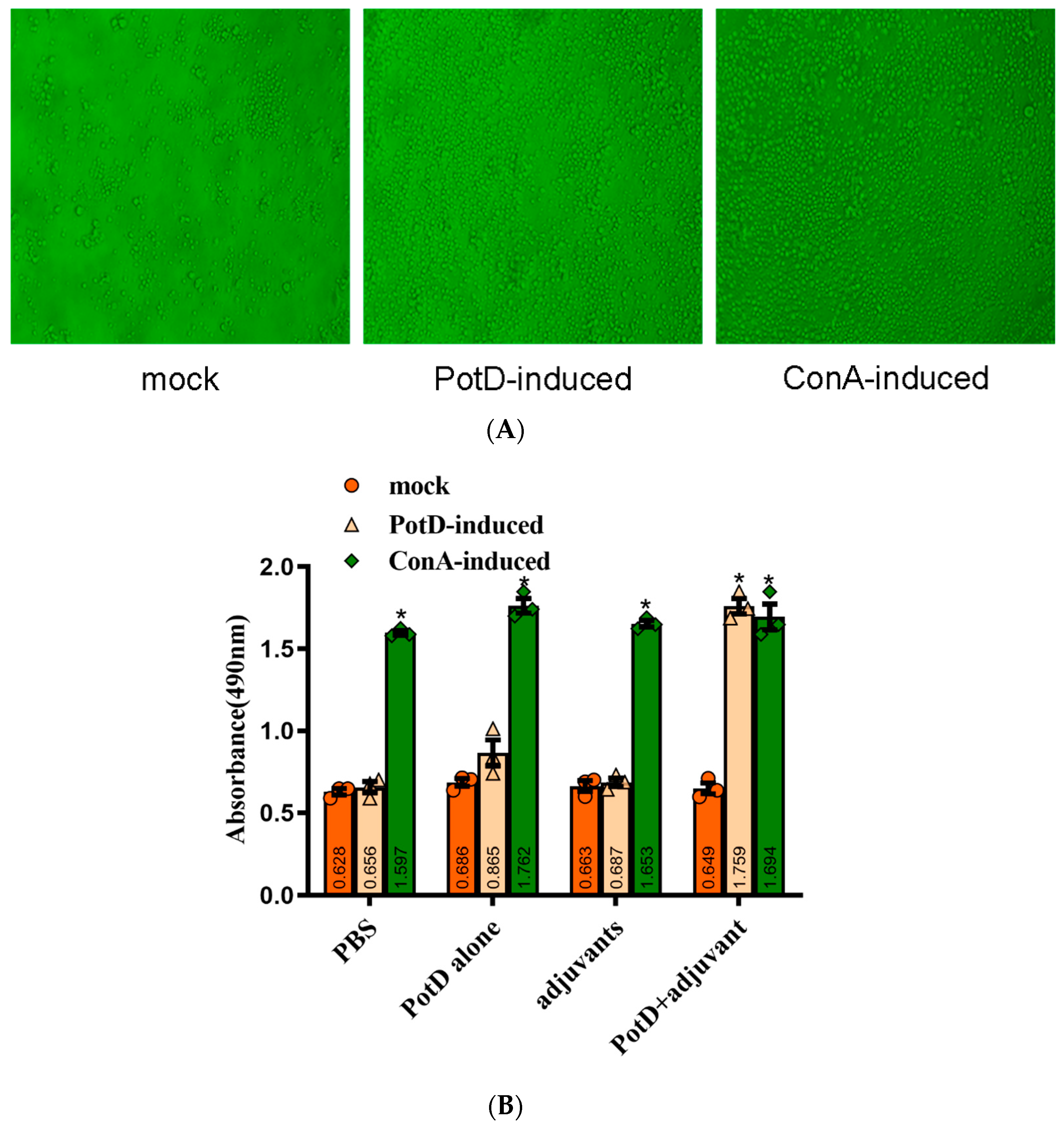
3.2. Recombinant PotD Protein Mediates a Cellular Immune Response
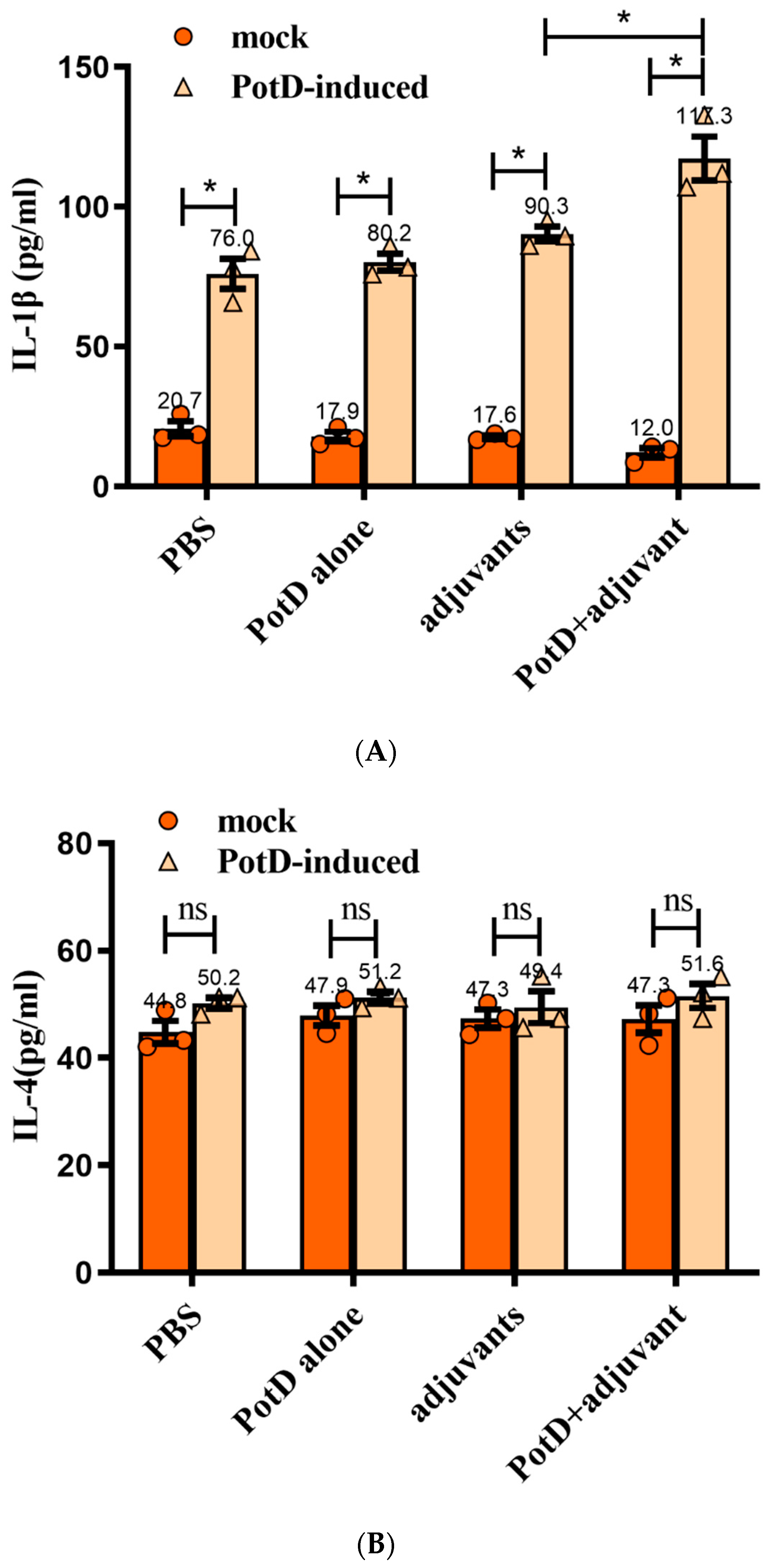
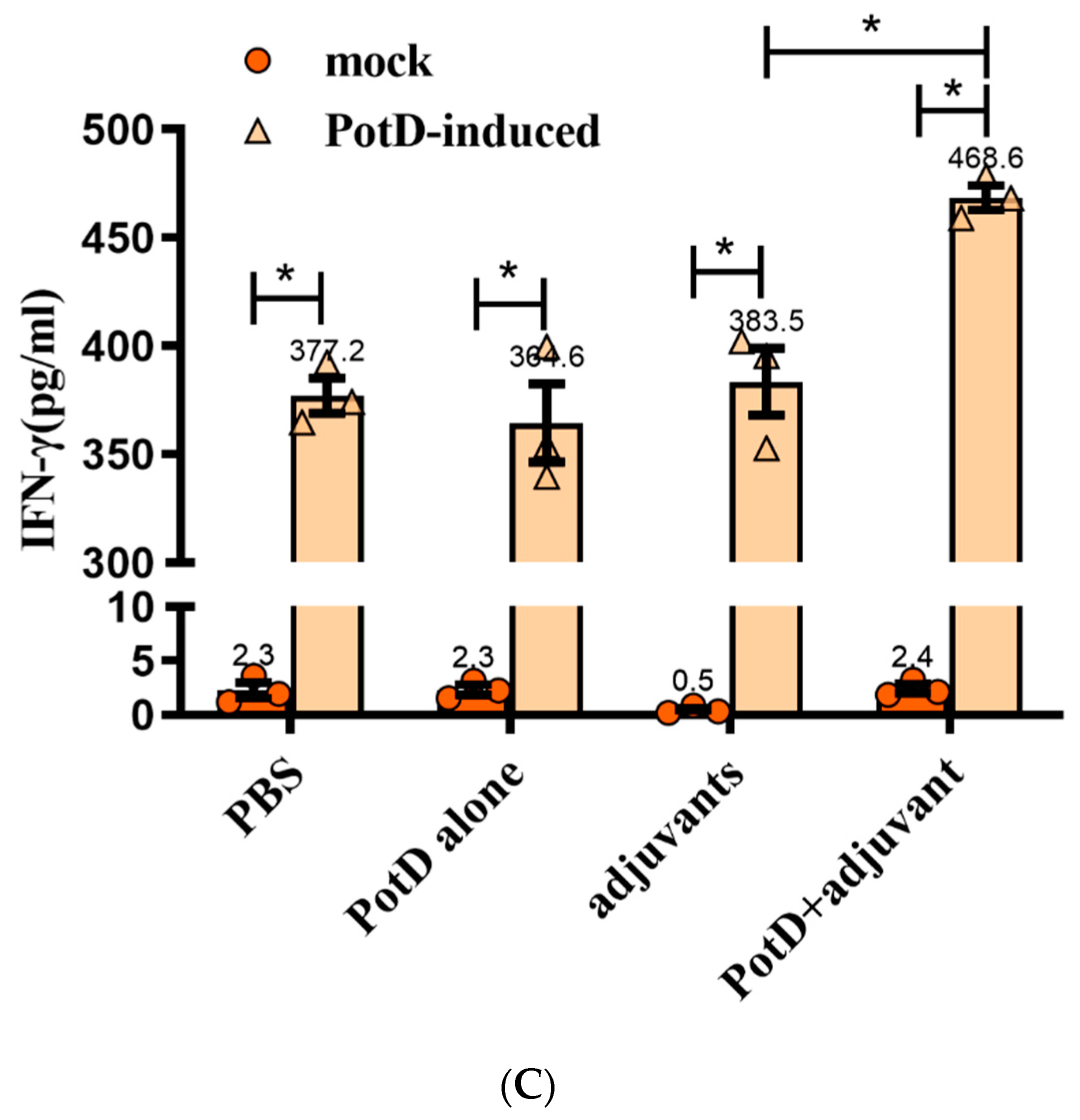
3.3. Lymphocyte Proliferation Assay and Detection of Cytokines
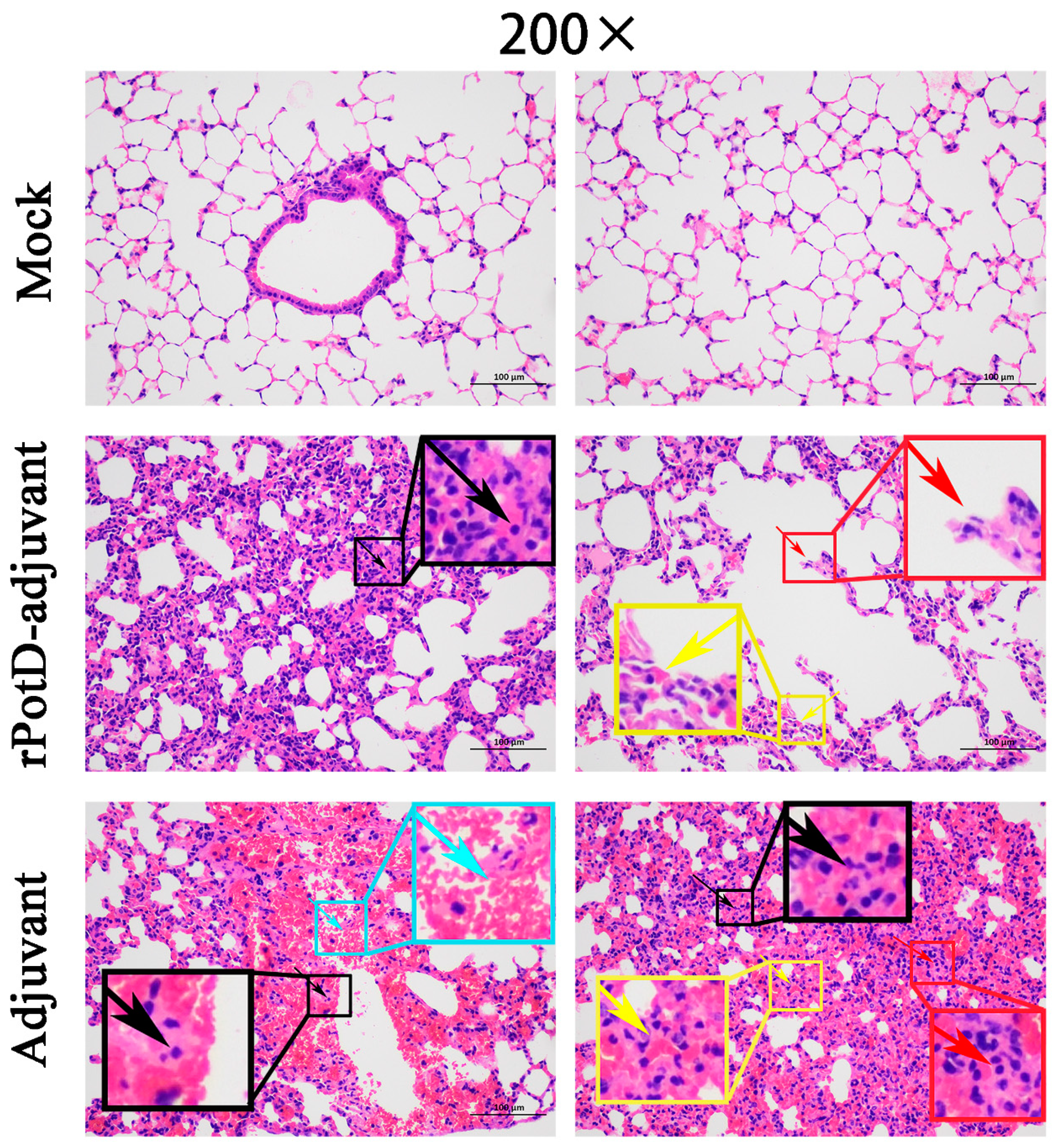
3.4. rPotD-Vaccinated Mice Show Less Severe Pathological Signs in Multiple Tissues
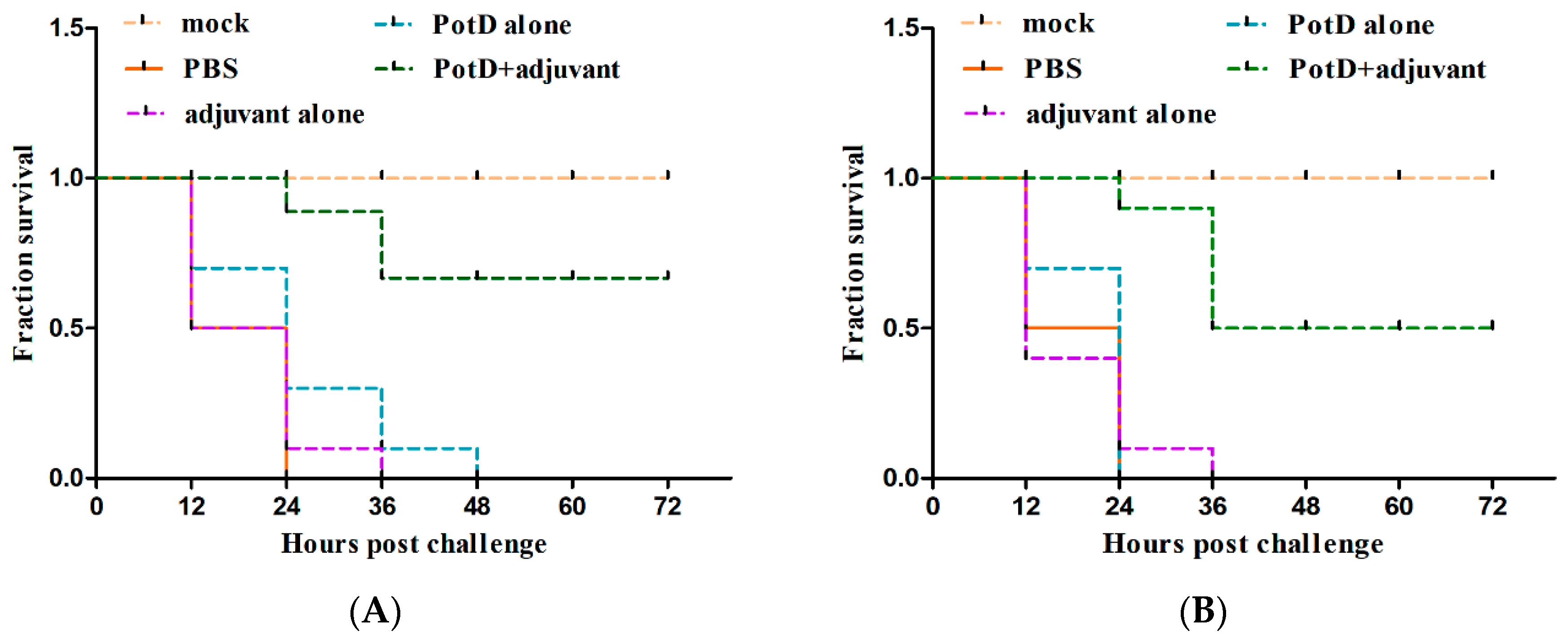
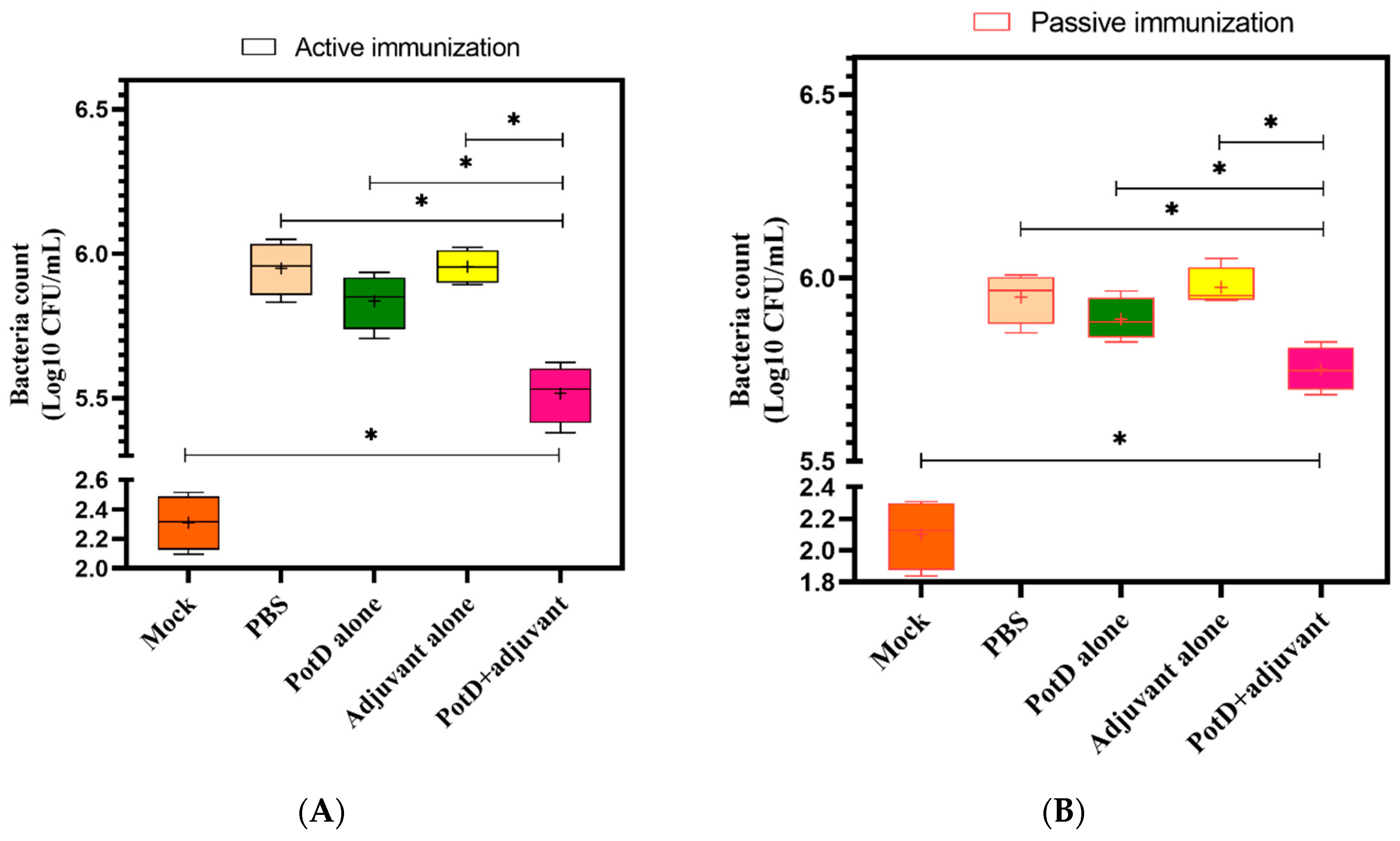
3.5. rPotD+Adjuvant-Vaccinated Mice Showed Significantly Elevated Bacterial Clearance Ability
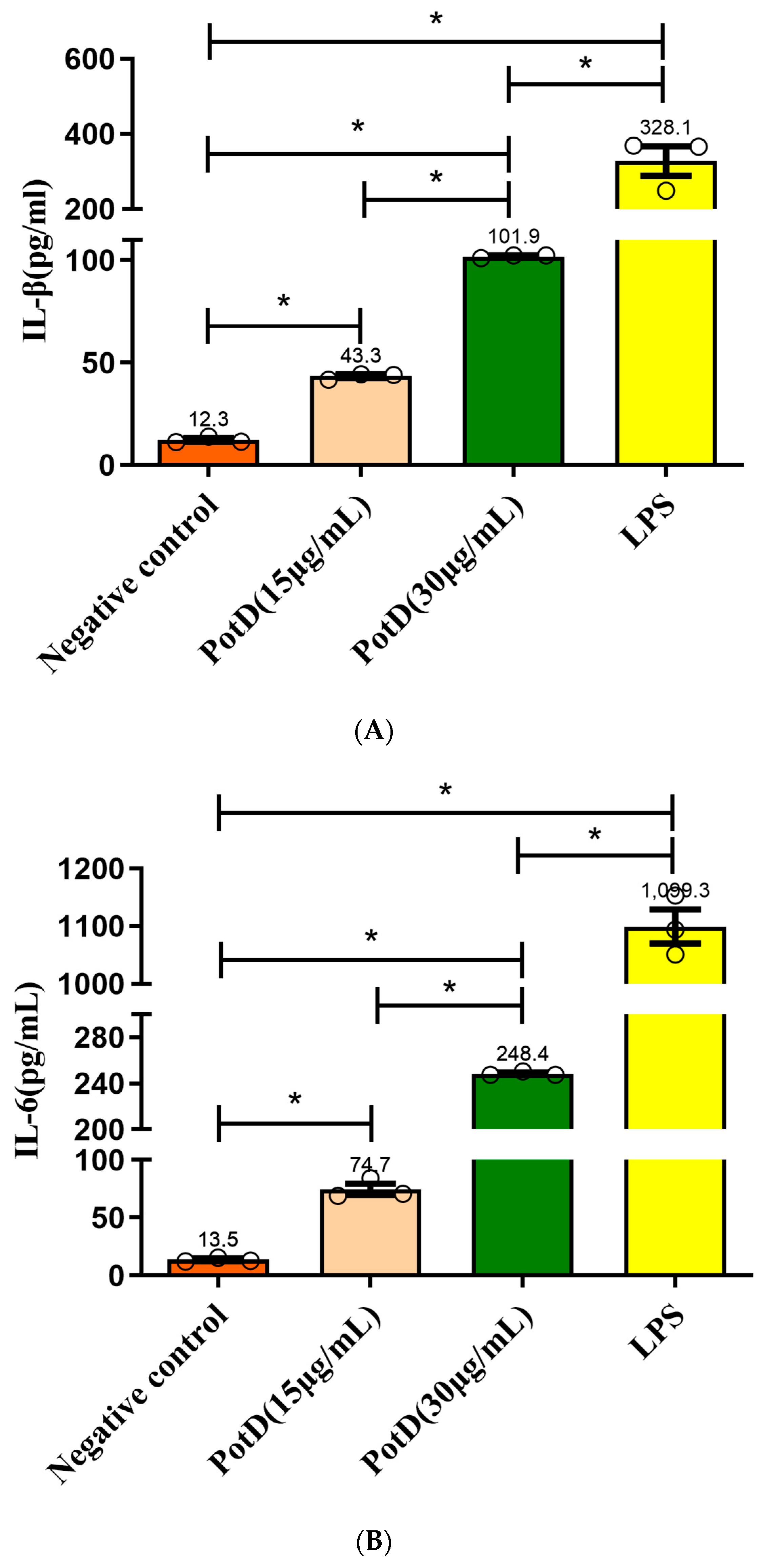
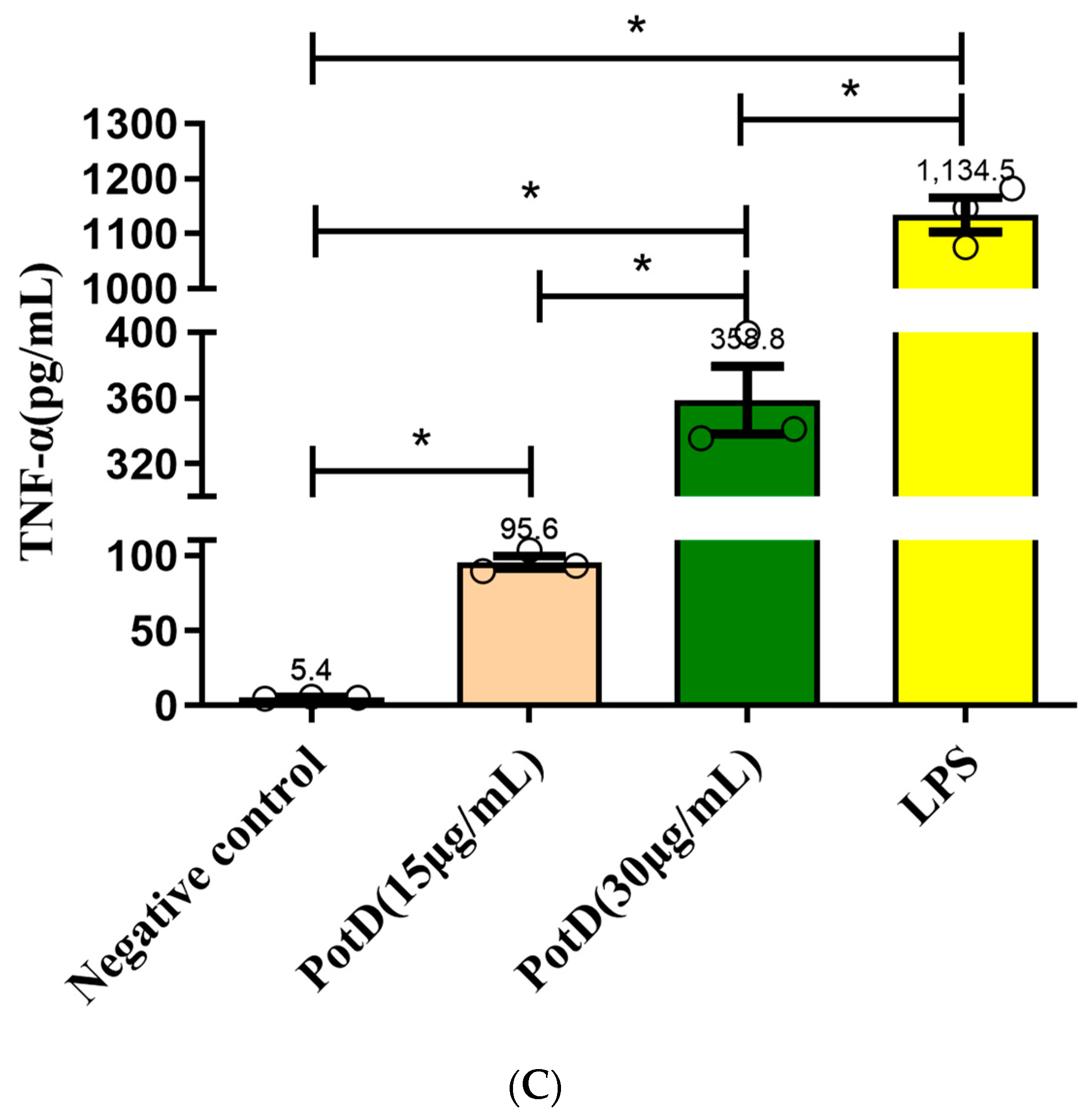
3.6. rPotD Actively Elevates the Levels of Pro-Inflammatory Cytokines IL-1β, IL-6, and TNF-α
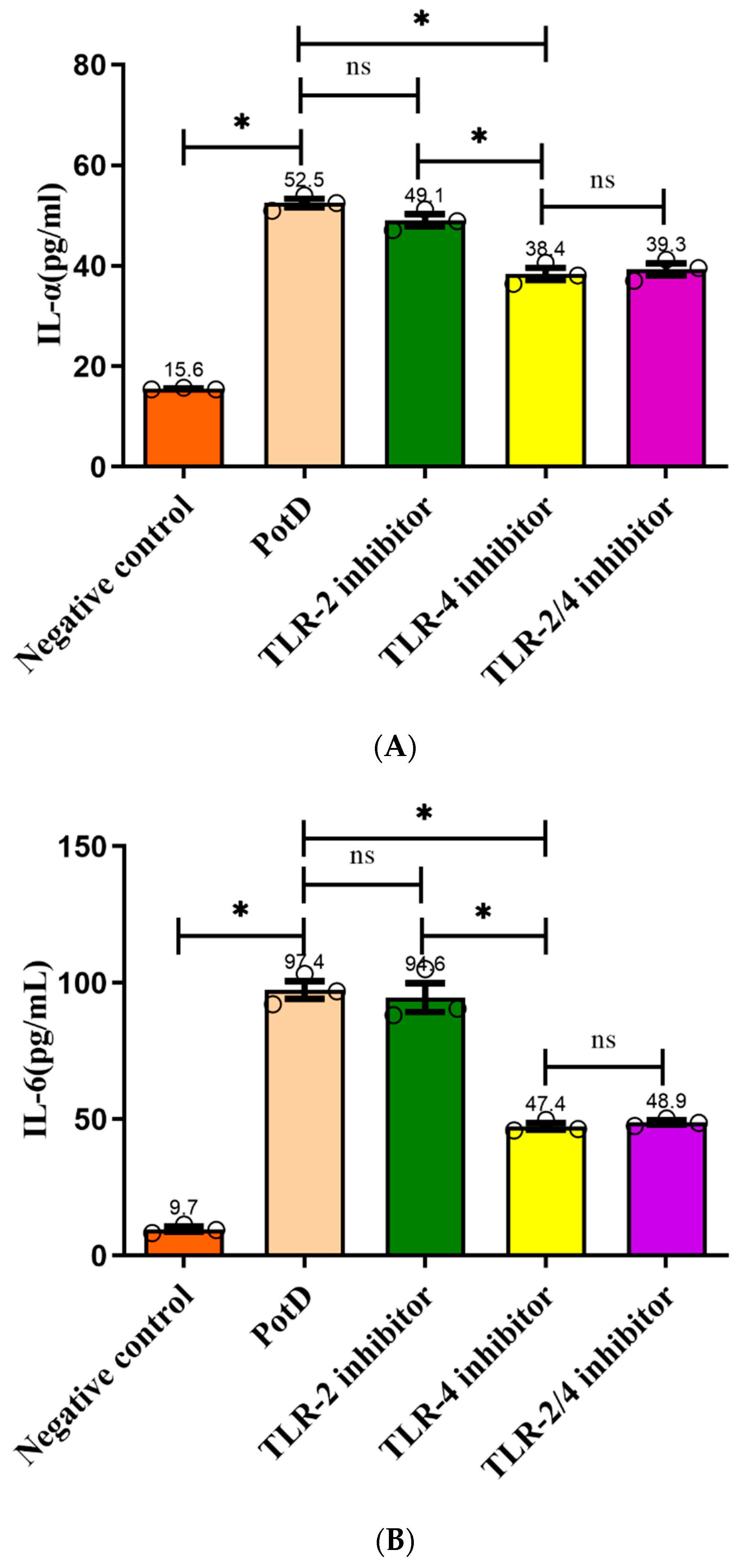
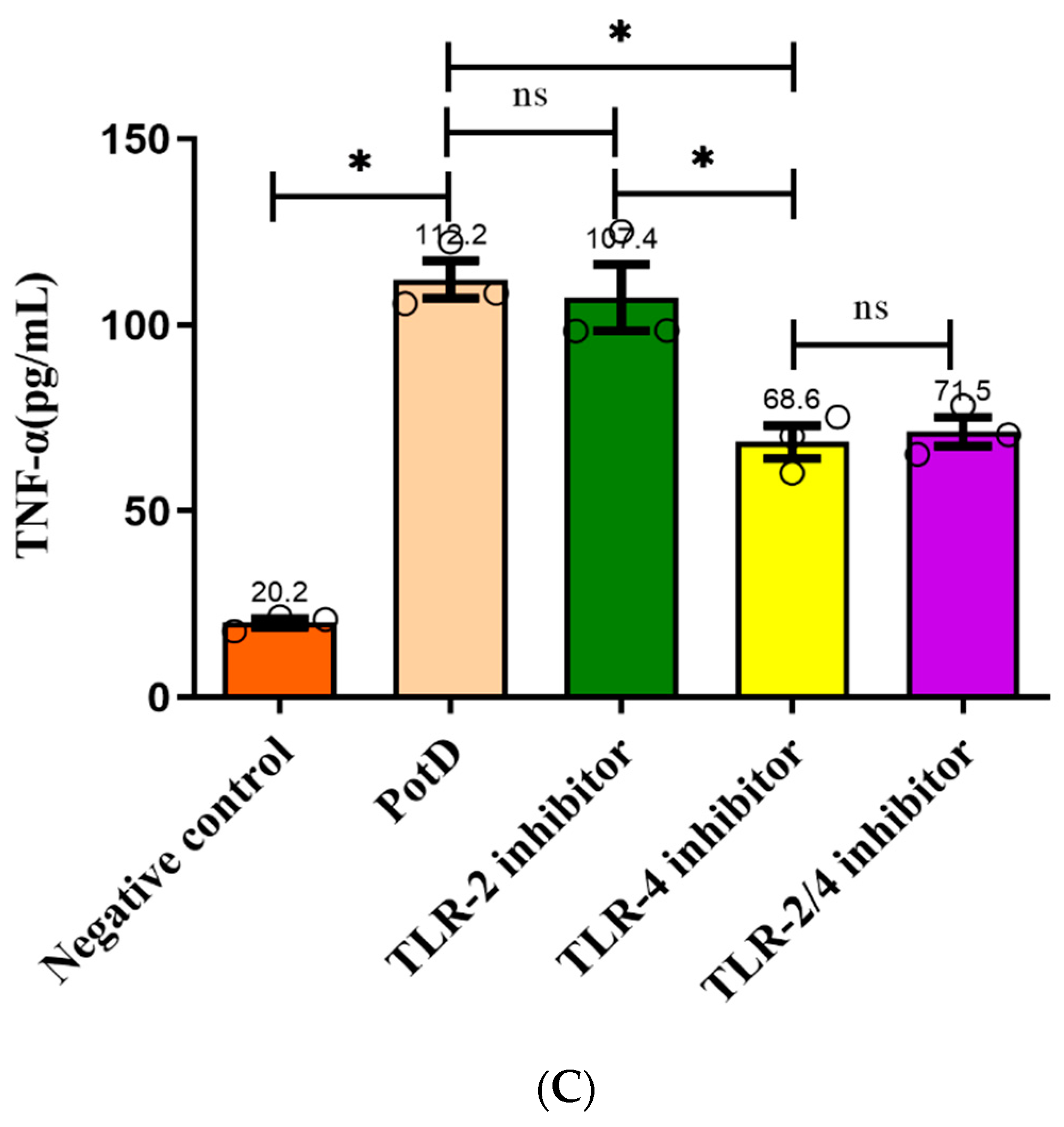
3.7. TLR4, Rather Than TLR2, Participates in Regulatory Role on Expression of Pro-Inflammatory Cytokines in Macrophages
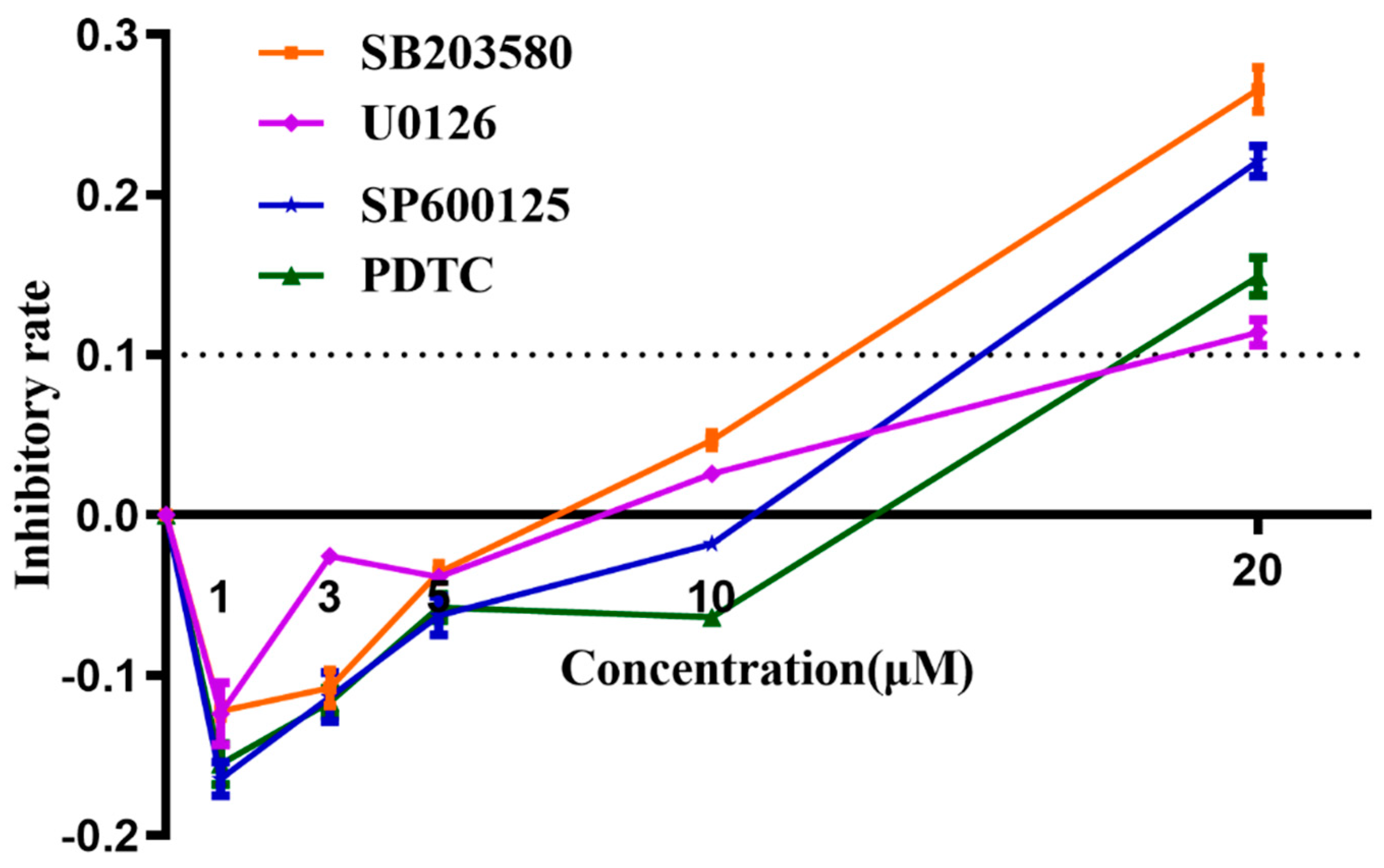
3.8. Determination of Working Concentrations of Inhibitors Using MTT Assay
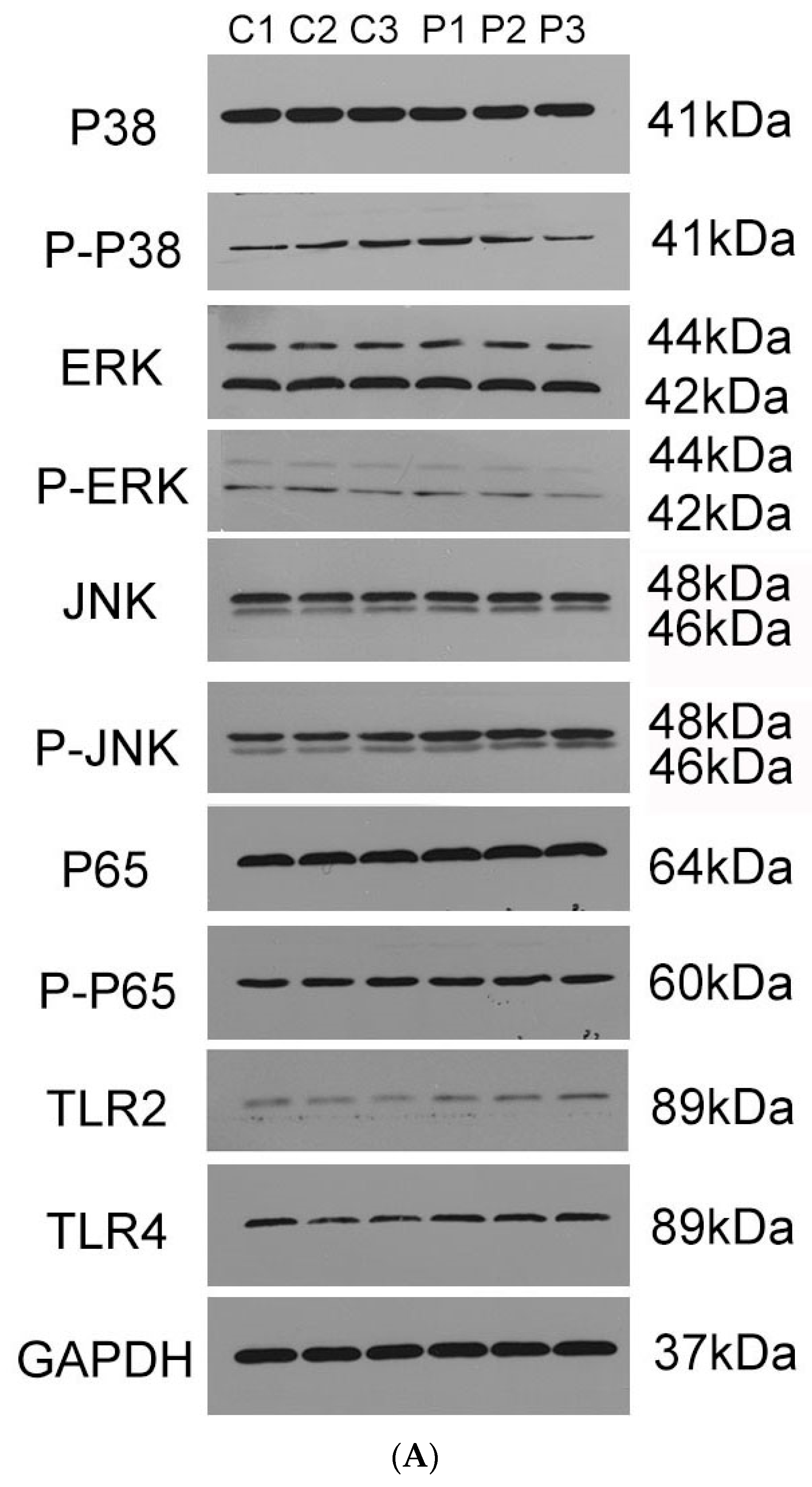
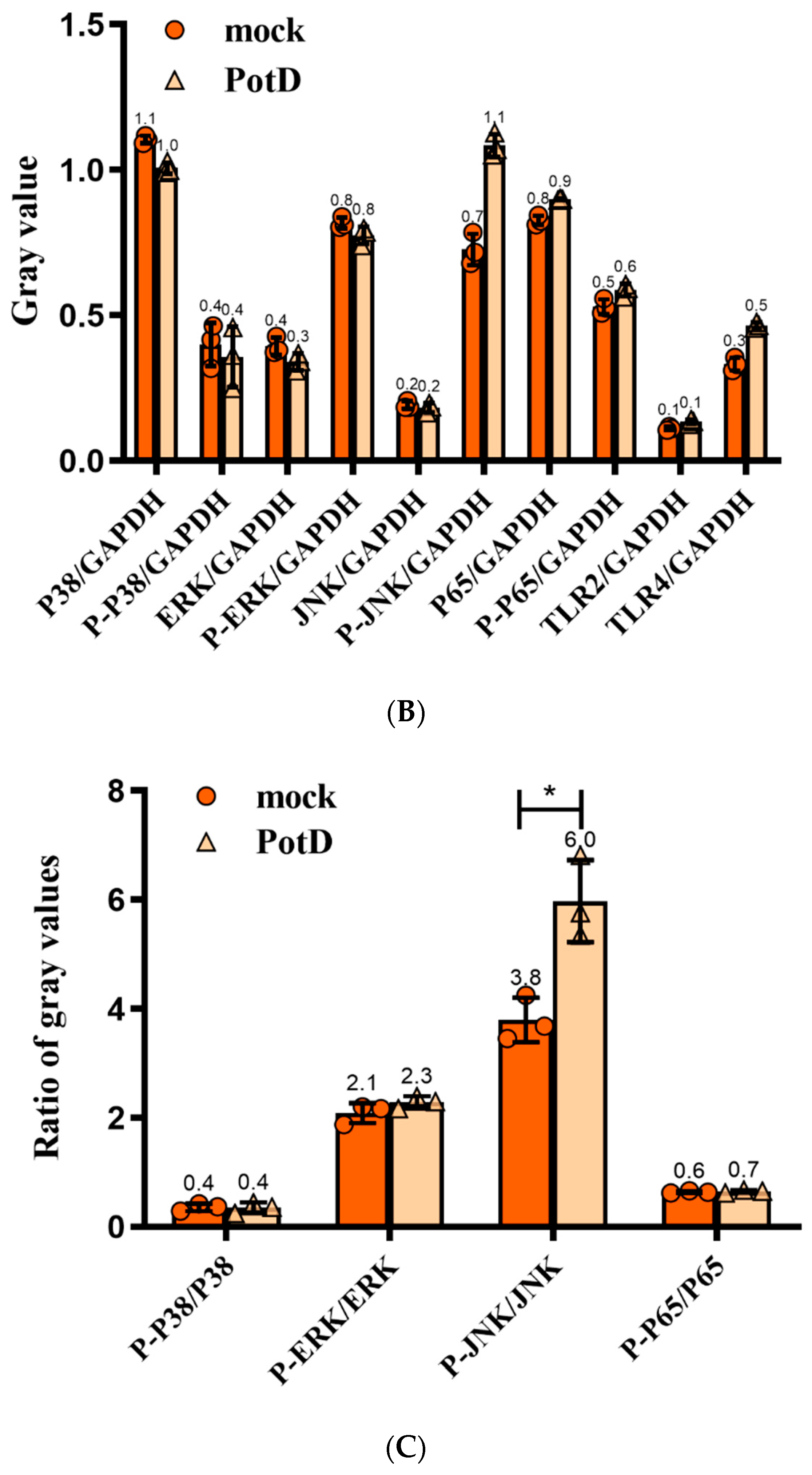
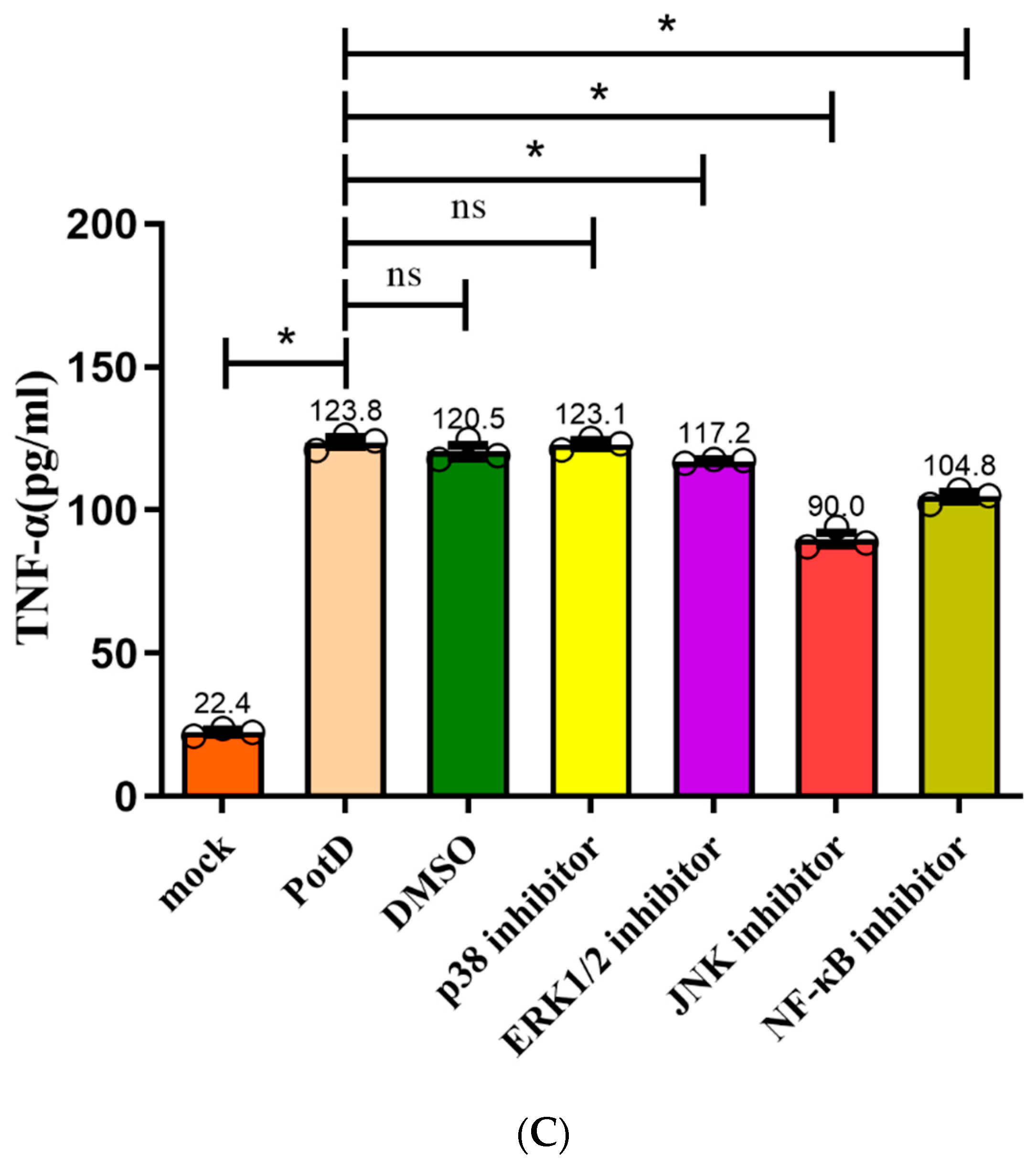
3.9. Production of Pro-Inflammatory Cytokines Mainly through JNK–MAPK and NF–κB Signal Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dai, K.; He, L.; Chang, Y.F.; Cao, S.; Zhao, Q.; Huang, X.; Wu, R.; Huang, Y.; Yan, Q.; Han, X.; et al. Basic Characterization of Natural Transformation in a Highly Transformable Haemophilus parasuis Strain SC1401. Front. Cell. Infect. Microbiol. 2018, 8, 32. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhang, C.; Aragon, V.; Zhou, X.; Zou, M.; Wu, C.; Shen, Z. Investigation of Haemophilus parasuis from healthy pigs in China. Vet. Microbiol. 2019, 231, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, Q.; Li, J.; Lin, X.; Huang, X.; Fang, B. Epidemiology of Haemophilus parasuis isolates from pigs in China using serotyping, antimicrobial susceptibility, biofilm formation and ERIC-PCR genotyping. PeerJ 2018, 6, e5040. [Google Scholar] [CrossRef] [PubMed]
- Dai, K.; Yang, Z.; Chang, Y.F.; He, L.; Cao, S.; Zhao, Q.; Huang, X.; Wu, R.; Huang, Y.; Yan, Q.; et al. Construction of targeted and integrative promoter-reporter plasmids pDK-K and pDK-G to measure gene expression activity in Haemophilus parasuis. Microb. Pathog. 2019, 103565. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Zhou, N.; An, J.; Zhang, J.; Wang, M.; Li, Y.; Jiang, P. Haemophilus parasuis infection in 3D4/21 cells induces autophagy through the AMPK pathway. Cell. Microbiol. 2019, 21, e13031. [Google Scholar] [CrossRef]
- Pires Espindola, J.; Balbinott, N.; Trevisan Gressler, L.; Machado, G.; Silene Klein, C.; Rebelatto, R.; Gutierrez Martin, C.B.; Kreutz, L.C.; Schryvers, A.B.; Frandoloso, R. Molecular serotyping of clinical strains of Haemophilus (Glaesserella) parasuis brings new insights regarding Glasser’s disease outbreaks in Brazil. PeerJ 2019, 7, e6817. [Google Scholar] [CrossRef]
- Zhao, Y.; Guo, L.; Li, J.; Huang, X.; Fang, B. Characterization of antimicrobial resistance genes in Haemophilus parasuis isolated from pigs in China. PeerJ 2018, 6, e4613. [Google Scholar] [CrossRef]
- Kielstein, P.; Rapp-Gabrielson, V. Designation of 15 serovars of Haemophilus parasuis on the basis of immunodiffusion using heat-stable antigen extracts. J. Clin. Microbiol. 1992, 30, 862. [Google Scholar]
- Spaic, A.; Seinige, D.; Muller, A.; Kehrenberg, C. First report of tetracycline resistance mediated by the tet(O) gene in Haemophilus parasuis. J. Glob. Antimicrob. Resistance 2019, 17, 21–22. [Google Scholar] [CrossRef]
- De la Fuente, A.J.; Tucker, A.W.; Navas, J.; Blanco, M.; Morris, S.J.; Gutierrez-Martin, C.B. Antimicrobial susceptibility patterns of Haemophilus parasuis from pigs in the United Kingdom and Spain. Vet. Microbiol. 2007, 120, 184–191. [Google Scholar] [CrossRef]
- Thomas, T.; Thomas, T.J. Polyamines in cell growth and cell death: Molecular mechanisms and therapeutic applications. Cell. Mol. Life Sci. CMLS 2001, 58, 244–258. [Google Scholar] [CrossRef] [PubMed]
- Wallace, H.M.; Fraser, A.V.; Hughes, A. A perspective of polyamine metabolism. Biochem. J. 2003, 376, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, S.; Matsushima, M.; Saisho, T.; Kashiwagi, K.; Igarashi, K.; Morikawa, K. Crystallization and preliminary X-ray analysis of the primary receptor (PotD) of the polyamine transport system in Escherichia coli. Acta Crystallogr. Sect. D Biol. Crystallogr. 1996, 52, 416–418. [Google Scholar] [CrossRef] [PubMed]
- Brandt, A.M.; Raksajit, W.; Yodsang, P.; Mulo, P.; Incharoensakdi, A.; Salminen, T.A.; Maenpaa, P. Characterization of the substrate-binding PotD subunit in Synechocystis sp. strain PCC 6803. Arch. Microbiol. 2010, 192, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Madeo, F.; Eisenberg, T.; Pietrocola, F.; Kroemer, G. Spermidine in health and disease. Science 2018, 359. [Google Scholar] [CrossRef]
- Kashiwagi, K.; Miyamoto, S.; Nukui, E.; Kobayashi, H.; Igarashi, K. Functions of potA and potD proteins in spermidine-preferential uptake system in Escherichia coli. J. Biol. Chem. 1993, 268, 19358–19363. [Google Scholar]
- Furuchi, T.; Kashiwagi, K.; Kobayashi, H.; Igarashi, K. Characteristics of the gene for a spermidine and putrescine transport system that maps at 15 min on the Escherichia coli chromosome. J. Biol. Chem. 1991, 266, 20928–20933. [Google Scholar]
- Yodsang, P.; Pothipongsa, A.; Maenpaa, P.; Incharoensakdi, A. Involvement of polyamine binding protein D (PotD) of Synechocystis sp. PCC 6803 in spermidine uptake and excretion. Curr. Microbiol. 2014, 69, 417–422. [Google Scholar] [CrossRef]
- Shah, P.; Marquart, M.; Quin, L.R.; Swiatlo, E. Cellular location of polyamine transport protein PotD in Streptococcus pneumoniae. FEMS Microbiol. Lett. 2006, 261, 235–237. [Google Scholar] [CrossRef]
- Kashiwagi, K.; Pistocchi, R.; Shibuya, S.; Sugiyama, S.; Morikawa, K.; Igarashi, K. Spermidine-preferential uptake system in Escherichia coli. Identification of amino acids involved in polyamine binding in PotD protein. J. Biol. Chem. 1996, 271, 12205–12208. [Google Scholar] [CrossRef]
- Urdiales, J.L.; Medina, M.A.; Sanchez-Jimenez, F. Polyamine metabolism revisited. Eur. J. Gastroenterol. Hepatol. 2001, 13, 1015–1019. [Google Scholar] [CrossRef] [PubMed]
- Antognoni, F.; Del Duca, S.; Kuraishi, A.; Kawabe, E.; Fukuchi-Shimogori, T.; Kashiwagi, K.; Igarashi, K. Transcriptional inhibition of the operon for the spermidine uptake system by the substrate-binding protein PotD. J. Biol. Chem. 1999, 274, 1942–1948. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, S.; Matsuo, Y.; Maenaka, K.; Vassylyev, D.G.; Matsushima, M.; Kashiwagi, K.; Igarashi, K.; Morikawa, K. The 1.8-A X-ray structure of the Escherichia coli PotD protein complexed with spermidine and the mechanism of polyamine binding. Protein Sci. Publ. Protein Soc. 1996, 5, 1984–1990. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, Y.; Liu, J.; Liu, H. PotD protein stimulates biofilm formation by Escherichia coli. Biotechnol. Lett. 2013, 35, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Ware, D.; Jiang, Y.; Lin, W.; Swiatlo, E. Involvement of potD in Streptococcus pneumoniae polyamine transport and pathogenesis. Infect. Immun. 2006, 74, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.; Romero, D.G.; Swiatlo, E. Role of polyamine transport in Streptococcus pneumoniae response to physiological stress and murine septicemia. Microb. Pathog. 2008, 45, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.; Swiatlo, E. Immunization with polyamine transport protein PotD protects mice against systemic infection with Streptococcus pneumoniae. Infect. Immun. 2006, 74, 5888–5892. [Google Scholar] [CrossRef]
- Shah, P.; Briles, D.E.; King, J.; Hale, Y.; Swiatlo, E. Mucosal immunization with polyamine transport protein D (PotD) protects mice against nasopharyngeal colonization with Streptococcus pneumoniae. Exp. Biol. Med. 2009, 234, 403–409. [Google Scholar] [CrossRef]
- Nasrallah, G.K.; Abdelhady, H.; Tompkins, N.P.; Carson, K.R.; Garduno, R.A. Deletion of potD, encoding a putative spermidine-binding protein, results in a complex phenotype in Legionella pneumophila. Int. J. Med. Microbiol. IJMM 2014, 304, 703–716. [Google Scholar] [CrossRef]
- Han, K.Y.; Seo, H.S.; Song, J.A.; Ahn, K.Y.; Park, J.S.; Lee, J. Transport proteins PotD and Crr of Escherichia coli, novel fusion partners for heterologous protein expression. Biochim. Biophys. Acta 2007, 1774, 1536–1543. [Google Scholar] [CrossRef]
- Zhang, L.; Wen, Y.; Li, Y.; Wei, X.; Yan, X.; Wen, X.; Wu, R.; Huang, X.; Huang, Y.; Yan, Q.; et al. Comparative proteomic analysis of the membrane proteins of two Haemophilus parasuis strains to identify proteins that may help in habitat adaptation and pathogenesis. Proteome Sci. 2014, 12, 38. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Mazmanian, S.K. Innate immune recognition of the microbiota promotes host-microbial symbiosis. Nat. Immunol. 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Deng, S.L.; Lian, Z.X.; Yu, K. Roles of Toll-Like Receptors in Nitroxidative Stress in Mammals. Cells 2019, 8, 576. [Google Scholar] [CrossRef] [PubMed]
- Motta, V.; Soares, F.; Sun, T.; Philpott, D.J. NOD-like receptors: Versatile cytosolic sentinels. Physiol. Rev. 2015, 95, 149–178. [Google Scholar] [CrossRef]
- Ma, B.; Hua, K.; Zhou, S.; Zhou, H.; Chen, Y.; Luo, R.; Bi, D.; Zhou, R.; He, Q.; Jin, H. Haemophilus parasuis infection activates NOD1/2-RIP2 signaling pathway in PK-15 cells. Dev. Comp. Immunol. 2018, 79, 158–165. [Google Scholar] [CrossRef]
- Kigerl, K.A.; de Rivero Vaccari, J.P.; Dietrich, W.D.; Popovich, P.G.; Keane, R.W. Pattern recognition receptors and central nervous system repair. Exp. Neurol. 2014, 258, 5–16. [Google Scholar] [CrossRef]
- Fischer, S. Pattern Recognition Receptors and Control of Innate Immunity: Role of Nucleic Acids. Curr. Pharm. Biotechnol. 2018, 19, 1203–1209. [Google Scholar] [CrossRef]
- Wang, F.; Wang, Z.; Pu, J.; Xie, X.; Gao, X.; Gu, Y.; Chen, S.; Zhang, J. Oscillating flow promotes inflammation through the TLR2-TAK1-IKK2 signalling pathway in human umbilical vein endothelial cell (HUVECs). Life Sci. 2019, 224, 212–221. [Google Scholar] [CrossRef]
- Chen, G.-Y.; Brown, N.K.; Wu, W.; Khedri, Z.; Yu, H.; Chen, X.; Vlekkert, D.v.d.; D’Azzo, A.; Zheng, P.; Liu, Y. Broad and direct interaction between TLR and Siglec families of pattern recognition receptors and its regulation by Neu1. eLife 2014, 3, e04066. [Google Scholar] [CrossRef]
- Alvarez-Estrada, A.; Rodriguez-Ferri, E.F.; Martinez-Martinez, S.; Alvarez, B.; Fernandez-Caballero, T.; Dominguez, J.; Gutierrez-Martin, C.B. TLR2, Siglec-3 and CD163 expressions on porcine peripheral blood monocytes are increased during sepsis caused by Haemophilus parasuis. Comp. Immunol. Microbiol. Infect. Dis. 2019, 64, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Kim, B.J.; Carmona, E.M.; Cutting, A.S.; Gurney, M.A.; Carlos, C.; Feuer, R.; Prasadarao, N.V.; Doran, K.S. Bacterial Pili exploit integrin machinery to promote immune activation and efficient blood-brain barrier penetration. Nat. Commun. 2011, 2, 462. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhao, Q.; Tian, J.; Chang, Y.F.; Wen, X.; Huang, X.; Wu, R.; Wen, Y.; Yan, Q.; Huang, Y.; et al. Effective Pro-Inflammatory Induced Activity of GALT, a Conserved Antigen in A. Pleuropneumoniae, Improves the Cytokines Secretion of Macrophage via p38, ERK1/2 and JNK MAPKs Signal Pathway. Front. Cell. Infect. Microbiol. 2018, 8, 337. [Google Scholar] [CrossRef] [PubMed]
- Quan, K.; Zhu, Z.; Cao, S.; Zhang, F.; Miao, C.; Wen, X.; Huang, X.; Wen, Y.; Wu, R.; Yan, Q.; et al. Escherichia coli-Derived Outer Membrane Vesicles Deliver Galactose-1-Phosphate Uridyltransferase and Yield Partial Protection against Actinobacillus pleuropneumoniae in Mice. J. Microbiol. Biotechnol. 2018, 28, 2095–2105. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, X.; Xia, X.; Zhu, C.; Qin, W.; Xu, Y.; Hang, B.; Sun, Y.; Chen, S.; Zhang, H.; et al. Antimicrobial Peptide JH-3 Effectively Kills Salmonella enterica Serovar Typhimurium Strain CVCC541 and Reduces Its Pathogenicity in Mice. Probiotics Antimicrob. Proteins 2019. [Google Scholar] [CrossRef]
- Lin, W.H.; Shih, H.C.; Lin, C.F.; Yang, C.Y.; Lin, C.N.; Chiou, M.T. Genotypic analyses and virulence characterization of Glaesserella parasuis isolates from Taiwan. PeerJ 2019, 7, e6960. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, Q.; Wei, H.; Wen, X.; Cao, S.; Huang, X.; Wu, R.; Yan, Q.; Huang, Y.; Wen, Y. Prevalence and seroepidemiology of Haemophilus parasuis in Sichuan province, China. PeerJ 2017, 5, e3379. [Google Scholar] [CrossRef]
- McCaig, W.D.; Loving, C.L.; Hughes, H.R.; Brockmeier, S.L. Characterization and Vaccine Potential of Outer Membrane Vesicles Produced by Haemophilus parasuis. PLoS ONE 2016, 11, e0149132. [Google Scholar] [CrossRef]
- Casero, R.A., Jr.; Murray Stewart, T.; Pegg, A.E. Polyamine metabolism and cancer: Treatments, challenges and opportunities. Nat. Rev. Cancer 2018, 18, 681–695. [Google Scholar] [CrossRef]
- Olsen, M.E.; Cressey, T.N.; Muhlberger, E.; Connor, J.H. Differential Mechanisms for the Involvement of Polyamines and Hypusinated eIF5A in Ebola Virus Gene Expression. J. Virol. 2018, 92. [Google Scholar] [CrossRef]
- Zhou, T.; Wang, P.; Yang, R.; Gu, Z. Polyamines regulating phytic acid degradation in mung bean sprouts. J. Sci. Food Agric. 2018, 98, 3299–3308. [Google Scholar] [CrossRef]
- Cho, H.E.; Kang, M.H. pH gradient-liquid chromatography tandem mass spectrometric assay for determination of underivatized polyamines in cancer cells. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1085, 21–29. [Google Scholar] [CrossRef]
- Igarashi, K.; Kashiwagi, K. Effects of polyamines on protein synthesis and growth of Escherichia coli. J. Biol. Chem. 2018, 293, 18702–18709. [Google Scholar] [CrossRef]
- Converso, T.R.; Goulart, C.; Darrieux, M.; Leite, L.C.C. A protein chimera including PspA in fusion with PotD is protective against invasive pneumococcal infection and reduces nasopharyngeal colonization in mice. Vaccine 2017, 35, 5140–5147. [Google Scholar] [CrossRef]
- Nagashima, H.; Yamaoka, Y. Importance of Toll-like Receptors in Pro-inflammatory and Anti-inflammatory Responses by Helicobacter pylori Infection. Curr. Top. Microbiol. Immunol. 2019, 421, 139–158. [Google Scholar] [CrossRef]
- Das, S.; Chowdhury, B.P.; Goswami, A.; Parveen, S.; Jawed, J.; Pal, N.; Majumdar, S. Mycobacterium indicus pranii (MIP) mediated host protective intracellular mechanisms against tuberculosis infection: Involvement of TLR-4 mediated signaling. Tuberculosis 2016, 101, 201–209. [Google Scholar] [CrossRef]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Primer Sequences (5′→3′) | Products (bp) |
|---|---|---|
| rPotD expression primers | ||
| P1 (potD-pET-F) | cagcaaatgggtcgcggatccAATGACACTGTACATCTTTATACTT | 1015 |
| P2 (potD-pET-R) | ctcgagtgcggccgcaagcttGCTTCGCAGCTTTTAACTCTTG | |
| qPCR primers | ||
| P3(GAPDH-F) | GTGTTCCTACCCCCAATGTG | 189 |
| P4(GAPDH-R) | CATCGAAGGTGGAAGAGTGG | |
| P5(IL-1β-F) | GGGCCTCAAAGGAAAGAATC | 183 |
| P6(IL-1β-R) | TACCAGTTGGGGAACTCTGC | |
| P7(IL-6-F) | GGGACTGATGCTGGTGACAA | 147 |
| P8(IL-6-R) | TCCACGATTTCCCAGAGAACA | |
| P9(TNF-α-F) | CGTCAGCCGATTTGCTATCT | 184 |
| P10(TNF-α-F) | CTTGGGCAGATTGACCTCAG | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, K.; Ma, X.; Yang, Z.; Chang, Y.-F.; Cao, S.; Zhao, Q.; Huang, X.; Wu, R.; Huang, Y.; Yan, Q.; et al. Polyamine Transport Protein PotD Protects Mice against Haemophilus parasuis and Elevates the Secretion of Pro-Inflammatory Cytokines of Macrophage via JNK–MAPK and NF–κB Signal Pathways through TLR4. Vaccines 2019, 7, 216. https://doi.org/10.3390/vaccines7040216
Dai K, Ma X, Yang Z, Chang Y-F, Cao S, Zhao Q, Huang X, Wu R, Huang Y, Yan Q, et al. Polyamine Transport Protein PotD Protects Mice against Haemophilus parasuis and Elevates the Secretion of Pro-Inflammatory Cytokines of Macrophage via JNK–MAPK and NF–κB Signal Pathways through TLR4. Vaccines. 2019; 7(4):216. https://doi.org/10.3390/vaccines7040216
Chicago/Turabian StyleDai, Ke, Xiaoyu Ma, Zhen Yang, Yung-Fu Chang, Sanjie Cao, Qin Zhao, Xiaobo Huang, Rui Wu, Yong Huang, Qigui Yan, and et al. 2019. "Polyamine Transport Protein PotD Protects Mice against Haemophilus parasuis and Elevates the Secretion of Pro-Inflammatory Cytokines of Macrophage via JNK–MAPK and NF–κB Signal Pathways through TLR4" Vaccines 7, no. 4: 216. https://doi.org/10.3390/vaccines7040216
APA StyleDai, K., Ma, X., Yang, Z., Chang, Y.-F., Cao, S., Zhao, Q., Huang, X., Wu, R., Huang, Y., Yan, Q., Han, X., Ma, X., Wen, X., & Wen, Y. (2019). Polyamine Transport Protein PotD Protects Mice against Haemophilus parasuis and Elevates the Secretion of Pro-Inflammatory Cytokines of Macrophage via JNK–MAPK and NF–κB Signal Pathways through TLR4. Vaccines, 7(4), 216. https://doi.org/10.3390/vaccines7040216