
Dissolving Microneedles Loaded with Nanoparticle Formulation of Respiratory Syncytial Virus Fusion Protein Virus-like Particles (F-VLPs) Elicits Cellular and Humoral Immune Responses

 ,
,  ,
,  ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Formulation of F-VLP-Loaded Polymeric Nanoparticles
2.2. Formulation of Dissolving Microneedles
2.3. Characterization of the Nanoparticle-Loaded Dissolving Microneedles
2.4. Immunization with Nanoparticle-Loaded Dissolving MNs and RSV Challenge
2.5. Measurement of Antigen-Specific Antibodies in Serum and Lung Homogenates Using ELISA
2.6. Measurement of T Cells in the Immune Organs Using Flow Cytometry
2.7. Statistical Analysis
3. Results
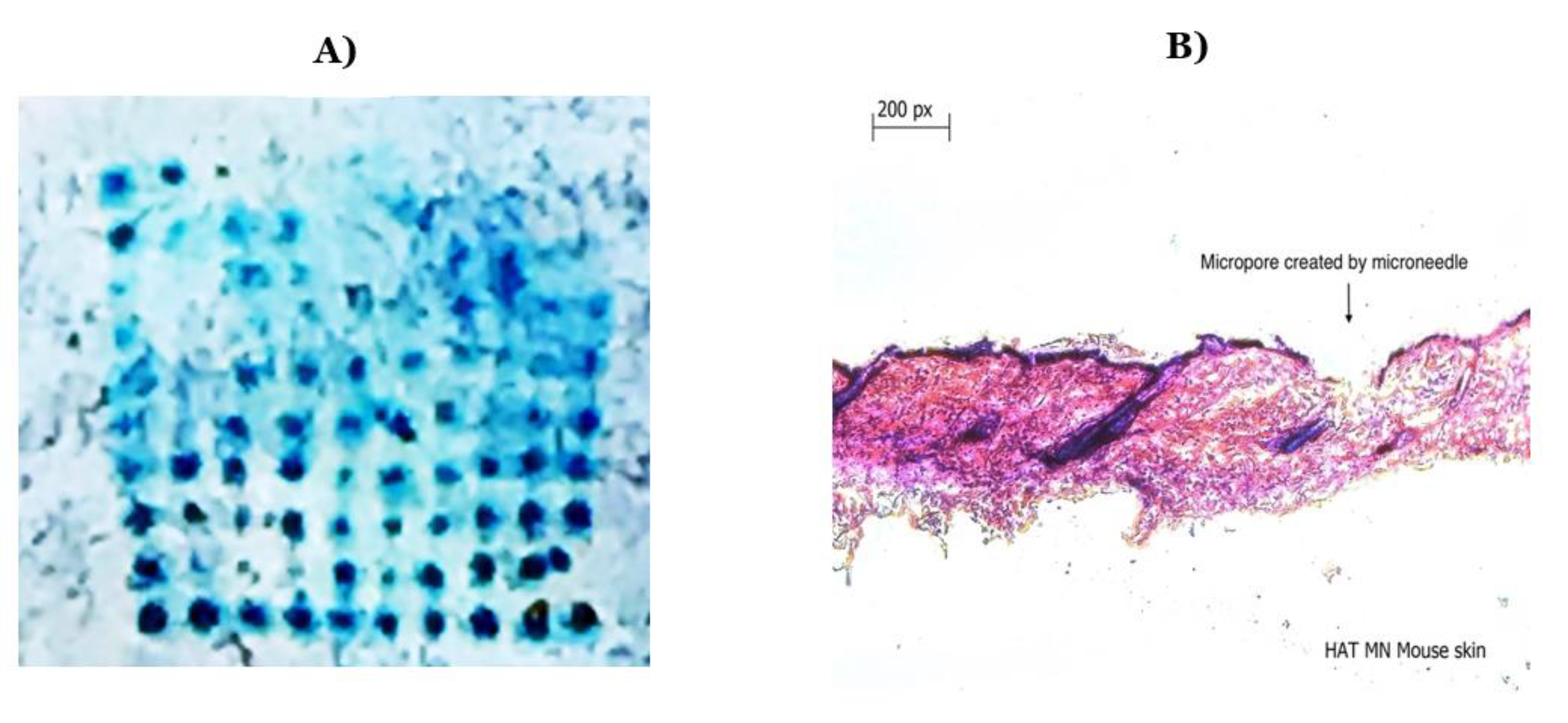
3.1. Characterization of Nanoparticle-Loaded Microneedles
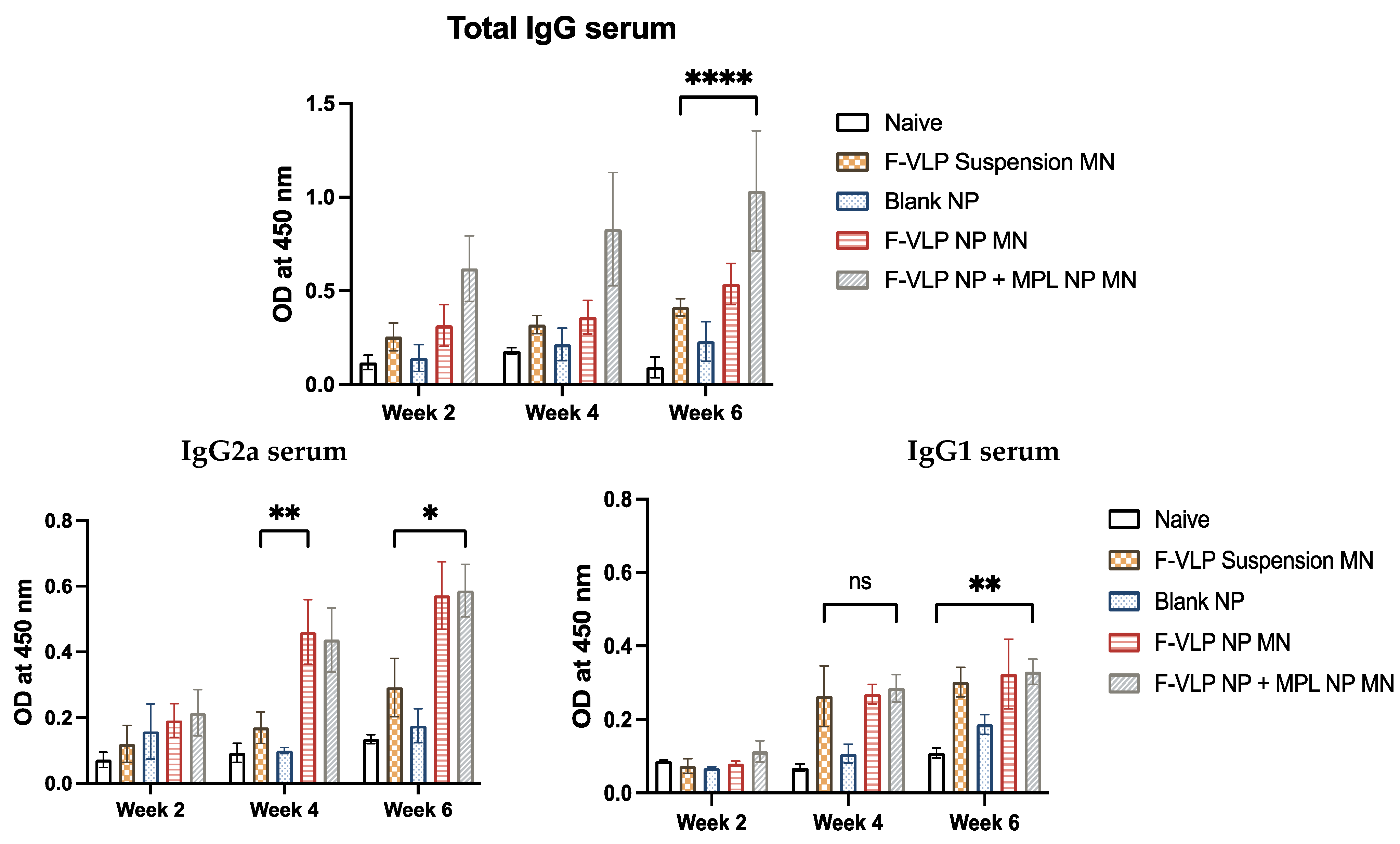
3.2. RSV F-Protein-Specific IgG and Subtype Antibodies Observed in the Serum of Vaccinated Mice
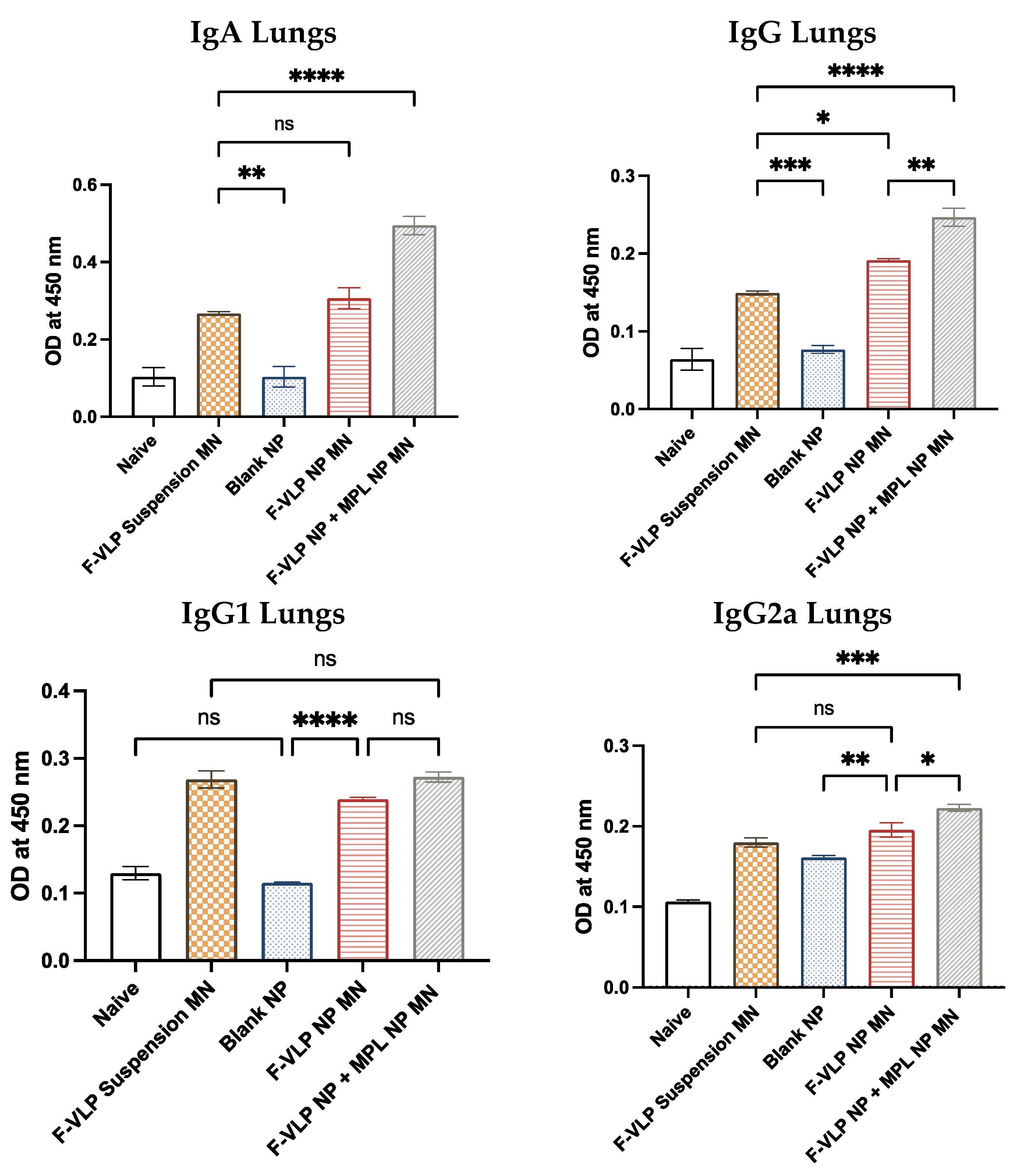
3.3. RSV F-Protein-Specific IgG, IgG Subtypes, and IgA Antibodies Observed in the Lung Homogenates of Vaccinated Mice after RSV Challenge
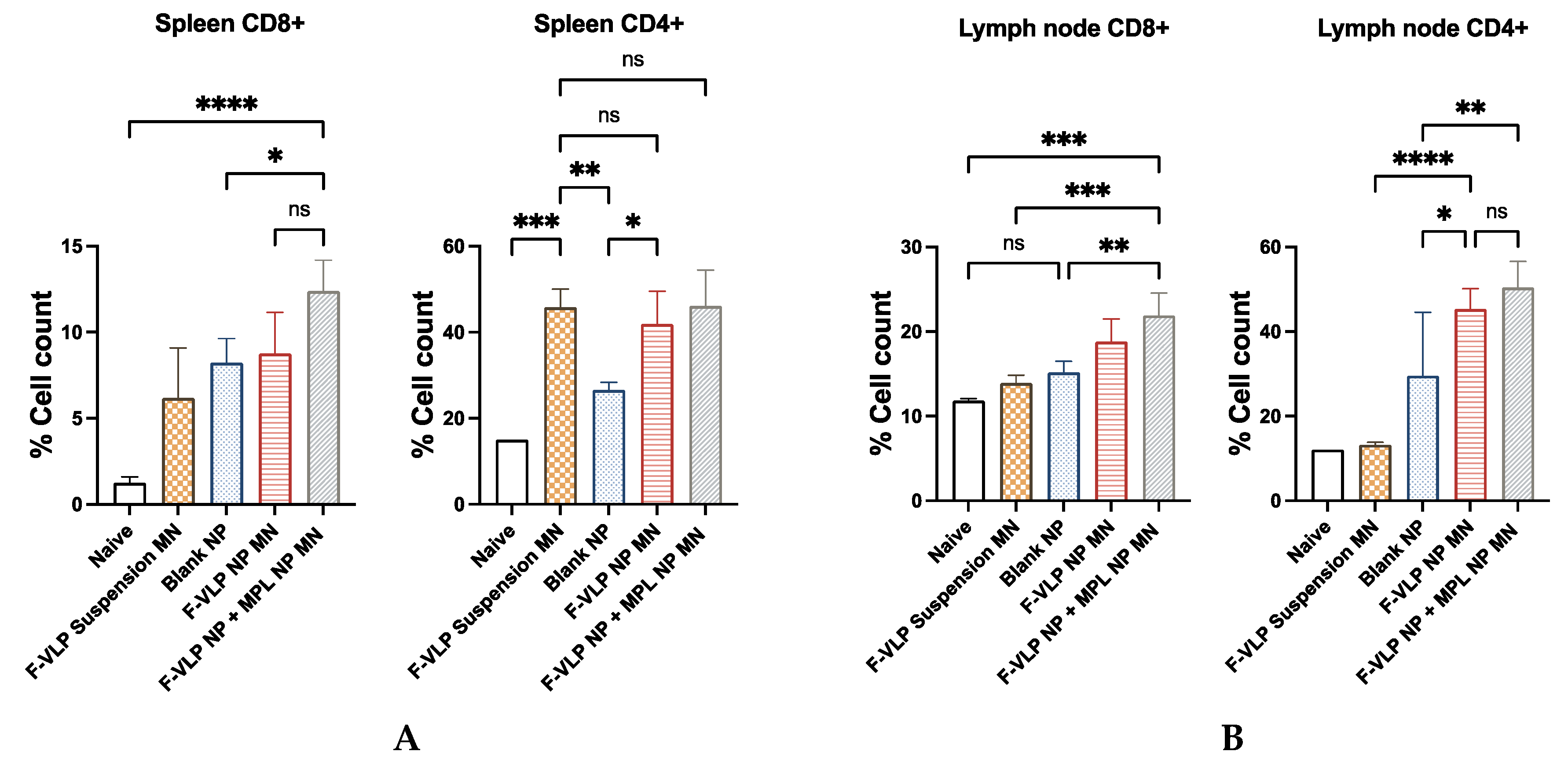
3.4. Enhanced Expression of T-Cell Markers Observed in Spleen and Lymph Nodes of Vaccinated Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Koltai, M.; Krauer, F.; Hodgson, D.; van Leeuwen, E.; Treskova-Schwarzbach, M.; Jit, M.; Flasche, S. Determinants of RSV Epidemiology Following Suppression through Pandemic Contact Restrictions. Epidemics 2022, 40, 100614. [Google Scholar] [CrossRef] [PubMed]
- Hansen, C.L.; Chaves, S.S.; Demont, C.; Viboud, C. Mortality Associated With Influenza and Respiratory Syncytial Virus in the US, 1999–2018. JAMA Netw. Open 2022, 5, e220527. [Google Scholar] [CrossRef]
- Mosscrop, L.G.; Williams, T.C.; Tregoning, J.S. Respiratory Syncytial Virus after the SARS-CoV-2 Pandemic—What Next? Nat. Rev. Immunol. 2022, 22, 589–590. [Google Scholar] [CrossRef] [PubMed]
- Olson, M.R.; Varga, S.M. CD8 T Cells Inhibit Respiratory Syncytial Virus (RSV) Vaccine-Enhanced Disease. J. Immunol. 2007, 179, 5415–5424. [Google Scholar] [CrossRef] [PubMed]
- Connors, M.; Kulkarni, A.B.; Firestone, C.Y.; Holmes, K.L.; Morse, H.C.; Sotnikov, A.V.; Murphy, B.R. Pulmonary Histopathology Induced by Respiratory Syncytial Virus (RSV) Challenge of Formalin-Inactivated RSV-Immunized BALB/c Mice Is Abrogated by Depletion of CD4+ T Cells. J. Virol. 1992, 66, 7444–7451. [Google Scholar] [CrossRef]
- Zhan, X.; Hurwitz, J.L.; Krishnamurthy, S.; Takimoto, T.; Boyd, K.; Scroggs, R.A.; Surman, S.; Portner, A.; Slobod, K.S. Respiratory Syncytial Virus (RSV) Fusion Protein Expressed by Recombinant Sendai Virus Elicits B-Cell and T-Cell Responses in Cotton Rats and Confers Protection against RSV Subtypes A and B. Vaccine 2007, 25, 8782–8793. [Google Scholar] [CrossRef] [PubMed]
- Gu, P.; Wusiman, A.; Zhang, Y.; Liu, Z.; Bo, R.; Hu, Y.; Liu, J.; Wang, D. Rational Design of PLGA Nanoparticle Vaccine Delivery Systems To Improve Immune Responses. Mol. Pharm. 2019, 16, 5000–5012. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.L.; Soema, P.C.; Slütter, B.; Ossendorp, F.; Jiskoot, W. PLGA Particulate Delivery Systems for Subunit Vaccines: Linking Particle Properties to Immunogenicity. Hum. Vaccines Immunother. 2016, 12, 1056–1069. [Google Scholar] [CrossRef]
- Kazzaz, J.; Singh, M.; Ugozzoli, M.; Chesko, J.; Soenawan, E.; O’Hagan, D.T. Encapsulation of the Immune Potentiators MPL and RC529 in PLG Microparticles Enhances Their Potency. J. Control. Release 2006, 110, 566–573. [Google Scholar] [CrossRef]
- Demento, S.L.; Cui, W.; Criscione, J.M.; Stern, E.; Tulipan, J.; Kaech, S.M.; Fahmy, T.M. Role of Sustained Antigen Release from Nanoparticle Vaccines in Shaping the T Cell Memory Phenotype. Biomaterials 2012, 33, 4957–4964. [Google Scholar] [CrossRef]
- Kasturi, S.P.; Skountzou, I.; Albrecht, R.A.; Koutsonanos, D.; Hua, T.; Nakaya, H.I.; Ravindran, R.; Stewart, S.; Alam, M.; Kwissa, M.; et al. Programming the Magnitude and Persistence of Antibody Responses with Innate Immunity. Nature 2011, 470, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Cluff, C.W. Monophosphoryl Lipid A (MPL) as an Adjuvant for Anti-Cancer Vaccines: Clinical Results; Landes Bioscience: Austin, TX, USA, 2013. [Google Scholar]
- Park, S.; Lee, Y.; Kwon, Y.-M.; Lee, Y.-T.; Kim, K.-H.; Ko, E.-J.; Jung, J.H.; Song, M.; Graham, B.; Prausnitz, M.R.; et al. Vaccination by Microneedle Patch with Inactivated Respiratory Syncytial Virus and Monophosphoryl Lipid A Enhances the Protective Efficacy and Diminishes Inflammatory Disease after Challenge. PLoS ONE 2018, 13, e0205071. [Google Scholar] [CrossRef] [PubMed]
- Hassan, J.; Haigh, C.; Ahmed, T.; Uddin, M.J.; Das, D.B. Potential of Microneedle Systems for COVID-19 Vaccination: Current Trends and Challenges. Pharmaceutics 2022, 14, 1066. [Google Scholar] [CrossRef] [PubMed]
- Menon, I.; Bagwe, P.; Gomes, K.B.; Bajaj, L.; Gala, R.; Uddin, M.N.; D’Souza, M.J.; Zughaier, S.M. Microneedles: A New Generation Vaccine Delivery System. Micromachines 2021, 12, 435. [Google Scholar] [CrossRef]
- Neuzil, K.M. Progress toward a Respiratory Syncytial Virus Vaccine. Clin. Vaccine Immunol. 2016, 23, 186–188. [Google Scholar] [CrossRef]
- Qiu, X.; Xu, S.; Lu, Y.; Luo, Z.; Yan, Y.; Wang, C.; Ji, J. Development of MRNA Vaccines against Respiratory Syncytial Virus (RSV). Cytokine Growth Factor Rev. 2022, 68, 37–53. [Google Scholar] [CrossRef]
- Braz Gomes, K.; D’Souza, B.; Vijayanand, S.; Menon, I.; D’Souza, M.J. A Dual-Delivery Platform for Vaccination Using Antigen-Loaded Nanoparticles in Dissolving Microneedles. Int. J. Pharm. 2022, 613, 121393. [Google Scholar] [CrossRef]
- Menon, I.; Kang, S.M.; Braz Gomes, K.; Uddin, M.N.; D’Souza, M. Laser-Assisted Intradermal Delivery of a Microparticle Vaccine for Respiratory Syncytial Virus Induces a Robust Immune Response. Vaccine 2023, 41, 1209–1222. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, H.; Guo, B.; Wang, B.; Wan, Z.; Zhang, Y.; Sun, L.; Yang, F. Microneedle-Based Two-Step Transdermal Delivery of Langerhans Cell-Targeting Immunoliposomes Induces a Th1-Biased Immune Response. Eur. J. Pharm. Biopharm. 2022, 177, 68–80. [Google Scholar] [CrossRef]
- Kim, Y.-C.; Park, J.-H.; Prausnitz, M.R. Microneedles for Drug and Vaccine Delivery. Adv. Drug Deliv. Rev. 2012, 64, 1547–1568. [Google Scholar] [CrossRef]
- Patil, S.; Vijayanand, S.; Joshi, D.; Menon, I.; Gomes, K.B.; Kale, A.; Bagwe, P.; Yacoub, S.; Uddin, M.N.; D’Souza, M.J. Subunit Microparticulate Vaccine Delivery Using Microneedles Trigger Significant SARS-Spike-Specific Humoral and Cellular Responses in a Preclinical Murine Model. Int. J. Pharm. 2023, 632, 122583. [Google Scholar] [CrossRef] [PubMed]
- Vijayanand, S.; Patil, S.; Joshi, D.; Menon, I.; Braz Gomes, K.; Kale, A.; Bagwe, P.; Yacoub, S.; Uddin, M.N.; D’Souza, M.J. Microneedle Delivery of an Adjuvanted Microparticulate Vaccine Induces High Antibody Levels in Mice Vaccinated against Coronavirus. Vaccines 2022, 10, 1491. [Google Scholar] [CrossRef] [PubMed]
- Kale, A.; Joshi, D.; Menon, I.; Bagwe, P.; Patil, S.; Vijayanand, S.; Braz Gomes, K.; Uddin, M.N.; D’Souza, M.J. Zika Vaccine Microparticles (MPs)-Loaded Dissolving Microneedles (MNs) Elicit a Significant Immune Response in a Pre-Clinical Murine Model. Vaccines 2023, 11, 583. [Google Scholar] [CrossRef] [PubMed]
- Menon, I.; Kang, S.M.; D’Souza, M. Nanoparticle Formulation of the Fusion Protein Virus like Particles of Respiratory Syncytial Virus Stimulates Enhanced in Vitro Antigen Presentation and Autophagy. Int. J. Pharm. 2022, 623, 121919. [Google Scholar] [CrossRef]
- Quan, F.-S.; Kim, Y.; Lee, S.; Yi, H.; Kang, S.-M.; Bozja, J.; Moore, M.L.; Compans, R.W. Viruslike Particle Vaccine Induces Protection against Respiratory Syncytial Virus Infection in Mice. J. Infect. Dis. 2011, 204, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yue, F.; Kuang, S. Muscle Histology Characterization Using H&E Staining and Muscle Fiber Type Classification Using Immunofluorescence Staining. Bio-Protocol 2017, 7, e2279. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Kwon, Y.-M.; Hwang, H.S.; Lee, Y.-N.; Ko, E.-J.; Yoo, S.-E.; Kim, M.-C.; Kim, K.-H.; Cho, M.-K.; Lee, Y.-T.; et al. Baculovirus-Expressed Virus-like Particle Vaccine in Combination with DNA Encoding the Fusion Protein Confers Protection against Respiratory Syncytial Virus. Vaccine 2014, 32, 5866–5874. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-T.; Kim, K.-H.; Hwang, H.S.; Lee, Y.; Kwon, Y.-M.; Ko, E.-J.; Jung, Y.-J.; Lee, Y.-N.; Kim, M.-C.; Kang, S.-M. Innate and Adaptive Cellular Phenotypes Contributing to Pulmonary Disease in Mice after Respiratory Syncytial Virus Immunization and Infection. Virology 2015, 485, 36–46. [Google Scholar] [CrossRef]
- Quan, F.-S.; Huang, C.; Compans, R.W.; Kang, S.-M. Virus-like Particle Vaccine Induces Protective Immunity against Homologous and Heterologous Strains of Influenza Virus. J. Virol. 2007, 81, 3514–3524. [Google Scholar] [CrossRef]
- Sigma-Aldrich. PARAFILM® M Roll Size 4 in. × 125 ft|Sigma-Aldrich. Available online: http://www.sigmaaldrich.com/ (accessed on 1 February 2022).
- Wei, J.C.J.; Edwards, G.A.; Martin, D.J.; Huang, H.; Crichton, M.L.; Kendall, M.A.F. Allometric Scaling of Skin Thickness, Elasticity, Viscoelasticity to Mass for Micro-Medical Device Translation: From Mice, Rats, Rabbits, Pigs to Humans. Sci. Rep. 2017, 7, 15885. [Google Scholar] [CrossRef]
- Koerner, J.; Horvath, D.; Groettrup, M. Harnessing Dendritic Cells for Poly (D,L-Lactide-Co-Glycolide) Microspheres (PLGA MS)—Mediated Anti-Tumor Therapy. Front. Immunol. 2019, 10, 707. [Google Scholar] [CrossRef] [PubMed]
- Pulendran, B.; Arunachalam, P.S.; O’Hagan, D.T. Emerging Concepts in the Science of Vaccine Adjuvants. Nat. Rev. Drug Discov. 2021, 20, 454–475. [Google Scholar] [CrossRef] [PubMed]
- Russell, C.D.; Unger, S.A.; Walton, M.; Schwarze, J. The Human Immune Response to Respiratory Syncytial Virus Infection. Clin. Microbiol. Rev. 2017, 30, 481–502. [Google Scholar] [CrossRef] [PubMed]
- Boopathy, A.V.; Mandal, A.; Kulp, D.W.; Menis, S.; Bennett, N.R.; Watkins, H.C.; Wang, W.; Martin, J.T.; Thai, N.T.; He, Y.; et al. Enhancing Humoral Immunity via Sustained-Release Implantable Microneedle Patch Vaccination. Proc. Natl. Acad. Sci. USA 2019, 116, 16473–16478. [Google Scholar] [CrossRef]
- Du, G.; Hathout, R.M.; Nasr, M.; Nejadnik, M.R.; Tu, J.; Koning, R.I.; Koster, A.J.; Slütter, B.; Kros, A.; Jiskoot, W.; et al. Intradermal Vaccination with Hollow Microneedles: A Comparative Study of Various Protein Antigen and Adjuvant Encapsulated Nanoparticles. J. Control. Release 2017, 266, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-C.; Prausnitz, M.R. Enabling Skin Vaccination Using New Delivery Technologies. Drug Deliv. Transl. Res. 2011, 1, 7–12. [Google Scholar] [CrossRef]
- Pielenhofer, J.; Sohl, J.; Windbergs, M.; Langguth, P.; Radsak, M.P. Current Progress in Particle-Based Systems for Transdermal Vaccine Delivery. Front. Immunol. 2020, 11, 266. [Google Scholar] [CrossRef]
- Macpherson, A.J.; McCoy, K.D.; Johansen, F.-E.; Brandtzaeg, P. The Immune Geography of IgA Induction and Function. Mucosal Immunol. 2008, 1, 11–22. [Google Scholar] [CrossRef]
- Boyaka, P.N. Inducing Mucosal IgA: A Challenge for Vaccine Adjuvants and Delivery Systems. J. Immunol. 2017, 199, 9–16. [Google Scholar] [CrossRef]
- Lycke, N. Recent Progress in Mucosal Vaccine Development: Potential and Limitations. Nat. Rev. Immunol. 2012, 12, 592–605. [Google Scholar] [CrossRef]
- Nizard, M.; Diniz, M.O.; Roussel, H.; Tran, T.; Ferreira, L.C.; Badoual, C.; Tartour, E. Mucosal Vaccines. Hum. Vaccines Immunother. 2014, 10, 2175–2187. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formulation Number | Hyaluronic Acid (% w/v) | Trehalose (% w/v) | Comments | Time Needed to Dissolve Needles (min) |
|---|---|---|---|---|
| #1 | 5 | 10 | Needles brittle, soft, pliable base, difficult to demold | 15 |
| #2 | 5 | 20 | Needles brittle, pliable base, difficult to demold | 15 |
| #3 | 5 | 30 | Needles brittle, soft, pliable base, difficult to demold | 15 |
| #4 | 10 | 5 | Needles formed, base nonpliable, easy to demold | 10 |
| #5 | 10 | 10 | Needles formed, base nonpliable, difficult to demold | 20 |
| #6 | 15 | 5 | Needles formed, base nonpliable, easy to demold | 15 |
| #7 | 15 | 10 | Needles formed, base nonpliable, easy to demold | 20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menon, I.; Patil, S.; Bagwe, P.; Vijayanand, S.; Kale, A.; Braz Gomes, K.; Kang, S.M.; D’Souza, M. Dissolving Microneedles Loaded with Nanoparticle Formulation of Respiratory Syncytial Virus Fusion Protein Virus-like Particles (F-VLPs) Elicits Cellular and Humoral Immune Responses. Vaccines 2023, 11, 866. https://doi.org/10.3390/vaccines11040866
Menon I, Patil S, Bagwe P, Vijayanand S, Kale A, Braz Gomes K, Kang SM, D’Souza M. Dissolving Microneedles Loaded with Nanoparticle Formulation of Respiratory Syncytial Virus Fusion Protein Virus-like Particles (F-VLPs) Elicits Cellular and Humoral Immune Responses. Vaccines. 2023; 11(4):866. https://doi.org/10.3390/vaccines11040866
Chicago/Turabian StyleMenon, Ipshita, Smital Patil, Priyal Bagwe, Sharon Vijayanand, Akanksha Kale, Keegan Braz Gomes, Sang Moo Kang, and Martin D’Souza. 2023. "Dissolving Microneedles Loaded with Nanoparticle Formulation of Respiratory Syncytial Virus Fusion Protein Virus-like Particles (F-VLPs) Elicits Cellular and Humoral Immune Responses" Vaccines 11, no. 4: 866. https://doi.org/10.3390/vaccines11040866
APA StyleMenon, I., Patil, S., Bagwe, P., Vijayanand, S., Kale, A., Braz Gomes, K., Kang, S. M., & D’Souza, M. (2023). Dissolving Microneedles Loaded with Nanoparticle Formulation of Respiratory Syncytial Virus Fusion Protein Virus-like Particles (F-VLPs) Elicits Cellular and Humoral Immune Responses. Vaccines, 11(4), 866. https://doi.org/10.3390/vaccines11040866