
Biological Characteristics of Feline Calicivirus Epidemic Strains in China and Screening of Broad-Spectrum Protective Vaccine Strains
 , , ,
, , ,
Abstract
:1. Introduction
2. Methods
2.1. Cells and Virus
2.2. Detection of FCV Whole-Genome Sequence Mutation
2.3. Phylogenetic Analyses
2.4. FCV Virus Titer Assay
2.5. Immunogenicity Assay of FCV Strains
2.5.1. Purification and Inactivation of FCV Virus
2.5.2. Antibody Titer Determination using Virus Neutralization Test
2.5.3. Immunogenicity Assay of FCV Strains
2.6. Determination of Dynamic Growth Curve of FCV Vaccine Strains
2.7. Pathogenicity Determination of FCV-HB7 and FCV-HB10 Strains in Cats
2.8. Cat Protective Efficacy Assay
2.9. Hematoxylin and Eosin (H&E) Staining
2.10. Statistical Analysis
3. Results
3.1. Homology Analysis of FCV Whole-Genome Sequence
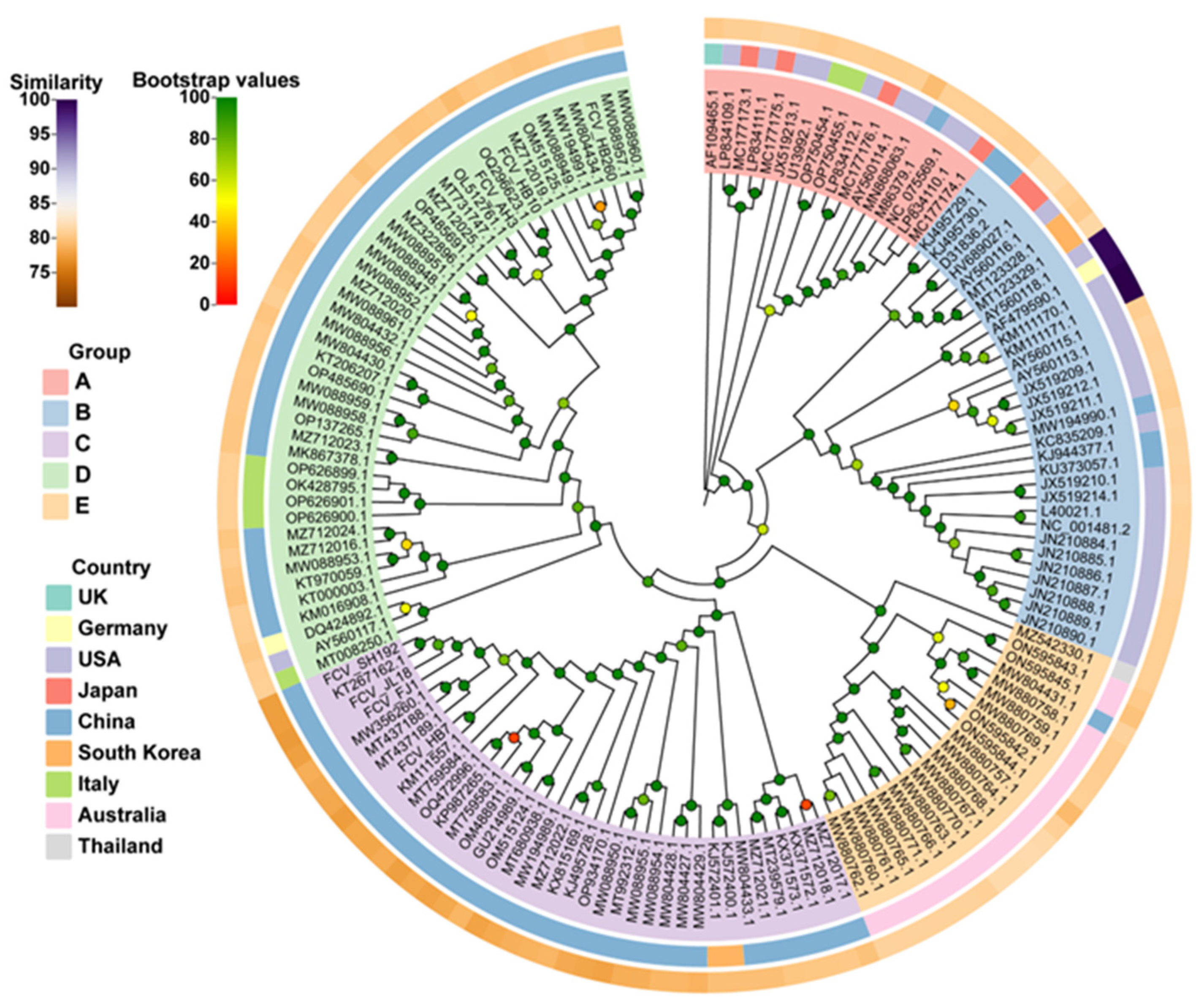
3.2. Phylogenetic Analyses of FCV Isolates
3.3. Determination of Virus Titer and Immunogenicity of FCV Strains
3.4. Growth Kinetics of FCV-Representative Strains
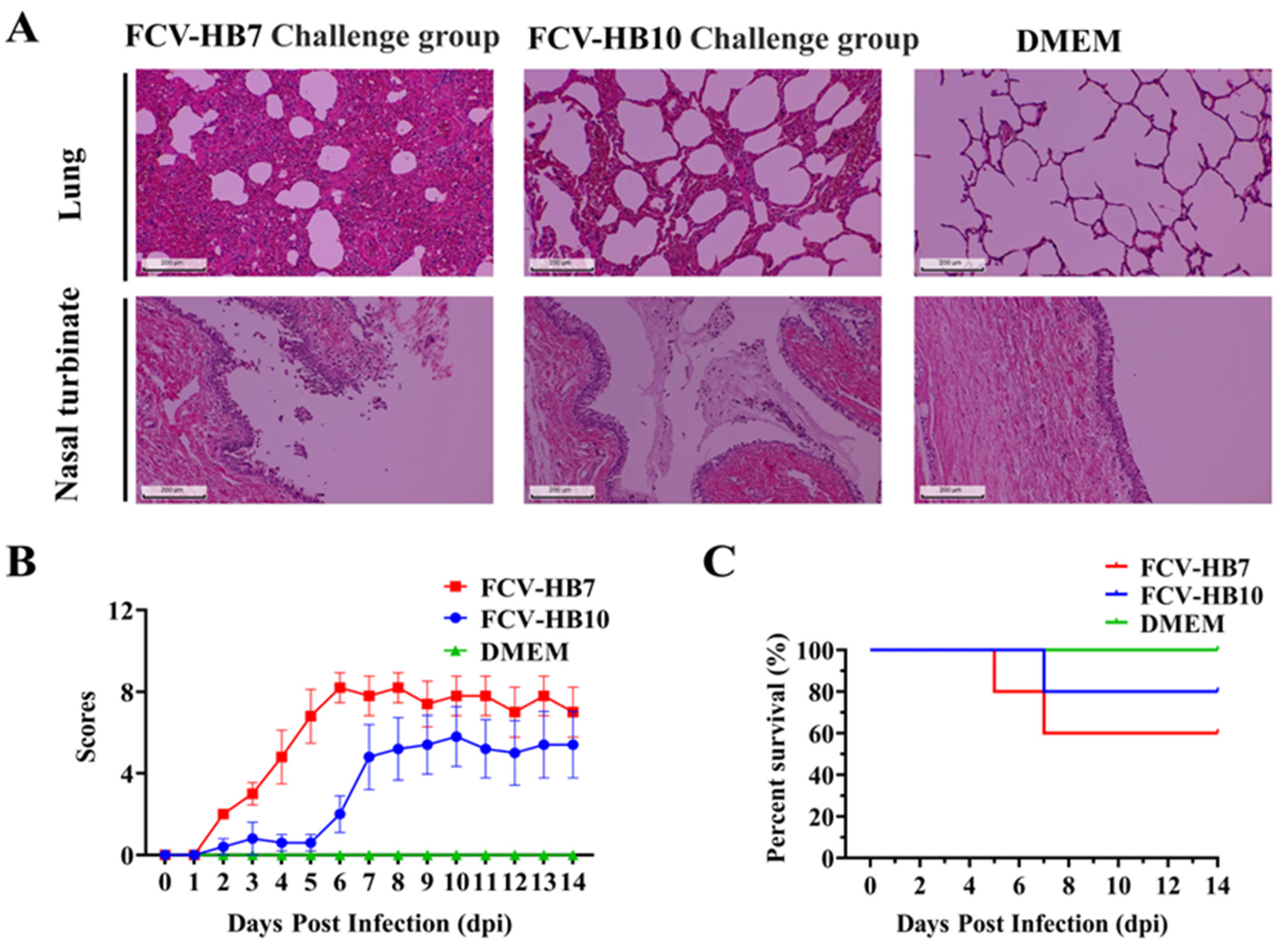
3.5. Pathogenicity of FCV-HB7 and FCV-HB10 in Cats
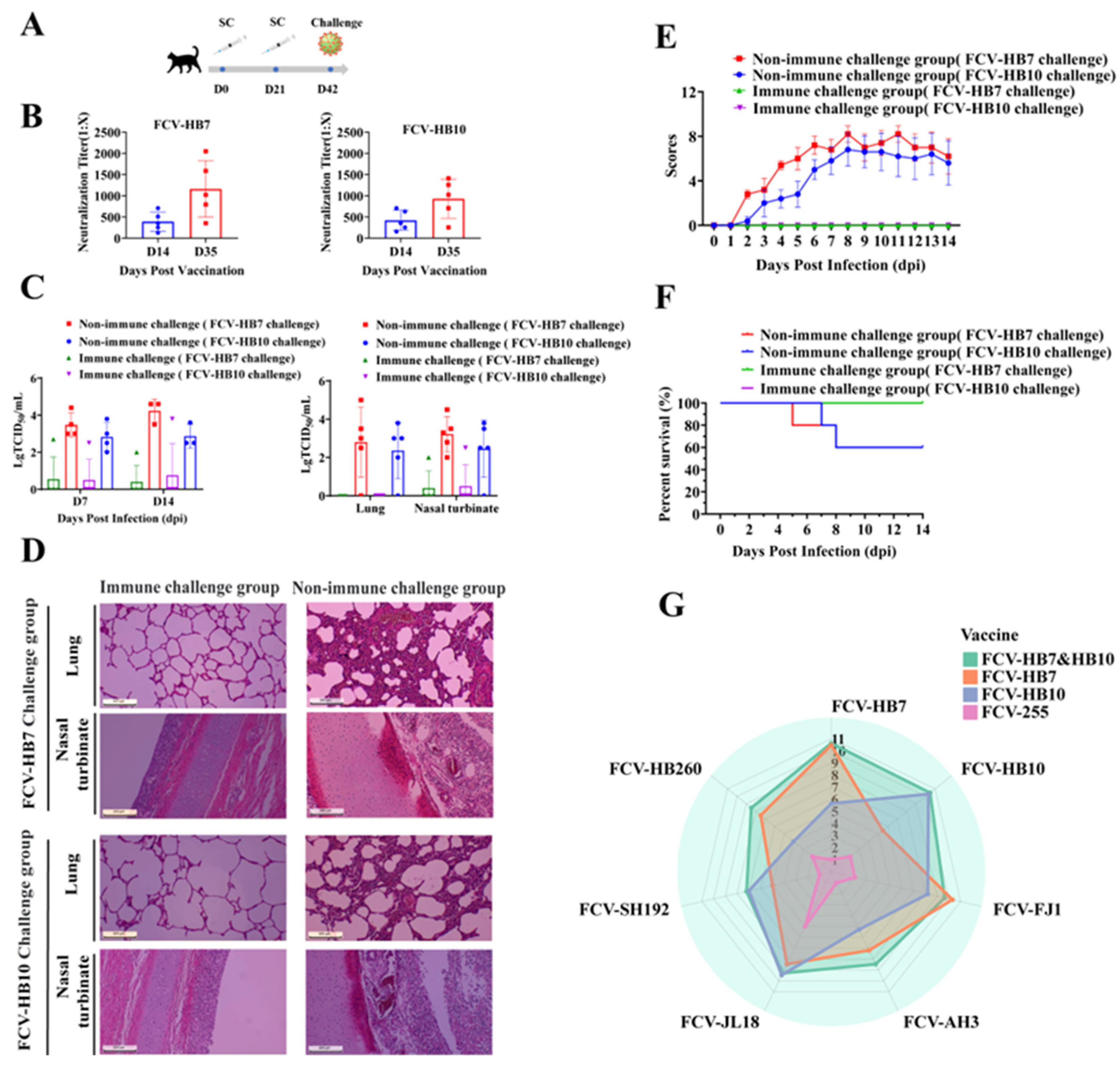
3.6. Cat Protective Efficacy of FCV Vaccine Strain and In Vitro Cross-Neutralization Test
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Ormerod, E.; Jarrett, O. A classification of feline calicivirus isolates based on plaque morphology. J. Gen. Virol. 1978, 39, 537–540. [Google Scholar] [CrossRef] [PubMed]
- Lauritzen, A.; Jarrett, O.; Sabara, M. Serological analysis of feline calicivirus isolates from the United States and United Kingdom. Vet. Microbiol. 1997, 56, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, N.C. Feline calicivirus infection. Tijdschr. Diergeneeskd. 2005, 130, 275–277. [Google Scholar]
- Ohe, K.; Sakai, S.; Sunaga, F.; Murakami, M.; Kiuchi, A.; Fukuyama, M.; Furuhata, K.; Hara, M.; Soma, T.; Ishikawa, Y.; et al. Detection of feline calicivirus (FCV) from vaccinated cats and phylogenetic analysis of its capsid genes. Vet. Res. Commun. 2006, 30, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Radford, A.D.; Coyne, K.P.; Dawson, S.; Porter, C.J.; Gaskell, R.M. Feline calicivirus. Vet. Res. 2007, 38, 319–335. [Google Scholar] [CrossRef] [PubMed]
- Pesavento, P.A.; Chang, K.O.; Parker, J.S. Molecular virology of feline calicivirus. Vet. Clin. N. Am. Small Anim. Pract. 2008, 38, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Ossiboff, R.J.; Zhou, Y.; Lightfoot, P.J.; Prasad, B.V.; Parker, J.S. Conformational changes in the capsid of a calicivirus upon interaction with its functional receptor. J. Virol. 2010, 84, 5550–5564. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Liu, D.; Liu, Y.; Wu, H.; Jiang, Y.; Zu, S.; Liu, C.; Sun, X.; Liu, J.; Qu, L. Molecular characterization of a feline calicivirus isolated from tiger and its pathogenesis in cats. Vet. Microbiol. 2016, 192, 110–117. [Google Scholar] [CrossRef]
- Seal, B.S.; Neill, J.D. Capsid protein gene sequence of feline calicivirus isolates 255 and LLK: Further evidence for capsid protein configuration among feline caliciviruses. Virus Genes 1995, 9, 183–187. [Google Scholar] [CrossRef]
- Geissler, K.; Schneider, K.; Truyen, U. Mapping neutralizing and non-neutralizing epitopes on the capsid protein of feline calicivirus. J. Vet. Med. B Infect. Dis. Vet. Public Health 2002, 49, 55–60. [Google Scholar] [CrossRef]
- Knowles, J.O.; Dawson, S.; Gaskell, R.M.; Gaskell, C.J.; Harvey, C.E. Neutralisation patterns among recent British and North American feline calicivirus isolates from different clinical origins. Vet. Rec. 1990, 127, 125–127. [Google Scholar]
- Michaela, C.; Marion, M.; Ian, G.; David, B. Assembly of a portal-like structure in feline calicivirus following receptor engagement. Access Microbiol. 2019, 1, 245. [Google Scholar]
- Lanave, G.; Buonavoglia, A.; Pellegrini, F.; Di Martino, B.; Di Profio, F.; Diakoudi, G.; Catella, C.; Omar, A.H.; Vasinioti, V.I.; Cardone, R.; et al. An Outbreak of Limping Syndrome Associated with Feline Calicivirus. Animals 2023, 13, 1778. [Google Scholar] [CrossRef] [PubMed]
- Barrera-Vazquez, O.S.; Cancio-Lonches, C.; Hernandez-Gonzalez, O.; Chavez-Munguia, B.; Villegas-Sepulveda, N.; Gutierrez-Escolano, A.L. The feline calicivirus leader of the capsid protein causes survivin and XIAP downregulation and apoptosis. Virology 2019, 527, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Spiri, A.M. An Update on Feline Calicivirus. Schweiz. Arch. Tierh. 2022, 164, 225–241. [Google Scholar] [CrossRef] [PubMed]
- Huimin, G.; Qiuhong, M.; Jie, Z.; Zhibiao, Y.; Guangqing, L. Isolation and molecular characterization of a virulent systemic feline calicivirus isolated in China. Infect. Genet. Evol. 2018, 65, 425–429. [Google Scholar]
- Francesca, C.; Gabriella, E.; Nicola, D.; Vito, M.; Gianvito, L.; Katia, V.; Cristiana, C.; Georgia, D.; Grazia, C.; Maria, L.C.; et al. Feline calicivirus infection in cats with virulent systemic disease, Italy. Res. Vet. Sci. 2019, 124, 46–51. [Google Scholar]
- Di Martino, B.; Lanave, G.; Di Profio, F.; Melegari, I.; Marsilio, F.; Camero, M.; Catella, C.; Capozza, P.; Bányai, K.; Barrs, V.R.; et al. Identification of feline calicivirus in cats with enteritis. Transbound. Emerg. Dis. 2020, 67, 2579–2588. [Google Scholar] [CrossRef]
- Spiri, A.M.; Novacco, M.; Meli, M.L.; Stirn, M.; Riond, B.; Fogle, J.E.; Boretti, F.S.; Herbert, I.; Hosie, M.J.; HofmannLehmann, R. Modified-Live Feline Calicivirus Vaccination Elicits Cellular Immunity against a Current Feline Calicivirus Field Strain in an Experimental Feline Challenge Study. Viruses 2021, 13, 1736. [Google Scholar] [CrossRef]
- Paola, D.A.; Chiara, L.; Elisabetta, S.; Eva, S.; Sonia, M.; Stefania, L. Prevalence of serum antibody titres against feline panleukopenia, herpesvirus and calicivirus infections in stray cats of Milan, Italy. Prev. Vet. Med. 2019, 167, 32–38. [Google Scholar]
- Andrea, M.S.; Marina, L.M.; Barbara, R.; Imogen, H.; Margaret, J.H.; Regina, H. Environmental Contamination and Hygienic Measures After Feline Calicivirus Field Strain Infections of Cats in a Research Facility. Viruses 2019, 11, 958. [Google Scholar]
- Zhou, L.; Fu, N.; Ding, L.; Li, Y.; Huang, J.; Sha, X.; Zhou, Q.; Song, X.; Zhang, B. Molecular Characterization and Cross-Reactivity of Feline Calicivirus Circulating in Southwestern China. Viruses 2021, 13, 1812. [Google Scholar] [CrossRef]
- Urban, C.; Luttermann, C. Major Capsid Protein Synthesis from the Genomic RNA of Feline Calicivirus. J. Virol. 2020, 94, 10–1128. [Google Scholar] [CrossRef]
- Smith, S.L.; Afonso, M.M.; Pinchbeck, G.L.; Gaskell, R.M.; Dawson, S.; Radford, A.D. Temporally separated feline calicivirus isolates do not cluster phylogenetically and are similarly neutralised by high-titre vaccine strain FCV-F9 antisera in vitro. J. Feline Med. Surg. 2019, 22, 602–607. [Google Scholar] [CrossRef]
- Bordicchia, M.; Fumian, T.M.; Van Brussel, K.; Russo, A.G.; Carrai, M.; Le, S.J.; Pesavento, P.A.; Holmes, E.C.; Martella, V.; White, P.; et al. Feline Calicivirus Virulent Systemic Disease: Clinical Epidemiology, Analysis of Viral Isolates and In Vitro Efficacy of Novel Antivirals in Australian Outbreaks. Viruses 2021, 13, 2040. [Google Scholar] [CrossRef]
- Zheng, M.; Li, Z.; Fu, X.; Lv, Q.; Yang, Y.; Shi, F. Prevalence of feline calicivirus and the distribution of serum neutralizing antibody against isolate strains in cats of Hangzhou, China. J. Vet. Sci. 2021, 22, e73. [Google Scholar] [CrossRef]
- Gao, J.; Li, Y.; Xie, Q.; Al-Zaban, M.I.; Al-Saeed, F.A.; Shati, A.A.; Al-Doaiss, A.A.; Ahmed, A.E.; Nawaz, S.; Ebrahem, H.; et al. Epidemiological Investigation of Feline Upper Respiratory Tract Infection Encourages a Geographically Specific FCV Vaccine. Vet. Sci. 2023, 10, 46. [Google Scholar] [CrossRef]
- Poulet, H.; Jas, D.; Lemeter, C.; Coupier, C.; Brunet, S. Efficacy of a bivalent inactivated non-adjuvanted feline calicivirus vaccine: Relation between in vitro cross-neutralization and heterologous protection in vivo. Vaccine 2008, 26, 3647–3654. [Google Scholar] [CrossRef]
- Cao, L.; Li, Q.; Shi, K.; Wei, L.; Ouyang, H.; Ye, Z.; Du, W.; Ye, J.; Hui, X.; Li, J.; et al. Isolation and phylogenetic analysis of feline calicivirus strains from various region of China. Anim. Dis. 2022, 2, 16. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, Z.; Chen, M.; Feng, K.; Qi, R.; Zheng, Y.; Wang, Y.; Kang, H.; Jiang, Q.; Yang, M.; et al. Classification of genotypes based on the VP1 gene of feline calicivirus and study of cross-protection between different genotypes. Front. Microbiol. 2023, 14, 1226877. [Google Scholar] [CrossRef]
- Guo, J.; Ding, Y.; Sun, F.; Zhou, H.; He, P.; Chen, J.; Guo, J.; Zeng, H.; Long, J.; Wei, Z.; et al. Co-circulation and evolution of genogroups I and II of respiratory and enteric feline calicivirus isolates in cats. Transbound. Emerg. Dis. 2022, 69, 2924–2937. [Google Scholar] [CrossRef]
- Coyne, K.P.; Gaskell, R.M.; Dawson, S.; Porter, C.J.; Radford, A.D. Evolutionary mechanisms of persistence and diversification of a calicivirus within endemically infected natural host populations. J. Virol. 2007, 81, 1961–1971. [Google Scholar] [CrossRef]
- CubillosZapata, C.; Angulo, I.; Almanza, H.; Borrego, B.; ZamoraCeballos, M.; Castón, J.R.; Mena, I.; Blanco, E.; Bárcena, J. Precise location of linear epitopes on the capsid surface of feline calicivirus recognized by neutralizing and non-neutralizing monoclonal antibodies. Vet. Res. 2020, 51, 59. [Google Scholar] [CrossRef]
- Wang, Z.; Xin, T.; Wei, J.; Jiang, Y.; Liu, X.; Song, W.; Guo, X.; Yuan, W.; Cui, Y.; Zhu, H.; et al. Isolation and phylogenetic analysis of strains of feline calicivirus in Beijing, China. Arch. Virol. 2021, 166, 2521–2527. [Google Scholar] [CrossRef]
- Liu, C.; Liu, Y.; Qian, P.; Cao, Y.; Wang, J.; Sun, C.; Huang, B.; Cui, N.; Huo, N.; Wu, H.; et al. Molecular and serological investigation of cat viral infectious diseases in China from 2016 to 2019. Transbound. Emerg. Dis. 2020, 67, 2329–2335. [Google Scholar] [CrossRef]
- Liang, J.; Zang, M.; Zhou, Z. Genetic and phylogenetic analysis of capsid gene of feline calicivirus in Nanjing, China. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2022, 103, 105323. [Google Scholar] [CrossRef]
- Mao, J.; Ye, S.; Li, Q.; Bai, Y.; Wu, J.; Xu, L.; Wang, Z.; Wang, J.; Zhou, P.; Li, S. Molecular Characterization and Phylogenetic Analysis of Feline Calicivirus Isolated in Guangdong Province, China from 2018 to 2022. Viruses 2022, 14, 2421. [Google Scholar] [CrossRef]
- Brunet, S.; Sigoillot-Claude, C.; Pialot, D.; Poulet, H. Multiple Correspondence Analysis on Amino Acid Properties within the Variable Region of the Capsid Protein Shows Differences between Classical and Virulent Systemic Feline Calicivirus Strains. Viruses 2019, 11, 1090. [Google Scholar] [CrossRef]
- Knowles, J.O.; McArdle, F.; Dawson, S.; Carter, S.D.; Gaskell, C.J.; Gaskell, R.M. Studies on the role of feline calicivirus in chronic stomatitis in cats. Vet. Microbiol. 1991, 27, 205–219. [Google Scholar] [CrossRef]
- Kahn, D.E.; Hoover, E.A.; Bittle, J.L. Induction of immunity to feline caliciviral disease. Infect. Immun. 1975, 11, 1003–1009. [Google Scholar] [CrossRef] [PubMed]
- Michèle, B.; Stephanie, S.; Anna, R.; Uwe, T.; Katrin, H. Antibody Response to Feline Calicivirus Vaccination in Healthy Adult Cats. Viruses 2019, 11, 702. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequences (5′−3′) | Tm (°C) | Product (bp) |
|---|---|---|---|
| FCV-Q1 | F-GTAAAAGAAATTTGAGACAATGT | 51 | 1–2468 |
| R-GAATTAACRGTTACCACATG | |||
| FCV-Q2 | F-GAACTACCCGCCAATCAACATGT | 63 | 2431–5362 |
| R-AGCACGTTAGCGCAGGTTGAGC | |||
| FCV-Q3 | F-TTGAGCATGTGCTCAACCTG | 55 | 5330–7684 |
| R-CCCTGGGGTTAGRCGC |
| Clinical Symptoms | Description of Clinical Symptoms | Score |
|---|---|---|
| Body temperature | 37.5–39.5 | 0 |
| ≥39.6 | 1 | |
| ≤37.4 | 2 | |
| Oral ulcers | None | 0 |
| Small and few | 1 | |
| Big and many | 3 | |
| Eye and nose secreta | None | 0 |
| Few | 1 | |
| Many | 2 | |
| Body weight | Increase or decrease <3% | 0 |
| Reduce ≥3% | 2 | |
| State of existence | Survival | 0 |
| Death | 10 |
| Isolated | Time | Area | Group | Length (bp) | GenBank No. |
|---|---|---|---|---|---|
| FCV-HB7 | 2020 | Hubei, China | C | 7683 | OR645484 |
| FCV-HB10 | 2020 | Hubei, China | D | 7706 | OR645485 |
| FCV-FJ1 | 2021 | Fujian, China | C | 7683 | OR645479 |
| FCV-AH3 | 2021 | Anhui, China | D | 7706 | OR645480 |
| FCV-JL18 | 2021 | Jilin, China | C | 7683 | OR645481 |
| FCV-SH192 | 2021 | Shanghai, China | C | 7666 | OR645482 |
| FCV-HB260 | 2021 | Hubei, China | D | 7704 | OR645483 |
| Strains | ORF1 | ORF2 | ORF3 | |||
|---|---|---|---|---|---|---|
| nt | aa | nt | aa | nt | aa | |
| FCV-HB7 | 76.02 | 87.65 | 74.54 | 83.53 | 78.19 | 85.85 |
| FCV-HB10 | 78.61 | 91.16 | 77.11 | 88.04 | 82.87 | 94.34 |
| FCV-FJ1 | 76.26 | 87.14 | 74.54 | 84.58 | 77.26 | 83.96 |
| FCV-AH3 | 78.80 | 91.10 | 77.21 | 87.00 | 85.36 | 94.34 |
| FCV-JL18 | 75.54 | 86.97 | 74.99 | 85.03 | 78.19 | 84.91 |
| FCV-SH192 | 76.69 | 87.25 | 75.04 | 86.92 | 78.82 | 83.18 |
| FCV-HB260 | 78.99 | 91.10 | 78.34 | 87.28 | 83.54 | 95.28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, L.; Liu, J.; Li, Y.; Xie, D.; Yan, Q.; Li, Q.; Cao, Y.; Du, W.; Li, J.; Ye, Z.; et al. Biological Characteristics of Feline Calicivirus Epidemic Strains in China and Screening of Broad-Spectrum Protective Vaccine Strains. Vaccines 2023, 11, 1858. https://doi.org/10.3390/vaccines11121858
Cao L, Liu J, Li Y, Xie D, Yan Q, Li Q, Cao Y, Du W, Li J, Ye Z, et al. Biological Characteristics of Feline Calicivirus Epidemic Strains in China and Screening of Broad-Spectrum Protective Vaccine Strains. Vaccines. 2023; 11(12):1858. https://doi.org/10.3390/vaccines11121858
Chicago/Turabian StyleCao, Longlong, Jian Liu, Yongfan Li, Denglong Xie, Quanhui Yan, Qiuyan Li, Yiran Cao, Wenxin Du, Jiakang Li, Zijun Ye, and et al. 2023. "Biological Characteristics of Feline Calicivirus Epidemic Strains in China and Screening of Broad-Spectrum Protective Vaccine Strains" Vaccines 11, no. 12: 1858. https://doi.org/10.3390/vaccines11121858
APA StyleCao, L., Liu, J., Li, Y., Xie, D., Yan, Q., Li, Q., Cao, Y., Du, W., Li, J., Ye, Z., Zhou, D., Kang, C., & Cao, S. (2023). Biological Characteristics of Feline Calicivirus Epidemic Strains in China and Screening of Broad-Spectrum Protective Vaccine Strains. Vaccines, 11(12), 1858. https://doi.org/10.3390/vaccines11121858