
Autoinflammation in Syndromic Hidradenitis Suppurativa: The Role of AIM2

 ,
,  ,
,  ,
,  , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Gene Sequencing
3. Results
3.1. Clinical Features
3.2. Genetic Analyses
4. Discussion
5. Limitations and Strengths
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Amarante-Mendes, G.P.; Adjemian, S.; Branco, L.M.; Zanetti, L.C.; Weinlich, R.; Bortoluci, K.R. Pattern Recognition Receptors and the Host Cell Death Molecular Machinery. Front. Immunol. 2018, 9, 2379. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed]
- Strowig, T.; Henao-Mejia, J.; Elinav, E.; Flavell, R. Inflammasomes in health and disease. Nature 2012, 481, 278–286. [Google Scholar] [CrossRef]
- Hornung, V.; Ablasser, A.; Charrel-Dennis, M.; Bauernfeind, F.; Horvath, G.; Caffrey, D.R.; Latz, E.; Fitzgerald, K.A. AIM2 recognizes cytosolic dsDNA and forms a caspase-1-activating inflammasome with ASC. Nature 2009, 458, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, M.; Choubey, D.; Lengauer, T. The HIN domain of IFI-200 proteins consists of two OB folds. Biochem. Biophys. Res. Commun. 2005, 327, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Dombrowski, Y.; Peric, M.; Koglin, S.; Kammerbauer, C.; Göss, C.; Anz, D.; Simanski, M.; Gläser, R.; Harder, J.; Hornung, V.; et al. Cytosolic DNA triggers inflammasome activation in keratinocytes in psoriatic lesions. Sci. Transl. Med. 2011, 3, 82ra38. [Google Scholar] [CrossRef]
- Javierre, B.M.; Fernandez, A.F.; Richter, J.; Al-Shahrour, F.; Martin-Subero, J.I.; Rodriguez-Ubreva, J.; Berdasco, M.; Fraga, M.F.; O’Hanlon, T.P.; Rider, L.G.; et al. Changes in the pattern of DNA methylation associate with twin discordance in systemic lupus erythematosus. Genome Res. 2010, 20, 170–179. [Google Scholar] [CrossRef]
- de Koning, H.D.; Bergboer, J.G.; van den Bogaard, E.H.; van Vlijmen-Willems, I.M.; Rodijk-Olthuis, D.; Simon, A.; Zeeuwen, P.L.; Schalkwijk, J. Strong induction of AIM2 expression in human epidermis in acute and chronic inflammatory skin conditions. Exp. Dermatol. 2012, 21, 961–964. [Google Scholar] [CrossRef]
- Jemec, G.B. Clinical practice. Hidradenitis suppurativa. N. Engl. J. Med. 2012, 366, 158–164. [Google Scholar] [CrossRef]
- Gasparic, J.; Theut Riis, P.; Jemec, G.B. Recognizing syndromic hidradenitis suppurativa: A review of the literature. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 1809–1816. [Google Scholar] [CrossRef]
- Marzano, A.V.; Damiani, G.; Ceccherini, I.; Berti, E.; Gattorno, M.; Cugno, M. Autoinflammation in pyoderma gangrenosum and its syndromic form (pyoderma gangrenosum, acne and suppurative hidradenitis). Br. J. Dermatol. 2017, 176, 1588–1598. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Xu, H.; Wang, B. Is SAPHO Syndrome Linked to PASH Syndrome and Hidradenitis Suppurativa by Nicastrin Mutation? A Case Report. J. Rheumatol. 2018, 45, 1605–1607. [Google Scholar] [CrossRef]
- Brandao, L.; Moura, R.; Tricarico, P.M.; Gratton, R.; Genovese, G.; Moltrasio, C.; Garcovich, S.; Boniotto, M.; Crovella, S.; Marzano, A.V. Altered keratinization and vitamin D metabolism may be key pathogenetic pathways in syndromic hidradenitis suppurativa: A novel whole exome sequencing approach. J. Dermatol. Sci. 2020, 99, 17–22. [Google Scholar] [CrossRef]
- Marzano, A.V.; Genovese, G.; Moltrasio, C.; Tricarico, P.M.; Gratton, R.; Piaserico, S.; Garcovich, S.; Boniotto, M.; Brandão, L.; Moura, R.; et al. Whole-Exome Sequencing in 10 Unrelated Patients with Syndromic Hidradenitis Suppurativa: A Preliminary Step for a Genotype-Phenotype Correlation. Dermatology 2022, 238, 860–869. [Google Scholar] [CrossRef]
- Moltrasio, C.; Tricarico, P.M.; Romagnuolo, M.; Marzano, A.V.; Crovella, S. Hidradenitis Suppurativa: A Perspective on Genetic Factors Involved in the Disease. Biomedicines 2022, 10, 2039. [Google Scholar] [CrossRef]
- Sabat, R.; Jemec, G.B.E.; Matusiak, Ł.; Kimball, A.B.; Prens, E.; Wolk, K. Hidradenitis suppurativa. Nat. Rev. Dis. Primers. 2020, 6, 18. [Google Scholar] [CrossRef]
- Ensemble. Available online: https://www.ensembl.org/ (accessed on 10 October 2022).
- GnomAD. Available online: https://gnomad.broadinstitute.org (accessed on 10 October 2022).
- dbSNP. Available online: http://www.ncbi.nlm.nih.gov/SNP (accessed on 10 October 2022).
- Lugrin, J.; Martinon, F. The AIM2 inflammasome: Sensor of pathogens and cellular perturbations. Immunol. Rev. 2018, 281, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Marzano, A.V.; Borghi, A.; Wallach, D.; Cugno, M. A Comprehensive Review of Neutrophilic Diseases. Clin. Rev. Allergy Immunol. 2018, 54, 114–130. [Google Scholar] [CrossRef] [PubMed]
- Marzano, A.V.; Ceccherini, I.; Gattorno, M.; Fanoni, D.; Caroli, F.; Rusmini, M.; Grossi, A.; De Simone, C.; Borghi, O.M.; Meroni, P.L.; et al. Association of pyoderma gangrenosum, acne, and suppurative hidradenitis (PASH) shares genetic and cytokine profiles with other autoinflammatory diseases. Medicine 2014, 93, e187. [Google Scholar] [CrossRef]
- Hurtado-Nedelec, M.; Chollet-Martin, S.; Chapeton, D.; Hugot, J.P.; Hayem, G.; Gérard, B. Genetic susceptibility factors in a cohort of 38 patients with SAPHO syndrome: A study of PSTPIP2, NOD2, and LPIN2 genes. J. Rheumatol. 2010, 37, 401–409. [Google Scholar] [CrossRef]
- DeFilippis, E.M.; Feldman, S.R.; Huang, W.W. The genetics of pyoderma gangrenosum and implications for treatment: A systematic review. Br. J. Dermatol. 2015, 172, 1487–1497. [Google Scholar] [CrossRef] [PubMed]
- Perko, D.; Debeljak, M.; Toplak, N.; Avčin, T. Clinical features and genetic background of the periodic Fever syndrome with aphthous stomatitis, pharyngitis, and adenitis: A single center longitudinal study of 81 patients. Mediat. Inflamm. 2015, 2015, 293417. [Google Scholar] [CrossRef] [PubMed]


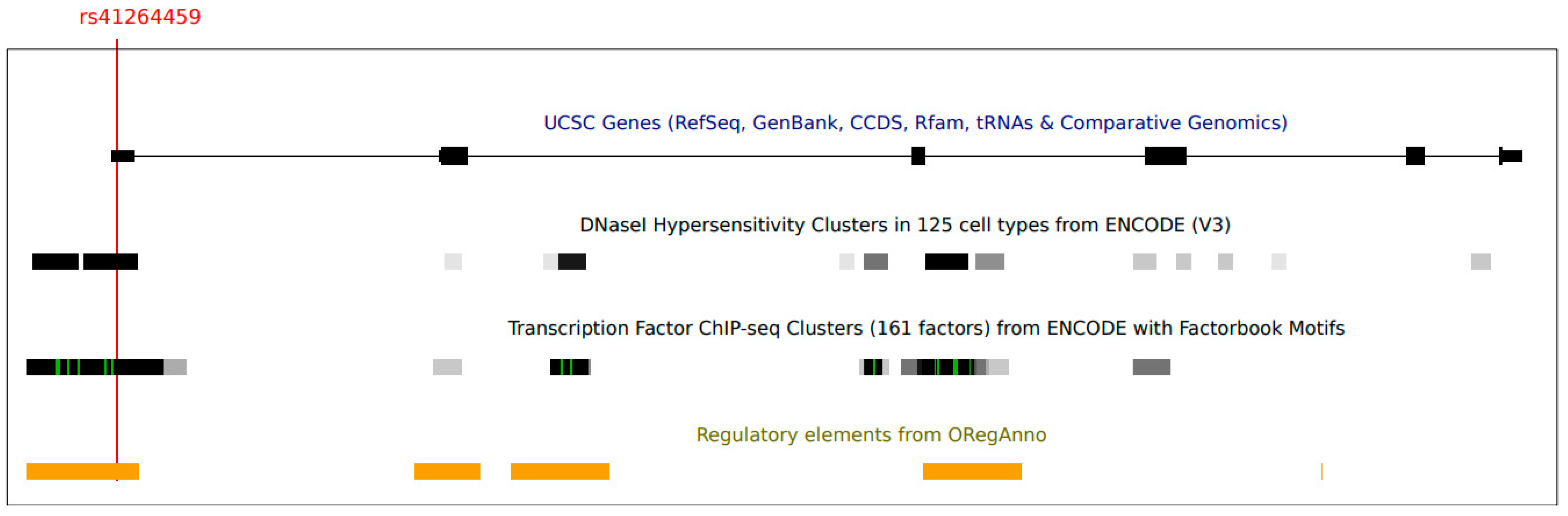{kind=link}
| ID | Sex | Diagnosis | Comorbidities | Hidradenitis suppurativa | Pyoderma gangrenosum | Acne | Treatments | Follow-up | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Age at onset (years) | Involved sites | IHS4 | Age at onset (years) | Involved sites | PG Score | Age at onset (years) | Involved sites | ||||||
| 1 | M | PAPASH | pyogenic arthritis | 14 | axillae, groins, back, nuchal region, scalp | 30 | 18 | trunk | multi-lesional | 13 | face, back | rifampicin, doxycycline, azitromycine, lymecycline, clindamycin, adalimumab, anakinra, prednisone | CR |
| 2 | M | PASH/SAPHO | SAPHO | 43 | axillae, groins, anogenital area | 27 | 43 | lower limbs, trunk, upper limbs | disseminated | 17 | face, back | clindamycin, infliximab | CR |
| 3 | M | PASH/SAPHO | SAPHO | 42 | axillae, groins, anogenital area | 19 | 46 | lower limbs | localized | 16 | face | clindamycin, methotrexate, infliximab, adalimumab | CR |
| 4 | M | PASH | Obesity, BBS, osteoporosis, PPPP | 36 | axillae, groins, anogenital area | 24 | 36 | lower limbs, trunk, upper limbs | disseminated | 34 | face | trimetoprim-sulfametoxazole, azitromycine, cyclosporine, prednisone, adalimumab, ustekinumab, | CR |
| 5 | F | PASH | Obesity, BBS, myocardial infarction, type II DM, BP | 51 | axillae, groins, submammary folds, anogenital area | 23 | 51 | perianal region | localized | 15 | face | rifampicin, doxycycline, infliximab, adalimumab | CR |
| 6 | F | PASH | PCOS | 15 | Axillae, groin | 20 | 24 | Lower limbs | multi-lesional | 14 | Face, back | infliximab | CR |
| 7 | F | PASH/SAPHO | SAPHO | 15 | axillae, groins, anogenital region, nuchal region | 26 | 15 | trunk | multi-lesional | 13 | face | rifampicin, doxycycline, azitromycine, lymecycline, clindamycin, intravenous immunoglobulin, methotrexate, isotretinoin, adalimumab, anakinra, ustekinumab, secukinumab | NR |
| 8 | F | PAPASH | Ulcerative colitis, PCOS, pyogenic arthritis | 28 | submammary folds, groins, anogenital area | 22 | 36 | peristomal | localized | 15 | face, back | doxycycline, infliximab, surgery for HS | CR |
| 9 | M | PASH | Crohn disease, osteoporosis | 18 | axillae, groins, anogenital area, nuchal region | 23 | 19 | lower limbs | multi-lesional | 16 | face, back | azitromycine, trimetoprim-sulfametoxazole, ceftriaxone, ciprofloxacine, lymecycline, rifampicin, amoxicillin clavulanate, methylprednisolone, dapsone, cyclosporine, adalimumab, anakinra, | PR |
| 10 | F | PASH | Crohn disease, coeliac disease, myasthenia gravis, PCOS | 18 | axillae, nuchal region, submammary folds, groins, anogenital area | 19 | 44 | peristomal | localized | 17 | face, back | clindamycin, methotrexate, infliximab, adalimumab, surgery for HS | CR |
| 11 | F | PAPASH | FMF, PCOS, pyogenic arthritis | 16 | axillae, groins, anogenital area, intermammary and submammary folds | 20 | 16 | trunk | multi-lesional | 14 | face | azitromycine, doxycycline, dapsone, adalimumab, anakinra, colchicine | CR |
| 12 | M | PASH | Pilonidal cyst | 15 | axillae and anogenital | 21 | 18 | lower back | localized | 14 | Face, back | Azithromycin, infliximab, adalimumab | PR/CR |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moltrasio, C.; Cagliani, R.; Sironi, M.; Clerici, M.; Pontremoli, C.; Maronese, C.A.; Tricarico, P.M.; Crovella, S.; Marzano, A.V. Autoinflammation in Syndromic Hidradenitis Suppurativa: The Role of AIM2. Vaccines 2023, 11, 162. https://doi.org/10.3390/vaccines11010162
Moltrasio C, Cagliani R, Sironi M, Clerici M, Pontremoli C, Maronese CA, Tricarico PM, Crovella S, Marzano AV. Autoinflammation in Syndromic Hidradenitis Suppurativa: The Role of AIM2. Vaccines. 2023; 11(1):162. https://doi.org/10.3390/vaccines11010162
Chicago/Turabian StyleMoltrasio, Chiara, Rachele Cagliani, Manuela Sironi, Mario Clerici, Chiara Pontremoli, Carlo Alberto Maronese, Paola Maura Tricarico, Sergio Crovella, and Angelo Valerio Marzano. 2023. "Autoinflammation in Syndromic Hidradenitis Suppurativa: The Role of AIM2" Vaccines 11, no. 1: 162. https://doi.org/10.3390/vaccines11010162
APA StyleMoltrasio, C., Cagliani, R., Sironi, M., Clerici, M., Pontremoli, C., Maronese, C. A., Tricarico, P. M., Crovella, S., & Marzano, A. V. (2023). Autoinflammation in Syndromic Hidradenitis Suppurativa: The Role of AIM2. Vaccines, 11(1), 162. https://doi.org/10.3390/vaccines11010162