Antioxidant Activity Modulated by Polyphenol Contents in Apple and Leaves during Fruit Development and Ripening



Abstract

1. Introduction
2. Materials and Methods
2.1. Plant Material and Sample Preparation
2.2. Extraction Procedure
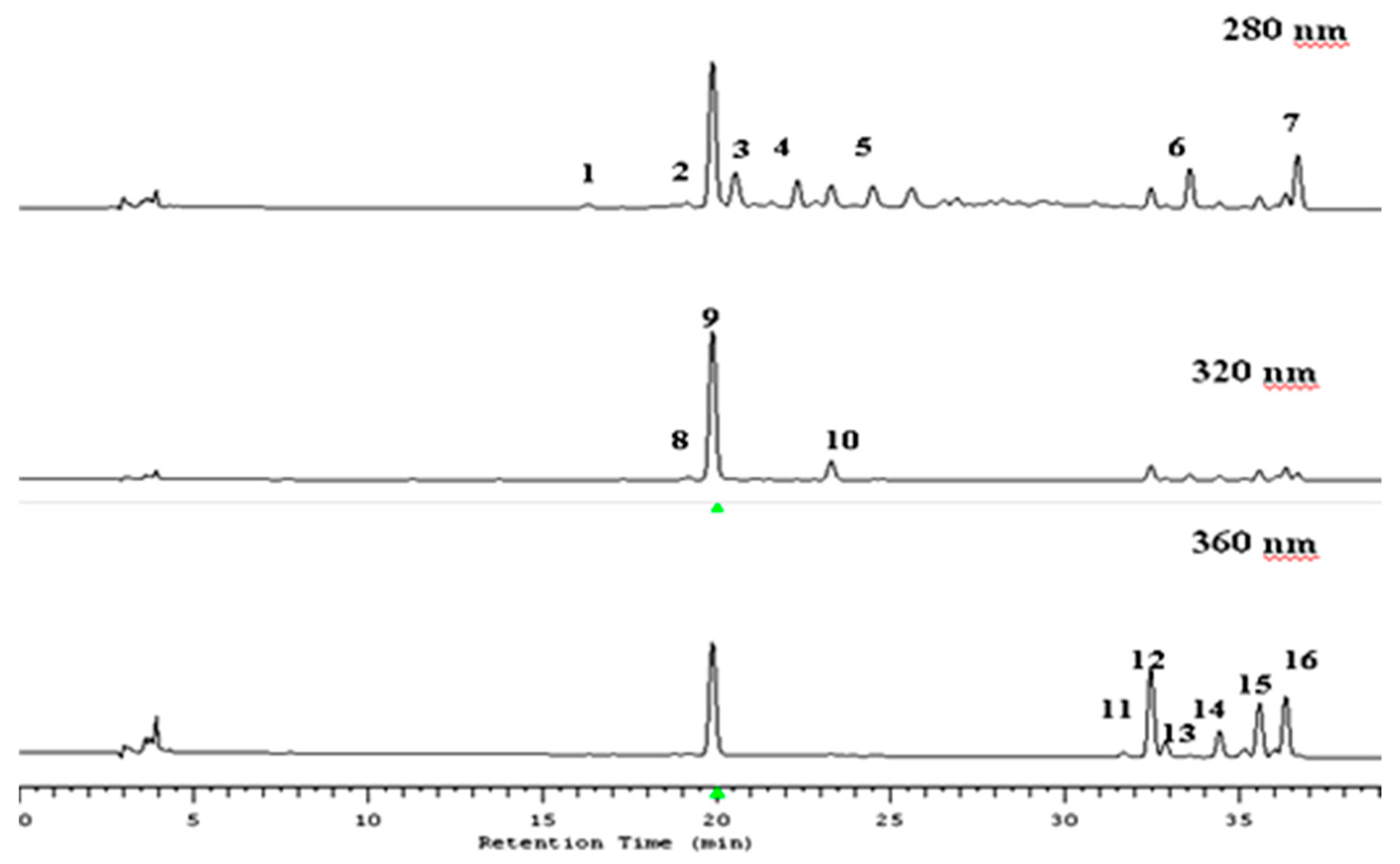
2.3. Identification of Polyphenols by LC–MS Method
2.4. Quantification of Polyphenols by HPLC Method
2.5. Polymeric Procyanidins Analysis by Thiolysis Method
2.6. Antioxidant Activities Measurements as 2,2-azinobis-(3-ethyl-benzothiazoline-6-sulfonic acid (ABTS) and 2,2-diphenyl-picrylhydrazyl (DPPH) Assay
2.7. Statistical Analysis
3. Results and Discussion
3.1. Qualitative Analysis of Polyphenol Compounds
3.2. Quantification Phenolic Compounds in Apple Fruits and Leaves
3.3. Antioxidant Capacity Fruits and Leaves of Apple cv. Apple
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wildman, R.E. Nutraceuticals: A brief review of historical and teleological aspects. In Handbook of Nutraceuticals and Functional Foods; CRC Press: Boca Raton, FL, USA, 2001; pp. 23–34. [Google Scholar]
- Guthrie, N.; Kurowska, E.M.N. Anticancer and cholesterol lowering activities of citrus flavonoids. In Handbook of Nutraceuticals and Functional Foods; CRC Press: Boca Raton, FL, USA, 2001; pp. 113–126. [Google Scholar]
- Lee, K.W.; Kim, Y.J.; Kim, D.O.; Lee, H.J.; Lee, C.Y. Major phenolics in apple and their contribution to the total antioxidant capacity. J. Agric. Food Chem. 2003, 51, 6516–6520. [Google Scholar] [CrossRef] [PubMed]
- Knekt, P.; Järvinen, R.; Seppänen, R.; Heliövaara, M.; Tempo, L.; Pukkala, E.; Aromat, A. Dietary flavonoids and the risk of lung cancer and other malignant neoplasms. Am. J. Epidemiol. 1997, 146, 223–230. [Google Scholar] [CrossRef]
- Knekt, P.; Kumpulainen, J.; Jarvinen, R.; Rissanen, H.; Heliovaara, M.; Reunanen, A.; Hakulinen, T.; Aromaa, A. Flavonoid intake and risk of chronic diseases. Am. J. Clin. Nutr. 2002, 76, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Boyer, J.; Liu, R.H. Apple phytochemicals and their health benefits. Nutr. J. 2004, 3, 5. [Google Scholar] [CrossRef] [PubMed]
- Hyson, D.; Studebaker-Hallman, D.; Davis, P.A.; Gershwin, M.E. Apple juice consumption reduces plasma low-density lipoprotein oxidation in healthy men and women. J. Med. Food 2000, 3, 159–166. [Google Scholar] [CrossRef]
- Shoji, T.; Mutsuga, M.; Nakamura, T.; Kanda, T.; Akiyama, H.; Goda, Y. Isolation and structural elucidation of some procyanidins from apple by low-temperature nuclear magnetic resonance. J. Agric. Food Chem. 2003, 51, 3806–3813. [Google Scholar] [CrossRef] [PubMed]
- Sanoner, P.; Guyot, S.; Marnet, N.; Molle, D.; Drilleau, J.F. Polyphenolic profiles of French cider apple varieties. J. Agric. Food Chem. 1999, 47, 4847–4853. [Google Scholar] [CrossRef]
- Guyot, S.; Le Bourvellec, C.; Marnet, N.; Drilleau, J.F. Procyanidins are the most abundant polyphenols in dessert apples at maturity. LWT-Food Sci. Technol. 2002, 35, 289–291. [Google Scholar] [CrossRef]
- Guyot, S.; Marnet, N.; Sanoner, P.; Drilleau, J.F. Variability of the polyphenolic composition of cider apple (Malus domestica) fruits and juices. J. Agric. Food Chem. 2003, 51, 6240–6247. [Google Scholar] [CrossRef]
- Lea, A.G.; Arnold, G.M. The phenolics of ciders: Bitterness and astringency. J. Sci. Food Agric. 1978, 29, 478–483. [Google Scholar] [CrossRef]
- Vidal, S.; Francis, L.; Guyot, S.; Marnet, N.; Kwiatkowski, M.; Gawel, R.; Cheynier, V.; Waters, E.J. The mouth-feel properties of grape and apple proanthocyanidins in a wine-like medium. J. Sci. Food Agric. 2003, 83, 564–573. [Google Scholar] [CrossRef]
- Awad, M.A.; Jager, A.; van Westing, L.M. Flavonoid and chlorogenic acid levels in apple fruit: Characterization of variation. Sci. Hortic. 2000, 83, 249–263. [Google Scholar] [CrossRef]
- Tsao, R.; Yang, R.; Young, J.C.; Zhu, H. Polyphenolic profiles in eight apple cultivars using high-performance liquid chromatography (HPLC). J. Agric. Food Chem. 2003, 51, 6347–6353. [Google Scholar] [CrossRef] [PubMed]
- Mosel, H.D.; Herrmann, K. Changes in catechins and hydroxycinnamic acid derivatives during development of apples and pears. J. Sci. Food Agric. 1974, 25, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Renard, C.M.; Dupont, N.; Guillermin, P. Concentrations and characteristics of procyanidins and other phenolics in apples during fruit growth. Phytochemistry 2007, 68, 1128–1138. [Google Scholar] [CrossRef]
- Oliveira, A.P.; Pereira, J.O.; Andrade, P.B.; Valentão, P.; Seabra, R.M.; Silva, B.M. Phenolic profile of Cydonia oblonga Miller leaves. J. Agric. Food Chem. 2007, 55, 7926–7930. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.L.; Juan, I.M.; Chen, Y.L.; Liang, Y.C.; Lin, J.K. Composition of polyphenols in fresh tea leaves and associations of their oxygen-radical-absorbing capacity with antiproliferative actions in fibroblast cells. J. Agric. Food Chem. 1996, 44, 1387–1394. [Google Scholar] [CrossRef]
- Osawa, T. Novel natural antioxidants for utilization in food and biological systems. In Postharvest Biochemistry of Plant Food-Materials in the Tropics; Uritani, I., Garcia, V.V., Mendoza, E.M.T., Eds.; Japan Scientific Society Press: Tokyo, Japan, 1994; pp. 241–251. [Google Scholar]
- Ridgway, T.; Tucker, G.; Wiseman, H. Novel bioconversions for the production of designer antioxidant and colourant flavonoids using polyphenol oxidases. Biotechnol. Genet. Eng. Rev. 1997, 14, 165–190. [Google Scholar] [CrossRef] [PubMed]
- Guyot, S.; Marnet, N.; Sanoner, P.; Drilleau, J.F. Direct thiolysis on crude apple materials for HPLC characterization and quantification of polyphenols in cider apple tissues and juices. Met. Enzymol. 2001, 335, 57–64. [Google Scholar]
- Yen, G.C.; Chen, H.Y. Antioxidant activity of various tea extracts in relation to their antimutagenicity. J. Agric. Food Chem. 1995, 43, 27–32. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Rad Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Mullen, W.; Marks, S.C.; Crozier, A. Evaluation of phenolic compounds in commercial fruit juices and fruit drinks. J. Agric. Food Chem. 2007, 55, 3148–3157. [Google Scholar] [CrossRef] [PubMed]
- Burda, S.; Oleszek, W.; Lee, C.Y. Phenolic compounds and their changes in apples during maturation and cold storage. J. Agric. Food Chem. 1999, 38, 945–948. [Google Scholar] [CrossRef]
- Wojdyło, A.; Oszmiański, J.; Laskowski, P. Polyphenolic compounds and antioxidant activity of new and old apple varieties. J. Agric. Food Chem. 2008, 56, 6520–6530. [Google Scholar] [CrossRef]
- Mayr, U.; Treutter, D.; Santos-Buelga, C.; Bauer, H.; Feucht, W. Developmental changes in the phenol concentrations of ‘‘Golden delicious’’ apple fruits and leaves. Phytochemistry 1995, 38, 1151–1155. [Google Scholar] [CrossRef]
- Takos, A.M.; Ubi, B.E.; Robinson, S.P.; Walkee, A.R. Condensed tannin biosynthesis genes are regulated separately from other flavonoid biosynthesis genes in apple fruit skin. Plant. Sci. 2006, 170, 487–499. [Google Scholar] [CrossRef]
- Shiow, Y.; Wang, S.Y.; Lin, H.S. Antioxidant activity in fruits and leaves of blackberry, raspberry, and strawberry varies with cultivar and developmental stage. J. Agric. Food Chem. 2000, 48, 140–146. [Google Scholar]
- Nuncio-Jáuregui, N.; Nowicka, P.; Munera-Picazo, S.; Hernández, F.; Carbonell-Barrachina, A.A.; Wojdyło, A. Identification and quantification of major derivatives of ellagic acid and antioxidant properties of thinning and ripe Spanish pomegranates. J. Funct. Food 2014, 12, 354–364. [Google Scholar] [CrossRef]
- Melgarejo, P.; Hernández, F.; Legua, P. El Granado. In Proceedings of the I Jornadas Nacionales sobre el Granado: Producción, Economía, Industrialización, Alimentación y Salud; Departamento de Producción Vegetal y Microbiología, Universidad Miguel Hernández de Elche: Elche, Spain, 2010; pp. 36–37. [Google Scholar]
- Fawole, O.A.; Opara, U.L. Changes in physical properties, chemical and elemental composition and antioxidant capacity of pomegranate (cv. Ruby) fruit at five maturity stages. Sci. Hortic. 2013, 150, 37–46. [Google Scholar] [CrossRef]
- Shwartz, E.; Glazer, I.; Bar-Yaakov, I.; Matityahu, I.; Bar-Ilan, I.; Holland, D.; Amir, R. Changes in chemical constituents during the maturation and ripening of two commercially important pomegranate accessions. Food Chem. 2009, 115, 965–973. [Google Scholar] [CrossRef]
- Walle, T.; Walle, U.K. The beta-d-glucoside and sodium-dependent glucose transporter 1 (SGLT1)-inhibitor phloridzin is transported by both SGLT1 and multidrug resistance-associated proteins 1/2. Drug Metab. Dispos. 2003, 31, 1288–1291. [Google Scholar] [CrossRef] [PubMed]
- Ehrenkranz, J.R.; Lewis, N.G.; Kahn, C.R.; Roth, J. Phlorizin: A review. Diabetes Metab. Res. Rev. 2005, 21, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Xing, N.; Chen, Y.; Mitchell, S.H.; Young, C.Y. Quercetin inhibits the expression and function of the androgen receptor in LNCaP prostate cancer cells. Carcinogenesis 2001, 22, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Liaudanskas, M.; Viskelis, P.; Raudonis, R.; Kviklys, D.; Uselis, N.; Janulis, V. Phenolic composition and antioxidant activity of Malus domestica leaves. Sci. World J. 2014. [Google Scholar] [CrossRef] [PubMed]
- Knekt, P.; Isotupa, S.; Rissanen, H.; Heliovaara, M.; Jarvinen, R.; Hakkinen, S.; Aroma, A.; Reunanen, A. Quercetin intake and the incidence of cerebrovascular disease. Eur. J. Clin. Nutr. 2000, 54, 415–417. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Group of Polyphenols | Rt (min) | λmax (nm) | Compound | Molecular Weight | MS (M−H)− | MS/MS Fragments |
|---|---|---|---|---|---|---|
| hydroxycinnamic acid | 19.1 | 305 | p-coumarylquinic acid | 338 | 337 | 191 |
| 19.9 | 320 | 5-O-caffeoylquinic acid | 354 | 353 | 191 | |
| 23.5 | 305 | caffeic acid hexose conjugate | 342 | 341 | 179 | |
| flavanol and procyanidins | 16.8 | 275 | procyanidin B1 | 578 | 577 | 289/245 |
| 19.5 | 280 | (+)-catechin | 290 | 289 | 245 | |
| 20.8 | 275 | procyanidin B2 | 578 | 577 | 289/245 | |
| 22.4 | 275 | (−)-epicatechin | 290 | 289 | 245 | |
| 24.5 | 280 | procyanidin C1 | 866 | 865 | 577/289/245 | |
| dihydrochalcones | 34.0 | 285 | phloretin-2′-O-xyloglucoside | 568 | 567 | 273 |
| 36.8 | 285 | phloretin-2′-O-glucoside | 436 | 435 | 273 | |
| flavonols | 31.6 | 350 | quercetin- 3-O-rutinoside | 610 | 609 | 301 |
| 32.4 | 355 | quercetin-3-O-galactoside | 464 | 463 | 301 | |
| 32.9 | 350 | quercetin-3-O-glucoside | 464 | 463 | 301 | |
| 34.3 | 355 | quercitin-3-O-arabinoside | 434 | 433 | 301 | |
| 35.5 | 350 | quercitin-3-O-xyloside | 434 | 433 | 301 | |
| 36.4 | 345 | quercetin-3-O-rhamnoside | 448 | 447 | 301 |
| Phenolic Compounds | Cultivars | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ozark Gold | Starkinson | Kosztela | ||||||||||
| Fruits after Full Bloom (Days) | ||||||||||||
| 60 | 80 | 130 | 145 | 60 | 80 | 130 | 145 | 60 | 80 | 130 | 145 | |
| CQA | 0.88 ± 0.18 d,* | 0.69 ± 0.03 c,d | 0.37 ± 0.02 d | 0.27 ± 0.08 e | 0.13 ± 0.02 c | 0.09 ± 0.03 c | 0.05 ± 0.01 c | 0.04 ± 0.01 c | 6.33 ± 0.08 b | 4.43 ± 0.06 b | 2.17 ± 0.04 b | 1.62 ± 0.03 b |
| CGLU | 0.17 ± 0.05 k | 0.12 ± 0.02 n,m | 0.04 ± 0.03 l,m | 0.03 ± 0.01 n | 0.02 ± 0.00 e | 0.02 ± 0.01 d | 0.04 ± 0.00 e | 0.05 ± 0.00 e | nd | nd | nd | nd |
| PCQ | 0.12 ± 0.01l | 0.12 ± 0.01 n | 0.08 ± 0.04 k | 0.04 ± 0.02 m | 0.02 ± 0.01 e | 0.01 ± 0.00 d | 0.01 ± 0.00 d | 0.01 ± 0.00 d | 0.17 ± 0.02 e | 0.12 ± 0.02 f | 0.05 ± 0.01 f | 0.02 ± 0.00 f |
| CAd | 0.06 ± 0.00 p | 0.06 ± 0.02 p | 0.02 ± 0.00 o | 0.03 ± 0.01 o | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.12 ± 0.00 e | 0.08 ± 0.00 g | 0.01 ± 0.00 g | 0.01 ± 0.00 f |
| EC | 1.08 ± 0.02 c | 0.70 ± 0.02 c | 0.41 ± 0.03 c | 0.36 ± 0.01 c | 0.16 ± 0.02 b | 0.09 ± 0.03 c | 0.05 ± 0.01 c | 0.05 ± 0.01 c | 2.35 ± 0.04 c | 1.65 ± 0.04 c | 0.61 ± 0.02 c | 0.60 ± 0.02 c |
| PB2 | 1.16 ± 0.34 b | 1.08 ± 0.04 b | 0.62 ± 0.04 b | 0.51 ± 0.01 b | 0.17 ± 0.01 b | 0.14 ± 0.02 b | 0.08 ± 0.01 b | 0.07 ± 0.01 b | 0.30 ± 0.00 e | 0.21 ± 0.01 e | 0.11 ± 0.00 e | 0.66 ± 0.03 c |
| PC1 | 0.84 ± 0.15 e | 0.58 ± 0.02 f | 0.40 ± 0.05 c | 0.27 ± 0.05 d | 0.12 ± 0.02 c | 0.07 ± 0.01 c | 0.05 ± 0.01 c | 0.03 ± 0.00 c | 0.81 ± 0.04 d | 0.57 ± 0.01 d | 0.61 ± 0.01 c | 0.35 ± 0.02 d |
| CAT | 0.33 ± 0.01 g | 0.42 ± 0.03 h | 0.14 ± 0.02 g | 0.07 ± 0.00 k | 0.05 ± 0.01 e | 0.05 ± 0.01 d | 0.02 ± 0.00 d | 0.01 ± 0.00 d | 0.76 ± 0.03 d | 0.53 ± 0.01 d | 0.13 ± 0.02 d | 0.08 ± 0.00 f |
| PB1 | 0.54 ± 0.04 e | 0.62 ± 0.03 e | 0.25 ± 0.04 e | 0.20 ± 0.02 g | 0.08 ± 0.01 d | 0.08 ± 0.01 c | 0.03 ± 0.00 c | 0.03 ± 0.00 c | 0.41 ± 0.03 d | 0.28 ± 0.01 e | 0.09 ± 0.02 e | 0.18 ± 0.02 e |
| PLXG | 0.29 ± 0.02 i | 0.24 ± 0.03 j | 0.12 ± 0.03 i | 0.10 ± 0.03 ij | 0.00 ± 0.01 f | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | 0.00 ± 0.00 g | 0.00 ± 0.00 h | 0.00 ± 0.00 h | 0.00 ± 0.00 g |
| PLG | 0.42 ± 0.02 f | 0.35 ± 0.02 i | 0.14 ± 0.05 f | 0.13 ± 0.01 h | 0.06 ± 0.01 d | 0.04 ± 0.01 d | 0.02 ± 0.00 d | 0.02 ± 0.00 c | 0.41 ± 0.01 d | 0.29 ± 0.01 e | 0.20 ± 0.01 e | 0.17 ± 0.00 e |
| QRUT | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd | nd |
| QGAL | 0.31 ± 0.01 h | 0.50 ± 0.03 g | 0.13 ± 0.02 h | 0.21 ± 0.02 f | 0.05 ± 0.01 d | 0.06 ± 0.01 c | 0.02 ± 0.00 d | 0.03 ± 0.01 c | 0.58 ± 0.03 d | 0.41 ± 0.00 d | 0.11 ± 0.02 e | 0.13 ± 0.00 e |
| QGLU | 0.05 ± 0.00 n | 0.14 ± 0.03 l | 0.02 ± 0.00 n | 0.04 ± 0.01 o | 0.01 ± 0.00 e | 0.02 ± 0.01 d | 0.00 ± 0.00 e | 0.01 ± 0.00 d | 0.02 ± 0.00 f | 0.02 ± 0.00 g | 0.02 ± 0.00 f | 0.01 ± 0.00 f |
| QARA | 0.13 ± 0.00 l | 0.11 ± 0.01 n | 0.04 ± 0.00 l | 0.05 ± 0.01 l | 0.02 ± 0.01 e | 0.01 ± 0.00 d | 0.01 ± 0.00 d | 0.01 ± 0.00 d | 0.07 ± 0.00 f | 0.05 ± 0.00 g | 0.06 ± 0.00 f | 0.06 ± 0.00 f |
| QXYL | 0.24 ± 0.00 j | 0.22 ± 0.04 j,k | 0.09 ± 0.01 j | 0.11 ± 0.04 i | 0.04 ± 0.01 e | 0.03 ± 0.00 d | 0.01 ± 0.00 d | 0.01 ± 0.00 d | 0.17 ± 0.00 e | 0.12 ± 0.02 f | 0.09 ± 0.02 e | 0.04 ± 0.00 f |
| QRHM | 0.11 ± 0.02 m | 0.09 ± 0.03 o | 0.03 ± 0.00 n | 0.04 ± 0.01 m | 0.02 ± 0.00 e | 0.01 ± 0.00 d | 0.00 ± 0.00 e | 0.01 ± 0.00 d | nd | nd | nd | nd |
| PO | 32.43 ± 1.6 a | 28.49 ± 5.14 a | 12.47 ± 3.34 a | 11.80 ± 2.11 a | 5.01 ± 0.06 a | 3.87 ± 0.10 a | 1.65 ± 0.06 a | 1.68 ± 0.16 a | 20.79 ± 1.11 a | 14.55 ± 1.21 a | 8.19 ± 0.07 a | 5.30 ± 0.05 a |
| DP | 4.57 | 4.42 | 4.99 | 4.19 | 4.57 | 4.27 | 4.99 | 4.19 | 5.72 | 5.55 | 5.72 | 4.05 |
| Total | 39.15 | 34.46 | 15.37 | 14.22 | 5.97 | 4.62 | 2.05 | 2.05 | 33.39 | 23.37 | 12.54 | 9.31 |
| Phenolic Compounds | Leaves after Full Bloom (Days) | |||
|---|---|---|---|---|
| 60 | 80 | 130 | 145 | |
| CQA | 1.17 ± 0.12 i,* | 1.13 ± 0.24 i | 1.46 ± 0.12 h | 16.6 ± 1.50 g |
| CGLU | nd | nd | nd | nd |
| PCQ | 0.19 ± 0.01 n | 0.25 ± 0.05 m | 0.22 ± 0.03 l | 0.22 ± 0.04 l |
| CAd | 0.20 ± 0.05 m | 0.24 ± 0.09 m | 0.20 ± 0.05 l,m | 0.19 ± 0.02 m |
| EC | 0.62 ± 0.07 j | 0.74 ± 0.06 j | 0.85 ± 0.11 j | 0.96 ± 0.04 h |
| PB2 | 0.20 ± 0.09 m | 0.20 ± 0.01 o | 0.22 ± 0.04 l | 0.29 ± 0.03 k |
| PC1 | 0.12 ± 0.01 p | 0.11 ± 0.03 p | 0.14 ± 0.01 n | 0.17 ± 0.01 n |
| CAT | 0.07 ± 0.05 r | 0.06 ± 0.02 r | 0.05 ± 0.00 o | 0.06 ± 0.01 p |
| PB1 | 0.05 ± 0.02 s | 0.05 ± 0.00 s | 0.04 ± 0.01 p | 0.05 ± 0.04 s |
| PLXG | 5.42 ± 0.16 f | 6.24 ± 0.18 c | 3.27 ± 0.34 f | 3.39 ± 0.03 f |
| PLG | 97.05 ± 3.34 a | 110.15 ± 2.43 a | 110.75 ± 1.16 a | 109.70 ± 2.57 a |
| QRUT | 0.13 ± 0.04 o | 0.69 ± 0.11 l | 0.60 ± 0.19 i | 0.76 ± 0.02 i |
| QGAL | 7.36 ± 0.13 c | 6.05 ± 0.35 d | 4.33 ± 0.23 c | 5.45 ± 0.18 c |
| QGLU | 6.77 ± 1.61 d | 5.66 ± 1.38 e | 3.87 ± 0.17 d | 4.75 ± 1.35 d |
| QARA | 3.51 ± 1.23 g | 3.93 ± 0.34 g | 3.74 ± 0.34 e | 3.80 ± 1.02 f |
| QXYL | 2.30 ± 0.91 h | 2.58 ± 0.68 h | 2.17 ± 0.21 g | 2.43 ± 0.24 g |
| QRHM | 5.52 ± 1.28 e | 4.86 ± 0.49 f | 3.85 ± 1.34 d | 4.18 ± 1.18 e |
| PO | 19.06 ± 2.16 b | 17.55 ± 1.16 b | 17.09 ± 2.51 b | 22.15 ± 3.48 b |
| DP | 9.41 | 10.06 | 9.18 | 8.42 |
| Total | 150.20 | 160.93 | 153.15 | 160.65 |
| Days after Full Bloom | DPPH | ABTS | ||||||
|---|---|---|---|---|---|---|---|---|
| Ozark Gold | Starkinson | Kosztela | Ozark Gold | Starkinson | Kosztela | |||
| Leaves | Fruits | Leaves | Fruits | |||||
| 60 | 164.7 ± 2.9 a,* | 46.1 ± 3.1 a | 20.1 ± 1.6 a | 43.9 ± 3.4 a | 17.5 ± 1.6 c | 4.8 ± 0.5 a | 2.6 ± 0.6 a | 4.5 ± 0.5 a |
| 80 | 158.3 ± 1.7 c | 31.1 ± 1.0 b | 11.7 ± 2.1 b | 30.4 ± 1.2 b | 19.9 ± 2.5 a | 4.3 ± 0.4 b | 2.0 ± 0.3 b | 4.0 ± 0.3 a |
| 130 | 153.0 ± 1.8 d | 14.4 ± 1.5 c | 8.8 ± 0.6 c | 15.7 ± 1.9 c | 18.5 ± 0.9 b | 1.4 ± 0.2 c | 1.5 ± 0.5 c | 2.6 ± 0.2 b |
| 145 | 146.7 ± 0.9 b | 13.9 ± 1.6 d | 3.6 ± 0.9 d | 11.7 ± 0.4 d | 18.7 ± 1.7 b | 1.4 ± 0.1 c | 0.9 ± 0.2 d | 1.4 ± 0.3 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojdyło, A.; Oszmiański, J. Antioxidant Activity Modulated by Polyphenol Contents in Apple and Leaves during Fruit Development and Ripening. Antioxidants 2020, 9, 567. https://doi.org/10.3390/antiox9070567
Wojdyło A, Oszmiański J. Antioxidant Activity Modulated by Polyphenol Contents in Apple and Leaves during Fruit Development and Ripening. Antioxidants. 2020; 9(7):567. https://doi.org/10.3390/antiox9070567
Chicago/Turabian StyleWojdyło, Aneta, and Jan Oszmiański. 2020. "Antioxidant Activity Modulated by Polyphenol Contents in Apple and Leaves during Fruit Development and Ripening" Antioxidants 9, no. 7: 567. https://doi.org/10.3390/antiox9070567
APA StyleWojdyło, A., & Oszmiański, J. (2020). Antioxidant Activity Modulated by Polyphenol Contents in Apple and Leaves during Fruit Development and Ripening. Antioxidants, 9(7), 567. https://doi.org/10.3390/antiox9070567