Neuroprotective Potential of Verbascoside Isolated from Acanthus mollis L. Leaves through Its Enzymatic Inhibition and Free Radical Scavenging Ability

 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. General Experimental Procedure and Chemicals
2.2. Plant Material and Extract Preparation
2.3. Extract Fractionation and Isolation
2.4. Cell Culture
2.5. In Vitro Antioxidant Activity Assay
2.5.1. DPPH• Scavenging Assay
2.5.2. Xanthine/Xanthine Oxidase Assay
2.5.3. Hydroxyl Radical Scavenging Capacity
2.5.4. Total Antioxidant Capacity (TAC)
2.5.5. Cell Culture Antioxidant Capacity. Intracellular ROS Measurement
2.6. Neuroprotective Capacity. In Vitro Enzymatic Inhibition Capacity
2.6.1. Tyrosinase Inhibition Assay
2.6.2. MAO-A Inhibition Assay
2.6.3. Acetylcholinesterase (AChE) Inhibition Assay
2.7. Statistical Analysis
3. Results
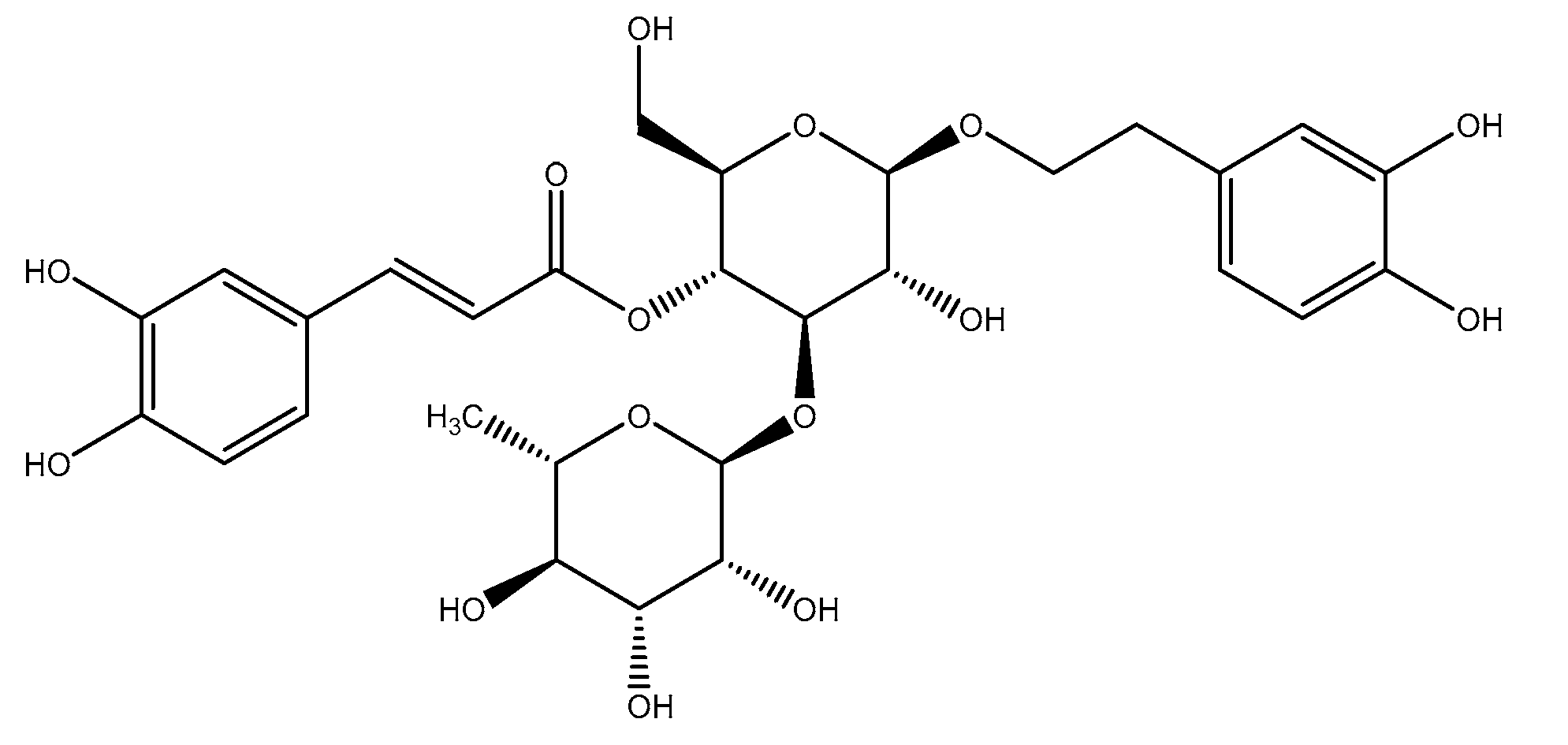
3.1. Isolation and Identification of Compound 1
3.2. In Vitro Antioxidant Activity
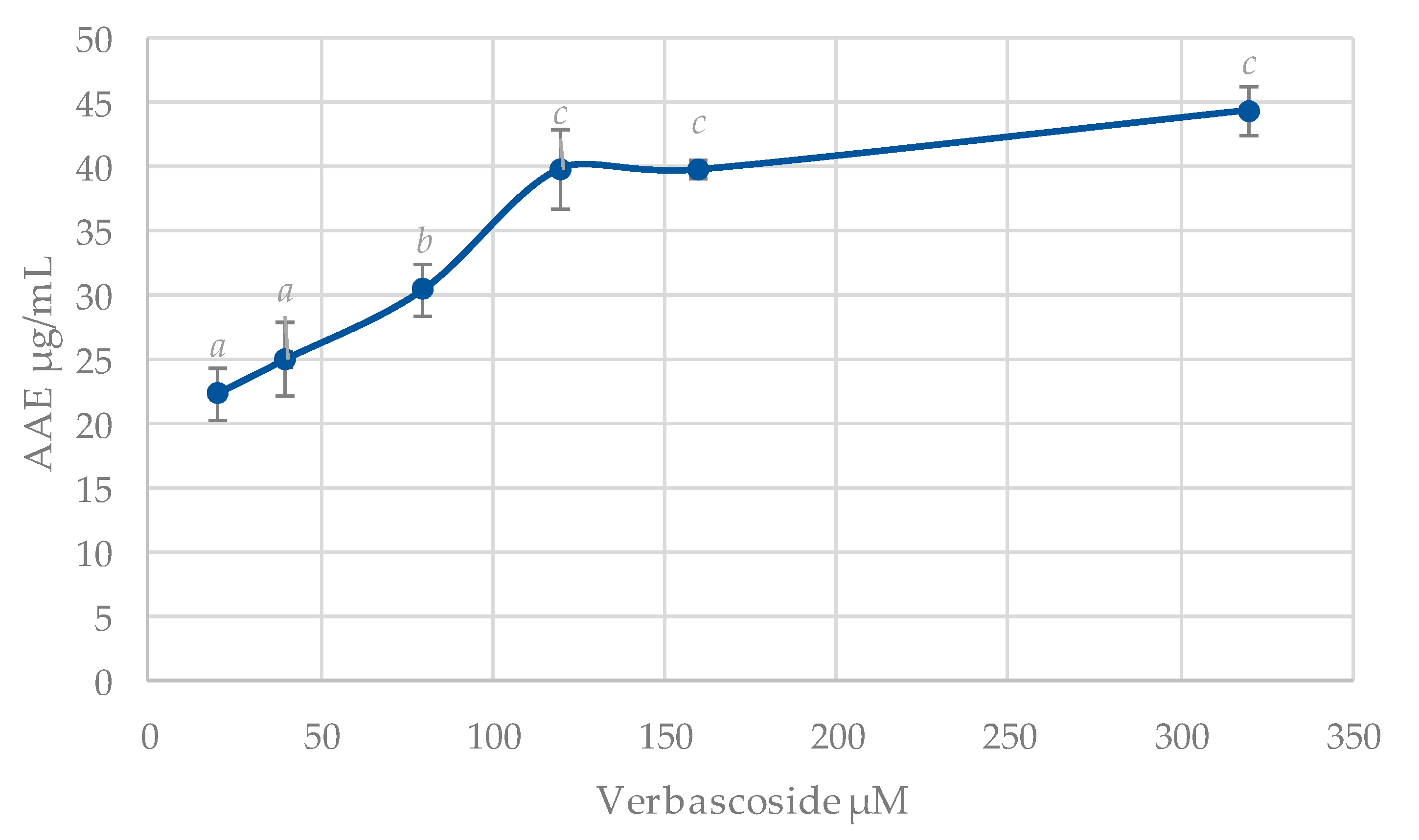
3.3. Total Antioxidant Capacity (TAC)
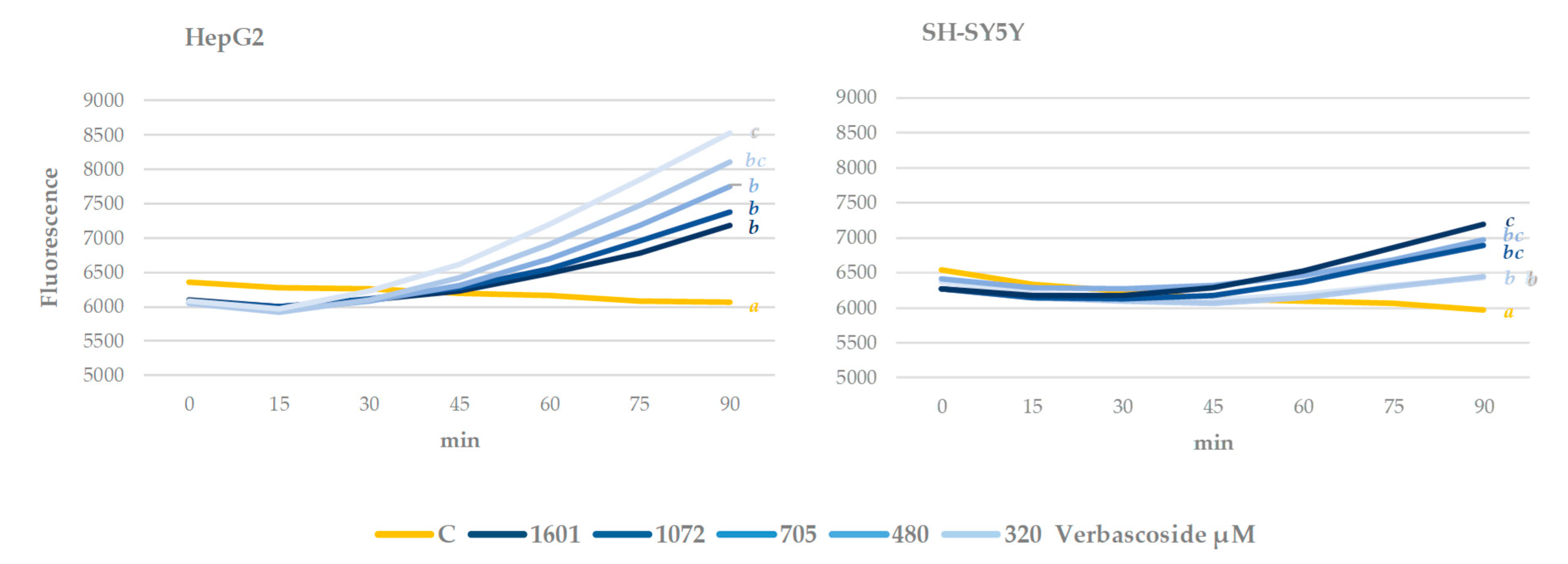
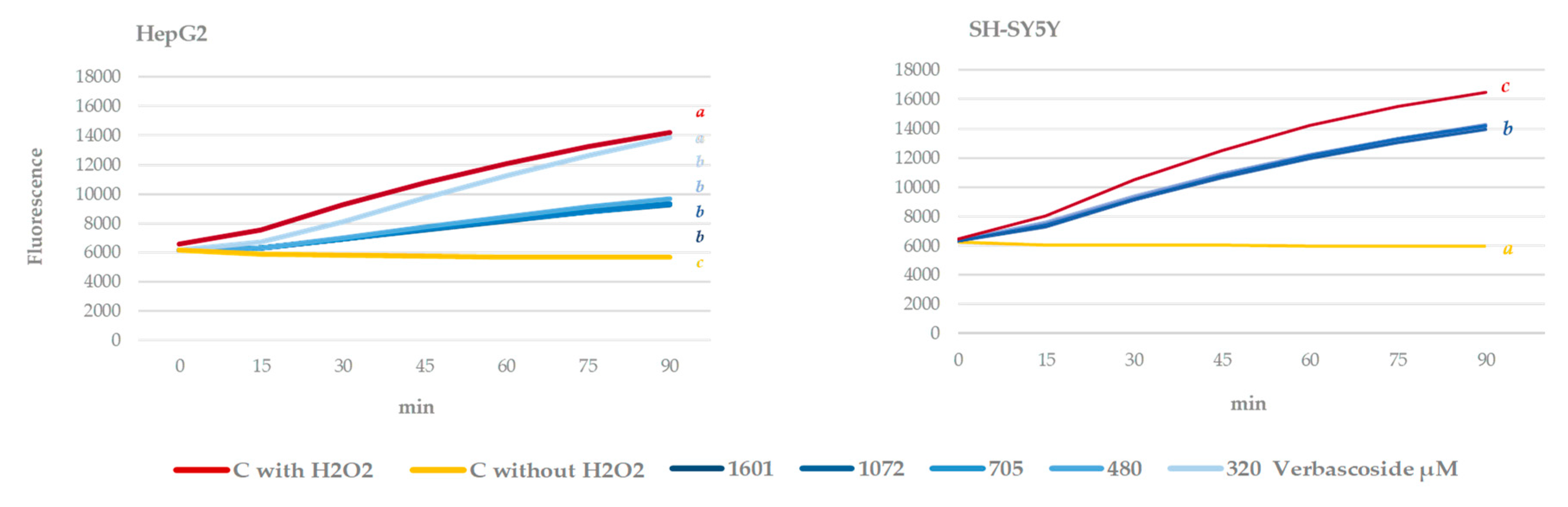
3.4. Cell Culture Antioxidant Capacity. Intracellular ROS Measurement
3.5. Neuroprotective Capacity. In Vitro Enzymatic Inhibition Capacity
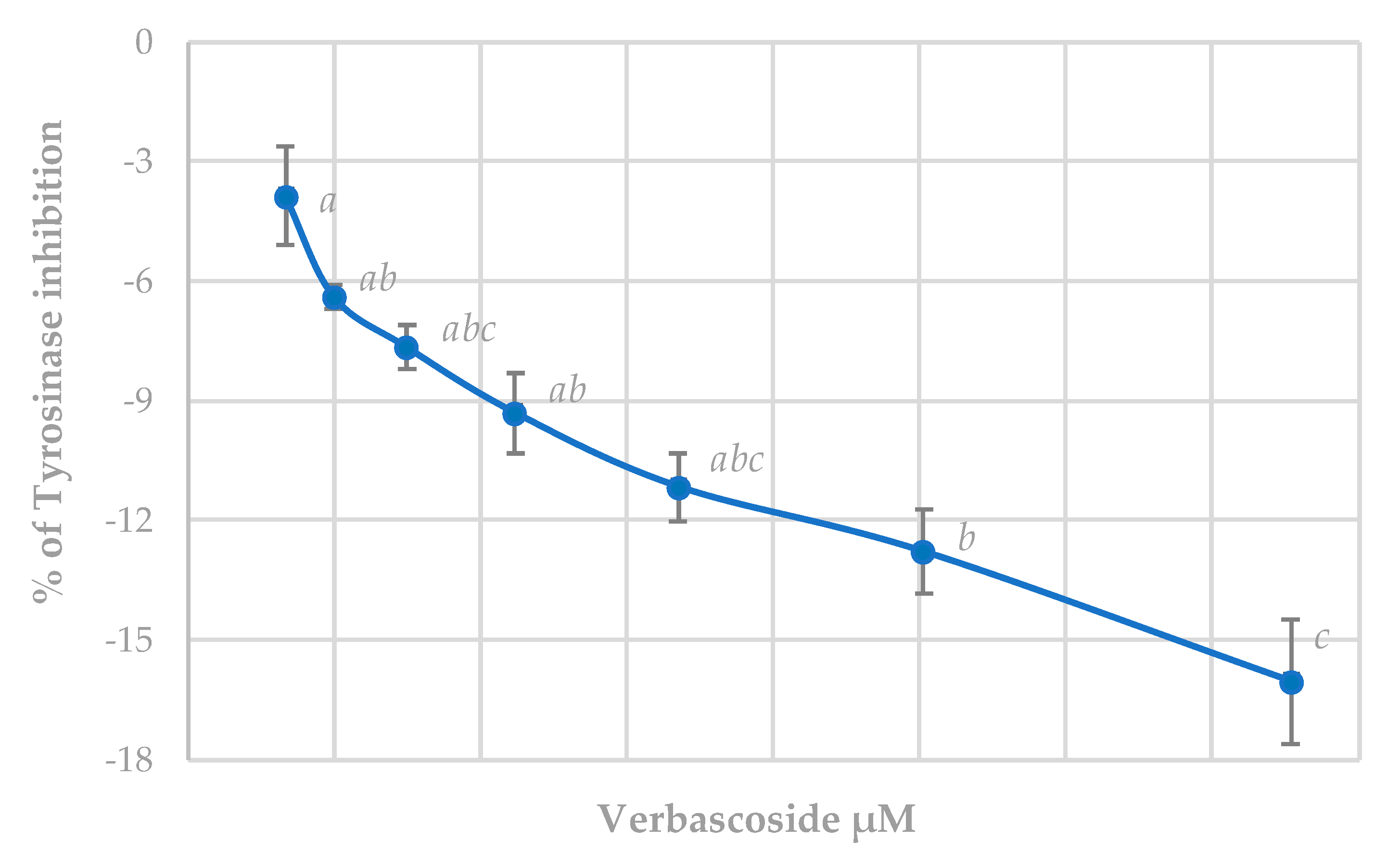
3.5.1. Tyrosinase Inhibition Assay
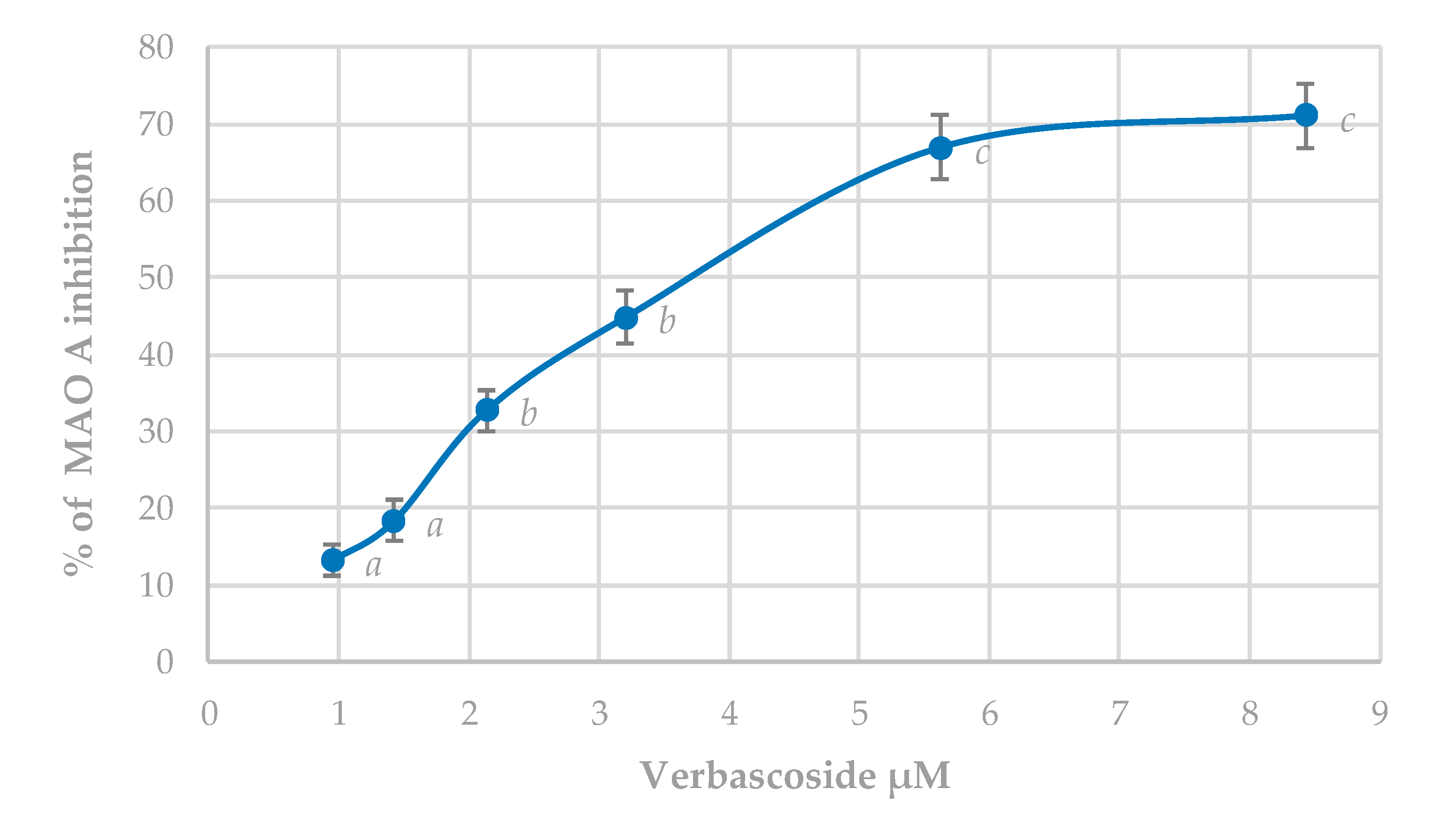
3.5.2. MAO-A Inhibition Assay
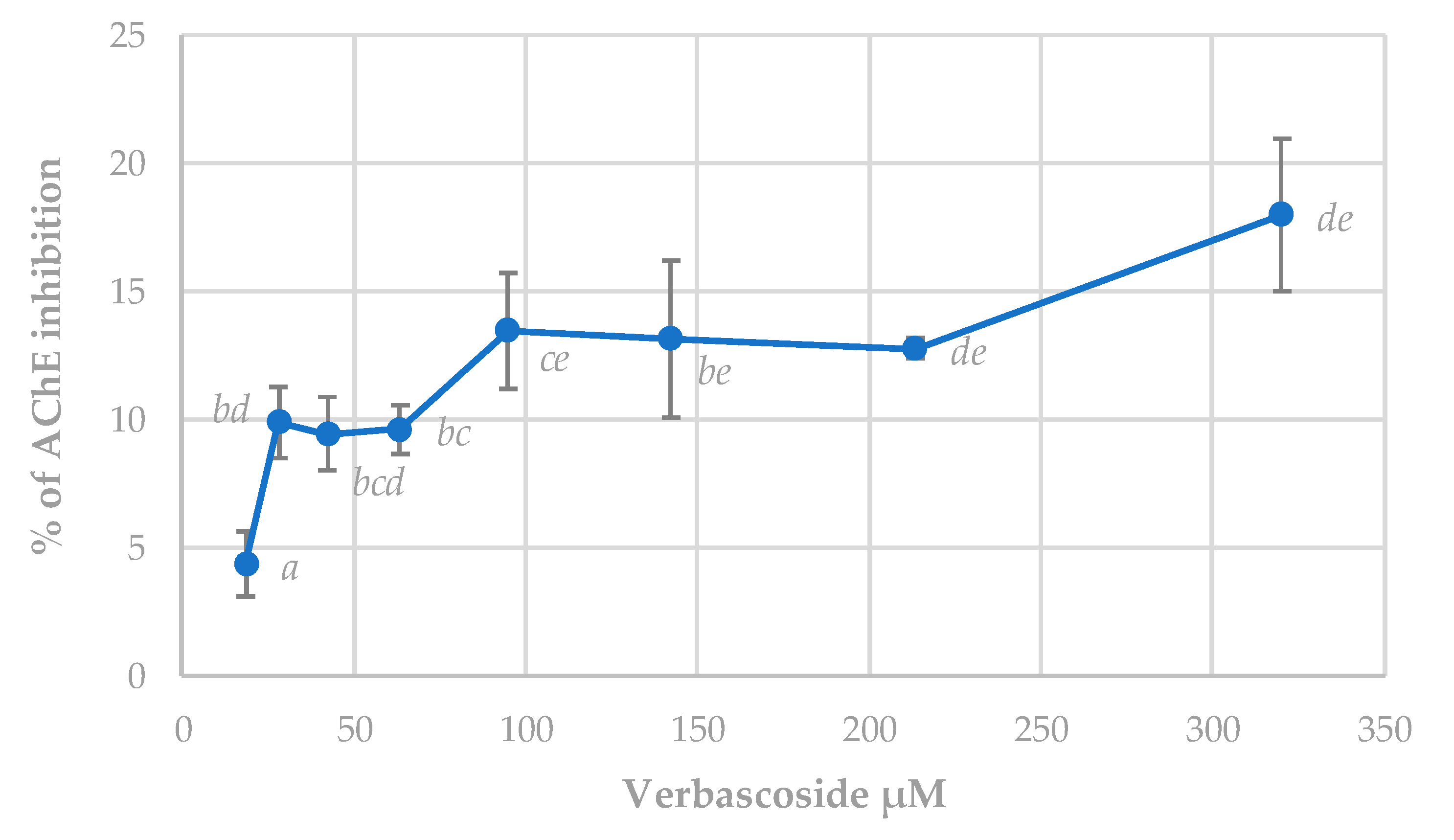
3.5.3. Acetylcholinesterase (AChE) Inhibition Assay
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hügel, H.M. Brain Food for Alzheimer-Free Ageing: Focus on Herbal Medicines. In Natural Compounds as Therapeutic Agents for Amyloidogenic Diseases; Springer: Cham, Switzerland, 2015; pp. 95–116. [Google Scholar]
- Hardy, J.; Selkoe, D.J. The Amyloid Hypothesis of Alzheimer’s Disease: Progress and Problems on the Road to Therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Mufson, E.J.; Counts, S.E.; Perez, S.E.; Ginsberg, S.D. Cholinergic system during the progression of Alzheimer’s disease: Therapeutic implications. Expert Rev. Neurother. 2008, 8, 1703–1718. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.H.; Cuong, T.D.; Hung, T.M.; Kim, J.A.; Woo, M.H.; Byeon, J.S.; Choi, J.S.; Min, B.S. Cholinesterase inhibitors from the roots of Harpagophytum procumbens. Arch. Pharmacol. Res. 2014, 37, 1124–1129. [Google Scholar] [CrossRef] [PubMed]
- Mendes, E.; Perry, M.d.J.; Francisco, A.P. Design and discovery of mushroom tyrosinase inhibitors and their therapeutic applications. Expert Opin. Drug Discov. 2014, 9, 533–554. [Google Scholar] [CrossRef]
- Pan, T.; Li, X.; Jankovic, J. The association between Parkinson’s disease and melanoma. Int. J. Cancer 2011, 128, 2251–2260. [Google Scholar] [CrossRef]
- Youdim, M.B.H.; Bakhle, Y.S. Monoamine oxidase: Isoforms and inhibitors in Parkinson’s disease and depressive illness. Br. J. Pharmacol. 2009, 147, S287–S296. [Google Scholar] [CrossRef]
- Joubert, J.; Petzer, J.P.; Prins, L.H.A.; Repsold, B.P.; Malan, S.F. Multifunctional enzyme inhibition for neuroprotection—A focus on MAO, NOS, and AChE inhibitors. In Drug Design and Discovery in Alzheimer’s Disease; Elsevier Inc.: Amsterdam, The Netherlands, 2014; pp. 291–365. ISBN 9780128039595. [Google Scholar]
- Tan, J.-L.; Li, Q.-X.; Ciccotosto, G.D.; Crouch, P.J.; Culvenor, J.G.; White, A.R.; Evin, G. Mild Oxidative Stress Induces Redistribution of BACE1 in Non-Apoptotic Conditions and Promotes the Amyloidogenic Processing of Alzheimer’s Disease Amyloid Precursor Protein. PLoS ONE 2013, 8, e61246. [Google Scholar] [CrossRef]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxid. Med. Cell. Longev. 2016, 2016. [Google Scholar] [CrossRef]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Ann. Rev. Biochem. 2017, 86. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative Stress and Antioxidant Defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef]
- Radak, Z.; Suzuki, K.; Higuchi, M.; Balogh, L.; Boldogh, I.; Koltai, E. Physical exercise, reactive oxygen species and neuroprotection. Free Radic. Biol. Med. 2016, 98, 187–196. [Google Scholar] [CrossRef]
- Gay, N.H.; Phopin, K.; Suwanjang, W.; Songtawee, N.; Ruankham, W.; Wongchitrat, P.; Prachayasittikul, S.; Prachayasittikul, V. Neuroprotective Effects of Phenolic and Carboxylic Acids on Oxidative Stress-Induced Toxicity in Human Neuroblastoma SH-SY5Y Cells. Neurochem. Res. 2018, 43, 619–636. [Google Scholar] [CrossRef]
- Wang, F.; Zhi, J.; Zhang, Z.; Wang, L.; Suo, Y.; Xie, C.; Li, M.; Zhang, B.; Du, J.; Gu, L.; et al. Transcriptome analysis of salicylic acid treatment in Rehmannia glutinosa hairy roots using RNA-seq technique for identification of genes involved in acteoside biosynthesis. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Georgiev, M.; Pastore, S.; Lulli, D.; Alipieva, K.; Kostyuk, V.; Potapovich, A.; Panetta, M.; Korkina, L. Verbascum xanthophoeniceum-derived phenylethanoid glycosides are potent inhibitors of inflammatory chemokines in dormant and interferon-gamma-stimulated human keratinocytes. J. Ethnopharmacol. 2012, 144, 754–760. [Google Scholar] [CrossRef]
- Alipieva, K.; Korkina, L.; Orhan, I.E.; Georgiev, M.I. Verbascoside—A review of its occurrence, (bio)synthesis and pharmacological significance. Biotechnol. Adv. 2014, 32, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Shiao, Y.J.; Su, M.H.; Lin, H.C.; Wu, C.R. Acteoside and isoacteoside protect amyloid β peptide induced cytotoxicity, cognitive deficit and neurochemical disturbances in vitro and in vivo. Int. J. Mol. Sci. 2017, 18, 895. [Google Scholar] [CrossRef]
- Quirantes-Piné, R.; Herranz-López, M.; Funes, L.; Borrás-Linares, I.; Micol, V.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Phenylpropanoids and their metabolites are the major compounds responsible for blood-cell protection against oxidative stress after administration of Lippia citriodora in rats. Phytomedicine 2013, 20, 1112–1118. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.L.; Cao, K.K.; Zhao, X.X.; Kang, N.X.; Zhang, Y.; Xu, Q.M.; Yang, S.L.; Liu, Y.L.; Wang, C. Antioxidant activity of phenylethanoid glycosides on glutamate-induced neurotoxicity. Biosci. Biotechnol. Biochem. 2019, 83, 2016–2026. [Google Scholar] [CrossRef]
- Matos, P.; Figueirinha, A.; Paranhos, A.; Nunes, F.; Cruz, P.; Geraldes, C.F.G.C.; Cruz, M.T.; Batista, M.T. Bioactivity of Acanthus mollis—Contribution of benzoxazinoids and phenylpropanoids. J. Ethnopharmacol. 2018, 227, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Jara, C.; Leyton, M.; Osorio, M.; Silva, V.; Fleming, F.; Paz, M.; Madrid, A.; Mellado, M. Antioxidant, phenolic and antifungal profiles of Acanthus mollis (Acanthaceae). Nat. Prod. Res. 2017, 31, 2325–2328. [Google Scholar] [CrossRef] [PubMed]
- Goya, L.; Martin, M.; Ramos, S.; Mateos, R.; Bravo, L. A Cell Culture Model for the Assessment of the Chemopreventive Potential of Dietary Compounds. Curr. Nutr. Food Sci. 2009, 5, 56–64. [Google Scholar] [CrossRef][Green Version]
- Marrazzo, P.; Angeloni, C.; Hrelia, S. Combined Treatment with Three Natural Antioxidants Enhances Neuroprotection in a SH-SY5Y 3D Culture Model. Antioxidants 2019, 8, 420. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.R.; Hu, L.S.; Li, G.Y. SH-SY5Y human neuroblastoma cell line: In vitro cell model of dopaminergic neurons in Parkinson’s disease. Chin. Med. J. (Engl.) 2010, 123, 1086–1092. [Google Scholar] [PubMed]
- Long, L.H.; Halliwell, B. Artefacts in cell culture: α-Ketoglutarate can scavenge hydrogen peroxide generated by ascorbate and epigallocatechin gallate in cell culture media. Biochem. Biophys. Res. Commun. 2011, 406, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Acero, N.; Muñoz-Mingarro, D. Effect on tumor necrosis factor-α production and antioxidant ability of black alder, as factors related to its anti-inflammatory properties. J. Med. Food 2012, 15. [Google Scholar] [CrossRef]
- Acero, N.; Gradillas, A.; Beltran, M.; García, A.; Muñoz Mingarro, D. Comparison of phenolic compounds profile and antioxidant properties of different sweet cherry (Prunus avium L.) varieties. Food Chem. 2019, 279, 260–271. [Google Scholar] [CrossRef]
- Sharma, P.; Ravikumar, G.; Kalaiselvi, M.; Gomathi, D.; Uma, C. In vitro antibacterial and free radical scavenging activity of green hull of Juglans regia. J. Pharm. Anal. 2013, 3, 298–302. [Google Scholar] [CrossRef]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric Quantitation of Antioxidant Capacity through the Formation of a Phosphomolybdenum Complex: Specific Application to the Determination of Vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar] [CrossRef]
- Muñoz Mingarro, D.; Plaza, A.; Galán, A.; Vicente, J.A.; Martínez, M.P.; Acero, N. The effect of five Taraxacum species on in vitro and in vivo antioxidant and antiproliferative activity. Food Funct. 2015, 6, 2787–2793. [Google Scholar] [CrossRef]
- Cásedas, G.; Les, F.; Gómez-Serranillos, M.P.; Smith, C.; López, V. Bioactive and functional properties of sour cherry juice (Prunus cerasus). Food Funct. 2016, 7, 4675–4682. [Google Scholar] [CrossRef]
- Olsen, H.T.; Stafford, G.I.; van Staden, J.; Christensen, S.B.; Jäger, A.K. Isolation of the MAO-inhibitor naringenin from Mentha aquatica L. J. Ethnopharmacol. 2008, 117, 500–502. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G.; Bae, Y.S.; Lee, S.-R.; Kwon, J. Hydrogen Peroxide: A Key Messenger That Modulates Protein Phosphorylation Through Cysteine Oxidation. Sci. Signal. 2000, 2000, pe1. [Google Scholar] [CrossRef] [PubMed]
- Lina, H.Z.; Samy, M.M.; Samir, A.E.B.; Fatma, A.M.; Kawther, M.T.; Abdelaaty, A.S. Hypoglycemic and antioxidant effects of Hibiscus rosa-sinensis L. leaves extract on liver and kidney damage in streptozotocin induced diabetic rats. Afr. J. Pharm. Pharmacol. 2017, 11, 161–169. [Google Scholar] [CrossRef]
- Govindan, P.; Muthukrishnan, S. Evaluation of total phenolic content and free radical scavenging activity of Boerhavia erecta. J. Acute Med. 2013, 3, 103–109. [Google Scholar] [CrossRef]
- Kaur, A.; Dhari, J.; Prakash Sharma, O.; Gupta, G.D.; Kharb, V. In vitro antioxidant and free radical scavenging activity of lycopene. Res. J. Pharm. Biol. Chem. Sci. 2012, 3, 1220–1228. [Google Scholar]
- Pavithra, K.; Vadivukkarasi, S. Evaluation of free radical scavenging activity of various extracts of leaves from Kedrostis foetidissima (Jacq.) Cogn. Food Sci. Hum. Wellness 2015, 4, 42–46. [Google Scholar] [CrossRef]
- Pietta, P.-G. Flavonoids as Antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef]
- Parejo, I.; Viladomat, F.; Bastida, J.; Rosas-Romero, A.; Flerlage, N.; Burillo, J.; Codina, C. Comparison between the Radical Scavenging Activity and Antioxidant Activity of Six Distilled and Nondistilled Mediterranean Herbs and Aromatic Plants. J. Agric. Food Chem. 2002, 50, 6882–6890. [Google Scholar] [CrossRef]
- Liu, F.; Ng, T.B. Antioxidative and free radical scavenging activities of selected medicinal herbs. Life Sci. 2000, 66, 725–735. [Google Scholar] [CrossRef]
- Lee, J.-C.; Kim, H.-R.; Kim, J.; Jang, Y.-S. Antioxidant Property of an Ethanol Extract of the Stem of Opuntia ficus-indica var. Saboten. J. Agric. Food Chem. 2002, 50, 6490–6496. [Google Scholar] [CrossRef]
- Battu, G.R.; Ethadi, S.R.; Veda Priya, G.; Swathi Priya, K.; Chandrika, K.; Venkateswara Rao, A.; Reddy, S.O. Evaluation of antioxidant and anti-inflammatory activity of Euphorbia heyneana Spreng. Asian Pac. J. Trop. Biomed. 2011, 1, S191–S194. [Google Scholar] [CrossRef]
- Harput, U.S.; Arihan, O.; Iskit, A.B.; Nagatsu, A.; Saracoglu, I. Antinociceptive, Free Radical–Scavenging, and Cytotoxic Activities of Acanthus hirsutus Boiss. J. Med. Food 2011, 14, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Asongalem, E.A.; Foyet, H.S.; Ekobo, S.; Dimo, T.; Kamtchouing, P. Antiinflammatory, lack of central analgesia and antipyretic properties of Acanthus montanus (Ness) T. Anderson. J. Ethnopharmacol. 2004, 95, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Marmitt, D.J.; Alves, C.; Silva, J.; Pinteus, S.; Schneider, T.; Christ Vianna Santos, R.; de Freitas, E.M.; Pedrosa, R.; Laufer, S.; Goettert, M.I. Neuroprotective potential of Myrciaria plinioides D. Legrand extract in an in vitro human neuroblastoma model. Inflammopharmacology 2020, 28, 737–748. [Google Scholar] [CrossRef]
- Dell’Aquila, M.E.; Bogliolo, L.; Russo, R.; Martino, N.A.; Filioli Uranio, M.; Ariu, F.; Amati, F.; Sardanelli, A.M.; Linsalata, V.; Ferruzzi, M.G.; et al. Prooxidant Effects of Verbascoside, a Bioactive Compound from Olive Oil Mill Wastewater, on In Vitro Developmental Potential of Ovine Prepubertal Oocytes and Bioenergetic/Oxidative Stress Parameters of Fresh and Vitrified Oocytes. Biomed. Res. Int. 2014, 2014, 1–14. [Google Scholar] [CrossRef]
- Hong, J.; Lu, H.; Meng, X.; Ryu, J.-H.; Hara, Y.; Hara, Y. Stability, cellular uptake, biotransformation, and efflux of tea polyphenol (−)-epigallocatechin-3-gallate in HT-29 human colon adenocarcinoma cells. Cancer Res. 2002, 62, 7241–7246. [Google Scholar]
- Uttara, B.; Singh, A.; Zamboni, P.; Mahajan, R. Oxidative Stress and Neurodegenerative Diseases: A Review of Upstream and Downstream Antioxidant Therapeutic Options. Curr. Neuropharmacol. 2009, 7, 65–74. [Google Scholar] [CrossRef]
- Rhee, I.K.; van de Meent, M.; Ingkaninan, K.; Verpoorte, R. Screening for acetylcholinesterase inhibitors from Amaryllidaceae using silica gel thin-layer chromatography in combination with bioactivity staining. J. Chromatogr. A 2001, 915, 217–223. [Google Scholar] [CrossRef]
- Stone, J.R.; Yang, S. Hydrogen Peroxide: A Signaling Messenger. Antioxid. Redox Signal. 2006, 8, 243–270. [Google Scholar] [CrossRef]
- Veal, E.A.; Day, A.M.; Morgan, B.A. Hydrogen Peroxide Sensing and Signaling. Mol. Cell 2007, 26, 1–14. [Google Scholar] [CrossRef]
- Huang, Y.Z.; McNamara, J.O. Neuroprotective Effects of Reactive Oxygen Species Mediated by BDNF-Independent Activation of TrkB. J. Neurosci. 2012, 32, 15521–15532. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.J.; Reichardt, L.F. Trk Receptors: Roles in Neuronal Signal Transduction. Annu. Rev. Biochem. 2003, 72, 609–642. [Google Scholar] [CrossRef] [PubMed]
- Chiaino, E.; Micucci, M.; Cosconati, S.; Novellino, E.; Budriesi, R.; Chiarini, A.; Frosini, M. Olive Leaves and Hibiscus Flowers Extracts-Based Preparation Protect Brain from Oxidative Stress-Induced Injury. Antioxidants 2020, 9, 806. [Google Scholar] [CrossRef] [PubMed]
- Das, J.; Ramani, R.; Suraju, M.O. Polyphenol compounds and PKC signaling. Biochim. Biophys. Acta (BBA) Gen. Subj. 2016, 1860, 2107–2121. [Google Scholar] [CrossRef]
- Herbert, J.M.; Maffrand, J.P.; Taoubi, K.; Augereau, J.M.; Fouraste, I.; Gleye, J. Verbascoside Isolated from Lantana camara, an Inhibitor of Protein Kinase C. J. Nat. Prod. 1991, 54. [Google Scholar] [CrossRef]
- An, Z.; Qi, Y.; Huang, D.; Gu, X.; Tian, Y.; Li, P.; Li, H.; Zhang, Y. EGCG inhibits Cd 2+ -induced apoptosis through scavenging ROS rather than chelating Cd 2+ in HL-7702 cells. Toxicol. Mech. Methods 2014, 24, 259–267. [Google Scholar] [CrossRef]
- Yang, J.; Yan, Y.; Liu, H.; Wang, J.; Hu, J. Protective effects of acteoside against X-ray-induced damage in human skin fibroblasts. Mol. Med. Rep. 2015, 12, 2301–2306. [Google Scholar] [CrossRef]
- Omar, S.; Kerr, P.; Scott, C.; Hamlin, A.; Obied, H. Olive (Olea europaea L.) Biophenols: A Nutriceutical against Oxidative Stress in SH-SY5Y Cells. Molecules 2017, 22, 1858. [Google Scholar] [CrossRef]
- Azimi, H.; Fallah-Tafti, M.; Khakshur, A.A.; Abdollahi, M. A review of phytotherapy of acne vulgaris: Perspective of new pharmacological treatments. Fitoterapia 2012, 83, 1306–1317. [Google Scholar] [CrossRef]
- Larit, F.; Elokely, K.M.; Chaurasiya, N.D.; Benyahia, S.; Nael, M.A.; León, F.; Abu-Darwish, M.S.; Efferth, T.; Wang, Y.-H.; Belouahem-Abed, D.; et al. Inhibition of human monoamine oxidase A and B by flavonoids isolated from two Algerian medicinal plants. Phytomedicine 2018, 40, 27–36. [Google Scholar] [CrossRef]
- Costa, F.H.d.R.; Rosso, A.L.Z.; Maultasch, H.; Nicaretta, D.H.; Vincent, M.B. Depression in Parkinson’s disease: Diagnosis and treatment. Arquivos De Neuro-Psiquiatria 2012, 70, 617–620. [Google Scholar] [CrossRef] [PubMed]
- Tougu, V. Acetylcholinesterase: Mechanism of Catalysis and Inhibition. Curr. Med. Chem. Cent. Nerv. Syst. Agents 2001, 1, 155–170. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DPPH• | Superoxide (O2•−) | Hydroxyl (•OH) | |
|---|---|---|---|
| Verbascoside | 58.1 µM ± 0.6 | 24.4 µM ± 1.4 | 357 µM ± 16.8 |
| Ascorbic acid | 284.9 µM ± 1.2 | 66.1 µM ± 1.8 | 1031 µM ± 19.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burgos, C.; Muñoz-Mingarro, D.; Navarro, I.; Martín-Cordero, C.; Acero, N. Neuroprotective Potential of Verbascoside Isolated from Acanthus mollis L. Leaves through Its Enzymatic Inhibition and Free Radical Scavenging Ability. Antioxidants 2020, 9, 1207. https://doi.org/10.3390/antiox9121207
Burgos C, Muñoz-Mingarro D, Navarro I, Martín-Cordero C, Acero N. Neuroprotective Potential of Verbascoside Isolated from Acanthus mollis L. Leaves through Its Enzymatic Inhibition and Free Radical Scavenging Ability. Antioxidants. 2020; 9(12):1207. https://doi.org/10.3390/antiox9121207
Chicago/Turabian StyleBurgos, Carmen, Dolores Muñoz-Mingarro, Inmaculada Navarro, Carmen Martín-Cordero, and Nuria Acero. 2020. "Neuroprotective Potential of Verbascoside Isolated from Acanthus mollis L. Leaves through Its Enzymatic Inhibition and Free Radical Scavenging Ability" Antioxidants 9, no. 12: 1207. https://doi.org/10.3390/antiox9121207
APA StyleBurgos, C., Muñoz-Mingarro, D., Navarro, I., Martín-Cordero, C., & Acero, N. (2020). Neuroprotective Potential of Verbascoside Isolated from Acanthus mollis L. Leaves through Its Enzymatic Inhibition and Free Radical Scavenging Ability. Antioxidants, 9(12), 1207. https://doi.org/10.3390/antiox9121207