
Circular Animal Protein Hydrolysates: A Comparative Approach of Functional Properties

 ,
,  ,
,  , ,
, ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. By-Products Source and Hydrolysate Production
2.2. Chemical Analysis
2.3. Amino Acid Profile and Peptide Molecular Weight Distribution
2.4. Protein Solubility
2.5. Antioxidant Activity
2.6. Impact on Bacterial Proliferation Dynamics
2.7. Statistical Analysis
3. Results
3.1. Hydrolysates Proximate Composition
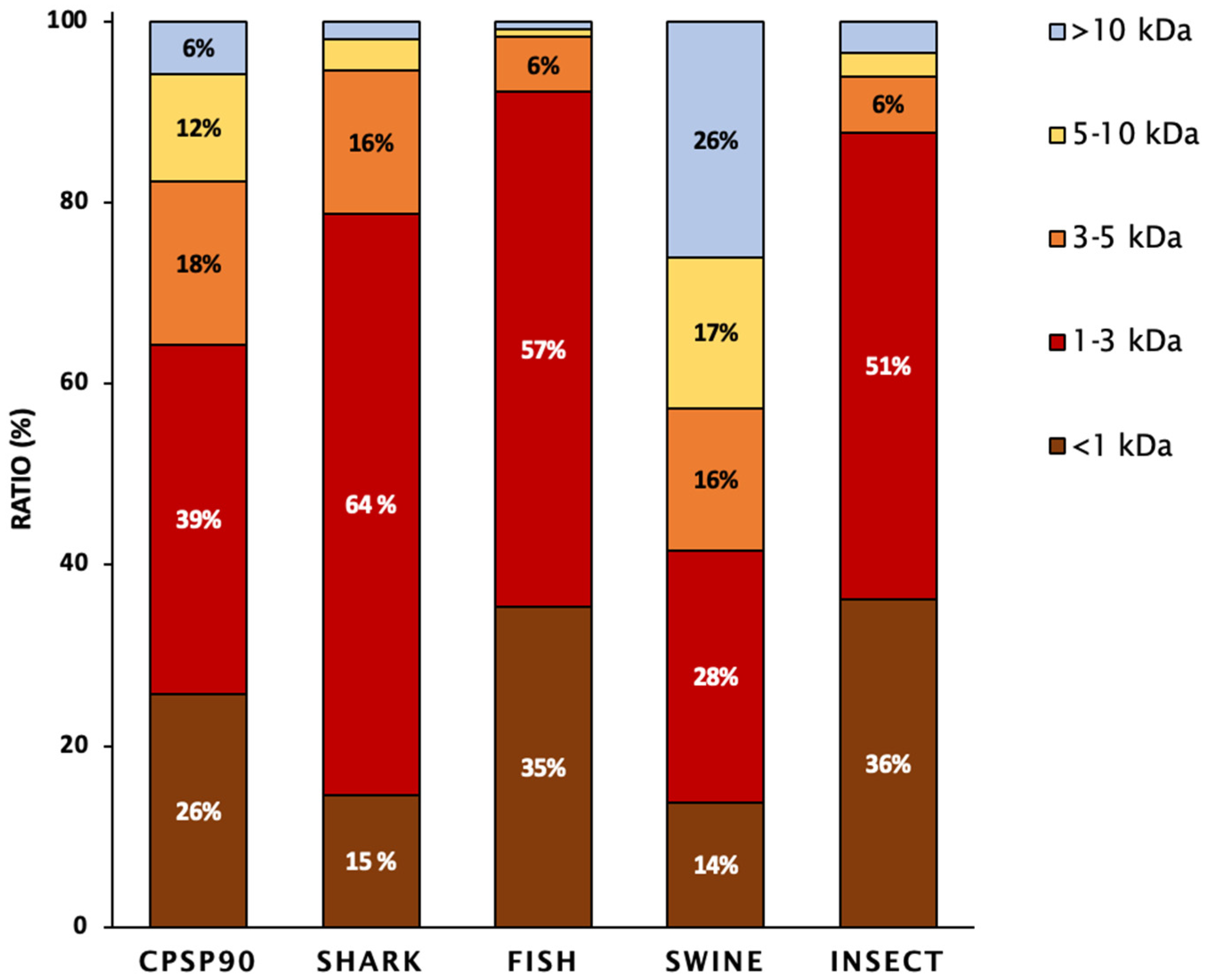
3.2. Amino Acid Profile and Peptide Molecular Weight Distribution
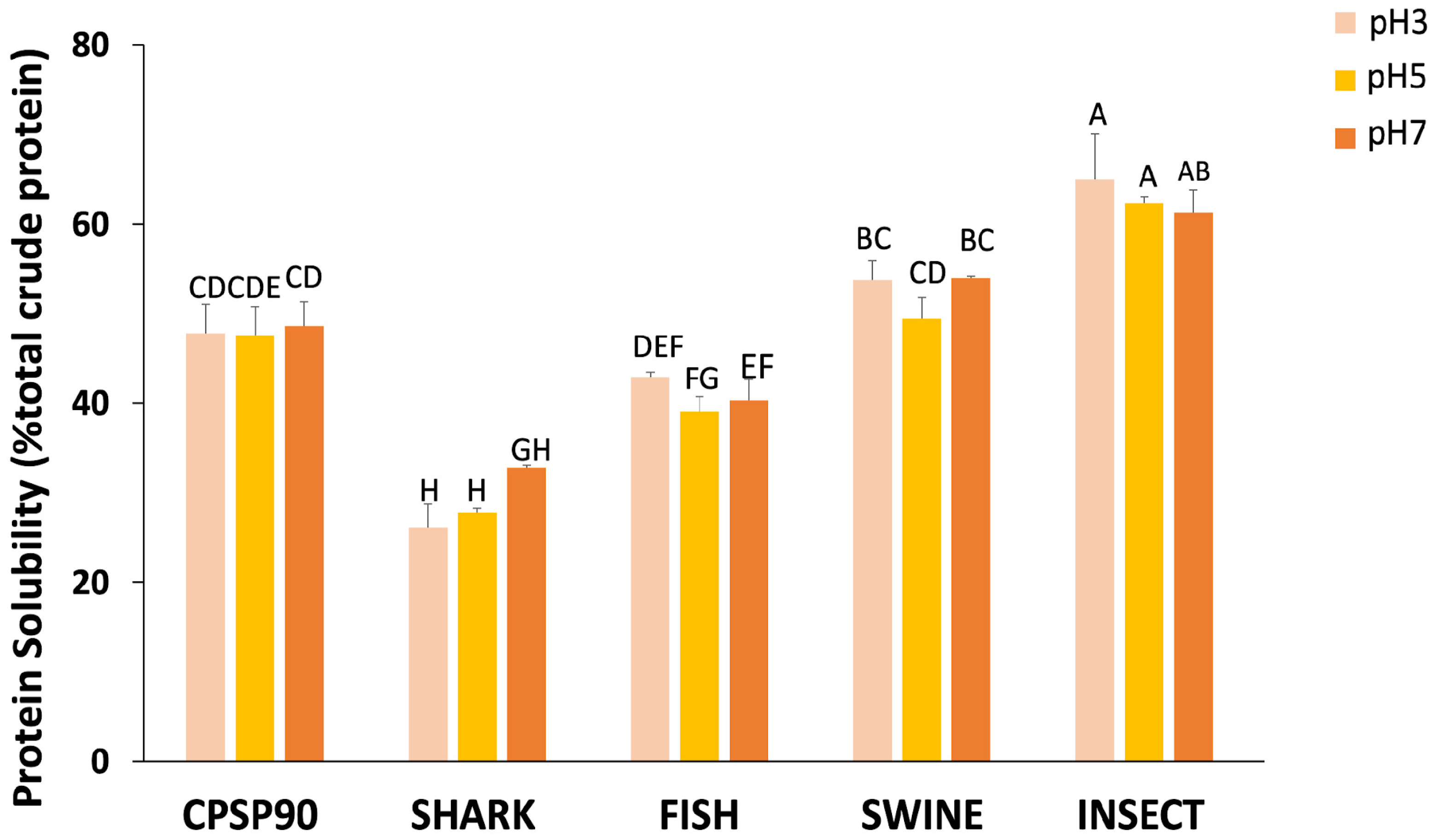
3.3. Protein Solubility
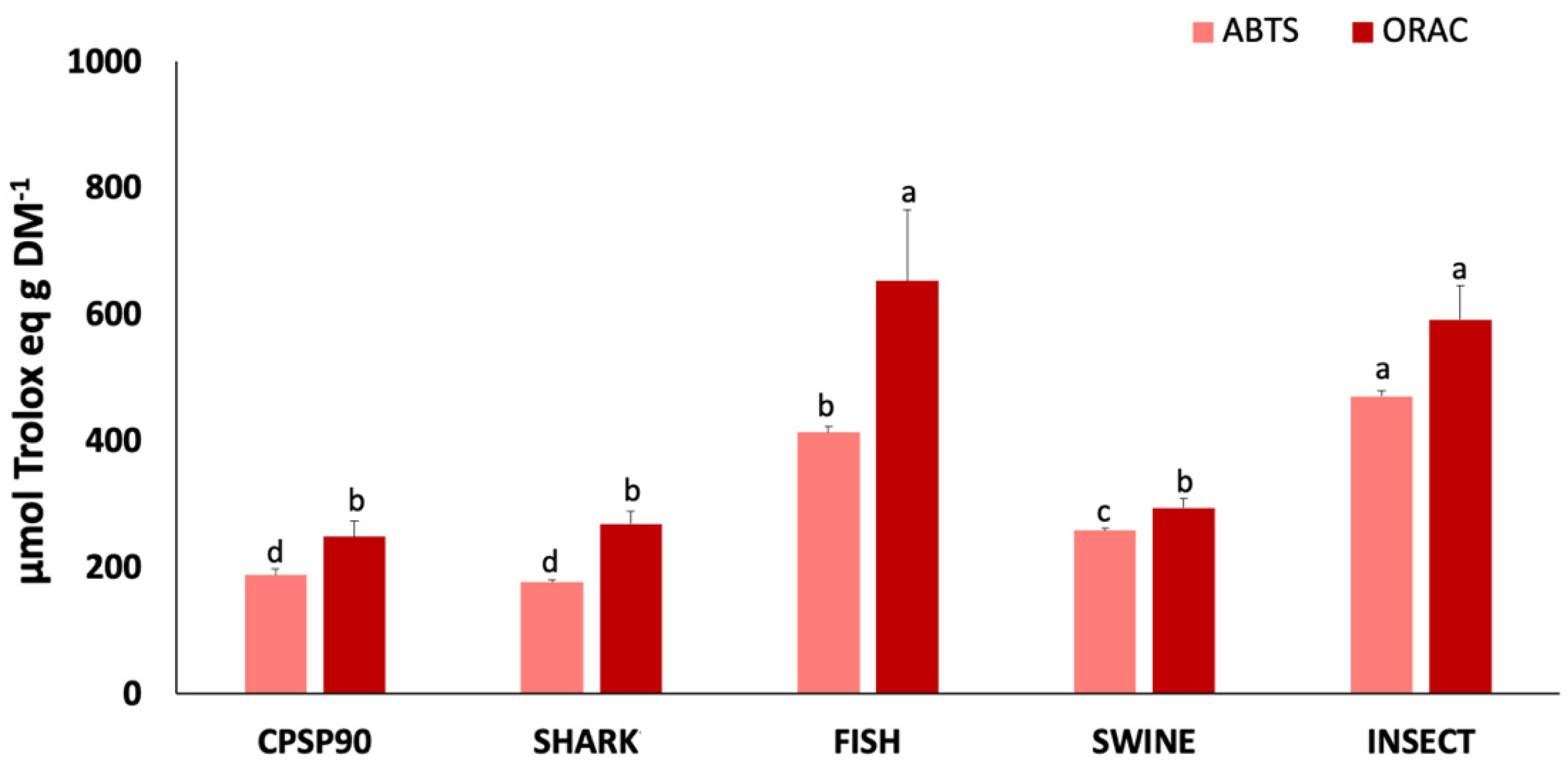
3.4. Antioxidant Activity
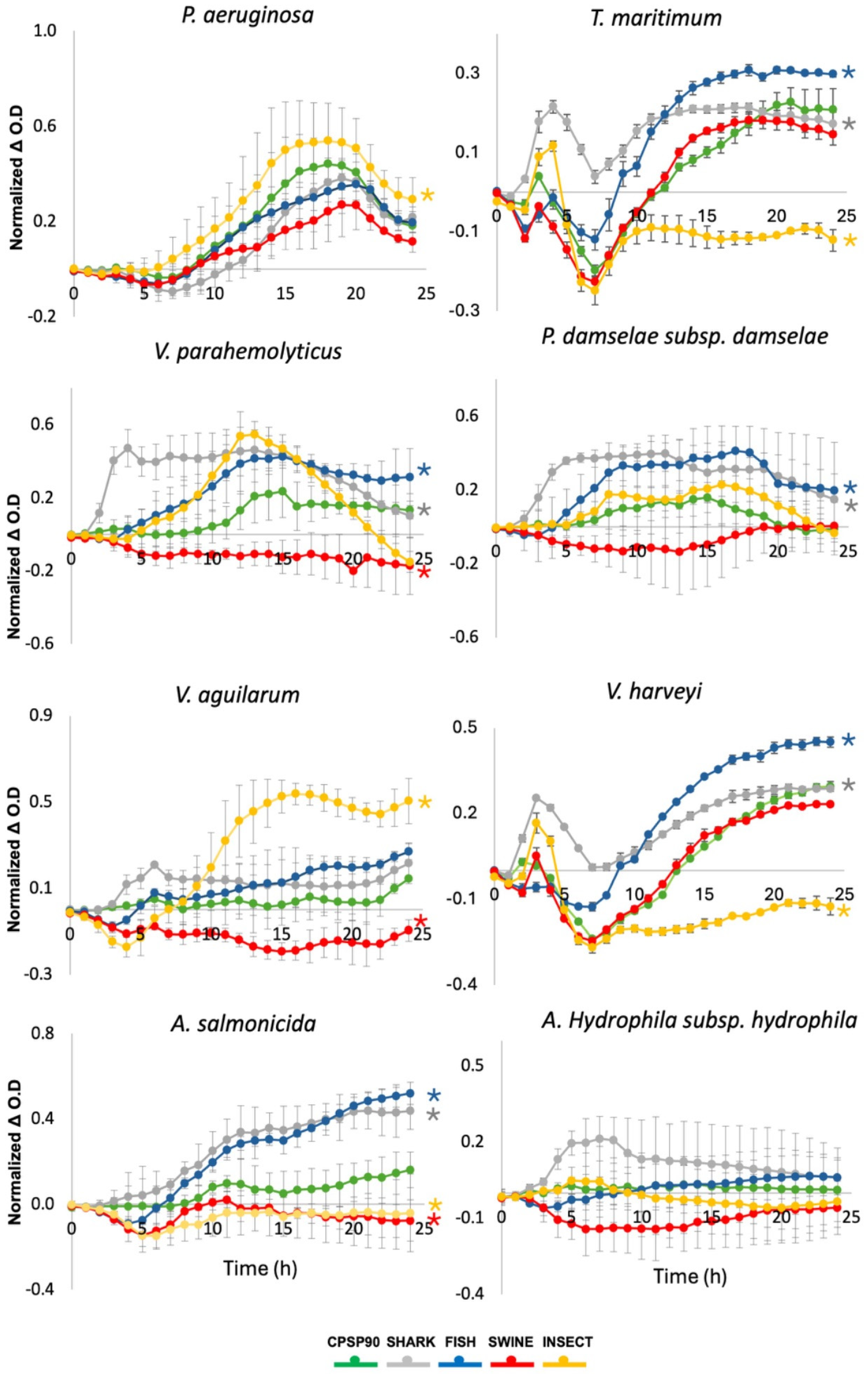
3.5. Bacterial Growth Dynamics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Etemadian, Y.; Ghaemi, V.; Shaviklo, A.R.; Pourashouri, P.; Sadeghi Mahoonak, A.R.; Rafipour, F. Development of animal/plant-based protein hydrolysate and its application in food, feed and nutraceutical industries: State of the art. J. Clean. Prod. 2021, 278, 123219. [Google Scholar] [CrossRef]
- Czelej, M.; Garbacz, K.; Czernecki, T.; Wawrzykowski, J.; Waśko, A. protein hydrolysates derived from animals and plants—A review of production methods and antioxidant activity. Foods 2022, 11, 1953. [Google Scholar] [CrossRef]
- Hou, Y.; Wu, Z.; Dai, Z.; Wang, G.; Wu, G. Protein hydrolysates in animal nutrition: Industrial production, bioactive peptides, and functional significance. J. Anim. Sci. Biotechnol. 2017, 8, 24. [Google Scholar] [CrossRef]
- Rezaee, N.; Hasanvand, P.; Bagheri Lotfabad, T.; Heydarinasab, A.; Khodabandeh, M.; Yaghmaei, S. Study on the use of bovine blood protein hydrolysate as a peptone in microbial culture media. Prep. Biochem. Biotechnol. 2023, 53, 622–633. [Google Scholar] [CrossRef]
- Hayes, M.; Hayes, M. Maximizing use of pelagic capture fisheries for global protein supply: Potential and caveats associated with fish and co-product conversion into value-add ingredients. Glob. Chall. 2023, 7, 2200098. [Google Scholar] [CrossRef] [PubMed]
- García-Garrido, S.; Hachero-Cruzado, I.; Domingues, P.; López, N.; Rosas, C. Effects of fish hydrolysate (CPSP®) on growth and digestive gland lipid composition of Octopus vulgaris (Cuvier, 1797) juveniles. Aquac. Nutr. 2011, 17, e825–e839. [Google Scholar] [CrossRef]
- Barrias, C.; Oliva-Teles, A. The use of locally produced fish meal and other dietary manipulations in practical diets for rainbow trout Oncorhynchus mykiss (walbaum). Aquac. Res. 2000, 31, 213–218. [Google Scholar] [CrossRef]
- Canada, P.; Engrola, S.; Richard, N.; Lopes, A.F.; Pinto, W.; Valente, L.M.P.; Conceição, L.E.C. Dietary indispensable amino acids profile affects protein utilization and growth of Senegalese sole larvae. Fish Physiol. Biochem. 2016, 42, 1493–1508. [Google Scholar] [CrossRef]
- Gisbert, E.; Skalli, A.; Fernández, I.; Kotzamanis, Y.; Zambonino-Infante, J.L.; Fabregat, R. Protein hydrolysates from yeast and pig blood as alternative raw materials in microdiets for gilthead sea bream (Sparus aurata) larvae. Aquaculture 2012, 338–341, 96–104. [Google Scholar] [CrossRef]
- Skalli, A.; Zambonino-Infante, J.L.; Kotzamanis, Y.; Fabregat, R.; Gisbert, E. Peptide molecular weight distribution of soluble protein fraction affects growth performance and quality in European sea bass (Dicentrarchus labrax) larvae. Aquac. Nutr. 2014, 20, 118–131. [Google Scholar] [CrossRef]
- Soudant, P.; Ventura, M.; Chauchat, L.; Guerreiro, M.; Mathieu-Resuge, M.; Le Grand, F.; Simon, V.; Collet, S.; Zambonino-Infante, J.L.; Le Goïc, N.; et al. Evaluation of aurantiochytrium mangrovei biomass grown on digestate as a sustainable feed ingredient of sea bass, Dicentrarchus labrax, juveniles and larvae. Sustainability 2022, 14, 14573. [Google Scholar] [CrossRef]
- Wojno, M.; Mandas, A.; Kwasek, K.; Dabrowski, K. The effect of dietary supplements of black pepper piper nigrum and turmeric Curcuma longa extracts on dietary amino acid utilization and growth performance in common carp. N. Am. J. Aquac. 2021, 83, 155–164. [Google Scholar] [CrossRef]
- Cabral, E.M.; Bacelar, M.; Batista, S.; Castro-Cunha, M.; Ozório, R.O.A.; Valente, L.M.P. Replacement of fishmeal by increasing levels of plant protein blends in diets for Senegalese sole (Solea senegalensis) juveniles. Aquaculture 2011, 322–323, 74–81. [Google Scholar] [CrossRef]
- Hicks, T.M.; Verbeek, C.J.R. Meat industry protein by-products: Sources and characteristics. Protein Byprod. Transform. Environ. Burd. Into Value-Added Prod. 2016, 37–61. [Google Scholar] [CrossRef]
- Livestock and Meat Domestic Data. 2025. Available online: https://www.ers.usda.gov/data-products/livestock-and-meat-domestic-data (accessed on 1 April 2025).
- Cheng, C.; Chen, L.; Zhang, D.; Yu, J.; Zhu, M.; Li, C.; Zheng, X.; Blecker, C.; Li, S. Value-added utilization of hemoglobin and its hydrolysis products from livestock and poultry blood processing by-products: A review. Trends Food Sci. Technol. 2024, 151, 104645. [Google Scholar] [CrossRef]
- Coscueta, E.R.; Cunha Fernandes, N.; Brassesco, M.E.; Rosa, A.; Almeida, A.; Pintado, M.M. Turning discarded blue shark (Prionace glauca) skin into a valuable nutraceutical resource: An enzymatic collagen hydrolysate. Food Biosci. 2024, 60, 104472. [Google Scholar] [CrossRef]
- Prandi, B.; Samaei, S.; Beninati, F.; Nardi, A.; Tedeschi, T.; Sforza, S. Exploitation of bones-rich poultry by-products to produce protein hydrolysates: Optimization of hydrolysis parameters and chemical characterization. Poult. Sci. 2024, 103, 103924. [Google Scholar] [CrossRef]
- Zhang, J.; Ahmmed, M.K.; Regenstein, J.M.; Wu, H. Recent advances of recycling proteins from seafood by-products: Industrial applications, challenges, and breakthroughs. Trends Food Sci. Technol. 2024, 149, 104533. [Google Scholar] [CrossRef]
- Falcon, W.P.; Naylor, R.L.; Shankar, N.D. Rethinking global food demand for 2050. Popul. Dev. Rev. 2022, 48, 921–957. [Google Scholar] [CrossRef]
- Kumar, P.; Abubakar, A.A.; Verma, A.K.; Umaraw, P.; Adewale Ahmed, M.; Mehta, N.; Nizam Hayat, M.; Kaka, U.; Sazili, A.Q. New insights in improving sustainability in meat production: Opportunities and challenges. Crit. Rev. Food Sci. Nutr. 2023, 63, 11830–11858. [Google Scholar] [CrossRef]
- Cain, K. The many challenges of disease management in aquaculture. J. World Aquac. Soc. 2022, 53, 1080. [Google Scholar] [CrossRef]
- Gisbert, E.; Ibarz, A.; Firmino, J.P.; Fernández-Alacid, L.; Salomón, R.; Vallejos-Vidal, E.; Ruiz, A.; Polo, J.; Sanahuja, I.; Reyes-López, F.E.; et al. Porcine protein hydrolysates (PEPTEIVA®) promote growth and enhance systemic immunity in gilthead sea bream (Sparus aurata). Animals 2021, 11, 2122. [Google Scholar] [CrossRef]
- Resende, D.; Costas, B.; Sá, T.; Golfetto, U.; Machado, M.; Pereira, M.; Pereira, C.; Marques, B.; Rocha, C.M.R.; Pintado, M.; et al. Innovative swine blood hydrolysates as promising ingredients for European seabass diets: Impact on growth performance and resistance to Tenacibaculum maritimum infection. Aquaculture 2022, 561, 738657. [Google Scholar] [CrossRef]
- Herault, M.; Gunathilaka, B.E.; Fournier, V.; Le Bris, H.; Lee, K.J.; Sadoul, B. Aquatic product hydrolysates increase rearing performance in red seabream (Pagrus major), fed a low fish meal diet, in both controlled and stressed conditions: From growth to stress responses. Aquaculture 2023, 576, 739830. [Google Scholar] [CrossRef]
- Borges, S.; Piccirillo, C.; Scalera, F.; Martins, R.; Rosa, A.; Couto, J.A.; Almeida, A.; Pintado, M. Valorization of porcine by-products: A combined process for protein hydrolysates and hydroxyapatite production. Bioresour. Bioprocess. 2022, 9, 30. [Google Scholar] [CrossRef] [PubMed]
- Borges, S.; Odila, J.; Voss, G.; Martins, R.; Rosa, A.; Couto, J.A.; Almeida, A.; Pintado, M. Fish by-products: A source of enzymes to generate circular bioactive hydrolysates. Molecules 2023, 28, 1155. [Google Scholar] [CrossRef]
- Borges, S.; Martins, R.; Rosa, A.; Almeida, A.; Pintado, M. Production of insect protein hydrolysates: A multifactorial study. Presented at 1st International Congress on Food, Nutrition and Public Health, Lisbon, Portugal, 17 November 2022. [Google Scholar]
- Marques, A.; Matos, E.; Aires, T.; Melo, D.; Oliveira, M.B.P.P.; Valente, L.M.P. Understanding the interaction between terrestrial animal fat sources and dietary emulsifier supplementation on muscle fatty acid profile and textural properties of European sea bass. Aquaculture 2022, 560, 738547. [Google Scholar] [CrossRef]
- Mæhre, H.K.; Dalheim, L.; Edvinsen, G.K.; Elvevoll, E.O.; Jensen, I.J. Protein determination—method matters. Foods 2018, 7, 5. [Google Scholar] [CrossRef]
- Sriperm, N.; Pesti, G.M.; Tillman, P.B. Evaluation of the fixed nitrogen-to-protein (N:P) conversion factor (6.25) versus ingredient specific N:P conversion factors in feedstuffs. J. Sci. Food Agric. 2011, 91, 1182–1186. [Google Scholar] [CrossRef]
- Guerreiro, I.; Castro, C.; Antunes, B.; Coutinho, F.; Rangel, F.; Couto, A.; Serra, C.R.; Peres, H.; Pousão-Ferreira, P.; Matos, E.; et al. Catching black soldier fly for meagre: Growth, whole-body fatty acid profile and metabolic responses. Aquaculture 2020, 516, 734613. [Google Scholar] [CrossRef]
- Monteiro, M.; Santos, R.A.; Iglesias, P.; Couto, A.; Serra, C.R.; Gouvinhas, I.; Barros, A.; Oliva-Teles, A.; Enes, P.; Díaz-Rosales, P. Effect of extraction method and solvent system on the phenolic content and antioxidant activity of selected macro- and microalgae extracts. J. Appl. Phycol. 2019, 32, 349–362. [Google Scholar] [CrossRef]
- Teodósio, R.; Aragão, C.; Colen, R.; Carrilho, R.; Dias, J.; Engrola, S. A nutritional strategy to promote gilthead seabream performance under low temperatures. Aquaculture 2021, 537, 736494. [Google Scholar] [CrossRef]
- Fernandez Cunha, M.; Coscueta, E.R.; Brassesco, M.E.; Marques, R.; Neto, J.; Almada, F.; Gonçalves, D.; Pintado, M. Exploring Bioactivities and Peptide Content of Body Mucus from the Lusitanian Toadfish Halobatrachus didactylus. Molecules 2023, 28, 6458. [Google Scholar] [CrossRef] [PubMed]
- Dent, T.; LeMinh, A.; Maleky, F. Comparison of Colorimetric Methods for Measuring the Solubility of Legume Proteins. Gels 2024, 10, 551. [Google Scholar] [CrossRef]
- Ribeiro, T.B.; Oliveira, A.; Campos, D.; Nunes, J.; Vicente, A.A.; Pintado, M. Simulated digestion of an olive pomace water-soluble ingredient: Relationship between the bioaccessibility of compounds and their potential health benefits. Food Funct. 2020, 11, 2238–2254. [Google Scholar] [CrossRef]
- Monteiro, M.; Lavrador, A.S.; Santos, R.A.; Rangel, F.; Iglesias, P.; Tárraga, M.; Couto, A.; Serra, C.S.; Tafalla, C.; da Costa, E.; et al. Evaluation of the potential of marine algae extracts as a source of functional ingredients using zebrafish as animal model for aquaculture. Mar. Biotechnol. 2021, 23, 529–545. [Google Scholar] [CrossRef]
- Hao, Y.; Xing, L.; Wang, Z.; Cai, J.; Toldrá, F.; Zhang, W. Study on the anti-inflammatory activity of the porcine bone collagen peptides prepared by ultrasound-assisted enzymatic hydrolysis. Ultrason. Sonochemistry 2023, 101, 106697. [Google Scholar] [CrossRef]
- Prajapati, S.; Koirala, S.; Anal, A.K. Bioutilization of chicken feather waste by newly isolated keratinolytic bacteria and conversion into protein hydrolysates with improved functionalities. Appl. Biochem. Biotechnol. 2021, 193, 2497–2515. [Google Scholar] [CrossRef]
- Siddaiah, G.M.; Kumar, R.; Kumari, R.; Damle, D.K.; Rasal, K.D.; Manohar, V.; Sundaray, J.K.; Pillai, B.R. Dietary supplementation of fish protein hydrolysate improves growth, feed efficiency and immune response in freshwater carnivore fish, Channa striata fingerlings. Aquac. Res. 2022, 53, 3401–3415. [Google Scholar] [CrossRef]
- dos Santos Cardoso, M.; Godoy, A.C.; Oxford, J.H.; Rodrigues, R.; dos Santos Cardoso, M.; Bittencourt, F.; Signor, A.; Boscolo, W.R.; Feiden, A. Apparent digestibility of protein hydrolysates from chicken and swine slaughter residues for Nile tilapia. Aquaculture 2021, 530, 735720. [Google Scholar] [CrossRef]
- Chemat, A.; Song, M.; Li, Y.; Fabiano-Tixier, A.S. Shade of innovative food processing techniques: Potential inducing factors of lipid oxidation. Molecules 2023, 28, 8138. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Qi, L.; Fan, F.; Guo, Z.; Wang, Z.; Song, W.; Du, M. Analysis of volatile compounds and nutritional properties of enzymatic hydrolysate of protein from cod bone. Food Chem. 2018, 264, 350–357. [Google Scholar] [CrossRef]
- Patel, S.; Goyal, A. Chitin and chitinase: Role in pathogenicity, allergenicity and health. Int. J. Biol. Macromol. 2017, 97, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Eggink, K.M.; Dalsgaard, J. Chitin contents in different black soldier fly (Hermetia illucens) life stages. J. Insects Food Feed 2023, 9, 855–863. [Google Scholar] [CrossRef]
- Basto, A.; Marques, A.; Silva, A.; Sá, T.; Sousa, V.; Oliveira, M.B.P.P.; Aires, T.; Valente, L.M.P. Nutritional, organoleptic and sensory quality of market-sized European sea bass (Dicentrarchus labrax) fed defatted Tenebrio molitor larvae meal as main protein source. Aquaculture 2023, 566, 739210. [Google Scholar] [CrossRef]
- Sevgili, H.; Kurtoğlu, A.; Oikawa, M.; Fedekar, D.; Emre, Y.; Takeno, N. Evaluation of nutritional values of selected commercial fish meal sources in turbot (Psetta maxima) diets. Aquac. Res. 2015, 46, 2332–2343. [Google Scholar] [CrossRef]
- Opheim, M.; Sterten, H.; Øverland, M.; Kjos, N.P. Atlantic salmon (Salmo salar) protein hydrolysate—Effect on growth performance and intestinal morphometry in broiler chickens. Livest. Sci. 2016, 187, 138–145. [Google Scholar] [CrossRef]
- Siddik, M.A.B.; Howieson, J.; Fotedar, R.; Partridge, G.J. Enzymatic fish protein hydrolysates in finfish aquaculture: A review. Rev. Aquac. 2021, 13, 406–430. [Google Scholar] [CrossRef]
- Khosravi, S.; Rahimnejad, S.; Herault, M.; Fournier, V.; Lee, C.R.; Dio Bui, H.T.; Jeong, J.B.; Lee, K.J. Effects of protein hydrolysates supplementation in low fish meal diets on growth performance, innate immunity and disease resistance of red sea bream Pagrus major. Fish Shellfish Immunol. 2015, 45, 858–868. [Google Scholar] [CrossRef]
- Egerton, S.; Wan, A.; Murphy, K.; Collins, F.; Ahern, G.; Sugrue, I.; Busca, K.; Egan, F.; Muller, N.; Whooley, J.; et al. Replacing fishmeal with plant protein in Atlantic salmon (Salmo salar) diets by supplementation with fish protein hydrolysate. Sci. Rep. 2020, 10, 4194. [Google Scholar] [CrossRef]
- Li, P.; Wu, G. Roles of dietary glycine, proline, and hydroxyproline in collagen synthesis and animal growth. Amino Acids 2018, 50, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Amiri, A.; Memarpoor-Yazdi, M.; Shanbedi, M.; Eshghi, H. Influence of different amino acid groups on the free radical scavenging capability of multi walled carbon nanotubes. J. Biomed. Mater. Res. Part A 2013, 101A, 2219–2228. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.H.; Karadeniz, F.; Yang, J.; Lee, H.; Choi, M.-N.; Jeon, S.; Park, G.; Kim, J.; Park, K.; Kong, C.-S. Antioxidant, anti-inflammatory, anti-adipogenesis activities and proximate composition of Hermetia illucens larvae reared on food waste enriched with different wastes. J. Anim. Sci. Technol. 2024, 66, 1034. [Google Scholar] [CrossRef] [PubMed]
- Pyo, S.J.; Kang, D.G.; Jung, C.; Sohn, H.Y. Anti-Thrombotic, Anti-Oxidant and Haemolysis Activities of Six Edible Insect Species. Foods 2020, 9, 401. [Google Scholar] [CrossRef]
- Aiello, D.; Barbera, M.; Bongiorno, D.; Cammarata, M.; Censi, V.; Indelicato, S.; Mazzotti, F.; Napoli, A.; Piazzese, D.; Saiano, F. Edible insects an alternative nutritional source of bioactive compounds: A review. Molecules 2023, 28, 699. [Google Scholar] [CrossRef]
- Zou, T.B.; He, T.P.; Li, H.B.; Tang, H.W.; Xia, E.Q. The structure-activity relationship of the antioxidant peptides from natural proteins. Molecules 2016, 21, 72. [Google Scholar] [CrossRef]
- Sun, Q. The Hydrophobic effects: Our current understanding. Molecules 2022, 27, 7009. [Google Scholar] [CrossRef]
- Kristinsson, H.G.; Rasco, B.A. Biochemical and functional properties of Atlantic salmon (Salmo salar) muscle proteins hydrolyzed with various alkaline proteases. J. Agric. Food Chem. 2000, 48, 657–666. [Google Scholar] [CrossRef]
- Sawicki, T.; Jabłońska, M.; Danielewicz, A.; Przybyłowicz, K.E. Phenolic compounds profile and antioxidant capacity of plant-based protein supplements. Molecules 2024, 29, 2101. [Google Scholar] [CrossRef]
- George, J.; Edwards, D.; Pun, S.; Williams, D. Evaluation of antioxidant capacity (abts and cuprac) and total phenolic content (folin-ciocalteu) assays of selected fruit, vegetables, and spices. Int. J. Food Sci. 2022, 2022, 2581470. [Google Scholar] [CrossRef]
- Fu, L.; Xu, B.T.; Xu, X.R.; Qin, X.S.; Gan, R.Y.; Li, H.B. Antioxidant capacities and total phenolic contents of 56 wild fruits from south China. Molecules 2010, 15, 8602–8617. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Rao, J.; Chen, B. Plant protein solubility: A challenge or insurmountable obstacle. Adv. Colloid Interface Sci. 2024, 324, 103074. [Google Scholar] [CrossRef]
- Grossmann, L.; McClements, D.J. Current insights into protein solubility: A review of its importance for alternative proteins. Food Hydrocoll. 2023, 137, 108416. [Google Scholar] [CrossRef]
- Forsyth, W.R.; Antosiewicz, J.M.; Robertson, A.D. Empirical relationships between protein structure and carboxyl pKa values in proteins. Proteins Struct. Funct. Bioinform. 2002, 48, 388–403. [Google Scholar] [CrossRef]
- León-López, A.; Morales-Peñaloza, A.; Martínez-Juárez, V.M.; Vargas-Torres, A.; Zeugolis, D.I.; Aguirre-Álvarez, G. Hydrolyzed collagen—sources and applications. Molecules 2019, 24, 4031. [Google Scholar] [CrossRef] [PubMed]
- Trevino, S.R.; Scholtz, J.M.; Pace, C.N. Amino acid contribution to protein solubility: Asp, glu, and ser contribute more favorably than the other hydrophilic amino acids in rnase sa. J. Mol. Biol. 2007, 366, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Hall, F.G.; Jones, O.G.; O’Haire, M.E.; Liceaga, A.M. Functional properties of tropical banded cricket (Gryllodes sigillatus) protein hydrolysates. Food Chem. 2017, 224, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Barea, P.; Melgosa, R.; Benito-Román, Ó.; Illera, A.E.; Beltrán, S.; Sanz, M.T. Green fractionation and hydrolysis of fish meal to improve their techno-functional properties. Food Chem. 2024, 452, 139550. [Google Scholar] [CrossRef]
- Tonheim, S.K.; Nordgreen, A.; Høgøy, I.; Hamre, K.; Rønnestad, I. In vitro digestibility of water-soluble and water-insoluble protein fractions of some common fish larval feeds and feed ingredients. Aquaculture 2007, 262, 426–435. [Google Scholar] [CrossRef]
- Gruppi, A.; Dermiki, M.; Spigno, G.; Fitzgerald, R.J. Impact of Enzymatic hydrolysis and heat inactivation on the physicochemical properties of milk protein hydrolysates. Foods 2022, 11, 516. [Google Scholar] [CrossRef]
- Shaviklo, G.R.; Thorkelsson, G.; Arason, S.; Kristinsson, H.G.; Sveinsdottir, K. The influence of additives and drying methods on quality attributes of fish protein powder made from saithe (Pollachius virens). J. Sci. Food Agric. 2010, 90, 2133–2143. [Google Scholar] [CrossRef]
- Verma, A.K.; Chatli, M.K.; Mehta, N.; Kumar, P. Efficacy of antioxidant and antimicrobial activity of whole porcine blood hydrolysates and its fractions under in-vitro conditions. Anim. Prod. Sci. 2018, 50, 2084–2090. [Google Scholar] [CrossRef]
- Bah, C.S.F.; Bekhit, A.E.D.A.; Carne, A.; McConnell, M.A. Slaughterhouse blood: An emerging source of bioactive compounds. Compr. Rev. Food Sci. Food Saf. 2013, 12, 314–331. [Google Scholar] [CrossRef]
- Huang, L.; Wu, Y.; Fan, Y.; Su, Y.; Liu, Z.; Bai, J.; Zhao, X.; Li, Y.; Xie, X.; Zhang, J.; et al. The growth-promoting effects of protein hydrolysates and their derived peptides on probiotics: Structure-activity relationships, mechanisms and future perspectives. Crit. Rev. Food Sci. Nutr. 2024, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Petrova, I.; Tolstorebrov, I.; Zhivlyantseva, I.; Eikevik, T.M. Utilization of fish protein hydrolysates as peptones for microbiological culture medias. Food Biosci. 2021, 42, 101063. [Google Scholar] [CrossRef]
- Kuo, I.P.; Liu, C.S.; Yang, S.D.; Liao, Z.H.; Hu, Y.F.; Nan, F.H. Tilapia processing byproduct treated with enzymatic hydrolysis and Rhodopseudomonas palustris is feasible for live feed culturing: Pathogen inhibition in vitro, live feeds microbiota, and Bidyanus bidyanus larviculture. Aquac. Rep. 2023, 28, 101439. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, C.; Wang, J.; Wen, Y.; Li, H.; Liu, X.; Liu, X. Can proteins, protein hydrolysates and peptides cooperate with probiotics to inhibit pathogens? Crit. Rev. Food Sci. Nutr. 2023, 6, 1023–1036. [Google Scholar] [CrossRef]
- Wu, Q.; Lin, H.; Shen, W.; Cao, W.; Qin, X.; Gao, J.; Chen, Z.; Zheng, H.; Zhong, S.; Huang, H. The Preventive Effect of low-molecular weight oyster peptides on lipopolysaccharide-induced acute colitis in mice by modulating intestinal microbiota communities. Foods 2024, 13, 2391. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains | Culture Media |
|---|---|
| Aeromonas hydrophila subsp. hydrophila (LMG 2844) | TSB |
| Aeromonas salmonicida (LMG 3780) | TSB |
| Pseudomonas aeruginosa (LMG 12228) | TSB |
| Photobacterium damselae subsp. damselae (LMG 19445) | TSB |
| Vibrio anguillarum (LMG 4437) | TSB |
| Vibrio parahaemolyticus (LMG 2850) | TSB |
| Vibrio harveyi (LMG 18298) | MB |
| Tenacibaculum maritimum (LMG 13040) | MB |
| CPSP90 | SHARK | FISH | SWINE | INSECT | |
|---|---|---|---|---|---|
| Dry Matter (DM) | 96.2 | 93.6 | 94.0 | 95.6 | 98.0 |
| Crude Protein | 83.7 | 89.5 | 80.5 | 87.6 | 55.1 |
| Crude Fat | 8.4 | 0.34 | 0.1 | 0.2 | 2.0 |
| Energy (kJ g−1 DM) | 19.6 | 19.4 | 20.8 | 20.5 | 19.7 |
| Ash | 7.9 | 5.8 | 6.4 | 5.4 | 11.2 |
| Phosphorus | 0.7 | 0.2 | 0.6 | 0.2 | 0.5 |
| Chitin | - | - | - | - | 0.2 |
| Total Phenolics (gallic acid eq.) | - | - | - | - | 2.7 |
| CPSP90 | SHARK | FISH | SWINE | INSECT | |
|---|---|---|---|---|---|
| Essential amino acids (EAA) | |||||
| Arginine | 6.8 | 7.5 | 5.0 | 6.6 | 4.0 |
| Histidine | 2.2 | 1.3 | 2.1 | 2.1 | 3.4 |
| Lysine | 7.9 | 4.7 | 8.3 | 5.5 | 5.7 |
| Threonine | 4.2 | 2.9 | 4.3 | 2.8 | 3.5 |
| Isoleucine | 3.8 | 2.8 | 4.1 | 2.5 | 4.3 |
| Leucine | 7.2 | 3.9 | 7.6 | 5.6 | 6.8 |
| Valine | 4.6 | 3.0 | 5.1 | 3.9 | 7.2 |
| Methionine | 2.9 | 2.2 | 2.9 | 1.4 | 1.3 |
| Phenylanine | 4.0 | 2.3 | 3.9 | 3.2 | 3.3 |
| Tryptophan | 0.9 | 0.2 | 0.8 | 0.5 | 1.4 |
| Σ EAA | 44.4 | 30.9 | 44.0 | 34.2 | 41.0 |
| Non-essential amino acids (EAA) | |||||
| Cysteine | 1.0 | 0.3 | 0.7 | 0.2 | 0.8 |
| Tyrosine | 2.9 | 0.7 | 1.6 | 1.9 | 4.2 |
| Aspartic acid + asparagine | 9.3 | 6.0 | 9.5 | 6.5 | 8.8 |
| Glutamic acid + glutamine | 13.7 | 11.0 | 15.6 | 13.3 | 14.8 |
| Alanine | 6.9 | 9.0 | 7.8 | 8.4 | 11.1 |
| Glycine | 9.8 | 20.7 | 9.3 | 16.1 | 6.7 |
| Proline | 5.2 | 10.0 | 5.4 | 9.5 | 7.5 |
| Serine | 5.0 | 3.8 | 3.7 | 3.2 | 3.7 |
| Hydroxyproline | 1.7 | 6.9 | 1.7 | 6.2 | 0.4 |
| Ornithine | - | 0.4 | 0.8 | 0.4 | 1.1 |
| Σ NEAA | 55.6 | 69.1 | 56.0 | 65.8 | 59.0 |
| NEAA/EAA | 1.3 | 2.2 | 1.3 | 1.9 | 1.4 |
| Amino acid Classes | |||||
| Positive | 16.9 | 13.9 | 16.2 | 14.6 | 14.3 |
| Hydrophilic | 53.0 | 38.8 | 51.6 | 42.6 | 50.1 |
| Collagenic | 23.6 | 46.7 | 24.1 | 40.2 | 25.7 |
| Aromatic | 10.0 | 4.6 | 8.4 | 7.7 | 12.3 |
| Branched-chain | 15.6 | 9.8 | 16.8 | 12.1 | 18.3 |
| Sulfur | 4.0 | 2.5 | 3.6 | 1.6 | 2.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteiro, M.; Rodrigues-dos-Santos, L.; Filipa-Silva, A.; Marques, D.A.; Pintado, M.; Almeida, A.; Valente, L.M.P. Circular Animal Protein Hydrolysates: A Comparative Approach of Functional Properties. Antioxidants 2025, 14, 782. https://doi.org/10.3390/antiox14070782
Monteiro M, Rodrigues-dos-Santos L, Filipa-Silva A, Marques DA, Pintado M, Almeida A, Valente LMP. Circular Animal Protein Hydrolysates: A Comparative Approach of Functional Properties. Antioxidants. 2025; 14(7):782. https://doi.org/10.3390/antiox14070782
Chicago/Turabian StyleMonteiro, Marta, Luciano Rodrigues-dos-Santos, Andreia Filipa-Silva, Diana A. Marques, Manuela Pintado, André Almeida, and Luisa M. P. Valente. 2025. "Circular Animal Protein Hydrolysates: A Comparative Approach of Functional Properties" Antioxidants 14, no. 7: 782. https://doi.org/10.3390/antiox14070782
APA StyleMonteiro, M., Rodrigues-dos-Santos, L., Filipa-Silva, A., Marques, D. A., Pintado, M., Almeida, A., & Valente, L. M. P. (2025). Circular Animal Protein Hydrolysates: A Comparative Approach of Functional Properties. Antioxidants, 14(7), 782. https://doi.org/10.3390/antiox14070782