
Unveiling the Impact of Peanut Consumption on Telomere Length in Young and Healthy Individuals: Insights from the ARISTOTLE Study: A Randomized Clinical Trial

 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
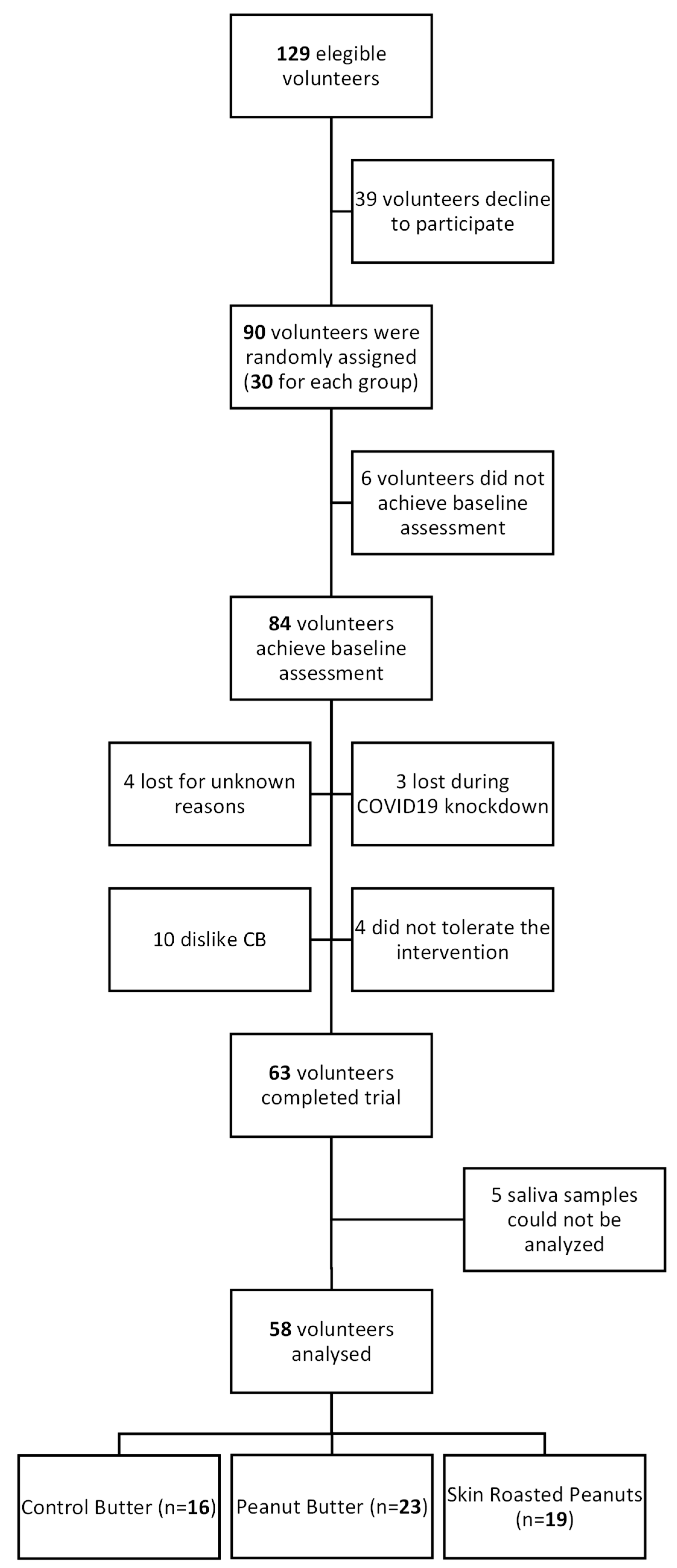
2.1. Study Population and Study Design
2.2. Intervention
2.3. Anthropometric, Biochemical, and Clinical Measurements
2.4. Dietary, Physical Activity, and Sociodemographic Variables
2.5. Telomere Length Assessment
2.6. Statistical Anaysis
3. Results
3.1. Baseline Characteristics of Participants
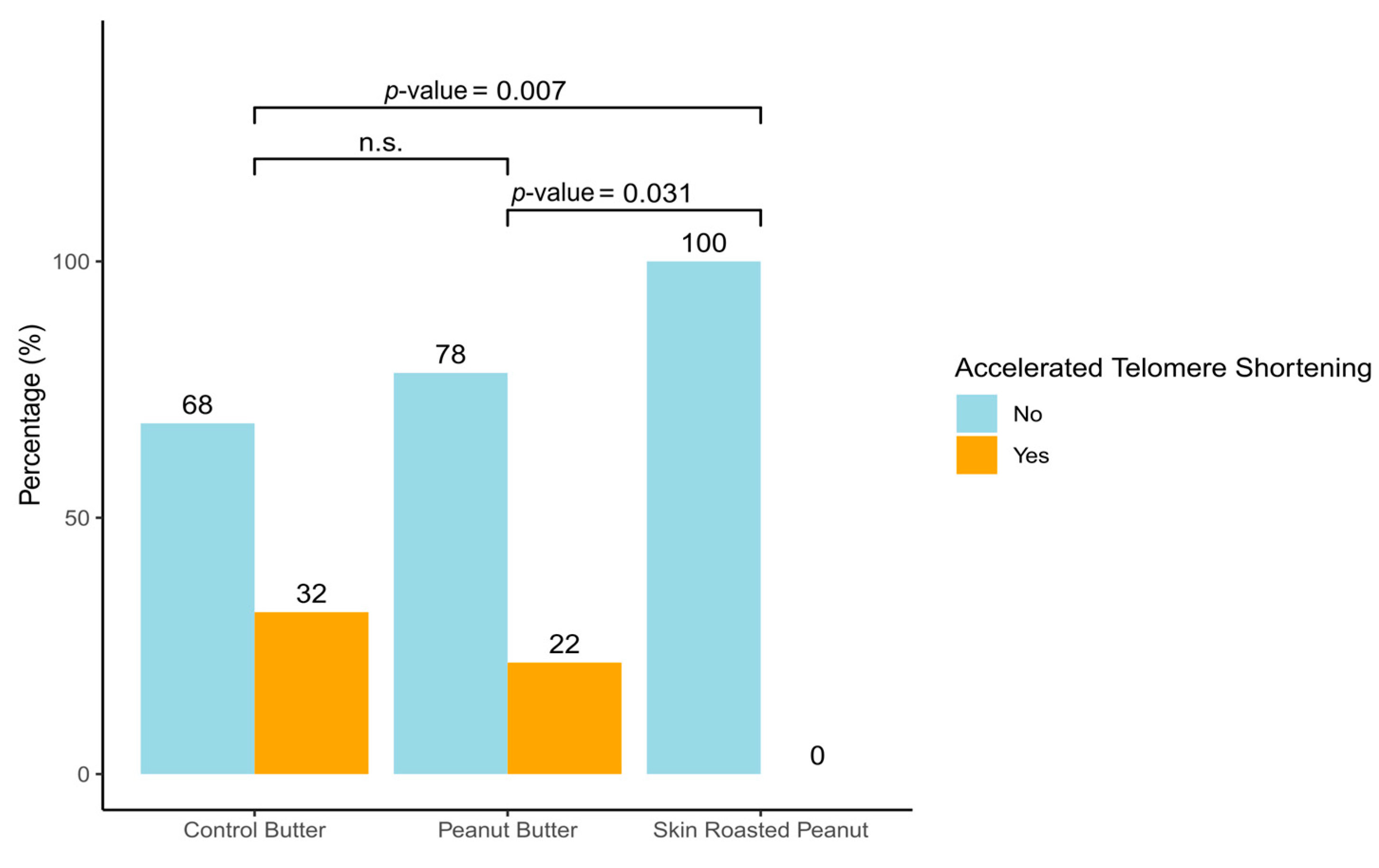
3.2. Effect of the Intervention on Telomere Length
3.3. Relationships Between Variables That Changed with the Intervention and Telomere Length
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Models | SRP vs. CB | p-Value | PB vs. CB | p-Value | |
|---|---|---|---|---|---|
| Mean Difference 6 Moth (95% CI) | Mean Difference 6 Moth (95% CI) | ||||
| Anthropometric measurements | |||||
| Weight (Kg) | Model 1 | −0.01 (−2.44, 2.42) | 0.991 | −0.69 (−2.94, 1.56) | 0.547 |
| Model 2 | 0.09 (−0.49, 0.68) | 0.752 | −0.24 (−0.73, 0.24) | 0.328 | |
| BMI (kg/m2) | Model 1 | −0.03 (−0.86, 0.79) | 0.935 | −0.16 (−0.91, 0.58) | 0.670 |
| Model 2 | −0.03 (−0.86, 0.79) | 0.939 | −0.20 (−0.95, 0.55) | 0.602 | |
| Waist circumference (cm) | Model 1 | −0.12 (−2.03, 1.79) | 0.902 | −0.14 (−2.36, 2.09) | 0.904 |
| Model 2 | −0.16 (−1.35, 1.04) | 0.797 | 0.19 (−1.72, 2.09) | 0.849 | |
| Waist-to-hip ratio | Model 1 | −0.03 (−0.07, 0.01) | 0.076 | −0.03 (−0.06, 0.01) | 0.086 |
| Model 2 | −0.03 (−0.05, −0.01) | 0.027 | −0.02 (−0.04, 0.01) | 0.139 | |
| Body fat (%) | Model 1 | −0.00 (−1.74, 1.74) | 0.999 | −0.34 (−2.10, 1.41) | 0.702 |
| Model 2 | 0.07 (−0.90, 1.05) | 0.880 | −0.11 (−0.99, 0.77) | 0.811 | |
| Visceral fat | Model 1 | −0.13 (−0.96, 0.70) | 0.762 | −0.11 (−0.86, 0.63) | 0.766 |
| Model 2 | −0.07 (−0.44, 0.30) | 0.707 | −0.03 (−0.39, 0.34) | 0.878 | |
| Muscle mass (%) | Model 1 | 0.37 (−0.73, 1.47) | 0.511 | 0.24 (−0.78,1.272) | 0.642 |
| Model 2 | 0.39 (−0.57, 1.33) | 0.436 | 0.15 (−0.60, 0.90) | 0.695 | |
| Blood pressure | |||||
| Systolic blood pressure (mmHg) | Model 1 | 0.74 (−7.86, 9.33) | 0.867 | −3.13 (−10.66, 4.39) | 0.415 |
| Model 2 | 0.80 (−7.19, 8.80) | 0.844 | −3.10 (−10.11, 3.90) | 0.385 | |
| Diastolic blood pressure (mmHg) | Model 1 | 2.60 (−4.61, 9.81) | 0.480 | 0.78 (−5.38, 6.93) | 0.805 |
| Model 2 | 2.64 (−4.20, 9.48) | 0.449 | 1.26 (−4.74, 7.25) | 0.681 | |
| Biochemical parameters | |||||
| Glucose (mmol/L) | Model 1 | 0.07 (−0.12, 0.27) | 0.456 | −0.05 (−0.23, 0.14) | 0.628 |
| Model 2 | 0.08 (−0.11, 0.27) | 0.387 | −0.03 (−0.21, 0.16) | 0.791 | |
| Triglyceride (mmol/L) | Model 1 | 0.08 (−0.13, 0.30) | 0.444 | −0.01 (−0.23, 0.21) | 0.938 |
| Model 2 | 0.09 (−0.13, 0.31) | 0.435 | 0.02 (−0.19, 0.23) | 0.857 | |
| Total cholesterol (mmol/L) | Model 1 | −0.06 (−0.73, 0.60) | 0.848 | −0.15 (−0.84, 0.54) | 0.676 |
| Model 2 | −0.05 (−0.72, 0.62) | 0.883 | −0.13 (−0.83, 0.57) | 0.717 | |
| HDL-cholesterol (mmol/L) | Model 1 | 0.20 (0.01, 0.39) | 0.035 | 0.16 (−0.06, 0.38) | 0.162 |
| Model 2 | 0.20 (0.02, 0.37) | 0.030 | 0.14 (−0.06, 0.34) | 0.168 | |
| LDL-cholesterol (mmol/L) | Model 1 | −0.07 (−0.55, 0.42) | 0.783 | −0.08 (−0.63, 0.47) | 0.771 |
| Model 2 | −0.04 (−0.53, 0.44) | 0.859 | −0.04 (−0.59, 0.50) | 0.877 | |
| Dietary Intake | Models | SRP vs. CB | p-Value | PB vs. CB | p-Value |
|---|---|---|---|---|---|
| Difference Time- Exposure (95% CI) | Difference Time-Exposure (95% CI) | ||||
| Nutrient intake | |||||
| Carbohydrates (g/day) | Model 1 | 0.85 (−25.80, 27.49) | 0.950 | 2.85 (−24.73, 30.43) | 0.840 |
| Model 2 | 9.56 (−4.49, 23.63) | 0.182 | 7.79 (−7.99, 23.58) | 0.334 | |
| Sugars (g/day) | Model 1 | 7.95 (−6.13, 22.04) | 0.269 | 5.45 (−8.01, 18.92) | 0.427 |
| Model 2 | 11.59 (0.04, 23.15) | 0.049 | 6.42 (−3.66, 16.50) | 0.212 | |
| Fiber (g/day) | Model 1 | 3.38 (−1.17, 7.94) | 0.146 | 2.78 (−2.13, 7.70) | 0.267 |
| Model 2 | 4.88 (1.77, 7.99) | 0.002 | 3.65 (0.45, 6.85) | 0.025 | |
| Protein (g/day) | Model 1 | −4.57 (−13.69, 4.54) | 0.325 | −5.25 (−14.96, 4.47) | 0.290 |
| Model 2 | −0.78 (−7.94, 6.39) | 0.832 | −3.56 (−10.47, 3.35) | 0.313 | |
| Total fat (g/day) | Model 1 | 11.12 (−1.24, 23.48) | 0.078 | −5.97 (−19.18, 7.23) | 0.375 |
| Model 2 | 6.27 (−0.10, 12.63) | 0.054 | −3.13 (−10.21, 3.95) | 0.386 | |
| Saturated fat (g/day) | Model 1 | −0.87 (−4.86, 3.12) | 0.670 | −1.50 (−5.66, 2.67) | 0.481 |
| Model 2 | 0.60 (−1.77, 2.96) | 0.622 | −0.71 (−2.85, 1.43) | 0.515 | |
| Monounsaturated fat (g/day) | Model 1 | 7.41 (0.79, 14.02) | 0.028 | −5.41 (−12.36, 1.54) | 0.127 |
| Model 2 | 5.29 (0.12, 10.71) | 0.056 | −4.37 8 (10.71, 0.12) | 0.139 | |
| Polyunsaturated fat (g/day) | Model 1 | 0.60 (−3.11, 4.31) | 0.752 | 1.59 (−2.22, 5.40) | 0.413 |
| Model 2 | 1.34 (−1.31, 4.00) | 0.322 | 2.28 (−0.57, 5.13) | 0.117 | |
| Energy, kcal/d | Model 1 | −92.80 (−299, 114) | 0.379 | −52.09 (−269, 165) | 0.638 |
| Model 2 | |||||
| Food consumption | |||||
| Fruits (g/day) | Model 1 | 26.23 (−48.62, 101) | 0.492 | −19.05 (−67.13, 29.03) | 0.437 |
| Model 2 | 34.04 (−37.49, 106) | 0.351 | −21.80 (−66.30, 22.70) | 0.337 | |
| Vegetables (g/day) | Model 1 | −20.07 (−50.37, 10.21) | 0.194 | −9.79 (−44.11, 24.54) | 0.576 |
| Model 2 | −14.83 (−45.01, 15.36) | 0.336 | −8.79 (−42.36, 24.78) | 0.608 | |
| Cereals (g/day) | Model 1 | −20.40 (−57.86, 17.05) | 0.286 | −22.72 (−60.76) | 0.242 |
| Model 2 | −12.90 (−43.74, 17.95) | 0.412 | −15.70 (−47.94, 17.95) | 0.412 | |
| Potatoes (g/day) | Model 1 | −3.25 (−27.33, 20.82) | 0.791 | 10.30 (−12.33, 33.53) | 0.385 |
| Model 2 | −2.43 (−26.79, 21.93) | 0.845 | 12.28 (−12.24, 36.80) | 0.326 | |
| Legumes (g/day) | Model 1 | 4.28 (−8.27, 16.83) | 0.504 | −0.77 (−11.87, 10.34) | 0.892 |
| Model 2 | 5.02 (−7.10, 17.16) | 0.417 | −0.48 (−11.50, 10.54) | 0.932 | |
| Dairy products (g/day) | Model 1 | 45.74 (−38.52, 130) | 0.287 | 25.21 (−63.53, 113) | 0.578 |
| Model 2 | 51.76 (−30.19, 134) | 0.216 | 31.07 (−57.34, 119) | 0.491 | |
| Eggs (g/day) | Model 1 | 7.68 (−4.45, 19.81) | 0.214 | −12.11 (−38.11, 13.89) | 0.361 |
| Model 2 | 10.09 (−2.60, 22.77) | 0.119 | −11.25 (−35.84, 13.35) | 0.370 | |
| Fish and sea food (g/day) | Model 1 | −11.02 (−24.52, 2.48) | 0.109 | −0.76 (−12.30, 10.78) | 0.897 |
| Model 2 | −7.89 (−22.90, 7.12) | 0.303 | 0.21 (13.41, 13.83) | 0.976 | |
| Meat/meat products (g/day) | Model 1 | −0.39 (−20.89, 20.11) | 0.970 | −18.35 (−39.39, 2.68) | 0.087 |
| Model 2 | 3.05 (−18.52, 24.62) | 0.782 | −18.49 (−38.76, 1.78) | 0.074 | |
| Oils and butter (g/day) | Model 1 | 8.42 (−7.76, 24.59) | 0.308 | 1.23 (−10.36, 12.83) | 0.834 |
| Model 2 | 10.28 (−6.51, 27.06) | 0.230 | 2.34 (−8.51, 13.21) | 0.672 | |
| EVOO (g/day) | Model 1 | 0.82 (−6.99, 8.62) | 0.838 | −6.15 (−13.16, 0.86) | 0.086 |
| Model 2 | 1.23 (−6.97, 9.43) | 0.769 | −6.08 (−13.49, 1.33) | 0.108 | |
| Pastries (g/day) | Model 1 | 0.32 (−15.83, 16.47) | 0.969 | 3.14 (−12.53, 18.81) | 0.694 |
| Model 2 | 3.28 (−10.53, 17.10) | 0.641 | 5.51 (−6.15, 17.17) | 0.354 | |
| Micronutrients | |||||
| B-group vitamins | Model 1 | 10.06 (−41.38, 61.49) | 0.702 | 2.39 (−51.75, 56.52) | 0.931 |
| Model 2 | 26.51 (−15.94, 68.97) | 0.221 | 11.62 (−31.53, 54.76) | 0.598 | |
| Vitamin A | Model 1 | 67.44 (−195, 330) | 0.614 | −28.12 (−399, 344) | 0.882 |
| Model 2 | 129 (−129, 386) | 0.327 | −9.07 (−369, 351) | 0.961 | |
| Vitamin D | Model 1 | 0.25 (−0.59, 1.09) | 0.554 | 0.31 (−0.66, 1.28) | 0.533 |
| Model 2 | 0.50 (−0.49, 1.49) | 0.329 | 0.35 (−0.66, 1.37) | 0.499 | |
| Vitamin E | Model 1 | 3.38 (0.96, 5.80) | 0.006 | 3.97 (1.10, 6.85) | 0.007 |
| Model 2 | 2.76 (0.85, 4.67) | 0.005 | 3.56 (1.25, 5.84) | 0.002 | |
| Vitamin C | Model 1 | −16.78 (−45.88, 12.36) | 0.259 | −10.17 (−40.49, 20.15) | 0.511 |
| Model 2 | −12.33 (−42.59, 17.93) | 0.425 | −11.47 (−42.59, 19.65) | 0.470 | |
| Sodium | Model 1 | −45.05 (−395, 305) | 0.801 | −135 (−475, 203) | 0.432 |
| Model 2 | 59.28 (−257, 375) | 0.713 | −88 (−353, 176) | 0.512 | |
| Magnesium | Model 1 | −6.43 (−64.39, 51.53) | 0.828 | −15.74 (−74.86, 43.38) | 0.602 |
| Model 2 | 14.59 (−15.89, 45.07) | 0.348 | −1.38 (−31.28, 28.52) | 0.928 | |
| Potassium | Model 1 | −15.41 (−192, 162) | 0.865 | −68.76 (−243, 106) | 0.440 |
| Model 2 | 51.18 (−68.50, 171) | 0.402 | −30.80 (−147, 85.22) | 0.603 | |
| Iron | Model 1 | −0.02 (−1.56, 1.52) | 0.981 | −0.53 (−2.25, 1.19) | 0.546 |
| Model 2 | 0.61 (−0.39, 1.60) | 0.230 | −0.16 (−1.14, 0.82) | 0.748 | |
| Zinc | Model 1 | 0.05 (−1.09, 1.19) | 0.933 | −1.02 (−2.36, 0.32) | 0.136 |
| Model 2 | 0.54 (−0.22, 1.29) | 0.165 | −0.75 (−1.51, 0.01) | 0.053 | |
| Polyphenols | |||||
| Flavanols | Model 1 | 89.34 (−218, 397) | 0.570 | −122 (−493, 250) | 0.521 |
| Model 2 | 113 (−182, 409) | 0.451 | −113 (−460, 234) | 0.523 | |
| Flavonols | Model 1 | −3.94 (−11.54, 3.66) | 0.310 | 0.50 (−5.37, 6.37) | 0.868 |
| Model 2 | −2.57 (−9.25, 4.10) | 0.450 | 1.57 (−3.55, 6.68) | 0.548 | |
| Flavanones | Model 1 | −12.73 (−34.81, 9.35) | 0.258 | −3.34 (−19.88, 13.21) | 0.692 |
| Model 2 | −12.27 (−20.90, 13.39) | 0.274 | −3.76 (−20.90, 13.39) | 0.667 | |
| Flavones | Model 1 | −23.68 (−46.04, −1.33) | 0.038 | −24.84 (−48.34, −1.34) | 0.038 |
| Model 2 | −18.72 (−41.08, 3.65) | 0.101 | −20.41 (−41.66, 0.83) | 0.060 | |
| Isoflavonoids | Model 1 | −2.74 (−8.65, 3.16) | 0.362 | −2.49 (−5.56, 0.58) | 0.112 |
| Model 2 | −2.49 (−8.03, 3.05) | 0.378 | −2.11 (−4.83, 0.62) | 0.130 | |
| Anthocyanins | Model 1 | −6.48 (−12.46, −0.50) | 0.034 | −2.18 (−7.58, 3.22) | 0.429 |
| Model 2 | −5.71 (−11.43, 0.01) | 0.051 | −2.01 (−7.43, 3.42) | 0.468 | |
| Phenolic acids | Model 1 | 18.75 (−91.99, 129) | 0.740 | 18.86 (−100, 138) | 0.757 |
| Model 2 | 42.41 (−57.45, 142) | 0.405 | 30.13 (−67.03, 127) | 0.543 | |
| Stilbenes | Model 1 | 0.09 (−0.01, 0.28) | 0.351 | 0.10 (−0.07, 0.21) | 0.265 |
| Model 2 | −0.08 (−0.27, 0.11) | 0.424 | −0.09 (−0.27, 0.08) | 0.272 | |
| Lignans | Model 1 | −0.19 (−2.90, 2.53) | 0.892 | 0.23 (−2.96, 3.43) | 0.885 |
| Model 2 | −0.10 (−2.69, 2.49) | 0.938 | 0.45 (−2.60, 3.50) | 0.771 | |
| Present in peanuts | |||||
| m-coumaric acid | Model 1 | 0.40 (0.18, 0.63) | <0.001 | 0.41 (0.25, 0.57) | <0.001 |
| Model 2 | 0.43 (0.21, 0.66) | <0.001 | 0.41 (0.25, 0.57) | <0.001 | |
| o-coumaric acid | Model 1 | 1.43 (0.81, 2.04) | <0.001 | 0.04 (−0.05, 0.14) | 0.376 |
| Model 2 | 1.45 (0.85, 2.04) | <0.001 | 0.03 (−0.08, 0.13) | 0.631 | |
| p-coumaric acid | Model 1 | 6.15 (5.95, 6.36) | <0.001 | 12.99 (12.42, 13.56) | <0.001 |
| Model 2 | 6.19 (6.02, 6.37) | <0.001 | 13.01 (12.43, 13.59) | <0.001 | |
| Resveratrol | Model 1 | 0.07 (0.05,0.08) | <0.001 | 0.08 (0.06, 0.09) | <0.001 |
| Model 2 | 0.07 (0.05, 0.09) | <0.001 | 0.08 (0.07, 0.09) | <0.001 | |
| n | Telomere Length | |||
|---|---|---|---|---|
| B (CI) | p-Value | |||
| Waist-to-hip ratio | Model 1 | 58 | −0.11 (−0.73;0.52) | 0.731 |
| Model 2 | 58 | −0.08 (−0.66, 0.50) | 0.791 | |
| HDL-cholesterol | Model 1 | 57 | 0.22 (−0.82; 1.32) | 0.643 |
| Model 2 | 57 | 0.03 (−0.94, 1.00) | 0.947 | |
| Carbohydrates intake | Model 1 | 58 | 0.08 (−0.66, 0.83) | 0.822 |
| Model 2 | 58 | 0.15 (−0.55, 0.85) | 0.660 | |
| Fiber intake | Model 1 | 58 | −0.01 (−0.78, 0.77) | 0.990 |
| Model 2 | 58 | 0.07 (−0.66,0.81) | 0.838 | |
| MUFA intake | Model 1 | 58 | 0.58 (0.11, 1.05) | 0.016 |
| Model 2 | 58 | 0.46 (0.02, 0.93) | 0.048 | |
| Vitamin E intake | Model 1 | 58 | −0.15 (−0.82, 0.52) | 0.663 |
| Model 2 | 58 | −0.09 (−0.65, 0.48) | 0.760 | |
| m-Coumaric acid | Model 1 | 58 | 0.24 (−0.31, 0.79) | 0.627 |
| Model 2 | 58 | 0.23 (−0.29, 0.75) | 0.630 | |
| o-Coumaric acid | Model 1 | 58 | −0.25 (−1.27, 0.76) | 0.619 |
| Model 2 | 58 | −0.27 (−1.21, 0.67) | 0.571 | |
| p-Coumaric acid | Model 1 | 58 | −0.06 (−1.90, 1.78) | 0.949 |
| Model 2 | 58 | 0.06 (−1.54, 1.67) | 0.936 | |
| Resveratrol | Model 1 | 58 | 1.20 (0.01, 2.39) | 0.048 |
| Model 2 | 58 | 1.52 (0.37, 2.67) | 0.011 | |
| Flavones | Model 1 | 58 | −0.07 (−0.80, 0.65) | 0.838 |
| Model 2 | 58 | −0.03 (−0.72, 0.66) | 0.929 | |
| Anthocyanins | Model 1 | 58 | 0.79 (0.02, 1.56) | 0.044 |
| Model 2 | 58 | 0.81 (−0.02, 1.66) | 0.057 | |
References
- Ros, E.; Hu, F.B. Consumption of Plant Seeds and Cardiovascular Health: Epidemiological and Clinical Trial Evidence. Circulation 2013, 128, 553–565. [Google Scholar] [CrossRef]
- Toomer, O.T. Nutritional Chemistry of the Peanut (Arachis Hypogaea). Crit. Rev. Food Sci. Nutr. 2018, 58, 3042–3053. [Google Scholar] [CrossRef]
- Gebicki, J.M.; Nauser, T. Fast Antioxidant Reaction of Polyphenols and Their Metabolites. Antioxidants 2021, 10, 1297. [Google Scholar] [CrossRef]
- Becerra-Tomás, N.; Paz-Graniel, I.; Kendall, C.; Kahleova, H.; Rahelić, D.; Sievenpiper, J.L.; Salas-Salvadó, J. Nut Consumption and Incidence of Cardiovascular Diseases and Cardiovascular Disease Mortality: A Meta-Analysis of Prospective Cohort Studies. Nutr. Rev. 2019, 77, 691–709. [Google Scholar] [CrossRef]
- Becerra-Tomás, N.; Paz-Graniel, I.; Hernández-Alonso, P.; Jenkins, D.J.A.; Kendall, C.W.C.; Sievenpiper, J.L.; Salas-Salvadó, J. Nut Consumption and Type 2 Diabetes Risk: A Systematic Review and Meta-Analysis of Observational Studies. Am. J. Clin. Nutr. 2021, 113, 960–971. [Google Scholar] [CrossRef]
- Ros, E.; Singh, A.; O’keefe, J.H. Nuts: Natural Pleiotropic Nutraceuticals. Nutrients 2021, 13, 3269. [Google Scholar] [CrossRef]
- Rusu, M.E.; Mocan, A.; Ferreira, I.C.F.R.; Popa, D.S. Health Benefits of Nut Consumption in Middle-Aged and Elderly Population. Antioxidants 2019, 8, 302. [Google Scholar] [CrossRef]
- Haycock, P.C.; Heydon, E.E.; Kaptoge, S.; Butterworth, A.S.; Thompson, A.; Willeit, P. Leucocyte Telomere Length and Risk of Cardiovascular Disease: Systematic Review and Meta-Analysis. BMJ 2014, 349, g4227. [Google Scholar] [CrossRef]
- D’Mello, M.J.J.; Ross, S.A.; Briel, M.; Anand, S.S.; Gerstein, H.; Paré, G. Association between Shortened Leukocyte Telomere Length and Cardiometabolic Outcomes: Systematic Review and Meta-Analysis. Circ. Cardiovasc. Genet. 2015, 8, 82–90. [Google Scholar] [CrossRef]
- Parilli-Moser, I.; Domínguez-López, I.; Vallverdú-Queralt, A.; Hurtado-Barroso, S.; Lamuela-Raventós, R.M. Urinary Phenolic Metabolites Associated with Peanut Consumption May Have a Beneficial Impact on Vascular Health Biomarkers. Antioxidants 2023, 12, 698. [Google Scholar] [CrossRef]
- Parilli-Moser, I.; Domínguez-López, I.; Trius-Soler, M.; Castellví, M.; Bosch, B.; Castro-Barquero, S.; Estruch, R.; Hurtado-Barroso, S.; Lamuela-Raventós, R.M. Consumption of Peanut Products Improves Memory and Stress Response in Healthy Adults from the ARISTOTLE Study: A 6-Month Randomized Controlled Trial. Clin. Nutr. 2021, 40, 5556–5567. [Google Scholar] [CrossRef]
- Blackburn, E.H. Structure and Function of Telomeres. Nature 1991, 350, 569–573. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of Aging: An Expanding Universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef]
- Frenck, R.W.; Blackburn, E.H.; Shannon, K.M. The Rate of Telomere Sequence Loss in Human Leukocytes Varies with Age. Proc. Natl. Acad. Sci. USA 1998, 95, 5607–5610. [Google Scholar] [CrossRef]
- Aviv, A.; Chen, W.; Gardner, J.P.; Kimura, M.; Brimacombe, M.; Cao, X.; Srinivasan, S.R.; Berenson, G.S. Leukocyte Telomere Dynamics: Longitudinal Findings among Young Adults in the Bogalusa Heart Study. Am. J. Epidemiol. 2009, 169, 323–329. [Google Scholar] [CrossRef]
- Shalev, I.; Entringer, S.; Wadhwa, P.D.; Wolkowitz, O.M.; Puterman, E.; Lin, J.; Epel, E.S. Stress and Telomere Biology: A Lifespan Perspective. Psychoneuroendocrinology 2013, 38, 1835–1842. [Google Scholar] [CrossRef]
- Gorenjak, V.; Petrelis, A.M.; Stathopoulou, M.G.; Visvikis-Siest, S. Telomere Length Determinants in Childhood. Clin. Chem. Lab. Med. 2020, 58, 162–177. [Google Scholar] [CrossRef]
- Okuda, K.; Bardeguez, A.; Gardner, J.P.; Rodriguez, P.; Ganesh, V.; Kimura, M.; Skurnick, J.; Awad, G.; Aviv, A. Telomere Length in the Newborn. Pediatr. Res. 2002, 52, 377–381. [Google Scholar] [CrossRef]
- Calado, R.T.; Dumitriu, B. Telomere Dynamics in Mice and Humans. Semin. Hematol. 2013, 50, 165–174. [Google Scholar] [CrossRef]
- Vidaček, N.Š.; Nanić, L.; Ravlić, S.; Sopta, M.; Gerić, M.; Gajski, G.; Garaj-Vrhovac, V.; Rubelj, I. Telomeres, Nutrition, and Longevity: Can We Really Navigate Our Aging? J. Gerontol.—Ser. A Biol. Sci. Med. Sci. 2018, 73, 39–47. [Google Scholar] [CrossRef]
- Galiè, S.; Canudas, S.; Muralidharan, J.; García-Gavilán, J.; Bulló, M.; Salas-Salvadó, J. Impact of Nutrition on Telomere Health: Systematic Review of Observational Cohort Studies and Randomized Clinical Trials. Adv. Nutr. 2020, 11, 576–601. [Google Scholar] [CrossRef]
- Factor-Litvak, P.; Susser, E. The Importance of Early Life Studies of Telomere Attrition. Paediatr. Perinat. Epidemiol. 2015, 29, 144–145. [Google Scholar] [CrossRef]
- Blackburn, E.H.; Epel, E.S.; Lin, J. Human Telomere Biology: A Contributory and Interactive Factor in Aging, Disease Risks, and Protection. Science 2015, 350, 1193–1198. [Google Scholar] [CrossRef]
- Tucker, L.A. Dietary Fiber and Telomere Length in 5674 U.S. Adults: An NHANES Study of Biological Aging. Nutrients 2018, 10, 400. [Google Scholar] [CrossRef]
- Juton, C.; Castro-barquero, S.; Casas, R.; Freitas, T.; Ruiz-león, A.M.; Crovetto, F.; Domenech, M.; Crispi, F.; Vieta, E.; Gratacós, E.; et al. Reliability and Concurrent and Construct Validity of a Food Frequency Questionnaire for Pregnant Women at High Risk to Develop Fetal Growth Restriction. Nutrients 2021, 13, 1629. [Google Scholar] [CrossRef]
- Elosua, R.; Marrugat, J.; Molina, L.; Pons, S.; Pujol, E. Validation of the Minnesota Leisure Time Physical Activity Questionnaire in Spanish Men. Am. J. Epidemiol. 1994, 139, 1197–1209. [Google Scholar] [CrossRef]
- Elosua, R.; Garcia, M.; Aguilar, A.; Molina, L. Validation of the Minnesota Leisure Time Physical Activity Questionnaire in Spanish Women. Med. Sci. Sports Exerc. 2000, 32, 1431–1437. [Google Scholar] [CrossRef]
- Lin, J.; Smith, D.L.; Esteves, K.; Drury, S. Telomere Length Measurement by QPCR—Summary of Critical Factors and Recommendations for Assay Design. Psychoneuroendocrinology 2019, 99, 271–278. [Google Scholar] [CrossRef]
- Stout, S.A.; Lin, J.; Hernandez, N.; Davis, E.P.; Blackburn, E.; Carroll, J.E.; Glynn, L.M. Validation of Minimally-Invasive Sample Collection Methods for Measurement of Telomere Length. Front. Aging Neurosci. 2017, 9, 397. [Google Scholar] [CrossRef]
- Goldman, E.A.; Eick, G.N.; Compton, D.; Kowal, P.; Snodgrass, J.J.; Eisenberg, D.T.A.; Sterner, K.N. Evaluating Minimally Invasive Sample Collection Methods for Telomere Length Measurement. Am. J. Hum. Biol. 2018, 30, e23062. [Google Scholar] [CrossRef]
- Mitchell, C.; Hobcraft, J.; McLanahan, S.S.; Siegeld, S.R.; Berg, A.; Brooks-Gunn, J.; Garfinkel, I.; Notterman, D. Social Disadvantage, Genetic Sensitivity, and Children’s Telomere Length. Proc. Natl. Acad. Sci. USA 2014, 111, 5944–5949. [Google Scholar] [CrossRef]
- Cawthon, R.M. Telomere Length Measurement by a Novel Monochrome Multiplex Quantitative PCR Method. Nucleic Acids Res. 2009, 37, e21. [Google Scholar] [CrossRef]
- O’Callaghan, N.J.; Fenech, M. A Quantitative PCR Method for Measuring Absolute Telomere Length. Biol. Proced. 2011, 13, 3. [Google Scholar] [CrossRef]
- Blom, G. Statistical Estimates and Transformed Beta Variables. Inc. Stat. 1960, 10, 53. [Google Scholar] [CrossRef]
- Saretzki, G. Telomeres, Telomerase and Ageing. Biochem. Cell Biol. Ageing Part I Biomed. Sci. 2018, 90, 221–308. [Google Scholar]
- Crous-Bou, M.; Molinuevo, J.L.; Sala-Vila, A. Plant-Rich Dietary Patterns, Plant Foods and Nutrients, and Telomere Length. Adv. Nutr. 2019, 10, S296–S303. [Google Scholar] [CrossRef]
- Canudas, S.; Becerra-Tomas, N.; Hernandez-Alonso, P.; Galie, S.; Leung, C.; Crous-Bou, M.; De Vivo, I.; Gao, Y.; Gu, Y.; Meinila, J.; et al. Mediterranean Diet and Telomere Length: A Systematic Review and Meta-Analysis. Adv. Nutr. 2020, 11, 1544–1554. [Google Scholar] [CrossRef]
- Tucker, L.A. Consumption of Nuts and Seeds and Telomere Length in 5,582 Men and Women of the National Health and Nutrition Examination Survey (NHANES). J. Nutr. Health Aging 2017, 21, 233–240. [Google Scholar] [CrossRef]
- Freitas-Simoes, T.M.; Cofán, M.; Blasco, M.A.; Soberón, N.; Foronda, M.; Serra-Mir, M.; Roth, I.; Valls-Pedret, C.; Doménech, M.; Ponferrada-Ariza, E.; et al. Walnut Consumption for Two Years and Leukocyte Telomere Attrition in Mediterranean Elders: Results of a Randomized Controlled Trial. Nutrients 2018, 10, 1907. [Google Scholar] [CrossRef]
- Canudas, S.; Hernández-Alonso, P.; Galié, S.; Muralidharan, J.; Morell-Azanza, L.; Zalba, G.; García-Gavilán, J.; Martí, A.; Salas-Salvadó, J.; Bulló, M. Pistachio Consumption Modulates DNA Oxidation and Genes Related to telomere Maintenance: A Crossover Randomized Clinical Trial. Am. J. Clin. Nutr. 2019, 109, 1738–1745. [Google Scholar] [CrossRef]
- Fernández de la Puente, M.; Hernández-Alonso, P.; Canudas, S.; Marti, A.; Fitó, M.; Razquin, C.; Salas-Salvadó, J. Modulation of Telomere Length by Mediterranean Diet, Caloric Restriction, and Exercise: Results from PREDIMED-Plus Study. Antioxidants 2021, 10, 1596. [Google Scholar] [CrossRef]
- García-Calzón, S.; Martínez-González, M.A.; Razquin, C.; Arós, F.; Lapetra, J.; Martínez, J.A.; Zalba, G.; Marti, A. Mediterranean Diet and Telomere Length in High Cardiovascular Risk Subjects from the PREDIMED-NAVARRA Study. Clin. Nutr. 2016, 35, 1399–1405. [Google Scholar] [CrossRef]
- Ros, E. Health Benefits of Nut Consumption. Nutrients 2010, 2, 652–682. [Google Scholar] [CrossRef]
- Cronin, P.; Joyce, S.A.; O’toole, P.W.; O’connor, E.M. Dietary Fibre Modulates the Gut Microbiota. Nutrients 2021, 13, 1655. [Google Scholar] [CrossRef]
- Ridout, K.K.; Ridout, S.J.; Price, L.H.; Sen, S.; Tyrka, A.R. Depression and Telomere Length: A Meta-Analysis. J. Affect. Disord. 2016, 191, 237–247. [Google Scholar] [CrossRef]
- González-Becerra, K.; Ramos-Lopez, O.; Barrón-Cabrera, E.; Riezu-Boj, J.I.; Milagro, F.I.; Martínez-López, E.; Martínez, J.A. Fatty Acids, Epigenetic Mechanisms and Chronic Diseases: A Systematic Review. Lipids Health Dis. 2019, 18, 178. [Google Scholar] [CrossRef]
- Alam, M.A.; Subhan, N.; Hossain, H.; Hossain, M.; Reza, H.M.; Rahman, M.M.; Ullah, M.O. Hydroxycinnamic Acid Derivatives: A Potential Class of Natural Compounds for the Management of Lipid Metabolism and Obesity. Nutr. Metab. 2016, 13, 27. [Google Scholar] [CrossRef]
- Crous-Bou, M.; Fung, T.T.; Prescott, J.; Julin, B.; Du, M.; Sun, Q.; Rexrode, K.M.; Hu, F.B.; De Vivo, I. Mediterranean Diet and Telomere Length in Nurses’ Health Study: Population Based Cohort Study. BMJ 2014, 349, g6674. [Google Scholar] [CrossRef]
- Song, Y.; You, N.C.Y.; Song, Y.; Kang, M.K.; Hou, L.; Wallace, R.; Eaton, C.B.; Tinker, L.F.; Liu, S. Intake of Small-to-Medium-Chain Saturated Fatty Acids Is Associated with Peripheral Leukocyte Telomere Length in Postmenopausal Women1-3. J. Nutr. 2013, 143, 907–914. [Google Scholar] [CrossRef]
- Tiainen, A.M.; Männistö, S.; Blomstedt, P.A.; Moltchanova, E.; Perälä, M.M.; Kaartinen, N.E.; Kajantie, E.; Kananen, L.; Hovatta, I.; Eriksson, J.G. Leukocyte Telomere Length and Its Relation to Food and Nutrient Intake in an Elderly Population. Eur. J. Clin. Nutr. 2012, 66, 1290–1294. [Google Scholar] [CrossRef]
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; Di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative Stress in Obesity: A Critical Component in Human Diseases. Int. J. Mol. Sci. 2014, 16, 378–400. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.L.; Goldfine, I.D.; Maddux, B.A.; Grodsky, G.M. Are Oxidative Stress−Activated Signaling Pathways Mediators of Insulin Resistance and β-Cell Dysfunction? Diabetes 2003, 52, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Senoner, T.; Dichtl, W. Oxidative Stress in Cardiovascular Diseases: Still a Therapeutic Target? Nutrients 2019, 11, 2090. [Google Scholar] [CrossRef] [PubMed]
- Aune, D.; Keum, N.N.; Giovannucci, E.; Fadnes, L.T.; Boffetta, P.; Greenwood, D.C.; Tonstad, S.; Vatten, L.J.; Riboli, E.; Norat, T. Nut Consumption and Risk of Cardiovascular Disease, Total Cancer, All-Cause and Cause-Specific Mortality: A Systematic Review and Dose-Response Meta-Analysis of Prospective Studies. BMC Med. 2016, 14, 207. [Google Scholar] [CrossRef]
- Naghshi, S.; Sadeghian, M.; Nasiri, M.; Mobarak, S.; Asadi, M.; Sadeghi, O. Association of Total Nut, Tree Nut, Peanut, and Peanut Butter Consumption with Cancer Incidence and Mortality: A Comprehensive Systematic Review and Dose-Response Meta-Analysis of Observational Studies. Adv. Nutr. 2021, 12, 793–808. [Google Scholar] [CrossRef]
| SRP (n = 19) | PB (n = 23) | CB (n = 16) | p-Value | |
|---|---|---|---|---|
| Female, n (%) | 12 (66) | 18 (78) | 10 (63) | 0.528 |
| Age (years) | 22.42 ± 3.32 | 23.43 ± 2.90 | 22.38 ± 3.50 | 0.308 |
| Educational level, n (%) | 0.512 | |||
| University students | 10 (52.60) | 11 (48) | 11 (68.80) | |
| Graduated | 9 (47.40) | 12 (52) | 5 (31.20) | |
| Physical activity (MET/week) | 4935 ± 2191 | 4703 ± 2381 | 4920 ± 1670 | 0.769 |
| Anthropometric measurements | ||||
| Weight (kg) | 64.47 ± 10.68 | 60.10 ± 7.72 | 64.68 ± 10.68 | 0.228 |
| BMI (kg/m2) | 22.38 ± 3.61 | 22.19 ± 2.60 | 22.79 ± 2.86 | 0.793 |
| Waist circumference (cm) | 73.68 ± 8.12 | 71.28 ± 5.53 | 75.38 ± 6.30 | 0.156 |
| Waist-to-hip ratio | 0.74 ± 0.06 | 0.74 ± 0.05 | 0.77 ± 0.05 | 0.166 |
| Body fat (%) | 26.71 ± 8.48 | 28.45 ± 7.88 | 26.51 ± 7.41 | 0.634 |
| Blood Pressure | ||||
| Systolic blood pressure | 112 ± 7.33 | 110 ± 8.87 | 111 ± 10.20 | 0.327 |
| Diastolic blood pressure | 72.28 ± 7.93 | 72.87 ± 6.20 | 69.98 ± 9.37 | 0.476 |
| Biochemical parameters | ||||
| Glucose (mmol/L) | 4.58 ± 0.44 | 4.59 ± 0.35 | 4.44 ± 0.24 | 0.307 |
| Triglycerides (mmol/L) | 0.71 ± 0.21 | 0.85 ± 0.35 | 0.78 ± 0.25 | 0.410 |
| Total cholesterol (mmol/L) | 4.34 ± 0.48 | 4.60 ± 0.88 | 4.04 ± 0.68 | 0.104 |
| LDL-cholesterol (mmol/L) | 2.24 ± 0.40 | 2.60 ± 0.69 | 2.23 ± 0.52 | 0.141 |
| HDL-cholesterol (mmol/L) | 1.73 ± 0.24 | 1.69 ± 0.40 | 1.53 ± 0.32 | 0.016 |
| Urinary cortisol (nmol/L) | 574 ± 270 | 497 ± 155 | 466 ± 164 | 0.604 |
| Dietary intake | ||||
| Energy (kcal/day) | 2756 ± 603 | 2705 ± 602 | 2656 ± 485 | 0.835 |
| Carbohydrates (g/day) | 258 ± 84.76 | 241 ± 73.92 | 251 ± 63.80 | 0.840 |
| Sugar (g/day) | 116 ± 35.98 | 112 ± 35.04 | 119 ± 42.24 | 0.989 |
| Fiber (g/day) | 45.02 ± 23.03 | 42.12 ± 14.65 | 40.76 ± 15.17 | 0.915 |
| Protein (g/day) | 103 ± 30.00 | 110 ± 31.86 | 109 ± 29.64 | 0.577 |
| Total fat (g/day) | 143 ± 28.01 | 142 ± 35.35 | 134 ± 28.61 | 0.249 |
| SFAs (g/day) | 37.67 ± 9.56 | 38.18 ± 11.04 | 38.11 ± 12.96 | 0.825 |
| MUFAs (g/day) | 69.35 ± 15.10 | 69.06 ± 17.17 | 62.13 ± 15.24 | 0.326 |
| PUFAs (g/day) | 25.70 ± 7.10 | 23.99 ± 7.25 | 23.37 ± 7.25 | 0.617 |
| Polyphenol intake | ||||
| Flavonoids (mg/day) | 659 ± 392 | 855 ± 676 | 664 ± 620 | 0.463 |
| Phenolic acids (mg/day) | 178 ± 113 | 208 ± 155 | 169 ± 79.87 | 0.978 |
| Stilbenes (mg/day) | 0.28 ± 0.44 | 0.31 ± 0.45 | 0.11 ± 0.13 | 0.067 |
| Lignans (mg/day) | 5.69 ± 7.03 | 4.76 ± 5.22 | 4.03 ± 4.06 | 0.716 |
| Present in peanuts | ||||
| m-coumaric acid (mg/day) | 0.79 ± 0.76 | 0.64 ± 0.73 | 0.29 ± 0.34 | 0.035 |
| o-coumaric acid (mg/day) | 0.49 ± 0.39 | 0.39 ± 0.38 | 0.23 ± 0.20 | 0.052 |
| p-coumaric acid (mg/day) | 0.94 ± 0.43 | 0.72 ± 0.36 | 0.70 ± 0.38 | 0.144 |
| Resveratrol (mg/day) | 0.05 ± 0.03 | 0.04± 0.02 | 0.03 ± 0.02 | 0.668 |
| SRP vs. CB | PB vs. CB | ||||
|---|---|---|---|---|---|
| Models | Difference Time-Exposure (95% CI) | p-Value | Difference Time-Exposure (95% CI) | p-Value | |
| Telomere length | Model 1 | 0.50 (−0.02, 1.02) | 0.058 | 0.12 (−0.40, 0.64) | 0.653 |
| Model 2 | 0.53 (0.01, 1.05) | 0.048 | 0.12 (−0.42, 0.66) | 0.661 |
| Telomere Length | ||||
|---|---|---|---|---|
| n | B (CI) | p-Value | ||
| Waist-to-hip Ratio | Model 3 | 58 | −0.17 (−0.72, 0.38) | 0.534 |
| HDL-cholesterol | Model 3 | 57 | 0.23 (−0.68, 1.13) | 0.615 |
| Carbohydrates intake | Model 3 | 58 | −0.03 (−0.46, 0.39) | 0.880 |
| Fiber intake | Model 3 | 58 | −0.08 (−0.65, 0.49) | 0.782 |
| MUFA intake | Model 3 | 58 | 0.35 (0.01, 0.69) | 0.046 |
| Vitamin E intake | Model 3 | 58 | 0.09 (−0.39, 0.56) | 0.714 |
| m-Coumaric acid | Model 3 | 58 | 0.42 (0.18, 0.67) | 0.012 |
| o-Coumaric acid | Model 3 | 58 | 0.24 (−0.32, 0.81) | 0.394 |
| p-Coumaric acid | Model 3 | 58 | −0.35 (−1.79, 1.09) | 0.626 |
| Resveratrol | Model 3 | 58 | 0.68 (−0.70, 2.05) | 0.082 |
| Flavones | Model 3 | 58 | 0.31 (−0.33, 0.94) | 0.333 |
| Anthocyanins | Model 3 | 58 | 0.46 (−0.30, 1.23) | 0.225 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Oteros, D.; Parilli-Moser, I.; Laveriano Santos, E.P.; Becerra-Tomás, N.; Sanz-Lamora, H.; Hurtado-Barroso, S.; Haro, D.; Marrero, P.F.; Lamuela-Raventos, R.M.; Relat, J.; et al. Unveiling the Impact of Peanut Consumption on Telomere Length in Young and Healthy Individuals: Insights from the ARISTOTLE Study: A Randomized Clinical Trial. Antioxidants 2025, 14, 467. https://doi.org/10.3390/antiox14040467
Torres-Oteros D, Parilli-Moser I, Laveriano Santos EP, Becerra-Tomás N, Sanz-Lamora H, Hurtado-Barroso S, Haro D, Marrero PF, Lamuela-Raventos RM, Relat J, et al. Unveiling the Impact of Peanut Consumption on Telomere Length in Young and Healthy Individuals: Insights from the ARISTOTLE Study: A Randomized Clinical Trial. Antioxidants. 2025; 14(4):467. https://doi.org/10.3390/antiox14040467
Chicago/Turabian StyleTorres-Oteros, Daniel, Isabella Parilli-Moser, Emily P. Laveriano Santos, Nerea Becerra-Tomás, Héctor Sanz-Lamora, Sara Hurtado-Barroso, Diego Haro, Pedro F. Marrero, Rosa M. Lamuela-Raventos, Joana Relat, and et al. 2025. "Unveiling the Impact of Peanut Consumption on Telomere Length in Young and Healthy Individuals: Insights from the ARISTOTLE Study: A Randomized Clinical Trial" Antioxidants 14, no. 4: 467. https://doi.org/10.3390/antiox14040467
APA StyleTorres-Oteros, D., Parilli-Moser, I., Laveriano Santos, E. P., Becerra-Tomás, N., Sanz-Lamora, H., Hurtado-Barroso, S., Haro, D., Marrero, P. F., Lamuela-Raventos, R. M., Relat, J., & Canudas, S. (2025). Unveiling the Impact of Peanut Consumption on Telomere Length in Young and Healthy Individuals: Insights from the ARISTOTLE Study: A Randomized Clinical Trial. Antioxidants, 14(4), 467. https://doi.org/10.3390/antiox14040467