
Tetraselmis chuii Supplementation Increases Skeletal Muscle Nuclear Factor Erythroid 2-Related Factor 2 and Antioxidant Enzyme Gene Expression, and Peak Oxygen Uptake in Healthy Adults: A Randomised Crossover Trial

 , ,
, ,  , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Participant Characteristics
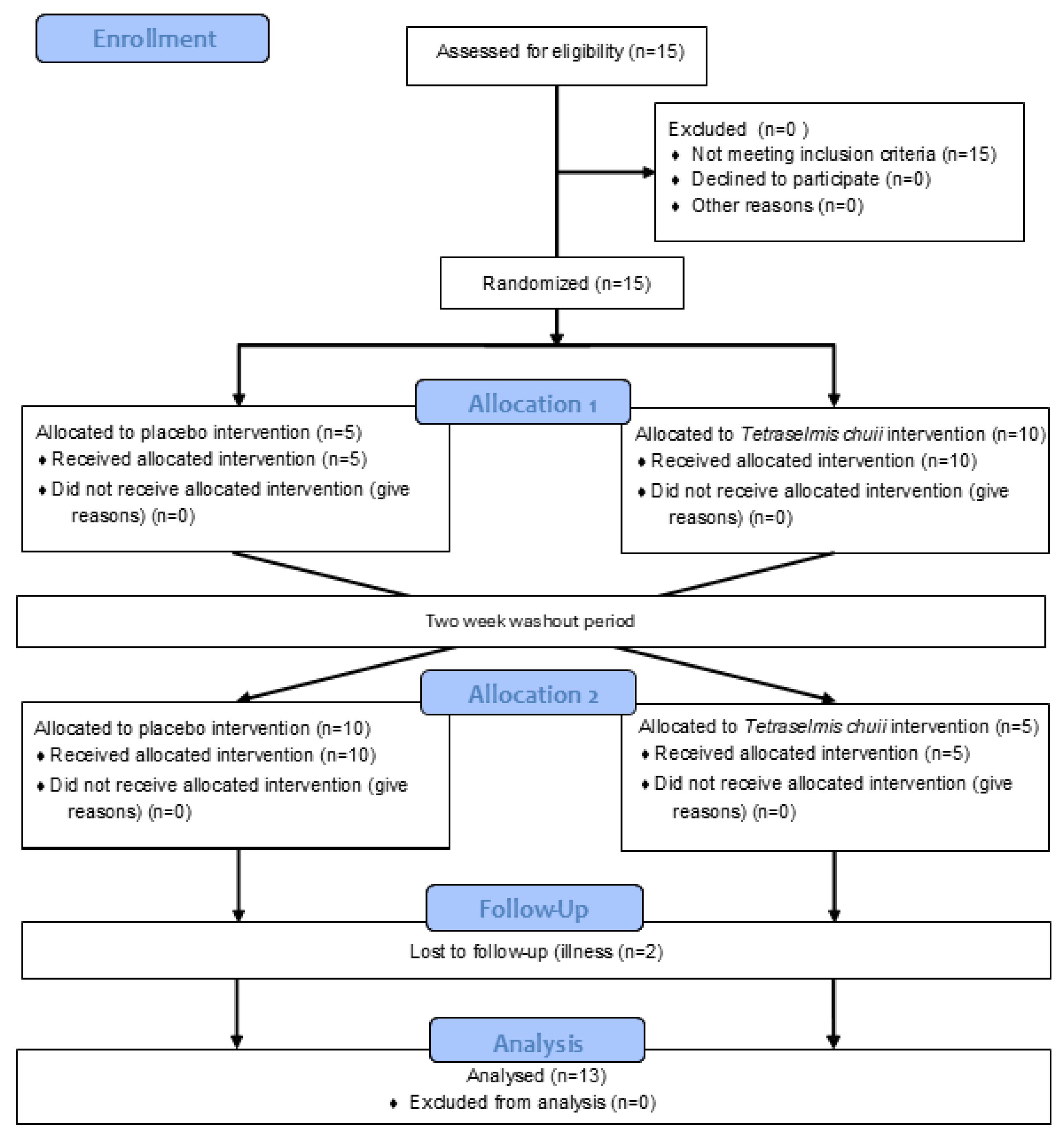
2.2. Experimental Design
2.3. Data Collection and Analysis Procedures
2.3.1. Skeletal Muscle Sampling
2.3.2. RNA Isolation
2.3.3. OpenArray™ Gene Expression Measurement
2.3.4. Gene Expression Data Analysis
2.3.5. Exercise Testing Procedures
2.3.6. Exercise Physiology and Tolerance Data Analysis Procedures
2.4. Statistical Analysis Procedures
3. Results
3.1. Human Skeletal Muscle Gene Expression
3.2. Exercise Physiology and Tolerance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rattan, S.I. Hormesis in aging. Ageing Res. Rev. 2008, 7, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Dhawan, G.; Kapoor, R.; Iavicoli, I.; Calabrese, V. HORMESIS: A Fundamental Concept with Widespread Biological and Biomedical Applications. Gerontology 2016, 62, 530–535. [Google Scholar] [PubMed]
- McCord, J.M.; Gao, B.; Hybertson, B.M. The Complex Genetic and Epigenetic Regulation of the Nrf2 Pathways: A Review. Antioxidants 2023, 12, 366. [Google Scholar] [CrossRef]
- Hayes, J.D.; Dinkova-Kostova, A.T. The Nrf2 Regulatory Network Provides an Interface between Redox and Intermediary Metabolism. Trends Biochem. Sci. 2014, 39, 199–218. [Google Scholar] [CrossRef]
- He, F.; Ru, X.; Wen, T. NRF2, a Transcription Factor for Stress Response and Beyond. Int. J. Mol. Sci. 2020, 21, 4777. [Google Scholar] [CrossRef]
- Onoki, T.; Izumi, Y.; Takahashi, M.; Murakami, S.; Matsumaru, D.; Ohta, N.; Wati, S.M.; Hatanaka, N.; Katsuoka, F.; Okutsu, M.; et al. Skeletal muscle-specific Keap1 disruption modulates fatty acid utilization and enhances exercise capacity in female mice. Redox Biol. 2021, 43, 101966. [Google Scholar] [CrossRef]
- Wang, L.; Yang, S.; Yan, L.; Wei, H.; Wang, J.; Yu, S.; Kong, A.T.; Zhang, Y. Hypoxia preconditioning promotes endurance exercise capacity of mice by activating skeletal muscle Nrf2. J. Appl. Physiol. 2019, 127, 1267–1277. [Google Scholar] [CrossRef]
- Gao, L.; Kumar, V.; Vellichirammal, N.N.; Park, S.Y.; Rudebush, T.L.; Yu, L.; Son, W.M.; Pekas, E.J.; Wafi, A.M.; Hong, J.; et al. Functional, proteomic and bioinformatic analyses of Nrf2- and Keap1- null skeletal muscle. J. Physiol. 2020, 598, 5427–5451. [Google Scholar] [CrossRef]
- Martinez-Canton, M.; Galvan-Alvarez, V.; Martin-Rincon, M.; Calbet, J.A.L.; Gallego-Selles, A. Unlocking peak performance: The role of Nrf2 in enhancing exercise outcomes and training adaptation in humans. Free Radic. Biol. Med. 2024, 224, 168–181. [Google Scholar] [CrossRef]
- Cokdinleyen, M.; Alvarez-Rivera, G.; Tejera, J.L.G.; Mendiola, J.A.; Valdés, A.; Kara, H.; Ibáñez, E.; Cifuentes, A. Tetraselmis chuii Edible Microalga as a New Source of Neuroprotective Compounds Obtained Using Fast Biosolvent Extraction. Int. J. Mol. Sci. 2024, 25, 3897. [Google Scholar] [CrossRef]
- Mantecón, L.; Moyano, R.; Cameán, A.M.; Jos, A. Safety assessment of a lyophilized biomass of Tetraselmis chuii (TetraSOD®) in a 90 day feeding study. Food Chem. Toxicol. 2019, 133, 110810. [Google Scholar] [CrossRef]
- Ramírez, P.; Torres, S.; Lama, C.; Mantecón, L.; Unamunzaga, C.; Infante, C. TetraSOD® activates the antioxidant response pathway in human cells: An in vitro approach. Afr. J. Biotechnol. 2020, 19, 367–373. [Google Scholar]
- Sharp, M.; Sahin, K.; Stefan, M.; Orhan, C.; Gheith, R.; Reber, D.; Sahin, N.; Tuzcu, M.; Lowery, R.; Durkee, S.; et al. Phytoplankton Supplementation Lowers Muscle Damage and Sustains Performance across Repeated Exercise Bouts in Humans and Improves Antioxidant Capacity in a Mechanistic Animal. Nutrients 2020, 12, 1990. [Google Scholar] [CrossRef]
- Sharp, M.H.; Sahin, K.; Stefan, M.W.; Gheith, R.H.; Reber, D.D.; Ottinger, C.R.; Orhan, C.; Tuzcu, M.; Sahin, N.; Lowery, R.P.; et al. Marine Phytoplankton Improves Exercise Recovery in Humans and Activates Repair Mechanisms in Rats. Int. J. Sports Med. 2021, 42, 1070–1082. [Google Scholar] [CrossRef]
- Sharp, M.; Wilson, J.; Stefan, M.; Gheith, R.; Lowery, R.; Ottinger, C.; Reber, D.; Orhan, C.; Sahin, N.; Tuzcu, M.; et al. Marine phytoplankton improves recovery and sustains immune function in humans and lowers proinflammatory immunoregulatory cytokines in a rat model. Phys. Act. Nutr. 2021, 25, 42–55. [Google Scholar] [CrossRef]
- Clifford, T.; Acton, J.P.; Cocksedge, S.P.; Davies, K.A.B.; Bailey, S.J. The effect of dietary phytochemicals on nuclear factor erythroid 2-related factor 2 (Nrf2) activation: A systematic review of human intervention trials. Mol. Biol. Rep. 2021, 48, 1745–1761. [Google Scholar] [CrossRef]
- Thorley, J.; Thomas, C.; Thon, N.; Nuttall, H.; Martin, N.R.W.; Bishop, N.; Bailey, S.J.; Clifford, T. Combined effects of green tea supplementation and eccentric exercise on nuclear factor erythroid 2-related factor 2 activity. Eur. J. Appl. Physiol. 2024, 124, 245–256. [Google Scholar] [CrossRef]
- Thorley, J.; Alhebshi, A.; Rodriguez-Mateos, A.; Zhang, Z.; Bailey, S.J.; Martin, N.R.W.; Bishop, N.C.; Clifford, T. Acute supplementation with a curcuminoid-based formulation fails to enhance resting or exercise-induced NRF2 activity in males and females. Food Funct. 2024, 15, 10782–10794. [Google Scholar] [CrossRef]
- Toro, V.; Siquier-Coll, J.; Bartolomé, I.; Robles-Gil, M.C.; Rodrigo, J.; Maynar-Mariño, M. Effects of Tetraselmis chuii Microalgae Supplementation on Ergospirometric, Haematological and Biochemical Parameters in Amateur Soccer Players. Int. J. Environ. Res. Public Health 2020, 17, 6885. [Google Scholar] [CrossRef]
- García, Á.; Toro-Román, V.; Siquier-Coll, J.; Bartolomé, I.; Muñoz, D.; Maynar-Mariño, M. Effects of Tetraselmis chuii Microalgae Supplementation on Anthropometric, Hormonal and Hematological Parameters in Healthy Young Men: A Double-Blind Study. Int. J. Environ. Res. Public Health 2022, 19, 6060. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Nikolaidis, M.G.; Margaritelis, N.V.; Matsakas, A. Quantitative Redox Biology of Exercise. Int. J. Sports Med. 2020, 41, 633–645. [Google Scholar] [CrossRef]
- Westerblad, H.; Allen, D.G. Emerging roles of ROS/RNS in muscle function and fatigue. Antioxid. Redox Signal. 2011, 15, 2487–2499. [Google Scholar] [CrossRef]
- Uhlen, M.; Oksvold, P.; Fagerberg, L.; Lundberg, E.; Jonasson, K.; Forsberg, M.; Zwahlen, M.; Kampf, C.; Wester, K.; Hober, S.; et al. Towards a knowledge-based Human Protein Atlas. Nat. Biotechnol. 2010, 28, 1248–1250. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−ΔΔC(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef]
- Kuang, J.; McGinley, C.; Lee, M.J.-C.; Saner, N.J.; Garnham, A.; Bishop, D.J. Interpretation of exericse-induced changes in human skeletal muscle mRNA expression depends on the timing of the post-exercise biopsies. Peer J. 2022, 10, e12856. [Google Scholar] [CrossRef]
- Bailey, S.J.; Wilkerson, D.P.; Dimenna, F.J.; Jones, A.M. Influence of repeated sprint training on pulmonary O2 uptake and muscle deoxygenation kinetics in humans. J. Appl. Physiol. 2009, 106, 1875–1887. [Google Scholar] [CrossRef]
- Whipp, B.J.; Davis, J.A.; Torres, F.; Wasserman, K. A test to determine parameters of aerobic function during exercise. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1981, 50, 217–221. [Google Scholar] [CrossRef]
- Vomund, S.; Schäfer, A.; Parnham, M.J.; Brüne, B.; von Knethen, A. Nrf2, the Master Regulator of Anti-Oxidative Responses. Int. J. Mol. Sci. 2017, 18, 2772. [Google Scholar] [CrossRef]
- Pei, J.; Pan, X.; Wei, G.; Hua, Y. Research Progress of Glutathione Peroxidase Family (GPX) in Redoxidation. Front. Pharmacol. 2023, 14, 1147414. [Google Scholar] [CrossRef]
- Manevich, Y.; Fisher, A.B. Peroxiredoxin 6, a 1-Cys peroxiredoxin, functions in antioxidant defense and lung phospholipid metabolism. Free Radic. Biol. Med. 2005, 38, 1422–1432. [Google Scholar] [CrossRef]
- Wadley, A.J.; Aldred, S.; Coles, S.J. An unexplored role for Peroxiredoxin in exercise-induced redox signalling? Redox Biol. 2016, 8, 51–58. [Google Scholar] [CrossRef]
- Averill-Bates, D.A. The antioxidant glutathione. Vitam. Horm. 2023, 121, 109–141. [Google Scholar] [CrossRef]
- Grillo, M.P. Bioactivation by Phase-II-Enzyme-Catalyzed Conjugation of Xenobiotics. In Encyclopedia of Drug Metabolism and Interactions; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012. [Google Scholar] [CrossRef]
- Hayes, J.D.; McLellan, L.I. Glutathione and glutathione-dependent enzymes represent a co-ordinately regulated defence against oxidative stress. Free Radic. Res. 1999, 31, 273–300. [Google Scholar] [CrossRef]
- Barbieri, E.; Sestili, P. Reactive oxygen species in skeletal muscle signaling. J. Signal Transduct. 2012, 2012, 982794. [Google Scholar] [CrossRef]
- Murakami, A. Non-specific protein modifications may be novel mechanism underlying bioactive phytochemicals. J. Clin. Biochem. Nutr. 2018, 62, 115–123. [Google Scholar] [CrossRef]
- Scuto, M.; Rampulla, F.; Reali, G.M.; Spanò, S.M.; Trovato Salinaro, A.; Calabrese, V. Hormetic Nutrition and Redox Regulation in Gut-Brain Axis Disorders. Antioxidants 2024, 13, 484. [Google Scholar] [CrossRef]
- L’honoré, A.; Commère, P.H.; Negroni, E.; Pallafacchina, G.; Friguet, B.; Drouin, J.; Buckingham, M.; Montarras, D. The role of Pitx2 and Pitx3 in muscle stem cells gives new insights into P38α MAP kinase and redox regulation of muscle regeneration. eLife 2018, 7, e32991. [Google Scholar] [CrossRef]
- Ji, L.L.; Kang, C.; Zhang, Y. Exercise-induced hormesis and skeletal muscle health. Free Radic. Biol. Med. 2016, 98, 113–122. [Google Scholar] [CrossRef]
- Fittipaldi, S.; Mercatelli, N.; Dimauro, I.; Jackson, M.J.; Paronetto, M.P.; Caporossi, D. Alpha B-crystallin induction in skeletal muscle cells under redox imbalance is mediated by a JNK-dependent regulatory mechanism. Free Radic. Biol. Med. 2015, 86, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Jeyapaul, J.; Jaiswal, A.K. Nrf2 and c-Jun regulation of antioxidant response element (ARE)-mediated expression and induction of gamma-glutamylcysteine synthetase heavy subunit gene. Biochem. Pharmacol. 2000, 59, 1433–1439. [Google Scholar] [CrossRef]
- Venugopal, R.; Jaiswal, A.K. Nrf2 and Nrf1 in association with Jun proteins regulate antioxidant response element-mediated expression and coordinated induction of genes encoding detoxifying enzymes. Oncogene 1998, 17, 3145–3156. [Google Scholar] [CrossRef]
- Singh, V.; Ubaid, S. Role of Silent Information Regulator 1 (SIRT1) in Regulating Oxidative Stress and Inflammation. Inflammation 2020, 43, 1589–1598. [Google Scholar] [CrossRef]
- Baird, L.; Yamamoto, M. The Molecular Mechanisms Regulating the NRF2 Pathway. Mol. Cell Biol. 2020, 40, e00099-20. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Holtzclaw, W.D.; Kensler, T.W. The role of Keap1 in cellular protective responses. Chem. Res. Toxicol. 2005, 18, 1779–1791. [Google Scholar] [CrossRef]
- Reid, M.B. Invited Review: Redox modulation of skeletal muscle contraction: What we know and what we don’t. J. Appl. Physiol. 2001, 90, 724–731. [Google Scholar] [CrossRef]
- Zhang, D.D.; Lo, S.C.; Sun, Z.; Habib, G.M.; Lieberman, M.W.; Hannink, M. Ubiquitination of Keap1, a BTB-Kelch Substrate Adaptor Protein for Cul3, Targets Keap1 for Degradation by a Proteasome-Independent Pathway. J. Biol. Chem. 2005, 280, 30091–30099. [Google Scholar] [CrossRef]
- Tranah, G.J.; Barnes, H.N.; Cawthon, P.M.; Coen, P.M.; Esser, K.A.; Hepple, R.T.; Huo, Z.; Kramer, P.A.; Toledo, F.G.S.; Zhang, X.; et al. Expression of mitochondrial oxidative stress response genes in muscle is associated with mitochondrial respiration, physical performance, and muscle mass in the Study of Muscle, Mobility, and Aging. Aging Cell 2024, 23, e14114. [Google Scholar] [CrossRef]
- Islam, H.; Bonafiglia, J.T.; Turnbull, P.C.; Simpson, C.A.; Perry, C.G.R.; Gurd, B.J. The impact of acute and chronic exercise on Nrf2 expression in relation to markers of mitochondrial biogenesis in human skeletal muscle. Eur. J. Appl. Physiol. 2020, 120, 149–160. [Google Scholar] [CrossRef]
- Russell, A.P.; Foletta, V.C.; Snow, R.J.; Wadley, G.D. Skeletal muscle mitochondria: A major player in exercise, health and disease. Biochim. Biophys. Acta 2014, 1840, 1276–1284. [Google Scholar] [CrossRef]
- Islam, H.; Hood, D.A.; Gurd, B.J. Looking beyond PGC-1α: Emerging regulators of exercise-induced skeletal muscle mitochondrial biogenesis and their activation by dietary compounds. Appl. Physiol. Nutr. Metab. 2020, 45, 11–23. [Google Scholar] [CrossRef]
- Kasiak, P.; Kowalski, T.; Rębiś, K.; Klusiewicz, A.; Ładyga, M.; Sadowska, D.; Wilk, A.; Wiecha, S.; Barylski, M.; Poliwczak, A.R.; et al. Is the Ventilatory Efficiency in Endurance Athletes Different?-Findings from the NOODLE Study. J. Clin. Med. 2024, 13, 490. [Google Scholar] [CrossRef]
- Joyner, M.J.; Coyle, E.F. Endurance exercise performance: The physiology of champions. J. Physiol. 2008, 586, 35–44. [Google Scholar]
- Kurl, S.; Hakkarainen, P.; Voutilainen, A.; Lönnroos, E. Combined effects of maximal oxygen uptake and glucose status on mortality: The Prospective KIHD cohort study. Scand. J. Med. Sci. Sports 2022, 32, 913–923. [Google Scholar]
- Ung, G.A.; Nguyen, K.H.; Hui, A.; Wong, N.D.; Dineen, E.H. Impact of cardiorespiratory fitness and diabetes status on cardiovascular disease and all-cause mortality: An NHANES retrospective cohort study. Am. Heart J. Plus 2024, 42, 100395. [Google Scholar] [CrossRef]
- Margaritelis, N.V.; Paschalis, V.; Theodorou, A.A.; Kyparos, A.; Nikolaidis, M.G. Antioxidant supplementation, redox deficiencies and exercise performance: A falsification design. Free Radic. Biol. Med. 2020, 158, 44–52. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Symbol | ThermoFisher Scientific Assay ID |
|---|---|---|
| Cellular Response to Chemical Stress | ||
| Catalase | CAT | Hs00156308_m1 |
| Ferritin Light Chain | FTL | Hs00830226_gH |
| Glutamate-Cysteine Ligase Catalytic Subunit | GCLC | Hs00155249_m1 |
| Glutamate-Cysteine Ligase Modifier Subunit | GCLM | Hs00978072_m1 |
| Glutaredoxin | GLRX | Hs00829752_g1 |
| Glutathione Peroxidase 1 | GPX1 | Hs00829989_gH |
| Glutathione Peroxidase 3 | GPX3 | Hs01078668_m1 |
| Glutathione Peroxidase 4 | GPX4 | Hs00989766_g1 |
| Glutathione Peroxidase 7 | GPX7 | Hs04194449_s1 |
| Glutathione Disulfide Reductase | GSR | Hs00167317_m1 |
| Heme Oxygenase 1 | HMOX1 | Hs01110250_m1 |
| Heme Oxygenase 2 | HMOX2 | Hs01558390_m1 |
| Kelch-Like ECH Associated Protein 1 | KEAP1 | Hs00202227_m1 |
| Nuclear Factor, Erythroid 2 Like 2 | NRF2 | Hs00975961_g1 |
| Peroxiredoxin 1 | PRDX1 | Hs00602020_mH |
| Peroxiredoxin 2 | PRDX2 | Hs00853603_s1 |
| Peroxiredoxin 3 | PRDX3 | Hs00428953_g1 |
| Peroxiredoxin 4 | PRDX4 | Hs01056076_m1 |
| Peroxiredoxin 5 | PRDX5 | Hs00738905_g1 |
| Peroxiredoxin 6 | PRDX6 | Hs00705355_s1 |
| Superoxide Dismutase 1 | SOD1 | Hs00533490_m1 |
| Superoxide Dismutase 2 | SOD2 | Hs00167309_m1 |
| Superoxide Dismutase 3 | SOD3 | Hs04973910_s1 |
| Sulfiredoxin 1 | SRXN1 | Hs00607800_m1 |
| Thioredoxin 2 | TXN2 | Hs00429399_g1 |
| Thioredoxin Reductase 1 | TXNRD1 | Hs00917067_m1 |
| Thioredoxin Reductase 2 | TXNRD2 | Hs00272352_m1 |
| Cellular Source of Chemical Stress | ||
| Phospholipase A2 Group XIIA | PLA2G12A | Hs04976755_s1 |
| Phospholipase A2 Group XV | PLA2G15 | Hs00202399_m1 |
| Phospholipase A2 Group XVI | PLA2G16 | Hs00912734_m1 |
| Phospholipase A2 Group IIA | PLA2G2A | Hs00179898_m1 |
| Phospholipase A2 Group IVC | PLA2G4C | Hs01003754_m1 |
| Phospholipase A2 Group IVF | PLA2G4F | Hs02577398_m1 |
| Biological Oxidations | ||
| Glutathione S-Transferase Kappa 1 | GSTK1 | Hs01114170_m1 |
| Glutathione S-Transferase Mu 1 | GSTM1 | Hs01683722_gH |
| Glutathione S-Transferase Mu 2 | GSTM2 | Hs00265266_g1 |
| Glutathione S-Transferase Mu 3 | GSTM3 | Hs00356079_m1 |
| Glutathione S-Transferase Mu 4 | GSTM4 | Hs00426432_m1 |
| Glutathione S-Transferase Mu 5 | GSTM5 | Hs00757076_m1 |
| Glutathione S-Transferase Omega 1 | GSTO1 | Hs02383465_s1 |
| Glutathione S-Transferase Pi 1 | GSTP1 | Hs00943350_g1 |
| Glutathione S-Transferase Theta 2 | GSTT2 | Hs04942283_g1 |
| Microsomal Glutathione S-Transferase 3 | MGST3 | Hs01058946_m1 |
| NAD(P)H Quinone Dehydrogenase 1 | NQO1 | Hs01045993_g1 |
| NAD(P)H Quinone Dehydrogenase 2 | NQO2 | Hs01056950_m1 |
| Signalling Pathways | ||
| C-C Motif Chemokine Ligand 2 | CCL2 | Hs00234140_m1 |
| Extracellular Signal-Regulated Kinase 1 | ERK1 (MAPK3) | Hs00385075_m1 |
| Extracellular Signal-Regulated Kinase 2 | ERK2 (MAPK1) | Hs01046830_m1 |
| Extracellular Signal-Regulated Kinase 3 | ERK3 (MAPK6) | Hs00833126_g1 |
| Extracellular Signal-Regulated Kinase 5 | ERK5 (MAPK7) | Hs00611114_g1 |
| Interleukin 10 | IL10 | Hs00961622_m1 |
| Interleukin 18 | IL18 | Hs01038788_m1 |
| Interleukin 1 Receptor Antagonist | IL1RN | Hs00893626_m1 |
| Interleukin 6 | IL6 | Hs00174131_m1 |
| Nuclear Factor Kappa B Subunit 1 | NFKB1 | Hs00765730_m1 |
| Nuclear Factor Kappa B Subunit 2 | NFKB2 | Hs01028890_g1 |
| NLR Family Pyrin Domain Containing 3 | NLRP3 | Hs00918082_m1 |
| p38 Alpha | p38a (MAPK14) | Hs01051152_m1 |
| p38 Gamma | p38g (MAPK12) | Hs00268060_m1 |
| Sirtuin 1 | SIRT1 | Hs01009006_m1 |
| Tumor Necrosis Factor Alpha | TNFa | Hs00174128_m1 |
| Antioxidant Response Elements | ||
| Activating Transcription Factor 1 | ATF1 | Hs00909673_m1 |
| Activating Transcription Factor 4 | ATF4 | Hs00909569_g1 |
| BTB Domain and CNC Homolog 1 | BACH1 | Hs00230917_m1 |
| BTB Domain and CNC Homolog 2 | BACH2 | Hs00935338_m1 |
| BTB Domain Containing 1 | BTBD1 | Hs00228832_m1 |
| BTB Domain Containing 10 | BTBD10 | Hs00900692_g1 |
| BTB Domain Containing 2 | BTBD2 | Hs00215064_m1 |
| BTB Domain Containing 6 | BTBD6 | Hs00746410_s1 |
| BTB Domain Containing 7 | BTBD7 | Hs01568248_m1 |
| BTB Domain Containing 9 | BTBD9 | Hs04987111_m1 |
| FOS Proto-Oncogene | FOS | Hs04194186_s1 |
| FOS Like 1 | FOSL1 | Hs00759776_s1 |
| FOS Like 2 | FOSL2 | Hs01050117_m1 |
| Forkhead Box O3 | FOXO3 | Hs00818121_m1 |
| JUN Proto-Oncogene | JUN | Hs01103582_s1 |
| JUNB Proto-Oncogene | JUNB | Hs00357891_s1 |
| JUND Proto-Oncogene | JUND | Hs04187679_s1 |
| Kruppel Like Factor 2 | KLF2 | Hs07291763_gH |
| MAF bZip Transcription Factor | MAF | Hs04185012_s1 |
| MAF bZip Transcription Factor A | MAFA | Hs04419852_s1 |
| MAF bZip Transcription Factor B | MAFB | Hs00534343_s1 |
| MAF bZip Transcription Factor F | MAFF | Hs05026540_g1 |
| MAF bZip Transcription Factor G | MAFG | Hs00536278_s1 |
| PPARG Coactivator 1 Alpha | PGC1a | Hs00173304_m1 |
| Cell Fate | ||
| Apoptosis Inducing Factor | AIFM1 | Hs00377585_m1 |
| BCL2 Antagonist/Killer 1 | BAK1 | Hs00832876_g1 |
| BCL2 Associated X | BAX | Hs00180269_m1 |
| BCL2 Apoptosis Regulator | BCL2 | Hs04986394_s1 |
| BH3 Interacting Domain Death Agonist | BID | Hs00609632_m1 |
| Caspase 1 | CASP1 | Hs00354836_m1 |
| Caspase 10 | CASP10 | Hs01017899_m1 |
| Caspase 3 | CASP3 | Hs00234387_m1 |
| Caspase 8 | CASP8 | Hs06630780_s1 |
| CASP8 and FADD Like Apoptosis Regulator | CFLAR | Hs01116280_m1 |
| Tumor Protein p53 | TP53 | Hs01034249_m1 |
| Miscellaneous | ||
| Apurinic/Apyrimidinic Endodeoxyribonuclease 1 | APEX1 | Hs00172396_m1 |
| Calpain 1 | CAPN1 | Hs00559804_m1 |
| Calpain 3 | CAPN3 | Hs00544982_m1 |
| Cullin 3 | CUL3 | Hs00180183_m1 |
| Metallothionein 2A | MT2A | Hs02379661_g1 |
| N-Terminal EF-Hand Calcium Binding Protein 1 | Necab1 | Hs00242747_m1 |
| Nitric Oxide Synthase 2 | NOS2 | Hs01075529_m1 |
| Nitric Oxide Synthase 3 | NOS3 | Hs01574665_m1 |
| Sequestosome 1 | SQSTM1 | Hs01061917_g1 |
| Uncoupling Protein 2 | UCP2 | Hs01075227_m1 |
| Vascular Endothelial Growth Factor A | VEGFA | Hs00900055_m1 |
| Reference Gene Candidates | ||
| Actin Beta | ACTB | Hs01060665_g1 |
| Beta 2 Microglobulin | B2M | Hs00187842_m1 |
| Glyceraldehyde-3-Phosphate Dehydrogenase | GAPDH | Hs02786624_g1 |
| Protein Phosphatase 1 Catalytic Subunit Alpha | PPP1CA | Hs00267568_m1 |
| TATA-Box Binding Protein | TBP | Hs00427620_m1 |
| Gene Name | Gene Symbol | 2−ΔΔCq (GM ± GSD) | t | p w | dz | ES r |
|---|---|---|---|---|---|---|
| PLA-T. chuii (p < 0.05, Medium to Large ES) | ||||||
| BTB domain containing 7 | BTBD7 | 1.38 ± 1.62 | 0.03 | 0.7 | ||
| Caspase 10 | CASP10 | 1.34 ± 1.59 | 0.04 | 0.6 | ||
| C-C Motif Chemokine Ligand 2 | CCL2 | 1.39 ± 1.39 | 0.00 | 1.0 | ||
| Cullin 3 | CUL3 | 1.77 ± 2.49 | 0.04 | 0.6 | ||
| Extracellular Signal-Regulated Kinase 3 | ERK3 (MAPK6) | 1.92 ± 2.42 | 0.02 | 0.7 | ||
| Glutathione Peroxidase 7 | GPX7 | 1.26 ± 1.37 | 0.02 | 0.7 | ||
| Glutathione-Disulfide Reductase | GSR | 1.22 ± 1.41 | 0.05 | 0.6 | ||
| Glutathione S-Transferase Mu 3 | GSTM3 | 1.34 ± 1.49 | 0.02 | 0.7 | ||
| JUN Proto-Oncogene | JUN | 1.53 ± 1.89 | 0.03 | 0.7 | ||
| Nuclear Factor, Erythroid 2 Like 2 | NRF2 | 1.62 ± 2.16 | 0.03 | 0.6 | ||
| p38 Alpha | p38a (MAPK14) | 1.33 ± 1.58 | 0.04 | 0.6 | ||
| Phospholipase A2 Group XIIA | PLA2G12A | 1.63 ± 2.05 | 0.03 | 0.7 | ||
| Phospholipase A2 Group XVI | PLA2G16 | 1.36 ± 1.42 | 0.01 | 0.9 | ||
| Peroxiredoxin 6 | PRDX6 | 1.36 ± 1.57 | 0.03 | 0.7 | ||
| Sirtuin 1 | SIRT1 | 1.73 ± 2.25 | 0.03 | 0.7 | ||
| PLA-T. chuii (p > 0.05, p < 0.1, Medium ES) | ||||||
| Apoptosis Inducing Factor | AIFM1 | 1.49 ± 2.14 | 0.08 | 0.5 | ||
| Activating Transcription Factor 1 | ATF1 | 1.54 ± 2.30 | 0.09 | 0.5 | ||
| BTB Domain and CNC Homolog 1 | BACH1 | 1.61 ± 2.39 | 0.07 | 0.5 | ||
| BTB domain containing 1 | BTBD1 | 1.37 ± 1.79 | 0.08 | 0.5 | ||
| Calpain 3 | CAPN3 | 1.29 ± 1.53 | 0.05 | 0.6 | ||
| Caspase 8 | CASP8 | 1.44 ± 1.95 | 0.07 | 0.5 | ||
| Catalase | CAT | 1.40 ± 1.87 | 0.07 | 0.5 | ||
| Extracellular Signal-Regulated Kinase 2 | ERK2 (MAPK1) | 1.32 ± 1.60 | 0.06 | 0.6 | ||
| Extracellular Signal-Regulated Kinase 5 | ERK5 (MAPK7) | 0.87 ± 1.27 | 0.05 | 0.6 | ||
| Glutamate-Cysteine Ligase Modifier Subunit | GCLM | 1.38 ± 1.78 | 0.07 | 0.6 | ||
| Interleukin 18 | IL18 | 2.05 ± 3.29 | 0.05 | 0.6 | ||
| Peroxiredoxin 3 | PRDX3 | 1.30 ± 1.56 | 0.06 | 0.6 | ||
| Superoxide Dismutase 2 | SOD2 | 1.40 ± 1.86 | 0.08 | 0.5 | ||
| Gene Name | Gene Symbol | r | CC p |
|---|---|---|---|
| PLA-(ΔPLA-T. chuii) (p < 0.05, Strong CC) | |||
| Apoptosis Inducing Factor | AIFM1 | −0.8 | <0.01 |
| Apurinic/Apyrimidinic Endodeoxyribonuclease 1 | APEX1 | −0.8 | <0.01 |
| Activating Transcription Factor 1 | ATF1 | −0.9 | <0.01 |
| BTB Domain and CNC Homolog 1 | BACH1 | −0.8 | <0.01 |
| Bcl2 Apoptosis Regulator | BCL2 | −0.9 | <0.01 |
| BTB Domain Containing 2 | BTBD2 | −0.8 | <0.01 |
| BTB Domain Containing 9 | BTBD9 | −0.8 | <0.01 |
| Caspase 1 | CASP1 | −0.9 | <0.01 |
| Caspase 3 | CASP3 | −0.9 | <0.01 |
| Caspase 8 | CASP8 | −0.8 | <0.01 |
| Catalase | CAT | −0.8 | <0.01 |
| Cullin 3 | CUL3 | −0.9 | <0.01 |
| Extracellular Signal-Regulated Kinase 3 | ERK3 (MAPK6) | −0.8 | <0.01 |
| Forkhead Box O3 | FOXO3 | −0.8 | <0.01 |
| Lipoxygenase Homology Domains 1 | LOXHD1 | −0.8 | <0.01 |
| MAF bZip Transcription Factor G | MAFG | −0.8 | <0.01 |
| Microtubule Associated Protein Tau | MAPT | −0.9 | <0.01 |
| N-Terminal EF-Hand Calcium Binding Protein 1 | Necab1 | −0.8 | <0.01 |
| Nuclear Factor Kappa B Subunit 1 | NFKB1 | −0.9 | <0.01 |
| PPARG Coactivator 1 Alpha | PGC1a | −0.9 | <0.01 |
| Phospholipase A2 Group XIIA | PLA2G12A | −0.8 | <0.01 |
| Peroxiredoxin 3 | PRDX3 | −0.8 | <0.01 |
| Superoxide Dismutase 2 | SOD2 | −0.8 | <0.01 |
| Thioredoxin Reductase 1 | TXNRD1 | −0.9 | <0.01 |
| Thioredoxin Reductase 2 | TXNRD2 | −0.8 | <0.01 |
| PLA-(ΔPLA-T. chuii) (p < 0.05, Moderate CC) | |||
| BCL2 Antagonist/Killer 1 | BAX | −0.7 | <0.01 |
| BCL2 Antagonist/Killer 1 | BAK1 | −0.6 | 0.03 |
| Calpain 3 | CAPN3 | −0.6 | 0.02 |
| Extracellular Signal-Regulated Kinase 5 | ERK5 (MAPK7) | −0.7 | 0.01 |
| FOS Like 2 | FOSL2 | −0.7 | 0.01 |
| Ferritin Light Chain | FTL | −0.6 | 0.04 |
| Glutamate-Cysteine Ligase Modifier Subunit | GCLM | −0.7 | 0.01 |
| Glutaredoxin | GLRX | −0.6 | 0.04 |
| Glutathione Peroxidase 1 | GPX1 | −0.7 | 0.01 |
| Glutathione S-Transferase Mu 4 | GSTM4 | −0.6 | 0.05 |
| Glutathione S-Transferase Pi 1 | GSTP1 | −0.6 | 0.02 |
| Heme Oxygenase 2 | HMOX2 | −0.6 | 0.02 |
| Interleukin 18 | IL18 | −0.7 | 0.01 |
| JUND Proto-Oncogene | JUND | −0.7 | 0.01 |
| Kruppel Like Factor 2 | KLF2 | −0.7 | 0.01 |
| MAF bZip Transcription Factor | MAF | −0.7 | 0.01 |
| MAF bZip Transcription Factor B | MAFB | −0.6 | 0.04 |
| Microsomal Glutathione S-Transferase 3 | MGST3 | −0.7 | 0.00 |
| Nitric Oxide Synthase 3 | NOS3 | −0.7 | 0.01 |
| Nuclear Factor Kappa B Subunit 2 | NFKB2 | −0.7 | 0.01 |
| P38 Gamma | p38g (MAPK12) | −0.7 | 0.01 |
| Phospholipase A2 Group XV | PLA2G15 | −0.7 | 0.02 |
| Phospholipase A2 Group IVC | PLA2G4C | −0.7 | 0.01 |
| Peroxiredoxin 5 | PRDX5 | −0.6 | 0.03 |
| Peroxiredoxin 6 | PRDX6 | −0.6 | 0.03 |
| Sequestosome 1 | SQSTM1 | −0.6 | 0.03 |
| Sirtuin 1 | SIRT1 | −0.7 | 0.00 |
| Superoxide Dismutase 3 | SOD3 | −0.7 | 0.01 |
| Thioredoxin 2 | TXN2 | −0.7 | 0.00 |
| PLA-Pre | PLA-Post | T. chuii-Pre | T. chuii-Post | |
|---|---|---|---|---|
| End-exercise (L/min) | 4.02 ± 0.38 | 3.99 ± 0.37 | 3.92 ± 0.28 | 4.06 ± 0.38 * |
| End-exercise RPE (AU) | 20 ± 0 | 20 ± 1 | 20 ± 1 | 20 ± 0 |
| End-exercise B[La] (mmol/L) | 10.46 ± 2.17 | 11.02 ± 1.34 | 10.85 ± 1.92 | 11.52 ± 1.91 |
| Tlim (s) | 410 ± 80 | 456 ± 134 | 411 ± 82 | 480 ± 124 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cocksedge, S.P.; Infante, C.; Torres, S.; Lama, C.; Mantecón, L.; Manchado, M.; Acton, J.P.; Alsharif, N.S.; Clifford, T.; Wadley, A.J.; et al. Tetraselmis chuii Supplementation Increases Skeletal Muscle Nuclear Factor Erythroid 2-Related Factor 2 and Antioxidant Enzyme Gene Expression, and Peak Oxygen Uptake in Healthy Adults: A Randomised Crossover Trial. Antioxidants 2025, 14, 435. https://doi.org/10.3390/antiox14040435
Cocksedge SP, Infante C, Torres S, Lama C, Mantecón L, Manchado M, Acton JP, Alsharif NS, Clifford T, Wadley AJ, et al. Tetraselmis chuii Supplementation Increases Skeletal Muscle Nuclear Factor Erythroid 2-Related Factor 2 and Antioxidant Enzyme Gene Expression, and Peak Oxygen Uptake in Healthy Adults: A Randomised Crossover Trial. Antioxidants. 2025; 14(4):435. https://doi.org/10.3390/antiox14040435
Chicago/Turabian StyleCocksedge, Stuart P., Carlos Infante, Sonia Torres, Carmen Lama, Lalia Mantecón, Manuel Manchado, Jarred P. Acton, Nehal S. Alsharif, Tom Clifford, Alex J. Wadley, and et al. 2025. "Tetraselmis chuii Supplementation Increases Skeletal Muscle Nuclear Factor Erythroid 2-Related Factor 2 and Antioxidant Enzyme Gene Expression, and Peak Oxygen Uptake in Healthy Adults: A Randomised Crossover Trial" Antioxidants 14, no. 4: 435. https://doi.org/10.3390/antiox14040435
APA StyleCocksedge, S. P., Infante, C., Torres, S., Lama, C., Mantecón, L., Manchado, M., Acton, J. P., Alsharif, N. S., Clifford, T., Wadley, A. J., Ferguson, R. A., Bishop, N. C., Martin, N. R. W., & Bailey, S. J. (2025). Tetraselmis chuii Supplementation Increases Skeletal Muscle Nuclear Factor Erythroid 2-Related Factor 2 and Antioxidant Enzyme Gene Expression, and Peak Oxygen Uptake in Healthy Adults: A Randomised Crossover Trial. Antioxidants, 14(4), 435. https://doi.org/10.3390/antiox14040435