

Evaluation of Bioactive Effects of Five Plant Extracts with Different Phenolic Compositions against Different Therapeutic Targets

 ,
, 
 , , ,
, , ,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
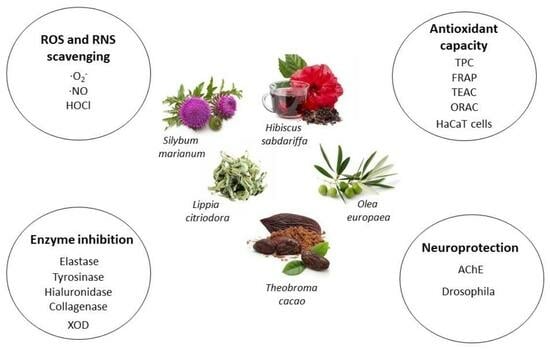
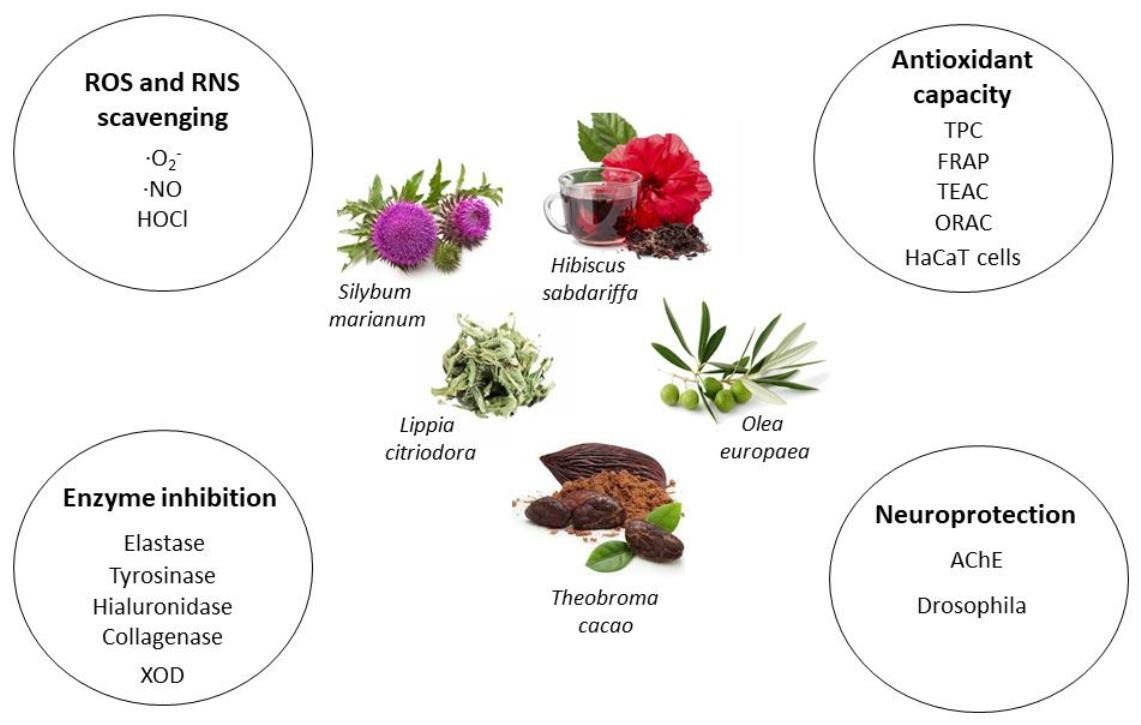
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Extraction Procedure from Plant Matrices
2.3. HPLC-ESI-TOF-MS Analysis
2.4. In Vitro Assays for Bioactive Determination of Phenolic Compounds in Extracts
2.4.1. Total Phenolic Content and Antioxidant Capacity Measurements
2.4.2. Evaluation of Free Radical and ROS/RNS Scavenging Potential
2.4.3. Evaluation of Enzymatic Inhibition Potential
2.4.4. Cellular Assays to Measure Antioxidant Capacity
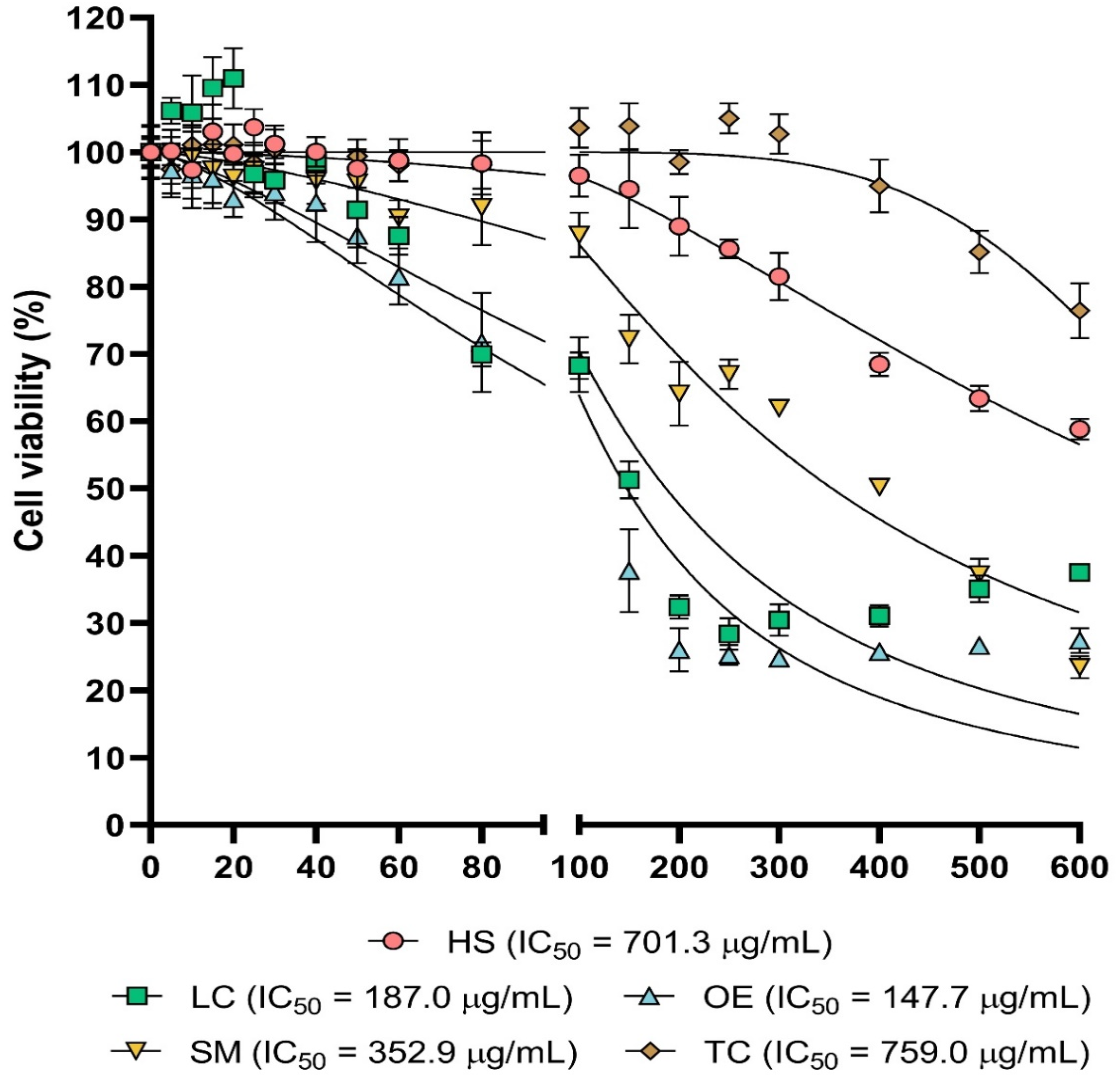
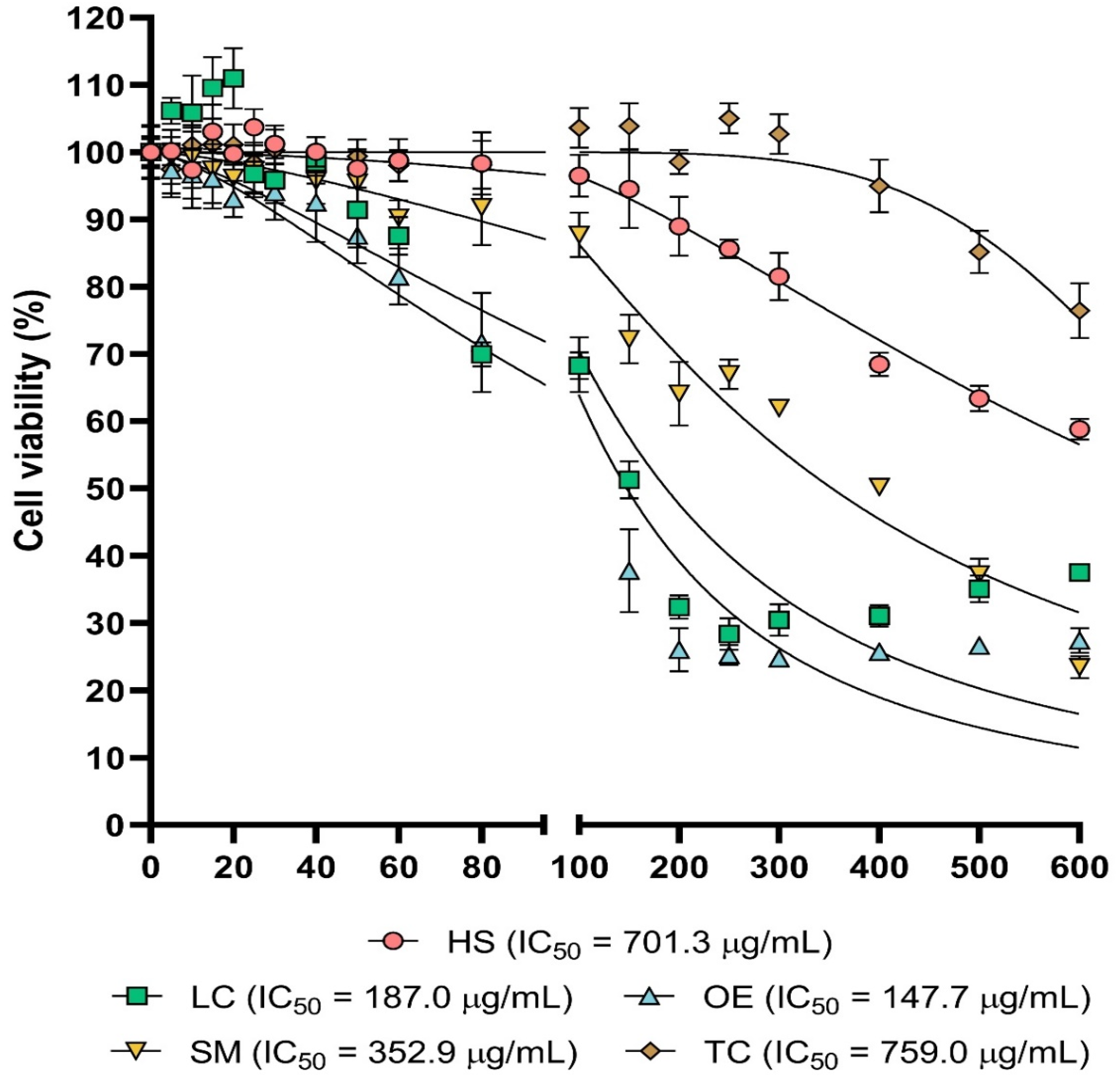
Cytotoxic Activity In Vitro
Antioxidant Activity In Vitro
2.5. In Vivo Neuroprotection Capacity
2.5.1. Drosophila Stock and Exposure to Hypoxia
2.5.2. Drosophila Locomotor Activity Monitoring
2.6. Statistical Analysis
3. Results
3.1. Characterisation of the Extracts by HPLC-ESI-qTOF-MS
3.2. Evaluation of the Antioxidant and Anti-Inflammatory Capacities of the Extracts
3.2.1. Evaluation of TPC, Antioxidant Capacity and ROS Scavenging Potential
3.2.2. Evaluation of Enzymatic Inhibition Capacity
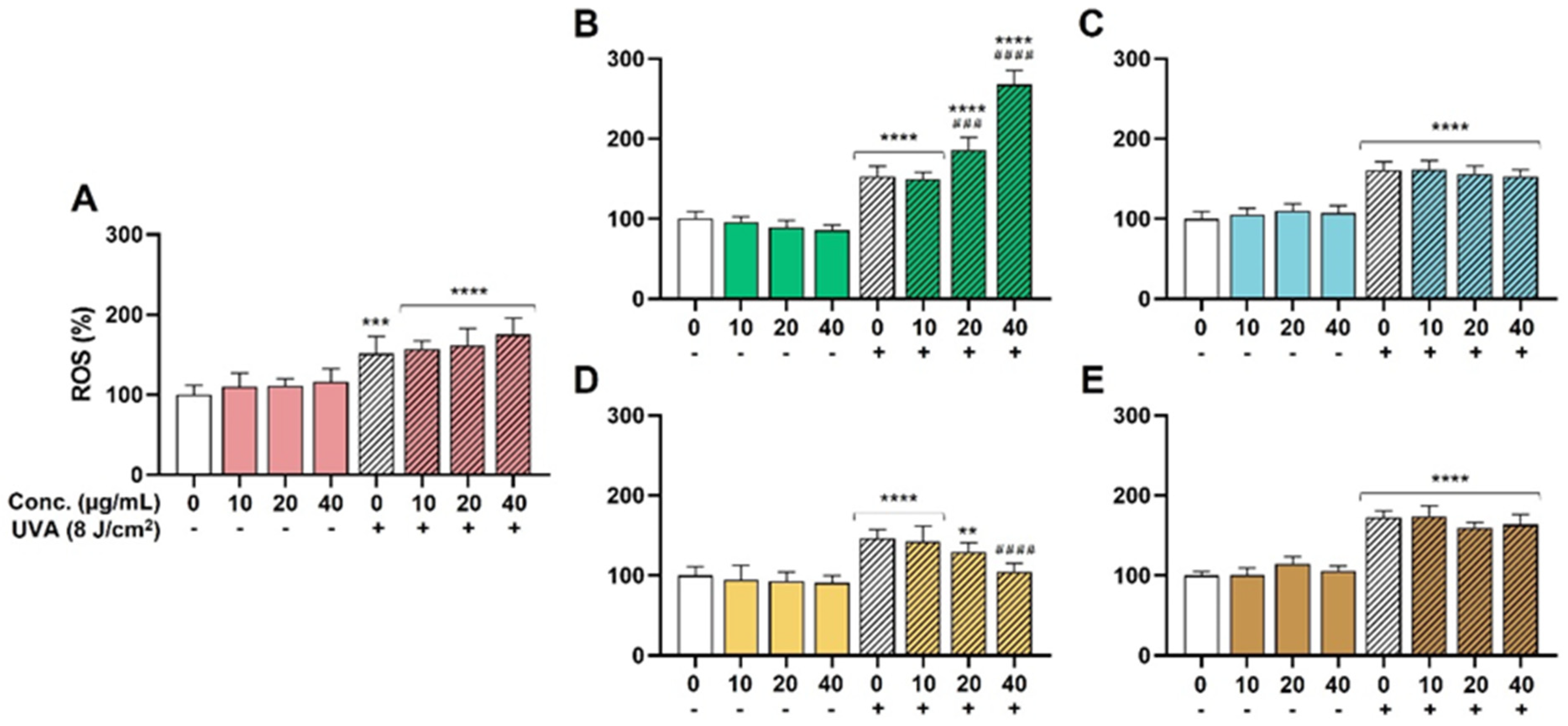
3.2.3. Cellular Assays to Measure Antioxidant Capacity
3.3. Evaluation of the Neuroprotective Effect of the Extracts
3.3.1. Evaluation of Acetylcholinesterase (AChE) Inhibition Capacity
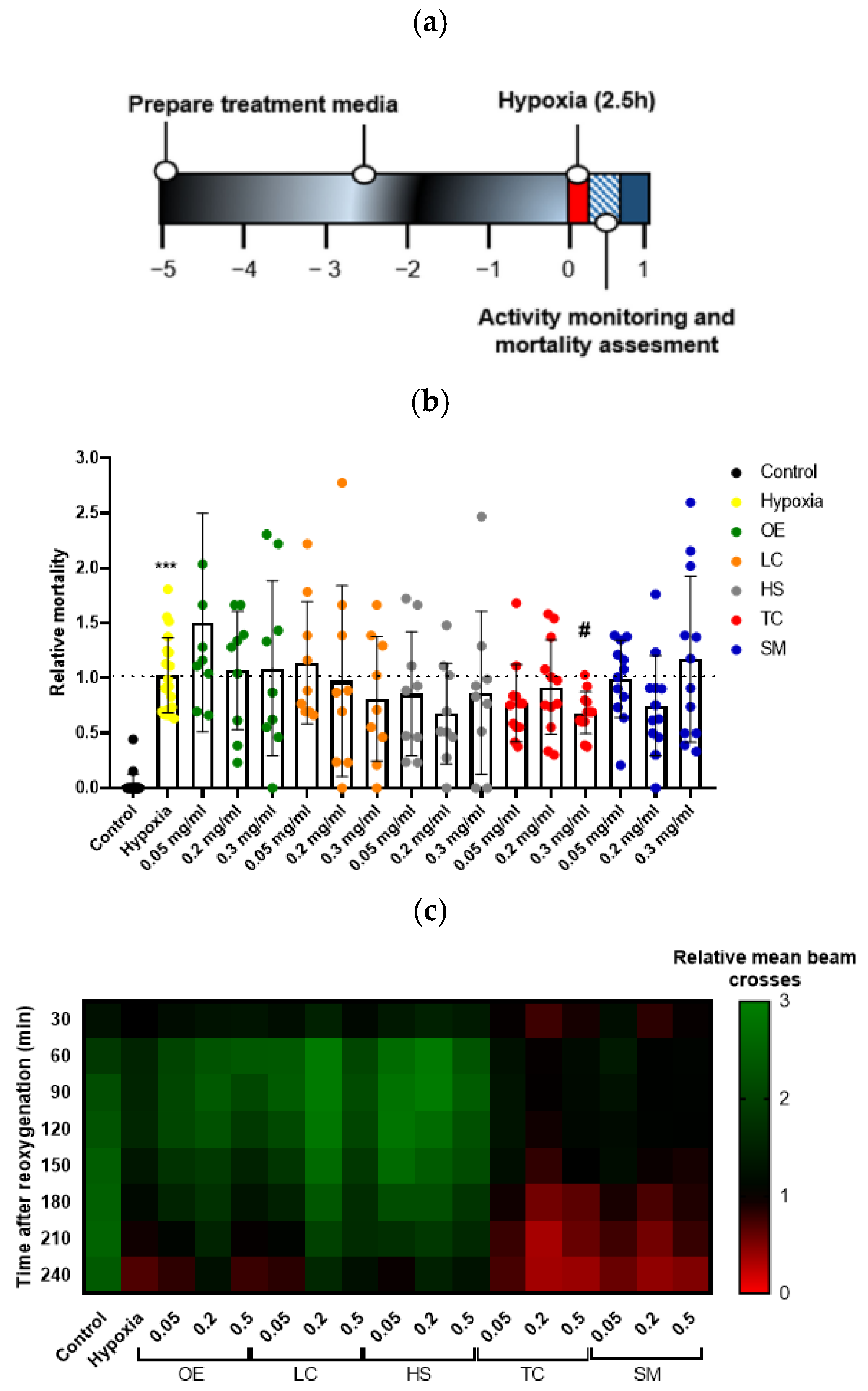
3.3.2. Effect of Supplementation with Different Plant Extracts on Hypoxia–Reoxygenation Injury in D. melanogaster
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ambra, R.; Pastore, G.; Lucchetti, S. The Role of Bioactive Phenolic Compounds on the Impact of Beer on Health. Molecules 2021, 26, 486. [Google Scholar] [CrossRef]
- Perez-Gregorio, R.; Simal-Gandara, J. A Critical Review of Bioactive Food Components, and of Their Functional Mechanisms, Biological Effects and Health Outcomes. Curr. Pharm. Des. 2017, 23, 2731–2741. [Google Scholar] [CrossRef]
- Vuolo, M.M.; Lima, V.S.; Maróstica Junior, M.R. Phenolic Compounds. In Bioactive Compounds; Elsevier: Amsterdam, The Netherlands, 2019; pp. 33–50. ISBN 9780128147757. [Google Scholar]
- Wojdylo, A.; Oszmianski, J.; Czemerys, R. Antioxidant Activity and Phenolic Compounds in 32 Selected Herbs. Food Chem. 2007, 105, 940–949. [Google Scholar] [CrossRef]
- Brahmi, F.; Nury, T.; Debbabi, M.; Hadj-Ahmed, S.; Zarrouk, A.; Prost, M.; Madani, K.; Boulekbache-Makhlouf, L.; Lizard, G. Evaluation of Antioxidant, Anti-Inflammatory and Cytoprotective Properties of Ethanolic Mint Extracts from Algeria on 7-Ketocholesterol-Treated Murine RAW 264.7 Macrophages. Antioxidants 2018, 7, 184. [Google Scholar] [CrossRef]
- Ambriz-Pérez, D.L.; Leyva-López, N.; Gutierrez-Grijalva, E.P.; Heredia, J.B. Phenolic Compounds: Natural Alternative in Inflammation Treatment. A Review. Cogent. Food Agric. 2016, 2, 1131412. [Google Scholar] [CrossRef]
- Luo, J.; Si, H.; Jia, Z.; Liu, D. Dietary Anti-Aging Polyphenols and Potential Mechanisms. Antioxidants 2021, 10, 283. [Google Scholar] [CrossRef]
- Kovacsova, M.; Barta, A.; Parohova, J.; Vrankova, S.; Pechanova, O. Neuroprotective Mechanisms of Natural Polyphenolic Compounds. Act. Nerv. Super. Rediviva 2010, 52, 181–186. [Google Scholar]
- Cai, Y.; Luo, Q.; Sun, M.; Corke, H. Antioxidant Activity and Phenolic Compounds of 112 Traditional Chinese Medicinal Plants Associated with Anticancer. Life Sci. 2004, 74, 2157–2184. [Google Scholar] [CrossRef] [PubMed]
- Coman, V.; Teleky, B.E.; Mitrea, L.; Martău, G.A.; Szabo, K.; Călinoiu, L.F.; Vodnar, D.C. Bioactive Potential of Fruit and Vegetable Wastes. In Advances in Food and Nutrition Research; Academic Press Inc.: Cambridge, MA, USA, 2020; Volume 91, pp. 157–225. ISBN 9780128204702. [Google Scholar]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural Products in Drug Discovery: Advances and Opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Gao, D.; Stoika, R.; Liu, K.; Sik, A.; Jin, M. Potential Implications of Polyphenolic Compounds in Neurodegenerative Diseases. Crit. Rev. Food Sci. Nutr. 2022, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yu, Z.; Xia, J.; Zhang, X.; Liu, K.; Sik, A.; Jin, M. Anti-Parkinson’s Disease Activity of Phenolic Acids from: Eucommia ulmoides Oliver Leaf Extracts and Their Autophagy Activation Mechanism. Food Funct. 2020, 11, 1425–1440. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Gao, D.; Paudel, Y.N.; Li, X.; Zheng, M.; Liu, G.; Ma, Y.; Chu, L.; He, F.; Jin, M. Anti-Parkinson’s Disease Activity of Sanghuangprous vaninii Extracts in the MPTP-Induced Zebrafish Model. ACS Chem. Neurosci. 2022, 13, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Hu, M.-J.; Wang, Y.-Q.; Cui, Y.-L. Antioxidant Activities of Quercetin and Its Complexes for Medicinal Application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Zhang, G.; Hu, X.; Xu, X.; Gong, D. Quercetin as a Tyrosinase Inhibitor: Inhibitory Activity, Conformational Change and Mechanism. Food Res. Int. 2017, 100, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Jan, R.; Khan, M.; Asaf, S.; Lubna; Asif, S.; Kim, K.-M. Bioactivity and Therapeutic Potential of Kaempferol and Quercetin: New Insights for Plant and Human Health. Plants 2022, 11, 2623. [Google Scholar] [CrossRef] [PubMed]
- Villegas-Aguilar, M.d.C.; Leyva-Jiménez, F.J.; Cádiz-Gurrea, M.d.l.L.; Segura-Carretero, A.; Arráez-Román, D. Comprehensive Analysis of Antioxidant Compounds from Lippia citriodora and Hibiscus sabdariffa Green Extracts Attained by Response Surface Methodology. Antioxidants 2020, 9, 1175. [Google Scholar] [CrossRef] [PubMed]
- Blainski, A.; Lopes, G.; de Mello, J. Application and Analysis of the Folin ciocalteu Method for the Determination of the Total Phenolic Content from Limonium brasiliense L. Molecules 2013, 18, 6852–6865. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Zulueta, A.; Esteve, M.J.; Frígola, A. ORAC and TEAC Assays Comparison to Measure the Antioxidant Capacity of Food Products. Food Chem. 2009, 114, 310–316. [Google Scholar] [CrossRef]
- Rojas-García, A.; Fuentes, E.; Cádiz-Gurrea, M.d.l.L.; Rodriguez, L.; Villegas-Aguilar, M.D.C.; Palomo, I.; Arráez-Román, D.; Segura-Carretero, A. Biological Evaluation of Avocado Residues as a Potential Source of Bioactive Compounds. Antioxidants 2022, 11, 1049. [Google Scholar] [CrossRef]
- Nema, N.K.; Maity, N.; Sarkar, B.K.; Mukherjee, P.K. Matrix Metalloproteinase, Hyaluronidase and Elastase Inhibitory Potential of Standardized Extract of Centella asiatica. Pharm. Biol. 2013, 51, 1182–1187. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.P.; Mandal, B.B. Inhibitory Role of Silk Cocoon Extract against Elastase, Hyaluronidase and UV Radiation-Induced Matrix Metalloproteinase Expression in Human Dermal Fibroblasts and Keratinocytes. Photochem. Photobiol. Sci. 2019, 18, 1259–1274. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A New and Rapid Colorimetric Determination of Acetylcholinesterase Activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Cooper, K.L.; Liu, K.J.; Hudson, L.G. Enhanced ROS Production and Redox Signaling with Combined arsenite and UVA Exposure: Contribution of NADPH Oxidase. Free Radic. Biol. Med. 2009, 47, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Habib, P.; Jung, J.; Wilms, G.M.; Kokott-Vuong, A.; Habib, S.; Schulz, J.B.; Voigt, A. Posthypoxic Behavioral Impairment and Mortality of Drosophila Melanogaster Are Associated with High Temperatures, Enhanced Predeath Activity and Oxidative Stress. Exp. Mol. Med. 2021, 53, 264–280. [Google Scholar] [CrossRef] [PubMed]
- Vigne, P.; Tauc, M.; Frelin, C. Strong Dietary Restrictions Protect Drosophila against Anoxia/Reoxygenation Injuries. PLoS ONE 2009, 4, e5422. [Google Scholar] [CrossRef]
- García-Rodríguez, P.; Ma, F.; del Río, C.; Romero-Bernal, M.; Najar, A.M.; Cádiz-Gurrea, M.d.l.L.; Leyva-Jimenez, F.J.; Ramiro, L.; Menéndez-Valladares, P.; Pérez-Sánchez, S.; et al. Diet Supplementation with Polyphenol-Rich Salicornia Ramosissima Extracts Protects against Tissue Damage in Experimental Models of Cerebral Ischemia. Nutrients 2022, 14, 5077. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed Minimum Reporting Standards for Chemical Analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef]
- Granato, D.; Shahidi, F.; Wrolstad, R.; Kilmartin, P.; Melton, L.D.; Hidalgo, F.J.; Miyashita, K.; van Camp, J.; Alasalvar, C.; Ismail, A.B.; et al. Antioxidant Activity, Total Phenolics and Flavonoids Contents: Should We Ban in Vitro Screening Methods? Food Chem. 2018, 264, 471–475. [Google Scholar] [CrossRef]
- Morales-Soto, A.; García-Salas, P.; Rodríguez-Pérez, C.; Jiménez-Sánchez, C.; Cádiz-Gurrea, M.d.l.L.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Antioxidant Capacity of 44 Cultivars of Fruits and Vegetables Grown in Andalusia (Spain). Food Res. Int. 2014, 58, 35–46. [Google Scholar] [CrossRef]
- Fischer, B.; Voynow, J.; Ghio, A. COPD: Balancing Oxidants and Antioxidants. Int. J. Chron. Obstruct. Pulmon. Dis. 2015, 10, 261–276. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative Stress, Inflammation, and Cancer: How Are They Linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhang, H.; Li, F.; Yang, F. Evaluation of Xanthine Oxidase Inhibitory Activity of Flavonoids by an Online Capillary Electrophoresis-based Immobilized Enzyme Microreactor. Electrophoresis 2020, 41, 1326–1332. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, Y.; Zhao, Z.; Qiu, J. Oxidative Stress in the Skin: Impact and Related Protection. Int. J. Cosmet. Sci. 2021, 43, 495–509. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Xu, W.; Meng, S.; Lim, N.K.H.; Wang, W.; Huang, F.-D. An Efficient and Reliable Assay for Investigating the Effects of Hypoxia/Anoxia on Drosophila. Neurosci. Bull. 2018, 34, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Aroso, I.M.; Araújo, A.R.; Fernandes, J.P.; Santos, T.; Batista, M.T.; Pires, R.A.; Mano, J.F.; Reis, R.L. Hydroalcoholic Extracts from the Bark of Quercus suber L. (Cork): Optimization of Extraction Conditions, Chemical Composition and Antioxidant Potential. Wood Sci. Technol. 2017, 51, 855–872. [Google Scholar] [CrossRef]
- Xu, J.Z.; Yeung, S.Y.V.; Chang, Q.; Huang, Y.; Chen, Z.-Y. Comparison of Antioxidant Activity and Bioavailability of Tea Epicatechins with Their Epimers. Br. J. Nutr. 2004, 91, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Viktorova, J.; Stranska-Zachariasova, M.; Fenclova, M.; Vitek, L.; Hajslova, J.; Kren, V.; Ruml, T. Complex Evaluation of Antioxidant Capacity of Milk Thistle Dietary Supplements. Antioxidants 2019, 8, 317. [Google Scholar] [CrossRef]
- Aldini, G.; Piccoli, A.; Beretta, G.; Morazzoni, P.; Riva, A.; Marinello, C.; Maffei Facino, R. Antioxidant Activity of Polyphenols from Solid Olive Residues of c.v. Coratina. Fitoterapia 2006, 77, 121–128. [Google Scholar] [CrossRef]
- Svobodová, A.; Zdařilová, A.; Walterová, D.; Vostálová, J. Flavonolignans from Silybum marianum Moderate UVA-Induced Oxidative Damage to HaCaT Keratinocytes. J. Dermatol. Sci. 2007, 48, 213–224. [Google Scholar] [CrossRef]
- Xi, Y.; Jiao, W.; Cao, J.; Jiang, W. Effects of Chlorogenic Acid on Capacity of Free Radicals Scavenging and Proteomic Changes in Postharvest Fruit of Nectarine. PLoS ONE 2017, 12, e0182494. [Google Scholar] [CrossRef]
- Suh, H.J.; Lee, J.M.; Cho, J.S.; Kim, Y.S.; Chung, S.H. Radical Scavenging Compounds in Onion Skin. Food Res. Int. 1999, 32, 659–664. [Google Scholar] [CrossRef]
- Furuno, K.; Akasako, T.; Sugihara, N. The Contribution of the Pyrogallol Moiety to the Superoxide Radical Scavenging Activity of Flavonoids. Biol. Pharm. Bull. 2002, 25, 19–23. [Google Scholar] [CrossRef]
- Ebuehi, O.A.T.; Anams, C.; Gbenle, O.D.; Ajagun-Ogunleye, M.O. Hydro-ethanol Seed Extract of Theobroma Cacao Exhibits Antioxidant Activities and Potential Anticancer Property. J. Food Biochem. 2019, 43, e12767. [Google Scholar] [CrossRef]
- He, J.; Xu, L.; Yang, L.; Wang, X. Epigallocatechin Gallate Is the Most Effective Catechin Against Antioxidant Stress via Hydrogen Peroxide and Radical Scavenging Activity. Med. Sci. Monit. 2018, 24, 8198–8206. [Google Scholar] [CrossRef] [PubMed]
- Surai, P. Silymarin as a Natural Antioxidant: An Overview of the Current Evidence and Perspectives. Antioxidants 2015, 4, 204–247. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Zou, B.; Zeng, H.; Zhang, L.; Chen, M.; Fu, G. Inhibitory Effect of Verbascoside on Xanthine Oxidase Activity. Int. J. Biol. Macromol. 2016, 93, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Zillich, O.V.; Schweiggert-Weisz, U.; Eisner, P.; Kerscher, M. Polyphenols as Active Ingredients for Cosmetic Products. Int. J. Cosmet. Sci. 2015, 37, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Vostálová, J.; Tinková, E.; Biedermann, D.; Kosina, P.; Ulrichová, J.; Rajnochová Svobodová, A. Skin Protective Activity of Silymarin and Its Flavonolignans. Molecules 2019, 24, 1022. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-Z.; Ren, Z.; Reddy, N.V.; Hou, T.; Zhang, Z.-J. In Silico Evaluation of Antimicrobial, Antihyaluronidase and Bioavailability Parameters of Rosmarinic Acid in Perilla Frutescens Leaf Extracts. SN Appl. Sci. 2020, 2, 1547. [Google Scholar] [CrossRef]
- García, G.A.; Hernández, S.V.; Mejía, Ó.R.; Baez, S.A.; Ananías García, Y.C. Biología y Patobiología Humana Del Ácido Hialurónico En La Estabilización de La Matriz Extracelular y La Inflamación. Revista Med. 2006, 14, 80–87. [Google Scholar]
- Eun Lee, K.; Bharadwaj, S.; Yadava, U.; Gu Kang, S. Evaluation of Caffeine as Inhibitor against Collagenase, Elastase and Tyrosinase Using in Silico and in Vitro Approach. J. Enzym. Inhib. Med. Chem. 2019, 34, 927–936. [Google Scholar] [CrossRef]
- Jiang, Y.; Gao, H.; Turdu, G. Traditional Chinese Medicinal Herbs as Potential AChE Inhibitors for Anti-Alzheimer’s Disease: A Review. Bioorg. Chem. 2017, 75, 50–61. [Google Scholar] [CrossRef]
- Zhang, L.; Cao, H.; Wen, J.; Xu, M. Green Tea Polyphenol (–)-Epigallocatechin-3-Gallate Enhances the Inhibitory Effect of Huperzine A on Acetylcholinesterase by Increasing the Affinity with Serum Albumin. Nutr. Neurosci. 2009, 12, 142–148. [Google Scholar] [CrossRef]
- Mottaghipisheh, J.; Taghrir, H.; Boveiri Dehsheikh, A.; Zomorodian, K.; Irajie, C.; Mahmoodi Sourestani, M.; Iraji, A. Linarin, a Glycosylated Flavonoid, with Potential Therapeutic Attributes: A Comprehensive Review. Pharmaceuticals 2021, 14, 1104. [Google Scholar] [CrossRef]
- Serrano, A.; Ros, G.; Nieto, G. Bioactive Compounds and Extracts from Traditional Herbs and Their Potential Anti-Inflammatory Health Effects. Medicines 2018, 5, 76. [Google Scholar] [CrossRef] [PubMed]
- Villegas-Aguilar, M.d.C.; Fernández-Ochoa, Á.; Cádiz-Gurrea, M.d.l.L.; Pimentel-Moral, S.; Lozano-Sánchez, J.; Arráez-Román, D.; Segura-Carretero, A. Pleiotropic Biological Effects of Dietary Phenolic Compounds and Their Metabolites on Energy Metabolism, Inflammation and Aging. Molecules 2020, 25, 596. [Google Scholar] [CrossRef] [PubMed]
- García-Villegas, A.; Rojas-García, A.; Villegas-Aguilar, M.d.C.; Fernández-Moreno, P.; Fernández-Ochoa, Á.; Cádiz-Gurrea, M.d.l.L.; Arráez-Román, D.; Segura-Carretero, A. Cosmeceutical Potential of Major Tropical and Subtropical Fruit By-Products for a Sustainable Revalorization. Antioxidants 2022, 11, 203. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Rt (min) | [M-H]− | Mol. Formula | T. cacao | H. sabdariffa | S. marianum | L. citriodora | O. europaea |
|---|---|---|---|---|---|---|---|---|
| Gluconic acid | 1.01 | 195.0519 | C6H12O7 | x | x | x | ||
| Malic acid | 1.07 | 133.0140 | C4H6O5 | x | x | |||
| Quinic acid | 1.13 | 191.0292 | C7H12O6 | x | x | |||
| Chlorogenic acid | 4.02 | 353.0867 | C16H18O9 | x | x | |||
| Epigallocatechin | 9.18 | 305.0690 | C15H14O7 | x | x | |||
| Verbascoside | 9.33 | 623.1978 | C29H36O15 | x | x | |||
| Quercetin 3-O-rutinoside | 9.60 | 609.1458 | C27H30O16 | x | x | |||
| Quercetin glucoside | 9.85 | 463.0878 | C21H20O12 | x | x | x | ||
| Unknown | 10.23 | 539.1751 | C25H32O13 | x | x | |||
| Quercetin | 12.16 | 301.0339 | C15H10O7 | x | x | |||
| Unknown | 12.95 | 329.0654 | C17H14O7 | x | x | |||
| Dihydrocapsiate | 13.53 | 307.192 | C18H28O4 | x | x | |||
| Eupatorin | 13.56 | 343.0818 | C18H16O7 | x | x | |||
| Gingerol | 13.92 | 293.1748 | C17H26O4 | x | x | x | x | |
| Hydroxylinolenic acid | 15.74 | 293.2113 | C18H30O3 | x | x | x | ||
| Unknown | 16.30 | 540.3288 | C29H49O9 | x | x | x | x | |
| Unknown | 16.60 | 566.3453 | C31H51O9 | x | x | x | ||
| Ethyl vanillate | 17.77 | 195.0686 | C10H12O4 | x | x | |||
| Linolenic acid | 18.46 | 277.2159 | C18H30O2 | x | x | x | x | |
| 10′-Apo-beta-carotenal | 18.73 | 375.2712 | C27H36O | x | x | x | x | x |
| Linoleic acid | 19.16 | 279.2328 | C18H32O2 | x | x | x | x | x |
| Palmitic acid | 19.82 | 255.2325 | C16H32O2 | x | x | x | ||
| Unkwnon | 19.87 | 403.3052 | C22H44O6 | x | x | x | x | |
| Oleic Acid | 19.95 | 281.2482 | C18H34O2 | x | x | x | x | x |
| Unknown | 20.98 | 383.1934 | C16H32O10 | x | x | x | x | |
| Ginsenoside Rh2 | 20.98 | 621.4417 | C36H62O8 | x | x |
| Sample | TPC (mg GAE/g DE) | FRAP (mmol Fe2+/g DE) | TEAC (mmol TE/g DE) | ORAC (mmol TE/g DE) |
|---|---|---|---|---|
| T. cacao | 255 ± 12 a | 1.38 ± 0.09 b,c | 1.25 ± 0.08 a | 2.35 ± 0.09 c |
| H. sabdariffa | 51 ± 1 d | 0.47 ± 0.05 d | 0.24 ± 0.02 c | 1.16 ± 0.03 d |
| S. marianum | 617 ± 8 b | 1.4 ± 0.3 a,b,c,d | 1.3 ± 0.1 a | 11.7 ± 0.3 b |
| L. citriodora | 344 ± 15 c | 2.4 ± 0.2 a | 1.15 ± 0.08 a | 5.9 ± 0.2 a |
| O. europaea | 216 ± 28 a | 1.65 ± 0.10 a,b | 0.84 ± 0.05 b | 6.0 ± 0.2 a |
| Sample | ·O2− (mg/L) 1 | ·NO (mg/L) 1 | HOCl (mg/L) 1 |
|---|---|---|---|
| T. cacao | 29.7 ± 0.4 c | 0.42 ± 0.02 b | 0.71 ± 0.02 b |
| H. sabdariffa | 50 ± 2 a | 10.3 ± 1.0 c | 1.32 ± 0.04 d |
| S. marianum | 57 ± 6 a | 5.0 ± 0.8 a | 0.70 ± 0.01 b |
| L. citriodora | n.d. | 3.76 ± 0.08 a | 3.5 ± 0.4 a |
| O. europaea | 269 ± 17 d | 3.0 ± 0.2 a | 16 ± 2 c |
| Gallic acid | 50 ± 3 a | 1.4 ± 0.3 b | 3.8 ± 0.3 a |
| Epicatechin | 70 ± 5 b | 0.87 ± 0.02 b | 0.18 ± 0.01 e |
| Sample | Hyaluronidase (mg/L) 1 | XOD (mg/L) 1 | Tyrosinase (% inh.) 2 | Elastase (% inh.) 3 | Collagenase (mg/L) 1 | AChE (mg/L) 1 |
|---|---|---|---|---|---|---|
| T. cacao | 12 ± 2 a | n.d. | 28 ± 8 a,b,c | 23 ± 4 b,c,d | 156 ±1 b | 244 ± 9 a |
| H. sabdariffa | 66 ± 6 d | n.d. | 5 ± 2 c | 44.1 ± 0.5 a,b,c,d | 1190 ± 35 b | n.d. |
| S. marianum | 4.9 ± 0.4 a | 4.4 ± 0.4 a | 39 ± 4 a | n.d. | 56 ± 3 b | 1259 ± 53 b |
| L. citriodora | 87 ± 5 c | 3.2 ± 0.4 a,b | 17 ± 2 b,c | 48 ± 3 a | 633 ± 27 a | 316 ± 4 c |
| O. europaea | 187 ± 5 b | 2.3 ± 0.3 b | 28 ± 3 a,b | 25 ± 3 b,c,d | 618 ± 9 a | 373 ± 6 d |
| Gallic acid | 102 ± 4 4 | n.d. | n.d. | n.d. | n.d. | n.d. |
| Epicatechin | 167 ± 6 4 | 9 ± 1 c | n.d. | n.d. | n.d. | n.d. |
| Physostigmine | n.d. | n.d. | n.d. | n.d. | n.d. | 0.043 ± 0.004 6 |
| 1,10-phenanthroline | n.d. | n.d. | n.d. | n.d. | 83 ± 2 5 | n.d. |
| Elastatinal (51.26 ppm) | n.d. | n.d. | n.d. | 53 ± 5 | n.d. | n.d. |
| Kojic acid (21.3 ppm) | n.d. | n.d. | 49 ± 6 | n.d. | n.d. | n.d. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villegas-Aguilar, M.d.C.; Sánchez-Marzo, N.; Fernández-Ochoa, Á.; Del Río, C.; Montaner, J.; Micol, V.; Herranz-López, M.; Barrajón-Catalán, E.; Arráez-Román, D.; Cádiz-Gurrea, M.d.l.L.; et al. Evaluation of Bioactive Effects of Five Plant Extracts with Different Phenolic Compositions against Different Therapeutic Targets. Antioxidants 2024, 13, 217. https://doi.org/10.3390/antiox13020217
Villegas-Aguilar MdC, Sánchez-Marzo N, Fernández-Ochoa Á, Del Río C, Montaner J, Micol V, Herranz-López M, Barrajón-Catalán E, Arráez-Román D, Cádiz-Gurrea MdlL, et al. Evaluation of Bioactive Effects of Five Plant Extracts with Different Phenolic Compositions against Different Therapeutic Targets. Antioxidants. 2024; 13(2):217. https://doi.org/10.3390/antiox13020217
Chicago/Turabian StyleVillegas-Aguilar, María del Carmen, Noelia Sánchez-Marzo, Álvaro Fernández-Ochoa, Carmen Del Río, Joan Montaner, Vicente Micol, María Herranz-López, Enrique Barrajón-Catalán, David Arráez-Román, María de la Luz Cádiz-Gurrea, and et al. 2024. "Evaluation of Bioactive Effects of Five Plant Extracts with Different Phenolic Compositions against Different Therapeutic Targets" Antioxidants 13, no. 2: 217. https://doi.org/10.3390/antiox13020217
APA StyleVillegas-Aguilar, M. d. C., Sánchez-Marzo, N., Fernández-Ochoa, Á., Del Río, C., Montaner, J., Micol, V., Herranz-López, M., Barrajón-Catalán, E., Arráez-Román, D., Cádiz-Gurrea, M. d. l. L., & Segura-Carretero, A. (2024). Evaluation of Bioactive Effects of Five Plant Extracts with Different Phenolic Compositions against Different Therapeutic Targets. Antioxidants, 13(2), 217. https://doi.org/10.3390/antiox13020217