
Thylakoid Rhodanese-like Protein–Ferredoxin:NADP+ Oxidoreductase Interaction Is Integrated into Plant Redox Homeostasis System


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Chemicals
2.3. Chloroplast Isolation from A. thaliana
2.4. EPR Measurements
2.5. Tandem Affinity Purification and MS Analysis
2.6. Co-Immunoprecipitation
2.7. Plant Material and Growth Conditions for Glutathione Determination
2.8. Sample Preparation for Transmission Electron Microscopy and Immunogold Labeling
2.9. Cytohistochemical Determination of Glutathione
2.10. Quantitative Analysis of Immunogold Labeling
3. Results
3.1. TROL May Form Transient Complexes with the tAPX
3.2. TROL OX Arabidopsis Chloroplasts Accumulate Elevated Levels of O2•− during the Exposure to High Light
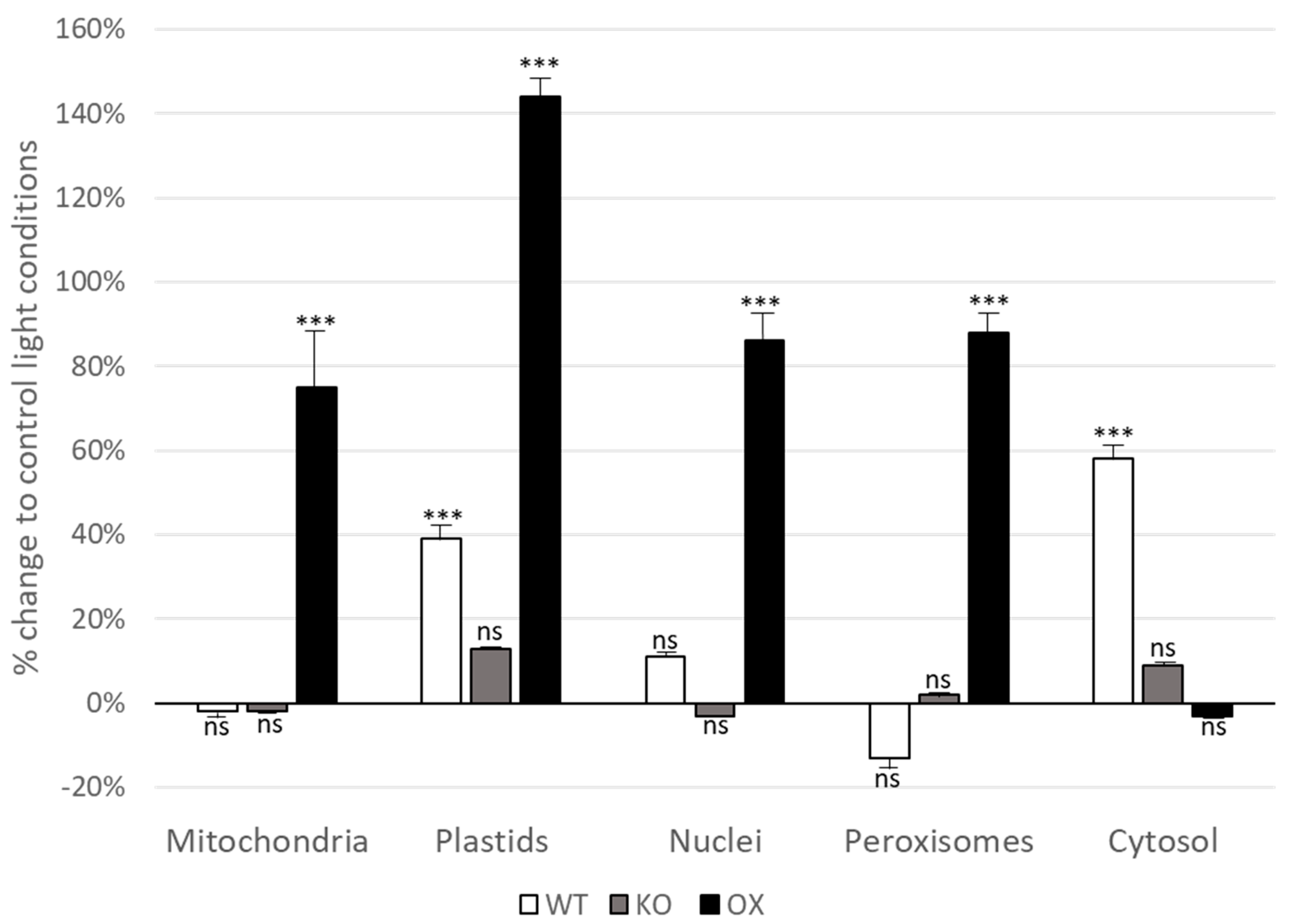
3.3. Glutathione Levels Are Elevated in All Cell Compartments in TROL OX Plants after Short-Term Light Stress
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TROL | thylakoid rhodanase-like protein |
| TROL OX | Arabidopsis line overproducing TROL protein |
| TROL KO | Arabidopsis line not producing TROL protein, trol |
| FNR | ferredoxin: NADP+ oxidoreductase |
| RHO | rhodanase-like domain of TROL |
| PEPE | Pro-Val-Pro repeat-rich region of TROL |
| ITEP | highly conserved module of TROL necessary for establishing high-affinity interaction with FNR |
| Tic62 | 62 kDa protein component of the translocon at the inner envelope of chloroplasts |
| tAPX | thylakoidal ascorbate peroxidase |
| SOD | superoxide dismutase |
| GSH | glutathione |
References
- Mulo, P. Chloroplast-targeted ferredoxin-NADP+ oxidoreductase (FNR): Structure, function and location. BBA Bioenerg. 2011, 1807, 927–934. [Google Scholar] [CrossRef]
- Jurić, S.; Hazler-Pilepić, K.; Tomašić, A.; Lepeduš, H.; Jeličić, B.; Puthiyaveetil, S.; Bionda, T.; Vojta, L.; Allen, J.F.; Schleiff, E.; et al. Tethering of ferredoxin: NADP + oxidoreductase to thylakoid membranes is mediated by novel chloroplast protein TROL. Plant J. 2009, 60, 783–794. [Google Scholar] [CrossRef]
- Vojta, L.; Fulgosi, H. Topology of TROL protein in thylakoid membranes of Arabidopsis thaliana. Physiol. Plantarum 2019, 166, 300–308. [Google Scholar] [CrossRef]
- Kekić, T.; Fulgosi, H.; Vojta, L.; Bertoša, B. Molecular basis of ferredoxin:NADP(+) reductase interactions with FNR binding domains from TROL and Tic62 proteins. J. Mol. Struct. 2020, 1215, 128281. [Google Scholar] [CrossRef]
- Vojta, L.; Fulgosi, H. Data supporting the absence of FNR dynamic photosynthetic membrane recruitment in trol mutants. Data Brief 2016, 7, 393–396. [Google Scholar] [CrossRef]
- Vojta, L.; Carić, D.; Cesar, V.; Antunović Dunić, J.; Lepeduš, H.; Kveder, M.; Fulgosi, H. TROL-FNR interaction reveals alternative pathways of electron partitioning in photosynthesis. Sci. Rep. 2015, 5, 10085. [Google Scholar] [CrossRef]
- Benz, J.P.; Lintala, M.; Soll, J.; Mulo, P.; Bölter, B. A new concept for ferredoxin-NADP(H) oxidoreductase binding to plant thylakoids. Trends Plant Sci. 2010, 15, 608–613. [Google Scholar] [CrossRef]
- Alte, F.; Stengel, A.; Benz, J.-P.; Petersen, E.; Soll, J.; Groll, M.; Bölter, B. Ferredoxin:NADPH oxidoreductase is recruited to thylakoids by binding to polyproline type II helix in a pH dependent manner. Proc. Natl. Acad. Sci. USA 2010, 107, 19260–19265. [Google Scholar] [CrossRef]
- Forti, G.; Bracale, M. Ferredoxin–ferredoxin NADP reductase interaction. FEBS Lett. 1984, 166, 81–84. [Google Scholar] [CrossRef]
- Hanke, G.T.; Endo, T.; Satoh, F.; Hase, T. Altered photosynthetic electron channelling into cyclic electron flow and nitrite assimilation in a mutant of ferredoxin: NADP(H) reductase. Plant Cell Environ. 2008, 31, 1017–1028. [Google Scholar] [CrossRef]
- Vojta, L.; Horvat, L.; Fulgosi, H. Balancing chloroplast redox status—Regulation of FNR binding and release. Period. Biol. 2012, 114, 25–31. [Google Scholar]
- Ogawa, K.; Kenematsu, K.; Takabe, K.; Asada, K. Attachment of CuZn superoxide dismutase to thylakoid membranes at the superoxide generation (PSI) in spinach chloroplasts: Detection by immuno-gold labeling after rapid freezing and substitution method. Plant Cell Physiol. 1995, 36, 565–573. [Google Scholar] [CrossRef][Green Version]
- Miyake, C.; Asada, K. Ferredoxin-Dependent Photoreduction of the Monodehydroascorbate Radical in Spinach Thylakoids. Plant Cell Physiol. 1994, 35, 539–549. [Google Scholar] [CrossRef]
- Miyake, C.; Asada, K. Thylakoid-Bound Ascorbate Peroxidase in Spinach Chloroplasts and Photoreduction of Its Primary Oxidation Product Monodehydroascorbate Radicals in Thylakoids. Plant Cell Physiol. 1992, 33, 541–553. [Google Scholar] [CrossRef]
- Miyake, C.; Cao, W.H.; Asada, K. Purification and molecular properties of thylakoid-bound ascorbate peroxidase from spinach chloroplasts. Plant Cell Physiol. 1993, 343, 881–889. [Google Scholar]
- Yoshimura, K.; Yabuta, Y.; Ishikawa, T.; Shigeoka, S. Identification of a cis element for tissue-specific alternative splicing of chloroplast ascorbate peroxidase pre-mRNA in higher plants. J. Biol. Chem. 2002, 277, 40623–40632. [Google Scholar] [CrossRef]
- Kangasjärvi, S.; Lepistö, A.; Hännikäinen, K.; Piippo, M.; Luomala, E.M.; Aro, E.M.; Rintamäki, E. Diverse roles for chloroplast stromal and thylakoid bound ascorbate peroxidases in plant stress responses. Biochem. J. 2008, 412, 275–285. [Google Scholar] [CrossRef]
- Asada, K. Production and Scavenging of Reactive Oxygen Species in Chloroplasts and Their Functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef]
- Kameoka, T.; Okayasub, T.; Kikuraku, K.; Ogawa, T.; Sawab, Y.; Yamamoto, H.; Ishikawa, T.; Maruta, T. Cooperation of chloroplast ascorbate peroxidases and proton gradient regulation 5 is critical for protecting Arabidopsis plants from photooxidative stress. Plant J. 2021, 107, 876–892. [Google Scholar] [CrossRef]
- Yabuta, Y.; Maruta, T.; Yoshimura, K.; Ishikawa, T.; Shigeoka, S. Two distinct redox signaling pathways for cytosolic APX induction under photooxidative stress. Plant Cell Physiol. 2004, 45, 1586–1594. [Google Scholar] [CrossRef]
- Exposito-Rodriguez, M.; Laissue, P.P.; Yvon-Durocher, G.; Smirnoff, N.; Mullineaux, P.M. Photosynthesis-dependent H2O2 transfer from chloroplasts to nuclei provides a high-light signalling mechanism. Nat. Commun. 2017, 8, 49. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A. The integration of glutathione homeostasis and redox signaling. J. Plant Physiol. 2009, 165, 1390–1403. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, A.; Hernandez, J.A.; Pastori, G.; del Rio, L.A.; Sevilla, F. Role of the Ascorbate-Glutathione Cycle of Mitochondria and Peroxisomes in the Senescence of Pea Leaves. Plant Physiol. 1998, 118, 1327–1335. [Google Scholar] [CrossRef] [PubMed]
- Rouhier, N.; Lemaire, S.D.; Jacquot, J.P. The role of glutathione in photosynthetic organisms: Emerging functions for glutaredoxins and glutathionylation. Annu. Rev. Plant Biol. 2008, 59, 143–166. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Mhamdi, A.; Chaouch, S.; Han, Y.; Neukermans, J.; Marquez-Garcia, B.; Queval, G.; Foyer, C.H. Glutathione in plants: An integrated overview. Plant Cell Environ. 2012, 35, 454–484. [Google Scholar] [CrossRef] [PubMed]
- Kovačević, J. Construction of Arabidopsis thaliana (Heynh.) Plants Containing the TROL Protein Labeled with HA and FLAG Protein Markers. Diploma Thesis, Faculty of Science, Zagreb, Croatia, 2013. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Zechmann, B.; Müller, M. Effects of ZYMV-infection on the subcellular distribution of glutathione and its precursors in a highly tolerant Cucurbita pepo cultivar. Botany 2008, 86, 1092–1100. [Google Scholar] [CrossRef]
- Bortz, J.; Lienert, G.A.; Boehnke, K. Verteilungsfreie Methoden in der Biostatistik; Springer: Berlin, Germany, 2000; ISBN 978-3-662-10786-7. [Google Scholar]
- De Gara, L.; Locato, V.; Dipierro, S.; DePinto, M.C. Redox homeostasis in plants. The challenge of living with endogenous oxygen production. Resp. Physiol. Neurobiol. 2010, 173S, S13–S19. [Google Scholar] [CrossRef]
- Heber, U.; French, C.S. Effects of oxygen on the transport chain of photosynthesis. Planta 1968, 79, 99–112. [Google Scholar] [CrossRef]
- McRae, D.G.; Thompson, J.E. Senescence-dependent changes in superoxide anion production by illuminated chloroplasts from bean leaves. Planta 1983, 158, 185–193. [Google Scholar] [CrossRef]
- Takahashi, Y.; Katoh, S. Triplet state in photosystem I reaction center complex. Inhibition of radical pair recombination by bipyridinium dyes and naphtoquinones. Plant Cell Physiol. 1984, 25, 785–794. [Google Scholar]
- Goss, T.; Hanke, G. The end of the line: Can ferredoxin and ferredoxin NADP(H) oxidoreductase determine the fate of photosynthetic electrons? Curr. Prot. Pept. Sci. 2014, 15, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef] [PubMed]
- Raven, E.L. Understanding functional diversity and substrate specificity in haem peroxidases: What can we learn from ascorbate peroxidase? Nat. Prod. Rep. 2003, 20, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Fartyal, D.; Agarwal, A.; Shukla, T.; James, D.; Kaul, T.; Negi, Y.K.; Arora, S.; Reddy, M.K. Abiotic Stress Tolerance in Plants: Myriad Roles of Ascorbate Peroxidase. Front. Plant Sci. 2017, 8, 581. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Halliwell, B. The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism. Planta 1976, 133, 21–25. [Google Scholar] [CrossRef]
- Foyer, C.H.; Halliwell, B. Purification and properties of dehydroascorbate reductase from spinach leaves. Phytochemistry 1977, 16, 1347–1350. [Google Scholar] [CrossRef]
- Mittler, R.; Zilinskas, B.A. Purification and characterization of pea cytosolic ascorbate peroxidase. Plant Physiol. 1991, 97, 962–968. [Google Scholar] [CrossRef]
- Jimenez, A.; Hernandez, J. Evidence for the presence of the ascorbate-glutathione cycle in mitochondria and peroxisomes of pea leaves. Plant Physiol. 1997, 114, 275–284. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Protein | Accession Number | MW | Organism |
|---|---|---|---|
| Rhodanese-like domain-containing protein 4, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = STR4 PE = 1 SV = 2 | sp|Q9M158|STR4_ARATH | 49 kDa | Arabidopsis thaliana |
| Chlorophyll a-b binding protein CP26, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = LHCB5 PE = 1 SV = 1 | sp|Q9XF89|CB5_ARATH (+1) | 30 kDa | Arabidopsis thaliana |
| Ferredoxin--NADP reductase, leaf isozyme 1, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = LFNR1 PE = 1 SV = 1 | sp|Q9FKW6|FNRL1_ARATH (+1) | 40 kDa | Arabidopsis thaliana |
| Ferredoxin--NADP reductase, leaf isozyme 2, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = LFNR2 PE = 1 SV = 1 | sp|Q8W493|FNRL2_ARATH (+1) | 41 kDa | Arabidopsis thaliana |
| Lipoxygenase 2, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = LOX2 PE = 1 SV = 1 | sp|P38418|LOX2_ARATH | 102 kDa | Arabidopsis thaliana |
| PSAF OS = Arabidopsis thaliana OX = 3702 GN = AXX17_At1g31980 PE = 4 SV = 1 | tr|A0A178WB32|A0A178WB32_ARATH | 24 kDa | Arabidopsis thaliana |
| Chlorophyll a-b binding protein 1, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = LHCB1.3 PE = 1 SV = 1 | sp|P04778|CB1C_ARATH (+5) | 28 kDa | Arabidopsis thaliana |
| Chlorophyll a-b binding protein CP29.1, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = LHCB4.1 PE = 1 SV = 1 | sp|Q07473|CB4A_ARATH (+2) | 31 kDa | Arabidopsis thaliana |
| L-ascorbate peroxidase T, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = APXT PE = 2 SV = 2 | sp|Q42593|APXT_ARATH (+1) | 46 kDa | Arabidopsis thaliana |
| Photosystem II D2 protein OS = Arabidopsis thaliana OX = 3702 GN = psbD PE = 1 SV = 3 | sp|P56761|PSBD_ARATH (+1) | 40 kDa | Arabidopsis thaliana |
| Chlorophyll a-b binding protein 4, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = LHCA4 PE = 1 SV = 1 | sp|P27521|CA4_ARATH (+1) | 28 kDa | Arabidopsis thaliana |
| Protein plastid transcriptionally active 16, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = PTAC16 PE = 1 SV = 1 | sp|Q9STF2|PTA16_ARATH (+1) | 54 kDa | Arabidopsis thaliana |
| Uncharacterized protein OS = Arabidopsis thaliana OX = 3702 GN = AXX17_At3g54180 PE = 4 SV = 1 | tr|A0A178VC12|A0A178VC12_ARATH (+1) | 74 kDa | unknown |
| CaS OS = Arabidopsis thaliana OX = 3702 GN = AXX17_At5g22580 PE = 4 SV = 1 | tr|A0A178UBZ3|A0A178UBZ3_ARATH | 41 kDa | Arabidopsis thaliana |
| Chlorophyll a-b binding protein, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = AXX17_At1g16590 PE = 3 SV = 1 | tr|A0A178WK60|A0A178WK60_ARATH | 27 kDa | Arabidopsis thaliana |
| Photosystem II CP43 reaction center protein OS = Arabidopsis thaliana OX = 3702 GN = psbC PE = 1 SV = 3 | sp|P56778|PSBC_ARATH (+1) | 52 kDa | Arabidopsis thaliana |
| Uncharacterized protein OS = Arabidopsis thaliana OX = 3702 GN = AXX17_At1g65710 PE = 4 SV = 1 | tr|A0A178W9Y8|A0A178W9Y8_ARATH (+1) | 32 kDa | Arabidopsis thaliana |
| Photosystem I chlorophyll a/b-binding protein 3-1, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = LHCA3 PE = 1 SV = 1 | sp|Q9SY97|LHCA3_ARATH (+1) | 29 kDa | Arabidopsis thaliana |
| FtsH extracellular protease family OS = Arabidopsis thaliana OX = 3702 GN = VAR2 PE = 3 SV = 1 | tr|A0A1P8AXC1|A0A1P8AXC1_ARATH (+1) | 75 kDa | unknown |
| APE1 OS = Arabidopsis thaliana OX = 3702 GN = AXX17_At5g36010 PE = 1 SV = 1 | tr|A0A178US93|A0A178US93_ARATH (+3) | 31 kDa | unknown |
| ATP-dependent zinc metalloprotease FTSH 1, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = FTSH1 PE = 1 SV = 2 | sp|Q39102|FTSH1_ARATH (+1) | 77 kDa | Arabidopsis thaliana |
| Photosystem II 10 kDa polypeptide, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = PSBR PE = 1 SV = 1 | sp|P27202|PSBR_ARATH (+2) | 15 kDa | Arabidopsis thaliana |
| Cytochrome b559 subunit alpha OS = Arabidopsis thaliana OX = 3702 GN = psbE PE = 1 SV = 4 | sp|P56779|PSBE_ARATH | 9 kDa | Arabidopsis thaliana |
| Photosystem II CP47 reaction center protein OS = Arabidopsis thaliana OX = 3702 GN = psbB PE = 1 SV = 1 | sp|P56777|PSBB_ARATH (+1) | 56 kDa | Arabidopsis thaliana |
| Photosystem I P700 chlorophyll a apoprotein A2 OS = Arabidopsis thaliana OX = 3702 GN = psaB PE = 3 SV = 1 | sp|P56767|PSAB_ARATH (+1) | 82 kDa | Arabidopsis thaliana |
| HCF244 OS = Arabidopsis thaliana OX = 3702 GN = AXX17_At4g40280 PE = 1 SV = 1 | tr|A0A178UXV2|A0A178UXV2_ARATH (+1) | 44 kDa | unknown |
| Cytochrome b6 OS = Arabidopsis thaliana OX = 3702 GN = petB PE = 1 SV = 1 | sp|P56773|CYB6_ARATH | 24 kDa | Arabidopsis thaliana |
| Photosystem II protein D1 OS = Arabidopsis thaliana OX = 3702 GN = psbA PE = 1 SV = 2 | sp|P83755|PSBA_ARATH (+1) | 39 kDa | Arabidopsis thaliana |
| Photosystem I reaction center subunit V, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = PSAG PE = 1 SV = 1 | sp|Q9S7N7|PSAG_ARATH (+1) | 17 kDa | Arabidopsis thaliana |
| UPF0603 protein At1g54780, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = At1g54780 PE = 1 SV = 1 | sp|Q9ZVL6|U603_ARATH (+1) | 31 kDa | Arabidopsis thaliana |
| Photosystem I chlorophyll a/b-binding protein 2, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = LHCA2 PE = 1 SV = 1 | sp|Q9SYW8|LHCA2_ARATH (+1) | 28 kDa | Arabidopsis thaliana |
| Chlorophyll a-b binding protein 6, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = LHCA1 PE = 1 SV = 1 | sp|Q01667|CAB6_ARATH (+2) | 26 kDa | Arabidopsis thaliana |
| Photosystem I P700 chlorophyll a apoprotein A1 OS = Arabidopsis thaliana OX = 3702 GN = psaA PE = 2 SV = 1 | sp|P56766|PSAA_ARATH (+1) | 83 kDa | Arabidopsis thaliana |
| Chlorophyll a-b binding protein CP29.2, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = LHCB4.2 PE = 1 SV = 1 | sp|Q9XF88|CB4B_ARATH | 31 kDa | Arabidopsis thaliana |
| Chlorophyll synthase, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = CHLG PE = 2 SV = 1 | sp|Q38833|CHLG_ARATH (+1) | 42 kDa | Arabidopsis thaliana |
| Photosystem I reaction center subunit N, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = PSAN PE = 1 SV = 2 | sp|P49107|PSAN_ARATH (+1) | 18 kDa | Arabidopsis thaliana |
| GTP binding Elongation factor Tu family protein OS = Arabidopsis thaliana OX = 3702 GN = At1g07930 PE = 1 SV = 1 | tr|F4HUA0|F4HUA0_ARATH | 41 kDa | Arabidopsis thaliana |
| Photosystem I subunit O OS = Arabidopsis thaliana OX = 3702 GN = PSAO PE = 1 SV = 1 | sp|Q949Q5|PSAO_ARATH (+1) | 15 kDa | Arabidopsis thaliana |
| Probable plastid-lipid-associated protein 8, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = PAP8 PE = 1 SV = 1 | sp|Q941D3|PAP8_ARATH (+1) | 26 kDa | Arabidopsis thaliana |
| ATP synthase subunit alpha, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = atpA PE = 1 SV = 1 | sp|P56757|ATPA_ARATH (+1) | 55 kDa | Arabidopsis thaliana |
| Light-regulated protein 1, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = LIR1 PE = 1 SV = 1 | sp|Q96500|LIRP1_ARATH (+1) | 15 kDa | Arabidopsis thaliana |
| Chlorophyll a-b binding protein 3, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = LHCB3 PE = 1 SV = 1 | sp|Q9S7M0|CB3_ARATH (+1) | 29 kDa | Arabidopsis thaliana |
| At5g08050/F13G24_250 OS = Arabidopsis thaliana OX = 3702 PE = 2 SV = 1 | tr|Q8VYV1|Q8VYV1_ARATH | 17 kDa | Arabidopsis thaliana |
| Protein CURVATURE THYLAKOID 1A, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = CURT1A PE = 1 SV = 1 | sp|O04616|CUT1A_ARATH (+1) | 18 kDa | Arabidopsis thaliana |
| PGR5-like protein 1A, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = PGRL1A PE = 1 SV = 1 | sp|Q8H112|PGL1A_ARATH (+1) | 36 kDa | Arabidopsis thaliana |
| Photosystem I reaction center subunit II-1, chloroplastic OS = Arabidopsis thaliana OX = 3702 GN = psaD1 PE = 1 SV = 1 | sp|Q9S7H1|PSAD1_ARATH (+3) | 23 kDa | Arabidopsis thaliana |
| Actin-7 OS = Arabidopsis thaliana OX = 3702 GN = ACT7 PE = 1 SV = 1 | sp|P53492|ACT7_ARATH (+1) | 42 kDa | unknown |
| AT4g28750 OS = Arabidopsis thaliana OX = 3702 GN = At4g28750/F16A16_140 PE = 2 SV = 1 | tr|Q7FY22|Q7FY22_ARATH (+1) | 12 kDa | Arabidopsis thaliana |
| Photosystem I iron-sulfur center OS = Arabidopsis thaliana OX = 3702 GN = psaC PE = 3 SV = 2 | sp|P62090|PSAC_ARATH (+1) | 9 kDa | Arabidopsis thaliana |
| Probable histone H2A variant 3 OS = Arabidopsis thaliana OX = 3702 GN = At1g52740 PE = 1 SV = 1 | sp|Q9C944|H2AV3_ARATH (+1) | 14 kDa | Arabidopsis thaliana |
| Tubulin beta chain OS = Arabidopsis thaliana OX = 3702 GN = AXX17_At5g62240 PE = 3 SV = 1 | tr|A0A178ULE0|A0A178ULE0_ARATH | 51 kDa | unknown |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vojta, L.; Rac-Justament, A.; Zechmann, B.; Fulgosi, H. Thylakoid Rhodanese-like Protein–Ferredoxin:NADP+ Oxidoreductase Interaction Is Integrated into Plant Redox Homeostasis System. Antioxidants 2023, 12, 1838. https://doi.org/10.3390/antiox12101838
Vojta L, Rac-Justament A, Zechmann B, Fulgosi H. Thylakoid Rhodanese-like Protein–Ferredoxin:NADP+ Oxidoreductase Interaction Is Integrated into Plant Redox Homeostasis System. Antioxidants. 2023; 12(10):1838. https://doi.org/10.3390/antiox12101838
Chicago/Turabian StyleVojta, Lea, Anja Rac-Justament, Bernd Zechmann, and Hrvoje Fulgosi. 2023. "Thylakoid Rhodanese-like Protein–Ferredoxin:NADP+ Oxidoreductase Interaction Is Integrated into Plant Redox Homeostasis System" Antioxidants 12, no. 10: 1838. https://doi.org/10.3390/antiox12101838
APA StyleVojta, L., Rac-Justament, A., Zechmann, B., & Fulgosi, H. (2023). Thylakoid Rhodanese-like Protein–Ferredoxin:NADP+ Oxidoreductase Interaction Is Integrated into Plant Redox Homeostasis System. Antioxidants, 12(10), 1838. https://doi.org/10.3390/antiox12101838