
Natural Compounds Affecting Inflammatory Pathways of Osteoarthritis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Inclusion Criteria
2.3. Exclusion Criteria
3. Results
4. Pathogenesis of OA
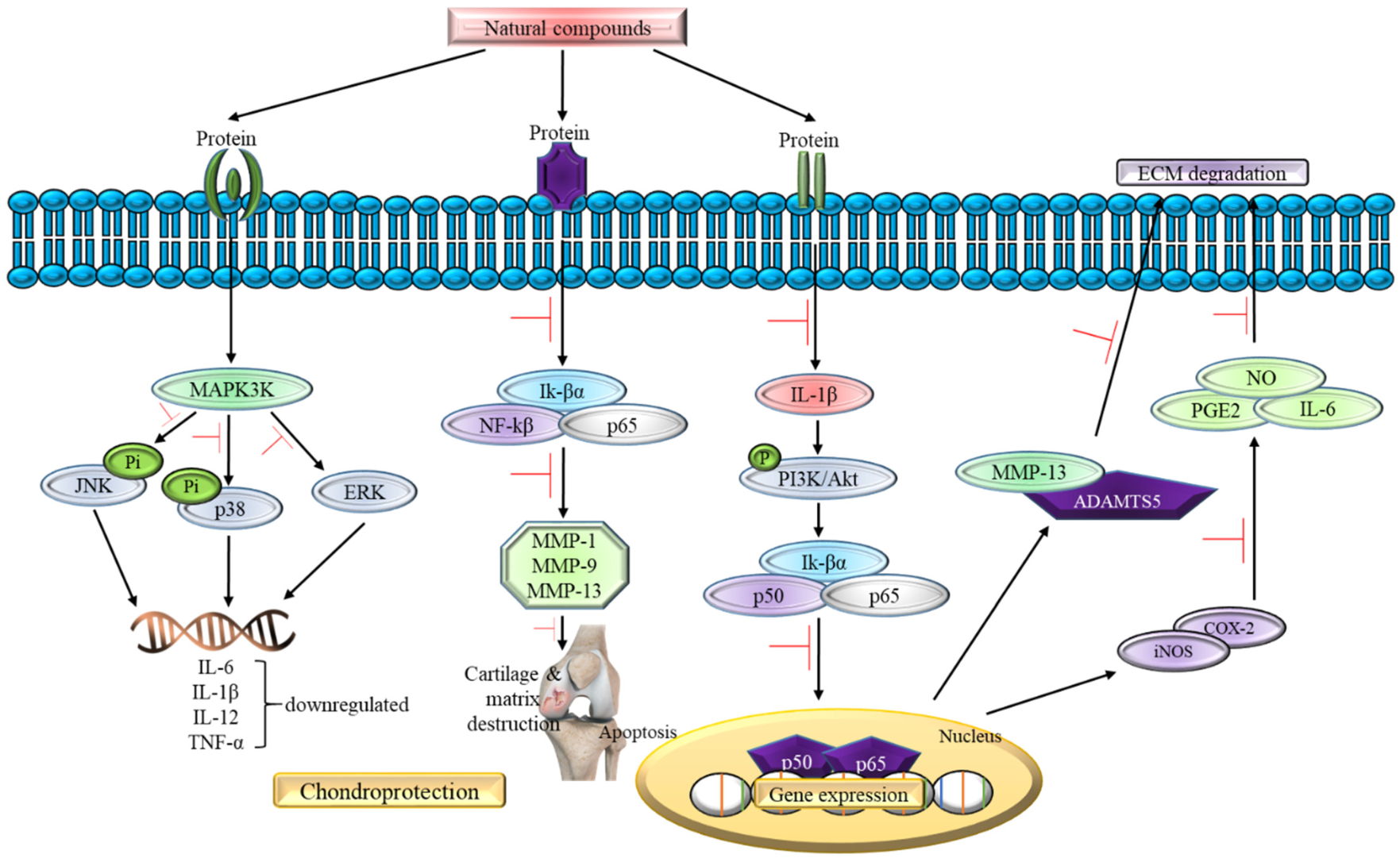
Inflammatory Pathway in Osteoarthritis Pathogenesis
5. In Vitro and In Vivo Evidence of Natural Compounds Affecting Inflammatory Pathways of Osteoarthritis
Role of Natural Compounds in Oxidative Stress Signaling Pathway in Osteoarthritis
6. Role of Natural Compounds on Oxidative Stress Signaling in OA
6.1. Aconitum carmichaelii Debx
6.2. Anthriscus sylvestris (L.) Hoffm
6.3. Artemisia argyi H. Lév. and Vaniot
6.4. Edible Bird’s Nest (EBN)
6.5. Stichopus chloronotus
6.6. Galangin
6.7. Astaxanthin
6.8. Berberine
6.9. Sesamin
6.10. Wogonin
7. Catabolic and Anabolic Effect of Natural Compounds in OA
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mora, J.C.; Przkora, R.; Cruz-Almeida, Y. Knee osteoarthritis: Pathophysiology and current treatment modalities. J. Pain Res. 2018, 11, 2189–2196. [Google Scholar] [CrossRef] [PubMed]
- Katz, J.N.; Arant, K.R.; Loeser, R.F. Diagnosis and Treatment of Hip and Knee Osteoarthritis: A Review. JAMA 2020, 325, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Musumeci, G.; Aiello, F.C.; Szychlinska, M.A.; Rosa, M.D.; Castrogiovanni, P.; Mobasheri, A. Osteoarthritis in the XXIst Century: Risk Factors and Behaviours that Influence Disease Onset and Progression. Int. J. Mol. Sci. 2015, 16, 6093–6112. [Google Scholar] [CrossRef]
- Hamood, R.; Tirosh, M.; Fallach, N.; Chodick, G.; Eisenberg, E.; Lubovsky, O. Prevalence and Incidence of Osteoarthritis: A Population-Based Retrospective Cohort Study. J. Clin. Med. 2021, 10, 4282. [Google Scholar] [CrossRef]
- Pérez-Lozano, M.L.; Cesaro, A.; Mazor, M.; Esteve, E.; Berteina-Raboin, S.; Best, T.M.; Lespessailles, E.; Toumi, H. Emerging Natural-Product-Based Treatments for the Management of Osteoarthritis. Antioxidants 2021, 10, 265. [Google Scholar] [CrossRef]
- Loeser, R.F.; Goldring, S.R.; Scanzello, C.R.; Goldring, M.B. Osteoarthritis: A disease of the joint as an organ. Arthritis Rheum. 2012, 64, 1697–1707. [Google Scholar] [CrossRef] [PubMed]
- Goldring, S.R.; Goldring, M.B. Changes in the osteochondral unit during osteoarthritis: Structure, function and cartilage–bone crosstalk. Nat. Rev. Rheumatol. 2016, 12, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Azzini, G.O.M.; Santos, G.S.; Visoni, S.B.C.; Azzini, V.O.M.; dos Santos, R.G.; Huber, S.C.; Lana, J.F. Metabolic syndrome and subchondral bone alterations: The rise of osteoarthritis—A review. J. Clin. Orthop. Trauma 2020, 11, S849–S855. [Google Scholar] [CrossRef] [PubMed]
- Kuyinu, E.L.; Narayanan, G.; Nair, L.S.; Laurencin, C.T. Animal models of osteoarthritis: Classification, update, and measurement of outcomes. J. Orthop. Surg. Res. 2016, 11, 19. [Google Scholar] [CrossRef]
- Yu, S.P.; Hunter, D.J. Managing osteoarthritis. Aust. Prescr. 2015, 38, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Uchôa, M.; Constantino, G. Viscosupplementation. Rev. Bras. Ortop. 2012, 47, 160–164. [Google Scholar] [CrossRef]
- De l’Escalopier, N.; Anract, P.; Biau, D. Surgical treatments for osteoarthritis. Ann. Phys. Rehabil. Med. 2016, 59, 227–233. [Google Scholar] [CrossRef]
- Patrignani, P.; Tacconelli, S.; Bruno, A.; Sostres, C.; Lanas, A. Managing the adverse effects of nonsteroidal anti-inflammatory drugs. Expert Rev. Clin. Pharmacol. 2011, 4, 605–621. [Google Scholar] [CrossRef] [PubMed]
- Sukhikh, S.; Noskova, S.; Ivanova, S.; Ulrikh, E.; Izgaryshev, A.; Babich, O. Chondroprotection and Molecular Mechanism of Action of Phytonutraceuticals on Osteoarthritis. Molecules 2021, 26, 2391. [Google Scholar] [CrossRef] [PubMed]
- Sukhikh, S.; Babich, O.; Prosekov, A.; Patyukov, N.; Ivanova, S. Future of Chondroprotectors in the Treatment of Degenerative Processes of Connective Tissue. Pharmaceuticals 2020, 13, 220. [Google Scholar] [CrossRef]
- Hafsi, K.; McKay, J.; Li, J.; Lana, J.F.; Macedo, A.; Santos, G.S.; Murrell, W.D. Nutritional, metabolic and genetic considerations to optimise regenerative medicine outcome for knee osteoarthritis. J. Clin. Orthop. Trauma 2018, 10, 2–8. [Google Scholar] [CrossRef]
- Jayakumar, T.; Bhavan, P.S.; Sheu, J.R. Molecular Targets of Natural Products for Chondroprotection in Destructive Joint Diseases. Int. J. Mol. Sci. 2020, 21, 4931. [Google Scholar] [CrossRef]
- Man, G.S.; Mologhianu, G. Osteoarthritis pathogenesis—A complex process that involves the entire joint. J. Med. Life 2014, 7, 41. [Google Scholar]
- Roughley, P.J.; Mort, J.S. The role of aggrecan in normal and osteoarthritic cartilage. J. Exp. Orthop. 2014, 1, 8. [Google Scholar] [CrossRef]
- Walimbe, T.; Panitch, A. Proteoglycans in biomedicine: Resurgence of an underexploited class of ECM molecules. Front. Pharmacol. 2019, 10, 1661. [Google Scholar] [CrossRef]
- Wang, M.; Peng, Z.; Vasilev, K.; Ketheesan, N. Investigation of Wear Particles Generated in Human Knee Joints Using Atomic Force Microscopy. Tribol. Lett. 2013, 51, 161–170. [Google Scholar] [CrossRef]
- Stannus, O.; Jones, G.; Cicuttini, F.; Parameswaran, V.; Quinn, S.; Burgess, J.; Ding, C. Circulating levels of IL-6 and TNF-α are associated with knee radiographic osteoarthritis and knee cartilage loss in older adults. Osteoarthr. Cartil. 2010, 18, 1441–1447. [Google Scholar] [CrossRef] [PubMed]
- Sonnenberg, G.F.; Artis, D. Innate lymphoid cells in the initiation, regulation and resolution of inflammation. Nat. Med. 2015, 21, 698–708. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Abu-Amer, Y.; O’Keefe, R.J.; McAlinden, A. Inflammation and epigenetic regulation in osteoarthritis. Connect. Tissue Res. 2016, 58, 49–63. [Google Scholar] [CrossRef]
- Wojdasiewicz, P.; Poniatowski, Ł.A.; Szukiewicz, D. The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis. Med. Inflamm. 2014, 2014, 561459. [Google Scholar] [CrossRef]
- Chow, Y.Y.; Chin, K.Y. The Role of Inflammation in the Pathogenesis of Osteoarthritis. Med. Inflamm. 2020, 2020, 8293921. [Google Scholar] [CrossRef]
- Scanzello, C.R. Chemokines and inflammation in osteoarthritis: Insights from patients and animal models. J. Orthop. Res. 2017, 35, 735–739. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. NF-κB, the first quarter-century: Remarkable progress and outstanding questions. Genes Dev. 2012, 26, 203–234. [Google Scholar] [CrossRef]
- Choi, M.C.; Jo, J.; Park, J.; Kang, H.K.; Park, Y. NF-κB Signaling Pathways in Osteoarthritic Cartilage Destruction. Cells 2019, 8, 734. [Google Scholar] [CrossRef]
- Kapoor, M.; Martel-Pelletier, J.; Lajeunesse, D.; Pelletier, J.P.; Fahmi, H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat. Rev. Rheumatol. 2010, 7, 33–42. [Google Scholar] [CrossRef]
- Ni, S.; Miao, K.; Zhou, X.; Xu, N.; Li, C.; Zhu, R.; Sun, R.; Wang, Y. The involvement of follistatin-like protein 1 in osteoarthritis by elevating NF-κB-mediated inflammatory cytokines and enhancing fibroblast like synoviocyte proliferation. Arthritis Res. Ther. 2015, 17, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigoglou, S.; Papavassiliou, A.G. The NF-κB signalling pathway in osteoarthritis. Int. J. Biochem. Cell Biol. 2013, 45, 2580–2584. [Google Scholar] [CrossRef] [PubMed]
- Viana, M.N.; Leiguez, E.; Gutiérrez, J.M.; Rucavado, A.; Markus, R.P.; Marçola, M.; Teixeira, C.; Fernandes, C.M. A representative metalloprotease induces PGE2 synthesis in fibroblast-like synoviocytes via the NF-κB/COX-2 pathway with amplification by IL-1β and the EP4 receptor. Sci. Rep. 2020, 10, 3269. [Google Scholar] [CrossRef] [PubMed]
- De Andrés, M.C.; Imagawa, K.; Hashimoto, K.; Gonzalez, A.; Roach, H.I.; Goldring, M.B.; Oreffo, R.O.C. Loss of methylation in CpG sites in the NF-κB enhancer elements of inducible nitric oxide synthase is responsible for gene induction in human articular chondrocytes. Arthritis Rheum. 2013, 65, 732–742. [Google Scholar] [CrossRef]
- Ahmad, N.; Ansari, M.Y.; Haqqi, T.M. Role of iNOS in osteoarthritis: Pathological and therapeutic aspects. J. Cell. Physiol. 2020, 235, 6366–6376. [Google Scholar] [CrossRef]
- Yunus, M.H.M.; Nordin, A.; Kamal, H. Pathophysiological Perspective of Osteoarthritis. Medicina 2020, 56, 614. [Google Scholar] [CrossRef]
- Xie, L.; Xie, H.; Chen, C.; Tao, Z.; Zhang, C.; Cai, L. Inhibiting the PI3K/AKT/NF-κB signal pathway with nobiletin for attenuating the development of osteoarthritis: In vitro and in vivo studies. Food Funct. 2019, 10, 2161–2175. [Google Scholar] [CrossRef]
- Chen, Y.; Shou, K.; Gong, C.; Yang, H.; Yang, Y.; Bao, T. Anti-Inflammatory Effect of Geniposide on Osteoarthritis by Suppressing the Activation of p38 MAPK Signaling Pathway. BioMed Res. Int. 2018, 2018, 8384576. [Google Scholar] [CrossRef]
- Keshet, Y.; Seger, R. The MAP kinase signaling cascades: A system of hundreds of components regulates a diverse array of physiological functions. Methods Mol. Biol. 2010, 661, 3–38. [Google Scholar] [CrossRef]
- Loeser, R.F.; Erickson, E.A.; Long, D.L. Mitogen-activated protein kinases as therapeutic targets in osteoarthritis. Curr. Opin. Rheumatol. 2008, 20, 581–586. [Google Scholar] [CrossRef]
- Zahan, O.M.; Serban, O.; Gherman, C.; Fodor, D. The evaluation of oxidative stress in osteoarthritis. Med. Pharm. Rep. 2020, 93, 22. [Google Scholar] [CrossRef] [PubMed]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef] [PubMed]
- Charlier, E.; Deroyer, C.; Ciregia, F.; Malaise, O.; Neuville, S.; Plener, Z.; Malaise, M.; de Seny, D. Chondrocyte dedifferentiation and osteoarthritis (OA). Biochem. Pharmacol. 2019, 165, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Csaki, C.; Mobasheri, A.; Shakibaei, M. Synergistic chondroprotective effects of curcumin and resveratrol in human articular chondrocytes: Inhibition of IL-1β-induced NF-κB-mediated inflammation and apoptosis. Arthritis Res. Ther. 2009, 11, R165. [Google Scholar] [CrossRef]
- Gilbert, S.J.; Bonnet, C.S.; Blain, E.J. Mechanical Cues: Bidirectional Reciprocity in the Extracellular Matrix Drives Mechano-Signalling in Articular Cartilage. Int. J. Mol. Sci. 2021, 22, 13595. [Google Scholar] [CrossRef]
- Na, J.Y.; Kim, S.; Song, K.; Lim, K.H.; Shin, G.W.; Kim, J.H.; Kim, B.; Kwon, Y.B.; Kwon, J. Anti-apoptotic Activity of Ginsenoside Rb1 in Hydrogen Peroxide-treated Chondrocytes: Stabilization of Mitochondria and the Inhibition of Caspase-3. J. Ginseng Res. 2012, 36, 247. [Google Scholar] [CrossRef]
- Marsden, V.S.; O’Connor, L.; O’Reilly, L.A.; Silke, J.; Metcalf, D.; Ekert, P.G.; Huang, D.C.S.; Cecconi, F.; Kuida, K.; Tomaselli, K.J.; et al. Apoptosis initiated by Bcl-2-regulated caspase activation independently of the cytochrome c/Apaf-1/caspase-9 apoptosome. Nature 2002, 419, 634–637. [Google Scholar] [CrossRef]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase Functions in Cell Death and Disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a008656. [Google Scholar] [CrossRef]
- Yang, Y.; Shen, P.; Yao, T.; Ma, J.; Chen, Z.; Zhu, J.; Gong, Z.; Shen, S.; Fang, X. Novel role of circRSU1 in the progression of osteoarthritis by adjusting oxidative stress. Theranostics 2021, 11, 1900. [Google Scholar] [CrossRef]
- Huang, X.; Pei, W.; Ni, B.; Zhang, R.; You, H. Chondroprotective and antiarthritic effects of galangin in osteoarthritis: An in vitro and in vivo study. Eur. J. Pharmacol. 2021, 906, 174232. [Google Scholar] [CrossRef]
- Su, Y.; Shen, L.; Xue, J.; Zou, J.; Wan, D.; Shi, Z. Therapeutic evaluation of galangin on cartilage protection and analgesic activity in a rat model of osteoarthritis. Electron. J. Biotechnol. 2021, 53, 8–13. [Google Scholar] [CrossRef]
- El-Ghazaly, M.A.; Abd El-Naby, D.H.; Khayyal, M.T. The influence of irradiation on the potential chondroprotective effect of aqueous extract of propolis in rats. Int. J. Radiat. Biol. 2011, 87, 254–262. [Google Scholar] [CrossRef]
- Chen, W.P.; Xiong, Y.; Shi, Y.X.; Hu, P.F.; Bao, J.P.; Wu, L.D. Astaxanthin reduces matrix metalloproteinase expression in human chondrocytes. Int. Immunopharmacol. 2013, 19, 174–177. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.A.; Moon, S.M.; Han, S.H.; Hwang, E.J.; Park, B.R.; Kim, J.S.; Kim, D.K.; Kim, C.S. Chondroprotective effects of aqueous extract of Anthriscus sylvestris leaves on osteoarthritis in vitro and in vivo through MAPKs and NF-κB signaling inhibition. Biomed. Pharmacother. 2018, 103, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.A.; Park, B.R.; Moon, S.M.; Hong, J.H.; Kim, D.K.; Kim, C.S. Chondroprotective Effect of Cynaroside in IL-1 β-Induced Primary Rat Chondrocytes and Organ Explants via NF-κ B and MAPK Signaling Inhibition. Oxid. Med. Cell. Longev. 2020, 2020, 9358080. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.C.; Lee, H.P.; Hung, C.Y.; Tsai, C.H.; Li, T.M.; Tang, C.H. Berberine attenuates CCN2-induced IL-1β expression and prevents cartilage degradation in a rat model of osteoarthritis. Toxicol. Appl. Pharmacol. 2015, 289, 20–29. [Google Scholar] [CrossRef]
- Wasik, U.; Milkiewicz, M.; Kempinska-Podhorodecka, A.; Milkiewicz, P. Protection against oxidative stress mediated by the Nrf2/Keap1 axis is impaired in Primary Biliary Cholangitis. Sci. Rep. 2017, 7, 44769. [Google Scholar] [CrossRef]
- Qiao, Y.Q.; Jiang, P.F.; Gao, Y.Z. Lutein prevents osteoarthritis through Nrf2 activation and downregulation of inflammation. Arch. Med. Sci. 2016, 14, 624. [Google Scholar] [CrossRef]
- Sun, K.; Luo, J.; Jing, X.; Guo, J.; Yao, X.; Hao, X.; Ye, Y.; Liang, S.; Lin, J.; Wang, G.; et al. Astaxanthin protects against osteoarthritis via Nrf2: A guardian of cartilage homeostasis. Aging 2019, 11, 10513–10531. [Google Scholar] [CrossRef]
- Sanada, Y.; Tan, S.J.O.; Adachi, N.; Miyaki, S. Pharmacological targeting of heme oxygenase-1 in osteoarthritis. Antioxidants 2021, 10, 419. [Google Scholar] [CrossRef]
- Drevet, S.; Gavazzi, G.; Grange, L.; Dupuy, C.; Lardy, B. Reactive oxygen species and NADPH oxidase 4 involvement in osteoarthritis. Exp. Gerontol. 2018, 111, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, A.; Kamrava, S.K.; Joghataei, M.T.; Darabi, R.; Shakeri-Zadeh, A.; Shahriari, M.; Reiter, R.J.; Ghaznavi, H.; Mehrzadi, S. Apoptosis signaling pathways in osteoarthritis and possible protective role of melatonin. J. Pineal Res. 2016, 61, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Leong, D.J.; Xu, L.; He, Z.; Wang, A.; Navati, M.; Kim, S.J.; Hirsh, D.M.; Hardin, J.A.; Cobelli, N.J.; et al. Curcumin slows osteoarthritis progression and relieves osteoarthritis-associated pain symptoms in a post-traumatic osteoarthritis mouse model. Arthritis Res. Ther. 2016, 18, 128. [Google Scholar] [CrossRef] [PubMed]
- Leong, D.J.; Choudhury, M.; Hanstein, R.; Hirsh, D.M.; Kim, J.J.; Majeska, R.J.; Schaffler, M.B.; Hardin, J.A.; Spray, D.C.; Goldring, M.B.; et al. Green tea polyphenol treatment is chondroprotective, anti-inflammatory and palliative in a mouse posttraumatic osteoarthritis model. Arthritis Res. Ther. 2014, 16, 508. [Google Scholar] [CrossRef]
- He, Z.; Leong, D.J.; Xu, L.; Hardin, J.A.; Majeska, R.J.; Schaffler, M.B.; Thi, M.M.; Yang, L.; Goldring, M.B.; Cobelli, N.J.; et al. CITED2 mediates the cross-talk between mechanical loading and IL-4 to promote chondroprotection. Ann. N. Y. Acad. Sci. 2019, 1442, 128–137. [Google Scholar] [CrossRef]
- Hanprasertpong, N.; Teekachunhatean, S.; Chaiwongsa, R.; Ongchai, S.; Kunanusorn, P.; Sangdee, C.; Panthong, A.; Bunteang, S.; Nathasaen, N.; Reutrakul, V. Analgesic, Anti-Inflammatory, and Chondroprotective Activities of Cryptolepis buchanani Extract: In Vitro and in Vivo Studies. BioMed Res. Int. 2014, 2014, 978582. [Google Scholar] [CrossRef]
- Zhang, L.; Li, T.; Wang, R.; Xu, J.; Zhou, L.; Yan, L.; Hu, Z.; Li, H.; Liu, F.; Du, W.; et al. Evaluation of Long-Time Decoction-Detoxicated Hei-Shun-Pian (Processed Aconitum carmichaeli Debeaux Lateral Root With Peel) for Its Acute Toxicity and Therapeutic Effect on Mono-Iodoacetate Induced Osteoarthritis. Front. Pharmacol. 2020, 11, 1053. [Google Scholar] [CrossRef]
- Jeong, J.H.; Moon, S.J.; Jhun, J.Y.; Yang, E.J.; Cho, M.L.; Min, J.K. Eupatilin exerts antinociceptive and chondroprotective properties in a rat model of osteoarthritis by downregulating oxidative damage and catabolic activity in chondrocytes. PLoS ONE 2015, 10, e0130882. [Google Scholar] [CrossRef]
- Miller, M.J.S.; Bobrowski, P.; Shukla, M.; Gupta, K.; Haqqi, T.M. Chondroprotective effects of a proanthocyanidin rich Amazonian genonutrient reflects direct inhibition of matrix metalloproteinases and upregulation of IGF-1 production by human chondrocytes. J. Inflamm. 2007, 4, 16. [Google Scholar] [CrossRef]
- Shakibaei, M.; John, T.; Seifarth, C.; Mobasheri, A. Resveratrol inhibits IL-1β-induced stimulation of caspase-3 and cleavage of PARP in human articular chondrocytes in vitro. Ann. N. Y. Acad. Sci. 2007, 1095, 554–563. [Google Scholar] [CrossRef]
- Sumantran, V.N.; Kulkarni, A.; Boddul, S.; Chinchwade, T.; Koppikar, S.J.; Harsulkar, A.; Patwardhan, B.; Chopra, A.; Wagh, U.V. Chondroprotective potential of root extracts of Withania somnifera in osteoarthritis. J. Biosci. 2007, 32, 299–307. [Google Scholar] [CrossRef]
- Sumantran, V.N.; Kulkarni, A.; Chandwaskar, R.; Harsulkar, A.; Patwardhan, B.; Chopra, A.; Wagh, U.V. Chondroprotective potential of fruit extracts of Phyllanthus emblica in osteoarthritis. Evidence-based Complement. Altern. Med. 2008, 5, 329–335. [Google Scholar] [CrossRef]
- Sanyachare, S.; Itghiarbha, A.; Kongtawele, P.; Meepowpan, P.; Nuntasaen, N.; Pompimon, W. A New Polyoxypregnane Glycoside from the Roots of Dregea volubilis (L.f) Benth. ex Hook. f and its Chondroprotective Effect. Am. J. Biochem. Biotechnol. 2009, 5, 202–209. [Google Scholar] [CrossRef]
- Liu, F.C.; Hung, L.F.; Wu, W.L.; Chang, D.M.; Huang, C.Y.; Lai, J.H.; Ho, L.J. Chondroprotective effects and mechanisms of resveratrol in advanced glycation end products-stimulated chondrocytes. Arthritis Res. Ther. 2010, 12, R167. [Google Scholar] [CrossRef]
- Hu, P.F.; Chen, W.P.; Tang, J.L.; Bao, J.P.; Wu, L.D. Protective effects of berberine in an experimental rat osteoarthritis model. Phyther. Res. 2011, 25, 878–885. [Google Scholar] [CrossRef]
- Rujirek, C.; Siriwan, O.; Siriwan, T.; Prachya, K.; Ampai, P.; Vichai, R. Chondroprotective potential of bioactive compounds of Zingiber cassumunar Roxb. against cytokine-induced cartilage degradation in explant culture. J. Med. Plants Res. 2012, 6, 5204–5213. [Google Scholar] [CrossRef]
- Itthiarbha, A.; Phitak, T.; Sanyacharernkul, S.; Pothacharoen, P.; Pompimon, W.; Kongtawelert, P. Polyoxypregnane glycoside from Dregea volubilis extract inhibits IL-1ß-induced expression of matrix metalloproteinase via activation of NF-kB in human chondrocytes. In Vitro Cell. Dev. Biol. Anim. 2012, 48, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Phitak, T.; Pothacharoen, P.; Settakorn, J.; Poompimol, W.; Caterson, B.; Kongtawelert, P. Chondroprotective and anti-inflammatory effects of sesamin. Phytochemistry 2012, 80, 77–88. [Google Scholar] [CrossRef]
- Chua, K.H.; Lee, T.H.; Nagandran, K.; Md Yahaya, N.H.; Lee, C.T.; Tjih, E.T.T.; Abdul Aziz, R. Edible Bird’s nest extract as a chondro-protective agent for human chondrocytes isolated from osteoarthritic knee: In vitro study. BMC Complement. Altern. Med. 2013, 13, 2–9. [Google Scholar] [CrossRef]
- Liu, L.; Gu, H.; Liu, H.; Jiao, Y.; Li, K.; Zhao, Y.; An, L.; Yang, J. Protective Effect of Resveratrol against IL-1β-Induced Inflammatory Response on Human Osteoarthritic Chondrocytes Partly via the TLR4/MyD88/NF-κB Signaling Pathway: An “in Vitro” Study. Int. J. Mol. Sci. 2014, 15, 6925–6940. [Google Scholar] [CrossRef]
- Tong, P.; Xu, S.; Cao, G.; Jin, W.; Guo, Y.; Cheng, Y.; Jin, H.; Shan, L.; Xiao, L. Chondroprotective activity of a detoxicated traditional Chinese medicine (Fuzi) of Aconitum carmichaeli Debx against severe-stage osteoarthritis model induced by mono-iodoacetate. J. Ethnopharmacol. 2013, 151, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Rufino, A.T.; Ribeiro, M.; Judas, F.; Salgueiro, L.; Lopes, M.C.; Cavaleiro, C.; Mendes, A.F. Anti-inflammatory and chondroprotective activity of (+)-α-pinene: Structural and enantiomeric selectivity. J. Nat. Prod. 2014, 77, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Pradit, W.; Chomdej, S.; Nganvongpanit, K.; Ongchai, S. Chondroprotective potential of Phyllanthus amarus Schum. Thonn. in experimentally induced cartilage degradation in the explants culture model. In Vitro Cell. Dev. Biol.-Anim. 2014, 51, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Qu, T.B.; Zhai, K.; Ding, J.; Hai, Y.; Zhou, J.L. Gallic acid can play a chondroprotective role against AGE-induced osteoarthritis progression. J. Orthop. Sci. 2015, 20, 734–741. [Google Scholar] [CrossRef]
- Park, J.S.; Lee, H.J.; Lee, D.Y.; Jo, H.S.; Jeong, J.H.; Kim, D.H.; Nam, D.C.; Lee, C.J.; Hwang, S.C. Chondroprotective Effects of Wogonin in Experimental Models of Osteoarthritis in vitro and in vivo. Biomol. Ther. 2015, 23, 442–448. [Google Scholar] [CrossRef]
- Buddhachat, K.; Chomdej, S.; Pradit, W.; Nganvongpanit, K.; Ongchai, S. In Vitro Chondroprotective Potential of Extracts Obtained from Various Phyllantus Species. Planta Med. 2016, 83, 87–96. [Google Scholar] [CrossRef]
- Kong, P.; Chen, G.; Jiang, A.; Wang, Y.; Song, C.; Zhuang, J.; Xi, C.; Wang, G.; Ji, Y.; Yan, J.; et al. Sesamin inhibits IL-1β stimulated inflammatory response in human osteoarthritis chondrocytes by activating Nrf2 signaling pathway. Oncotarget 2016, 7, 83720–83726. [Google Scholar] [CrossRef]
- Liu, S.; Yang, H.; Hu, B.; Zhang, M. Sirt1 regulates apoptosis and extracellular matrix degradation in resveratrol-treated osteoarthritis chondrocytes via the wnt/β-catenin signaling pathways. Exp. Ther. Med. 2017, 14, 5057–5062. [Google Scholar] [CrossRef]
- Gu, H.; Jiao, Y.; Yu, X.; Li, X.; Wang, W.; Ding, L.; Liu, L. Resveratrol inhibits the IL-1β-induced expression ofMMP-13 and IL-6 in human articular chondrocytes viaTLR4/MyD88-dependent and-independent signaling cascades. Int. J. Mol. Med. 2017, 39, 734–740. [Google Scholar] [CrossRef]
- Khan, N.M.; Haseeb, A.; Ansari, M.Y.; Devarapalli, P.; Haynie, S.; Haqqi, T.M. Wogonin, a plant derived small molecule, exerts potent anti-inflammatory and chondroprotective effects through the activation of ROS/ERK/Nrf2 signaling pathways in human Osteoarthritis chondrocytes. Free Radic. Biol. Med. 2017, 106, 288–301. [Google Scholar] [CrossRef]
- Liu, F.C.; Wang, C.C.; Lu, J.W.; Lee, C.H.; Chen, S.C.; Ho, Y.J.; Peng, Y.J. Chondroprotective effects of genistein against osteoarthritis induced joint inflammation. Nutrients 2019, 11, 1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ongchai, S.; Chokchaitaweesuk, C.; Kongdang, P.; Chomdej, S.; Buddhachat, K. In vitro chondroprotective potential of Senna alata and Senna tora in porcine cartilage explants and their species differentiation by DNA barcoding-high resolution melting (Bar-HRM) analysis. PLoS ONE 2019, 14, e0215664. [Google Scholar] [CrossRef]
- Mohd Heikal, M.Y.; Ahmad Nazrun, S.; Chua, K.H.; Norzana, A.G. Stichopus chloronotus aqueous extract as a chondroprotective agent for human chondrocytes isolated from osteoarthitis articular cartilage in vitro. Cytotechnology 2019, 71, 521–537. [Google Scholar] [CrossRef]
- D’Ascola, A.; Irrera, N.; Ettari, R.; Bitto, A.; Pallio, G.; Mannino, F.; Atteritano, M.; Campo, G.M.; Minutoli, L.; Arcoraci, V.; et al. Exploiting curcumin synergy with natural products using quantitative analysis of dose-effect relationships in an experimental in vitro model of osteoarthritis. Front. Pharmacol. 2019, 10, 1347. [Google Scholar] [CrossRef]
- Lee, H.; Jang, D.; Jeon, J.; Cho, C.; Choi, S.; Han, S.J.; Oh, E.; Nam, J.; Park, C.H.; Shin, Y.S.; et al. Seomae mugwort and jaceosidin attenuate osteoarthritic cartilage damage by blocking IκB degradation in mice. J. Cell. Mol. Med. 2020, 24, 8126–8137. [Google Scholar] [CrossRef] [PubMed]
- You, H.; Zhang, R.; Wang, L.; Pan, Q.; Mao, Z.; Huang, X. Chondro-Protective Effects of Shikimic Acid on Osteoarthritis via Restoring Impaired Autophagy and Suppressing the MAPK/NF-κB Signaling Pathway. Front. Pharmacol. 2021, 12, 1702. [Google Scholar] [CrossRef] [PubMed]
- Buhrmann, C.; Brockmueller, A.; Mueller, A.L.; Shayan, P.; Shakibaei, M. Curcumin attenuates environment-derived osteoarthritis by Sox9/NF-kB signaling axis. Int. J. Mol. Sci. 2021, 22, 7645. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zhai, Z.; Ying, H.; Lu, L.; Zhang, J.; Zeng, Y. Curcumin primed ADMSCs derived small extracellular vesicle exert enhanced protective effects on osteoarthritis by inhibiting oxidative stress and chondrocyte apoptosis. J. Nanobiotechnol. 2022, 20, 123. [Google Scholar] [CrossRef]
- Wang, J.; Gao, J.S.; Chen, J.W.; Li, F.; Tian, J. Effect of resveratrol on cartilage protection and apoptosis inhibition in experimental osteoarthritis of rabbit. Rheumatol. Int. 2011, 32, 1541–1548. [Google Scholar] [CrossRef]
- Gu, H.; Li, K.; Li, X.; Yu, X.; Wang, W.; Ding, L.; Liu, L. Oral resveratrol prevents osteoarthritis progression in C57BL/6J mice fed a High-Fat Diet. Nutrients 2016, 8, 233. [Google Scholar] [CrossRef]
- Wei, Y.; Jia, J.; Jin, X.; Tong, W.; Tian, H. Resveratrol ameliorates inflammatory damage and protects against osteoarthritis in a rat model of osteoarthritis. Mol. Med. Rep. 2017, 17, 1493–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Cao, J.; Yang, E.; Liang, B.; Ding, J.; Liang, J.; Xu, J. Curcumin improves age-related and surgically induced osteoarthritis by promoting autophagy in mice. Biosci. Rep. 2017, 38, 20171691. [Google Scholar] [CrossRef]
- Jiang, M.; Li, X.; Yu, X.; Liu, X.; Xu, X.; He, J.; Gu, H.; Liu, L. Oral Administration of Resveratrol Alleviates Osteoarthritis Pathology in C57BL/6J Mice Model Induced by a High-Fat Diet. Mediat. Inflamm. 2017, 2017, 7659023. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, W.; Zhang, H.; Li, H.; Liu, J.; Zhang, F.; Jiang, T.; Jiang, S. Curcumin prevents osteoarthritis by inhibiting the activation of inflammasome NLRP3. J. Interf. Cytokine Res. 2017, 37, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; He, B.; Guo, J.; Li, S.; Wang, J. Involvement of TLR4 in the protective effect of intra-articular administration of curcumin on rat experimental osteoarthritis. Acta Cir. Bras. 2019, 34. [Google Scholar] [CrossRef]
- Feng, K.; Ge, Y.; Chen, Z.; Li, X.; Liu, Z.; Li, X.; Li, H.; Tang, T.; Yang, F.; Wang, X. Curcumin inhibits the PERK-eIF2α-CHOP pathway through promoting SIRT1 expression in oxidative stress-induced rat chondrocytes and ameliorates osteoarthritis progression in a rat model. Oxid. Med. Cell. Longev. 2019, 2019, 8574386. [Google Scholar] [CrossRef]
- Jiang, C.; Luo, P.; Li, X.; Liu, P.; Li, Y.; Xu, J. Nrf2/ARE is a key pathway for curcumin-mediated protection of TMJ chondrocytes from oxidative stress and inflammation. Cell Stress Chaperones 2020, 25, 395–406. [Google Scholar] [CrossRef]
- Susmiarsih, T.P.; Hadi, R.S.; Sofwan, A.; Razari, I. Effect of Green Tea Extract to the Degree of Knee Joint Damage and Nitric Oxide Levels in the Rabbit Osteoarthitis Model. In Proceedings of the 2019 International Conference on Science, Technology, and Humanity (ISETH), Surakarta, Indonesia, 3–4 December 2019; pp. 648–657. [Google Scholar]
- Xu, W.; Zhang, M.; Liu, H.; Wei, K.; He, M.; Li, X.; Hu, D.; Yang, S.; Zheng, Y. Antiviral activity of aconite alkaloids from Aconitum carmichaelii Debx. Nat. Prod. Res. 2017, 33, 1486–1490. [Google Scholar] [CrossRef]
- Liu, X.X.; Jian, X.X.; Cai, X.F.; Chao, R.B.; Chen, Q.H.; Chen, D.L.; Wang, X.L.; Wang, F.P. Cardioactive C19-Diterpenoid Alkaloids from the Lateral Roots of Aconitum carmichaeli “Fu Zi”. Chem. Pharm. Bull. 2012, 60, 144–149. [Google Scholar] [CrossRef]
- Shamsul, B.; Chowdhury, S.; Hamdan, M.Y.; Ruszymah, B.H.I. Effect of cell density on formation of three-dimensional cartilaginous constructs using fibrin human osteoarthritic chondrocytes. Indian J. Med. Res. 2019, 149, 649. [Google Scholar] [CrossRef]
- Lajayer, H.M.; Norouzi, R.; Shahi-Gharahlar, A. Essential oil components, phenolic content and antioxidant activity of Anthriscus cerefolium and Anthriscus sylvestris from Iran. J. Hortic. Postharvest Res. 2020, 3, 355–366. [Google Scholar] [CrossRef]
- Jang, G.; Lee, S.A.; Hong, J.H.; Park, B.R.; Kim, D.K.; Kim, C.S. Chondroprotective Effects of 4,5-Dicaffeoylquinic Acid in Osteoarthritis through NF-κB Signaling Inhibition. Antioxidants 2022, 11, 487. [Google Scholar] [CrossRef] [PubMed]
- Abad, M.J.; Bedoya, L.M.; Apaza, L.; Bermejo, P. The Artemisia L. Genus: A Review of Bioactive Essential Oils. Molecules 2012, 17, 2542–2566. [Google Scholar] [CrossRef]
- Lee, T.H.; Wani, W.A.; Lee, C.H.; Cheng, K.K.; Shreaz, S.; Wong, S.; Hamdan, N.; Azmi, N.A. Edible Bird’s Nest: The Functional Values of the Prized Animal-Based Bioproduct From Southeast Asia—A Review. Front. Pharmacol. 2021, 12, 871. [Google Scholar] [CrossRef]
- Mou, J.; Li, Q.; Qi, X.; Yang, J. Structural comparison, antioxidant and anti-inflammatory properties of fucosylated chondroitin sulfate of three edible sea cucumbers. Carbohydr. Polym. 2018, 185, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Yin, H.; Qi, X.; Song, W.; Shi, W.; Mou, J.; Yang, J. Immunomodulatory effects of fucosylated chondroitin sulfate from Stichopus chloronotus on RAW 264.7 cells. Carbohydr. Polym. 2020, 251, 117088. [Google Scholar] [CrossRef] [PubMed]
- Bishnoi, M.; Jain, A.; Hurkat, P.; Jain, S.K. Chondroitin sulphate: A focus on osteoarthritis. Glycoconj. J. 2016, 33, 693–705. [Google Scholar] [CrossRef]
- Michel, B.A.; Stucki, G.; Frey, D.; De Vathaire, F.; Vignon, E.; Bruehlmann, P.; Uebelhart, D. Chondroitins 4 and 6 sulfate in osteoarthritis of the knee: A randomized, controlled trial. Arthritis Rheum. 2005, 52, 779–786. [Google Scholar] [CrossRef]
- Volpi, N. Analytical aspects of pharmaceutical grade chondroitin sulfates. J. Pharm. Sci. 2007, 96, 3168–3180. [Google Scholar] [CrossRef]
- Schiraldi, C.; Cimini, D.; Rosa, M. De Production of chondroitin sulfate and chondroitin. Appl. Microbiol. Biotechnol. 2010, 87, 1209–1220. [Google Scholar] [CrossRef]
- Mak, K.K.; Tan, J.J.; Marappan, P.; Balijepalli, M.K.; Choudhury, H.; Ramamurthy, S.; Pichika, M.R. Galangin’s potential as a functional food ingredient. J. Funct. Foods 2018, 46, 490–503. [Google Scholar] [CrossRef]
- Khan, N.M.; Ahmad, I.; Ansari, M.Y.; Haqqi, T.M. Wogonin, a natural flavonoid, intercalates with genomic DNA and exhibits protective effects in IL-1β stimulated osteoarthritis chondrocytes. Chem. Biol. Interact. 2017, 274, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, J.; Herrera-Bravo, J.; Salazar, L.A.; Shaheen, S.; Abdulmajid Ayatollahi, S.; Kobarfard, F.; Imran, M.; Imran, A.; Custódio, L.; Dolores López, M.; et al. The Therapeutic Potential of Wogonin Observed in Preclinical Studies. Evidence-based Complement. Altern. Med. 2021, 2021, 9935451. [Google Scholar] [CrossRef]
- Jiang, W.; Jin, Y.; Zhang, S.; Ding, Y.; Huo, K.; Yang, J.; Zhao, L.; Nian, B.; Zhong, T.P.; Lu, W.; et al. PGE2 activates EP4 in subchondral bone osteoclasts to regulate osteoarthritis. Bone Res. 2022, 10, 27. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, J.E.; Aspden, R.M. Cyclooxygenase inhibition lowers prostaglandin E2 release from articular cartilage and reduces apoptosis but not proteoglycan degradation following an impact load in vitro. Arthritis Res. Ther. 2007, 9, R129. [Google Scholar] [CrossRef] [PubMed]
- Murphy, G.; Lee, M.H. What are the roles of metalloproteinases in cartilage and bone damage? Ann. Rheum. Dis. 2005, 64, iv44–iv47. [Google Scholar] [CrossRef] [PubMed]
- Vincenti, M.P.; Brinckerhoff, C.E. Transcriptional regulation of collagenase (MMP-1, MMP-13) genes in arthritis: Integration of complex signaling pathways for the recruitment of gene-specific transcription factors. Arthritis Res. 2002, 4, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Matsuda, H.; Tanioka, M.; Kuwabara, K.; Itohara, S.; Suzuki, R. The Role of Matrix Metalloproteinase-2 and Matrix Metalloproteinase-9 in Antibody-Induced Arthritis. J. Immunol. 2002, 169, 2643–2647. [Google Scholar] [CrossRef] [PubMed]
- Malfait, A.M.; Liu, R.Q.; Ijiri, K.; Komiya, S.; Tortorella, M.D. Inhibition of ADAM-TS4 and ADAM-TS5 prevents aggrecan degradation in osteoarthritic cartilage. J. Biol. Chem. 2002, 277, 22201–22208. [Google Scholar] [CrossRef]
- Akeson, G.; Malemud, C.J. A Role for Soluble IL-6 Receptor in Osteoarthritis. J. Funct. Morphol. Kinesiol. 2017, 2, 27. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author | Subject | Active Compound | Route of Treatment | Duration of Treatment | Dosage | Cell Lines/Types | Types | Biological Effects/Findings |
|---|---|---|---|---|---|---|---|---|
| Miller et al., 2007 [69] | Croton palanostigma | Progrado (proanthocy-anidin) | Cell seeding | 72 h | 2 and 10 μg/mL | IL-1β treated human chondrocytes | Monolayer culture | -Reduce glycosaminoglycan release, block cartilage catabolism -Block gelatinolytic activity of MMP2, MMP9 -Promotes production of cartilage repair factor, insulin-like growth factor 1 (IGF-1) |
| Shakibaei et al., 2007 [70] | Not stated | Resveratrol | Cell seeding | 0, 1, 12, and 24 h | 100 μM | IL-1β treated human articular chondrocytes | Monolayer culture | -Increased level of collagen type II and β1-integrin synthesize -Deactivation of caspase-3 and cleavage of PARP |
| Sumantran et al., 2007 [71] | Withania somnifera (root) | Withania somnifera | Cell seeding | 2, 4, 6, and 8 days | 0.05, 0.1, 0.5, or 1 mg/mL | Human explant cartilage | Monolayer culture | -Reduce proteoglycan release -Inhibit collagenase type II |
| Sumantran et al., 2008 [72] | Phyllanthis emblicai (fruit) | Phyllanthis emblicai | Cell seeding | 2,4,6, and 8 days | 0.05 or 0.10 mg/mL | Human explant cartilage | Monolayer culture | -Inhibit hyaluronidase and collagenase type II activity -Reduce sGAG release |
| Sanyacharernkul et al., 2009 [73] | Dregea volubilis Benth. ex Hook. f (roots) | Polyoxypregnane glycoside | Cell seeding | 3 days | 6.25, 12.5, and 25 µg mL−1 | IL-1β treated porcine cartilage | Monolayer culture | -Reduce sGAG, HA, and MMP2 activity -Reverse effect of IL-1β-reduced UA level |
| Liu et al., 2010 [74] | Grape (skin) | Resveratrol | Cell seeding | 24 h | 25, 50, 75, and 100 μM | Advanced glycation end products (AGE)-stimulated OA porcine chondrocytes and cartilage explants | Monolayer culture | -Suppress iNOS-NO and COX-2-PGE2 production -Inhibit IKK-IκBα-NF-κβ and JNK/ERK-activator protein-1 (AP-1) signaling pathways -Decrease MMP13 and prevent destruction of collage type II -Prevent degradation of proteoglycan and aggrecan in explant cartilage |
| Hu et al., 2011 [75] | Hydrastis canadensis (goldenseal), Cortex phellodendri (Huang bai), and Rhizoma coptidis (Huang lian) | Barberine | Cell seeding | 3 days | 10, 25, 50, 100, µm | IL-1β treated rat articular chondrocytes and cartilage explants | Monolayer culture | -Reduce sGAG release -Inhibit NO production -Suppress production of MMP1, MMP3, and MMP13 -Upregulate TIMP-1 -Decrease lesions severity |
| Chai et al., 2012 [76] | Zingiber cassumunar Roxb. | cis-3-(2′,4′,5′-trimethoxyphenyl)-4-{(E)-2′′′,4′′′,5′′′-trimethoxystyryl}cyclohex-1-ene (Compound C) and (E)-4-(3′,4′-dimethoxyphenyl)but-3-en-1-ol (Compound D) | Cell seeding | 3 days | 1, 10, or 100 µM | IL-1β treated porcine cartilage explant | Monolayer culture | -Conserve collagen and UA -Reduce sGAG and HA release -Inhibit MMP2 and MMP13 activity |
| Itthiarbha et al., 2012 [77] | Dregea volubilis | Polyoxypregnane glycoside | Cell seeding | 3 days | 6.25, 12.5, and 25 µg mL−1 | IL-1β treated human articular chondrocytes | Monolayer | -Decrease HA released -Inhibit expression of MMP1, MMP3, and MMP13 -Reverse effect of IL-1β-inhibited type II collagen expression -Inhibit IL-1β-induced HA and MMPs mRNA upregulation -Inhibit phosphorylation of IKK and IκBα degradation of NF-κβ signaling pathway |
| Phitak et al., 2012 [78] | Sesamun indicum Linn. | Sesamin | Cell seeding | 24 h | 0.25, 0.5, and 1.0 μM | IL-1β treated porcine cartilage explants | Monolayer culture | -Inhibit release of sGAG and hydroxyproline -Inhibit loss of UA, prevent proteoglycan degradation -Decrease expression and activity of MMP1, MMP3, and MMP13 -Inhibit p38 and JNK signaling pathway |
| Chua et al., 2013 [79] | Edible bird’s nest (swiftlets’ saliva) | Not stated | Cell seeding | 7 days | 0.05–3.00% | Human articular chondrocytes | Monolayer culture | -Promote human articular cartilage proliferation -Reduce catabolic genes’ expression (MMP1, MMP3, IL-1, IL-6, IL-8, COX-2, iNOS, PGE2) -Increase type II collagen, aggrecan, and transcription factor SOX-9 gene expression -Increase sGAG production |
| Liu et al., 2014 [80] | Resveratrol | Resveratrol | Cell seeding | 24 h | 6.25, 12.5, 25, 50, 100, or 200 μM | IL-1β treated human knee articular chondrocytes | Monolayer culture | -Decreased level of IL-1β and TNFα -Decreased expression of TLR4 mRNA levels -Downregulation of TLR4/NF-κβ signaling pathway -Decreased expression of MyD88 and TRAF6 |
| Tong et al., 2014 [81] | Aconitum carmichaelii Debx. (Fuzi) | Not stated | Cell seeding | 24, 48, and 72 h | 5% to 20% | Mono-iodoacetate (MIA) treated rat chondrocytes | Monolayer culture | -Promote chondrocyte proliferation, inhibit chondrocyte damage |
| Rufino et al., 2014 [82] | Juniperus oxycedrus L. subsp. Oxycedrus (leaves) | α-pinene | Cell seeding | 24 h | 10, 25, 50, 100, 125, and 200 μg/mL | IL-1β treated human articular chondrocytes and human chondrocytes cell line (C28/12) | Monolayer culture | -Inhibit phosphorylation of IκBα, inhibit NF-κβ and JNK activation -Inhibit expression of iNOS, MMP1, and MMP13 genes |
| Pradit et al., 2014 [83] | Phyllanthus amarus Schum. and Thonn. | Phyllanthin and hypophyllanthin | Cell seeding | 14 days | 50, 100, and 200 μg/mL | IL-1β treated porcine articular cartilage explants | Monolayer culture | -Decrease sGAG level and MMP2 activity -Increase UA and proteoglycan contents |
| Wen et al., 2015 [84] | Gallnuts, grapes, tea leaves, and oak bark | Gallic acid | Cell seeding | 48 h | 0, 10, 20, 40, 80 μM | AGE-treated rabbit chondrocytes | Monolayer culture | -Inhibit ROS release -Suppress SOD and GSH activities -Reverse collagen type II and aggrecan degradation -Inhibit expression of iNOS-NO and COX-2-PGE2 |
| Liu et al., 2015 [56] | Rhizoma coptidis (Huang Lian) | Berberine | Cell seeding | 24 h | 1, 3, 10, and 30 ng/mL | Connective tissue growth factor (CNN2)-induced IL-1β expression treated human synovial fibroblast | Monolayer culture | -Attenuate CCN2-induced IL-1β expression -Reduce ROS generation -Reduce phosphorylation of apoptosis signal-regulating kinase 1 (ASK1), p38, JNK, and p65 |
| Park et al., 2015 [85] | Scutellariae radix | Wogonin | Cell seeding | 24 h | 1, 10, 50, or 100 μM | IL-1β treated rabbit articular chondrocytes | Monolayer culture | -Inhibit MMP1, MMP3, MMP13, and ADAMTS4 gene expression -Increase collagen type II expression -Inhibit MMP3 production and activity |
| Zhang et al., 2016 [63] | Turmeric | Curcumin | Cell seeding | 6 h | 100 μM | IL-1β treated human chondrocytes | Monolayer culture | -Suppress mRNA expression of IL-1β, TNF-α, MMP1, MMP3, MMP13, and ADAMTS5 -Upregulate chondroprotective transcriptional regulator, Cbp/p300 interacting transactivator with ED-rich tail 2 (CITED2) -Reduce OA disease progression |
| Buddhachat et al., 2016 [86] | Phyllanthus amarus, Phyllanthus urinaria L., Phyllanthus urinaria subsp. chamaepeuce, Phyllanthus debilis, and Phyllanthus airy-shawii | Phyllanthin and hypophyllanthin | Cell seeding | 2 weeks | 50, 100, and 250 µg/mL. | IL-1β treated porcine cartilage cartilage explants | Monolayer culture | -Inhibit sGAG release -Reduce loss of UA level -Reduce proteoglycan degradation |
| Kong et al., 2016 [87] | Sesame | Sesamin | Cell seeding | 24 h | 2.5, 5, or 10 μM | IL-1β treated human chondrocytes | Monolayer culture | -Inhibit PGE2 and NO production -Inhibit production of MMP1, MMP3, and MMP13 -Block NF-κβ p65 and IκBα phosphorylation -Increase expression of Nrf-2 and HO-1 |
| Liu et al., 2017 [88] | Resveratrol | Resveratrol | Cell seeding | 48 h | 10 µM | IL-1β treated human chondrocytes | Pellet culture | -Increased expression of Sirtuin 1 (Sirt1) -Decreased level of cell apoptosis -Decreased expression of procaspase-3, -9, MMP1, MMP3, MMP13, Wnt3α, Wnt5 α, Wnt7 α, and β-catenin |
| Gu et al., 2017 [89] | Resveratrol | Resveratrol | Cell seeding | 24 h | 0, 6.25, 12.5, 25, 50, 100, and 200 µM | IL-1β treated human chondrocytes | Monolayer culture | -Decreased level of MMP-13 and IL-6 -Suppression of TLR4/MyD88 dependent and independent downstream pathways |
| Khan et al., 2017 [90] | Scutellaria baicalensis (root) | Wogonin | Cell seeding | 2 h | 10, 25, 50, or 100 µM | IL-1β-stimulated human OA chondrocytes and cartilage explants | Monolayer culture | -Inhibit expression of IL-6, iNOS-NO, and COX-2-PGE2 -Suppress ROS production -Reduce MMP3, MMP9, MMP13, and ADAMTS4 mRNA expression and protein production -Enhance collagen type II and aggrecan mRNA and protein expression -Inhibit release of collagen type II and sGAG by cartilage explants -Reduce proteoglycan release from cartilage explants -Increase expression of Nrf-2-dependent antioxidant and cytoprotective enzymes (SOD-2, NQO1, glutamate-cysteine ligase catalytic subunit (GCLC),HO-1) |
| Sun et al., 2019 [59] | Astaxanthin | Astaxanthin | Cell seeding | 2 h | 5, 10, and 20 μM | IL-1β, TNF-α, and tert-butyl hydroperoxide (TBHP) treated chondrocytes | Monolayer culture | -Reduce ADAMTS5, MMP3, and MMP13 mRNA level -Increase expression of collagen type II -Upregulation of nuclear factor (erythroid-derived 2)-like 2 (Nrf-2) signaling and downstream chondroprotective gene, heme oxygenase-1 (HO-1) and reduced nicotinamide adenine dinucleotide phosphate (NADPH) quinone oxidoreductase (NQO1) -Reduce iNOS and COX-2 protein level -Block phosphorylation of ERK and JNK, block MAPK signaling -Inhibit p65 and IκBα phosphorylation, inhibit NF-κβ signaling -Reduce ROS level |
| Liu et al., 2019 [91] | Glycine max (soybean) | Genistein | Cell seeding | 24 h | 5, 10, and 50 μM | IL-1β treated human articular chondrocytes | Monolayer culture | -Inhibit expression of catabolic factors (nitric oxide synthase 2 (NOS2), COX-2, MMP1, MMP2, MMP3, MMP9, and MMP13 -Stimulate HO-1expression in Nrf-2 pathway activation |
| Ongchai et al., 2019 [92] | Senna alata (L.) Roxb. and Senna tora (L.) Roxb. (leaves) | Rhein / aloe-emodin | Cell seeding | 7, 14, and 21 days | 1.25, 2.5, or 5 μg/mL | IL-17α and IL-1β treated porcine cartilage explants | Monolayer culture | -Prevent cartilage degradation -Reduce sGAG and HA release, preserve proteoglycan |
| Heikal et al., 2019 [93] | Stichopus chloronotus (sea cucumber) | Stichopus chloronotus | Cell seeding | 7 days | 0.1, 0.2, 0.3, 0.4, 0.5, 0.6, 0.7, 0.8, 0.9, and 1.0% | Human articular chondrocytes | Monolayer culture | -Maintain chondrocytes characteristics -Increase collagen type II, aggrecan, and SOX-9 gene expression -Reduce gene expression of collagen type I, IL-1, IL-6, IL-8, MMP1, MMP3, MMP13, COX-2, iNOS, and proteinase-activated receptor 2 (PAR-2) -Increase production of sGAG -Reduce production of PGE2 and NO |
| D’Ascola et al., 2019 [94] | Curcumin | Curcumin | Cell seeding | 16 h | 0.65, 1.25, 2.5, 5, and 10 µg/mL | Human articular chondrocytes | Monolayer culture | -Decreased expression of IL-1β -Decreased expression of NF-κβ and STAT3 -Increased expression of collagen type II |
| Zhang et al., 2020 [67] | Aconitum carmichaelii Debeaux derived Hei-shun-pian (Hsp) | Benzoylacon-itine and benzoylhypa-conitine | Cell seeding | 24, 48, and 72 h | 2.5, 5, 10, 15% | TNF-α treated rat articular chondrocytes | Monolayer culture | -Stimulate chondrocytes proliferation -Regulate collagen type II, MMP1, ADAMTS9, and aggrecan gene expression |
| Lee et al., 2020 [55] | Cynaroside | Cynaroside | Cell seeding | 24 h | 0, 40, 80, and 160μM | IL-1β treated chondrocytes | Monolayer culture | -Inhibit expression of catabolic factors (nitrite, iNOS, ROS, PGE2, COX-2, MMP1, MMP3, MMP13, ADAMTS4) -Inhibit degradation of anabolic factors (collage type II and aggrecan) -Suppress phosphorylation of MAPKS and translocation of NF-κβ p65 subunit -Inhibit production of nitrite and PGE2 -Suppress protein expression of iNOS, COX-2, MMP13, and ADAMTS4 -Inhibit proteoglycan degradation |
| Lee et al., 2020 [95] | Artemisia argyi (seomae mugwort) | Jaceosidin | Cell seeding | 24 h | 10, 50, and 100 μg/mL | In vitro—IL-1β, IL-6 and TNF-α treated chondrocytes | Monolayer culture | -Suppress MMP3, MMP13, ADAMTS4, and ADAMTS5 expression -Reduce aggrecanase activity -Protect against sulfated proteoglycan degradation |
| You et al., 2021 [96] | Illicium verum | Shikimic acid (SA) | Cell seeding | 24 h | 0.1, 1, 5, 10, and 20 mM | IL-1β treated human chondrocytes (SW1353) | Monolayer culture | -Decrease expression of iNOS, COX-2, MMP3, MMP13, ADAMTS5, collagen type X and p63 -Increase collagen type II, autophagy related 7 (ATG7), beclin-1, and microtubule-associated light chain-3 (LC3) expression -Increase autophagic flux -Suppress activation of MAPK and NF-κβ pathway |
| Buhrmann et al., 2020 [97] | Curcumin | Curcumin | Cell seeding | 10 days | 1, 2, 5, and 10 μM | 3D-chondrocyte-alginate beads | Monolayer culture | -Suppress activation of NF-κβ pathway -Increased expression of collagen type II-, cartilage-specific proteoglycans, β1-integrin, and Sox9 -Downregulation of MMP-9, cox-2, and caspase-3 |
| Xu et al., 2022 [98] | Curcumin | Curcumin | Cell seeding | 2 days | 10 µM/L | Tert-butyl hydroperoxide (TBHP) treated C57 mice chondrocytes | Monolayer culture | -Increased chondrocytes proliferation -Increased extracellular matrix anabolism -Decreased level of (aggrecan, collagen II, ADAMTS5, MMP13, IL-1β, and TNFα -Decreased level of 8-OHdG positive cells and MDA |
| Author | Subject | Active Compound | Route of Treatment | Duration of Treatment | Dosage | Model Type | Anatomical Location | Biological Effects / Findings |
|---|---|---|---|---|---|---|---|---|
| El-Ghazaly et al., 2011 [52] | Propolis | Propolis | Intraarticular injection | Once daily for 2 weeks | 5 mL/kg | Male wistar rats injected with Freud’s adjuvant injected | Right hind paw | -Reduce breakdown of cartilage -Prevent reduction sulfated glycosaminoglycan (sGAG) and hydroxyproline level -Reduce TNF-α level -Normalize NO and the oxidative stress biomarkers (GSH, MDA) |
| Hu et al., 2011 [75] | Hydrastis canadensis (goldenseal), Cortex phellodendri (Huang bai), and Rhizoma coptidis (Huang lian) | Barberine | Intraarticular injection | Not specified | 10, 50, or 100 µm | Male SD rats injected with IL-1β | Right knee joint | -Reduce MMP1, MMP3, and MMP13 gene expression and protein production -Increase TIMP-1 mRNA levels and protein expression |
| Wang et al., 2011 [99] | Resveratrol | Resveratrol | Intraarticular injection | 2 weeks | 50, 20, and 10 µmol/kg | 1.5 to 2.5 kg male and female rabbits subjected to medial parapatellar arthrotomy | Right knee | -Decreased level of proteoglycan loss -Decreased level of cell apoptosis in chondrocytes -Decreased level of nitric oxide |
| Phitak et al., 2012 [78] | Sesamun indicum Linn. | Sesamin | Intraarticular injection | 5 days for 5 weeks | 1 μM or 10 μM | 8-week-old Wistar rats injected with papain | Right knee | -Reverse pathological changes -Reduce chondrocyte disorganization -Increase cartilage thickness -Decrease collagen type II and proteoglycan loss -Increase formation of collagen type II and proteoglycan |
| Leong et al., 2014 [64] | Green tea | Epigallocatechin 3-gallate (EGCG) | Intraperitoneal injection | 4 weeks | 25 mg/kg | 5- to 6-month-old male SD rats surgically transecting the medial meniscotibial ligament (MMTL) | Destabilization of the medial meniscus (DMM) at the right hind limb | -Slows progression of early and midstage OA -Reduce degradation of aggrecan and collagen type II -Reduce MMP13 and ADAMTS5 -Reduce MMP1, MMP3, MMP8, MMP13, ADAMTS5, IL-1β, and TNF-α mRNA levels -Elevate gene expression of CITED2 -Reduce expression of IL-1β, TNF-α, and chemokine receptor Ccr2 in dorsal root ganglion (DRG) |
| Hanprasertpong et al., 2014 [66] | Cryptolepis buchanani Roem. and Schult. | Cryptolepis buchanani | Intraperitoneal injection | 5 h | 100, 250, and 500 mg/kg | Male SD rats and Swiss albino mice injected with carrageenan | Hind paw | -Reduced MMP-2 activity |
| Tong et al., 2014 [81] | Aconitum carmichaelii Debx (Fuzi) | Not stated | Oral administration | 28 days | Daily 14 g/kg | Male rats injected with MIA | Right knee joint | -Prevent alteration in joint morphology (osteolysis, swelling, and irregular catilage surface) and bone density -Prevent subchondral abonormalities (loss of cartilage, cyst, fibrillation of bone marrow, subchondral sclerosis, and disorganized chondrocutes culture) -Show chondroprotective activity |
| Jeong et al., 2015 [68] | Artemsia princeps Pampanini | Eupatilin | Oral administration | 14 days | 100 mg/kg | 6-week-old male Wistar rats injected with MIA | Patellar ligament of right knee | -Reduce osteoclasts number -Reduce expression of IL-1β, IL-6, nitrotyrosine, iNOS -Reduce mRNA of MMP3, MMP13, and ADAMTS5 by increasing tissue inhibitor of metalloproteinases-1 (TIMP-1) -Inhibit phosphorylation of JNK protein -Suppress oxidative damage, enhance ECM production |
| Wen et al., 2015 [84] | Gallnuts, grapes, tea leaves, and oak bark | Gallic acid | Intra-articular injection | Once every 72 h for 8 weeks | 80 μM | Rabbits injected with collagenase type II | Right knee | -Reduce cartilage degradation |
| Liu et al., 2015 [56] | Rhizoma coptidis (Huang Lian) | Berberine | Intraperitoneal injections | 30 days | 10 or 30 mg/kg | Rats injected with collagenase | Right knee joint | -Inhibit OA development -Lower IL-1β serum level -Reduce proteoglycan break down |
| Park et al., 2015 [85] | Scutellariae radix | Wogonin | Intraarticular injection | 7 h | 50 or 100 μM | Male SD rats injected with IL-1β | Right knee joint | -Inhibit production of MMP3 |
| Zhang et al., 2016 [63] | Turmeric | Curcumin | Oral administration | 8 weeks | 50 mg/kg | 5- to 6-week-old male mice subjected to MMTL surgery | DMM at the right hind limb | -Reduce proteoglycan loss and cartilage erosion -Reduce synovitis and subchondral plate thickness -Reduce type II collagen and aggrecan cleavage and MMP13 and ADAMTS positive chondrocytes |
| Gu et al., 2016 [100] | Resveratrol | Resveratrol | Oral administration | 12 weeks | 5, 22.5, or 45 mg/kg | 7-week-old male C57BL/6J mice high fat diet (HFD) induced OA | Not specified | -Improvement in arrangement of matrix, and tide lines in chondrocytes -Decreased level of CTX-II -Increased expression of type II collagen -Decreased level of TUNEL-positive cells |
| Wei et al., 2017 [101] | Resveratrol | Resveratrol | Oral administration | 8 weeks | 50 mg/kg | 6-week-old male Wistar rats subjected to MIA injection | Patellar ligament of right knee | -Decreased expression of TNF-α, IL-1β, IL-6, and IL-18 -Suppression of iNOS, NF-κβ, p-AMPK, and SIRT1 protein -Increased level of SOD -Decreased level of MDA -Increased leve of HO-1 and Nrf-2 -Decreased activity of caspase-3/9 |
| Zhang et al., 2017 [102] | Curcumin | Curcumin | Oral administration | 8 weeks | 50 mg/kg | 8-week-old male C57BL/6 mice subjected to MMTL surgery | DMM at right knee | -Decreased cartilage loss and chondrocytes apoptotis -Suppression of caspase-3 expression -Decreased expression of LC-I/II and beclin 1 -Decreased expression of mTOR, Akt, and p-P70S6k |
| Jiang et al., 2017 [103] | Resveratrol | Resveratrol | Oral administration | 12 weeks | 22.5 or 50 mg/kg | 7-week-old male C57BL/6J mice HFD induced OA | Not specified | -Increased cartilage thickness and improvement on chrondrocytes arrangements -Decrease expression of TLR4 and tumor necrosis factor receptor-associated factor 6 (TRAF6) |
| Sun et al., 2017 [104] | Curcumin | Curcumin | Intraperitoneal injection | 8 weeks | 50 mM | Male C57BL/6 mice subjected to MMTL surgery | DMM | -Decreased level of IL-1b, IFN-g, IL-17A, IL-18, TNF-a, and VCAM1 -Increased interaction of pro-caspase-1 to NLRP3 -Decreased level of caspase-1 activation |
| Lee et al., 2018 [54] | Anthriscus sylvestris (leaves) | Anthriscus sylvestris | Oral administration | 8 weeks | 50, 100, and 200 mg/kg body | 8-week-old male rats subjected to MMTL surgery | DMM at right knee | -Inhibit cartilage destruction and proteolycan loss |
| Liu et al., 2019 [91] | Glycine max (soybean) | Genistein | Oral administration | 12 weeks | 40 mg/kg | 8-weeks-old male SD rats subjected to anterior cruciate ligament transection (ACLT) | Not specified | -Slow down progression of OA |
| Yan et al., 2019 [105] | Curcumin | Curcumin | Intra-articular injection | 8 weeks | 50 μl | Male SD rats subjected to ACLT | Right knee | -Suppression of TLR4 activation -Decreased level of IL-1β and TNFα -Suppression of NF-κβ |
| Feng et al., 2019 [106] | Curcumin | Curcumin | Intraperitoneal injection | 8 weeks | 50 and 150 mg/kg | 8-week-old male SD rats subjected ACLT | Right knee | -Downregulation of caspase3 and PARP protein -Increased expression of SIRT1 -Reverse the loss of chondrocytes and proteoglycan |
| Jiang et al., 2020 [107] | Curcumin | Curcumin | Intra-articular injection | 4 weeks | 40 μM | 8-week-old male SD rats injected with Freud’s adjuvant | Temporomandibular joint osteoarthritis | -Decreased expression of mRNA levels of COL2A1 and ACAN -Increased level of Nrf2 and p-Nrf2 -Decreased level of proteoglycan loss -Decreased level of iNOS, COX-2, IL-1β, MMP-9, and MMP-13 |
| Susmiarsih et al., 2019 [108] | Green Tea | EGCG | Oral administration | 8 weeks | 200 mg/kg | Rabbits injected with Freud’s adjuvant | Not specified | -Decrease degree of knee joint damage -Decrease NO level |
| Sun et al., 2019 [59] | Astaxanthin | Astaxanthin | Intra-articular injection | 2 times a week for 8 weeks | 20 mg/kg | 8-week-old male C57BL/6 mice subjected to MMTL to surgery | DMM at the right knee | -Reduce cartilage degradation |
| Lee et al., 2020 [95] | Artemisia argyi (seomae mugwort) | Jaceosidin | Oral administration | 10 weeks | 50, 100, and 250 mg/kg | 10-week-old male C57BL/6 mice subjected to MMTL surgery | DMM | -Suppress cartilage destruction and reduce subchondral bone thickness |
| Zhang et al., 2020 [67] | Aconitum carmichaelii Debeaux derived Hei-shun-pian (Hsp) | Benzoylacon-itine and benzoylhypa-conitine | Oral administration | 28 days | 14 g/kg dHsp | Male and female rats injceted with MIA | Patellar ligament of rat knee | -Prevent articular degradation -Restore abnormal expression of collagen type X, MMP2, SOX-5, ADAMTS4, ADAMTS5 and ADAMTS9 -Increase collagen type II expression |
| Huang et al., 2021 [50] | Propolis | Galangin | Intra-articular injection | 8 weeks | 5 mg/ml | 8-week-old male rats subjected to ACLT | Right knee | -Reduce cartilage degradation |
| Su et al., 2021 [51] | Galangin | Galangin | Oral administration | 14 days | 10 and 100 mg/kg | Male SD rats induced with MIA | Left femorotibial joint | -Reduce urinary type II collagen -Reduce reactive oxygen species (ROS), lipid peroxidation, IL-1β, IL-6, and TNF-α -Increase catalase, superoxide dismutase (SOD), glutathione peroxidase (Gpx), and reduce glutathione (GSH) level -Reduce collagen type II mRNA and protein expression level indicate inhibit cartilage degradation |
| You et al., 2021 [96] | Illicium verum | Shikimic acid (SA) | Intra-articular injection | 2 times a week for 1 month | 20 mM in 100 µL | 6-week-old SD male rats subjected to ACLT | Right knee | -Mitigates cartilage degradation -Alleviate OA progression |
| Xu et al., 2022 [98] | Curcumin | Curcumin | Intra-articular injection | Biweekly for 4 weeks | 1 × 109 p/mL | Male C57 mice subjected to ACLT | Right knee | -Alleviate cartiage degeneration and abrasion -Decreased level of aggrecan and collagen II -Decreased level of cleaved caspase 3 and 8-OHdG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.T.; Yunus, M.H.M.; Ugusman, A.; Yazid, M.D. Natural Compounds Affecting Inflammatory Pathways of Osteoarthritis. Antioxidants 2022, 11, 1722. https://doi.org/10.3390/antiox11091722
Lee YT, Yunus MHM, Ugusman A, Yazid MD. Natural Compounds Affecting Inflammatory Pathways of Osteoarthritis. Antioxidants. 2022; 11(9):1722. https://doi.org/10.3390/antiox11091722
Chicago/Turabian StyleLee, Yi Ting, Mohd Heikal Mohd Yunus, Azizah Ugusman, and Muhammad Dain Yazid. 2022. "Natural Compounds Affecting Inflammatory Pathways of Osteoarthritis" Antioxidants 11, no. 9: 1722. https://doi.org/10.3390/antiox11091722
APA StyleLee, Y. T., Yunus, M. H. M., Ugusman, A., & Yazid, M. D. (2022). Natural Compounds Affecting Inflammatory Pathways of Osteoarthritis. Antioxidants, 11(9), 1722. https://doi.org/10.3390/antiox11091722