

Transcriptomic Analysis of Taar5 Expression and Co-Expression Networks in the Cerebellum During Perinatal Development

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Public Transcriptomic Data
2.2. Data Normalization and Statistical Analysis
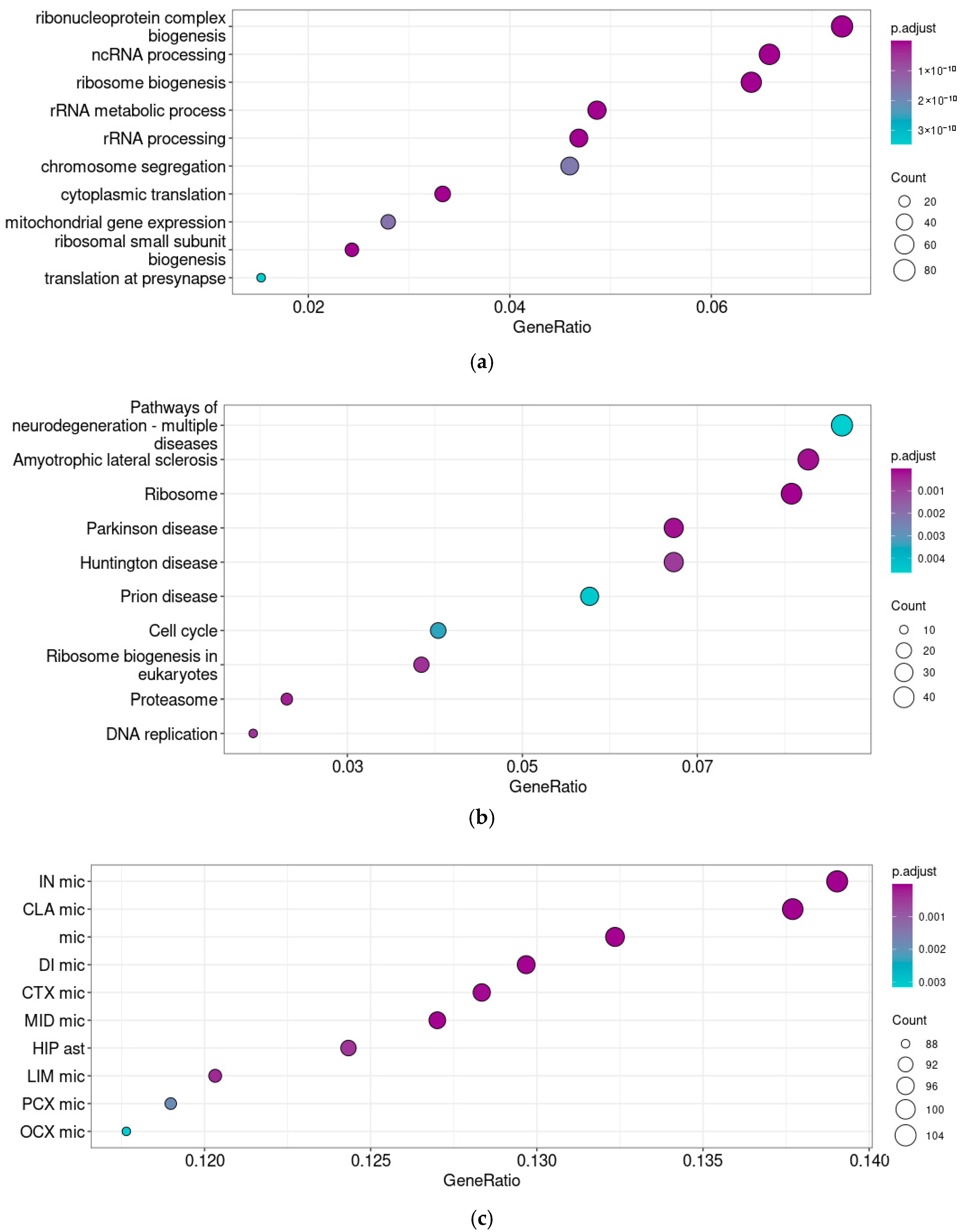
2.3. Gene Co-Expression Measurement and Pathway Enrichment Analysis
2.4. Taar5 mRNA Expression Analysis by RT-PCR
3. Results
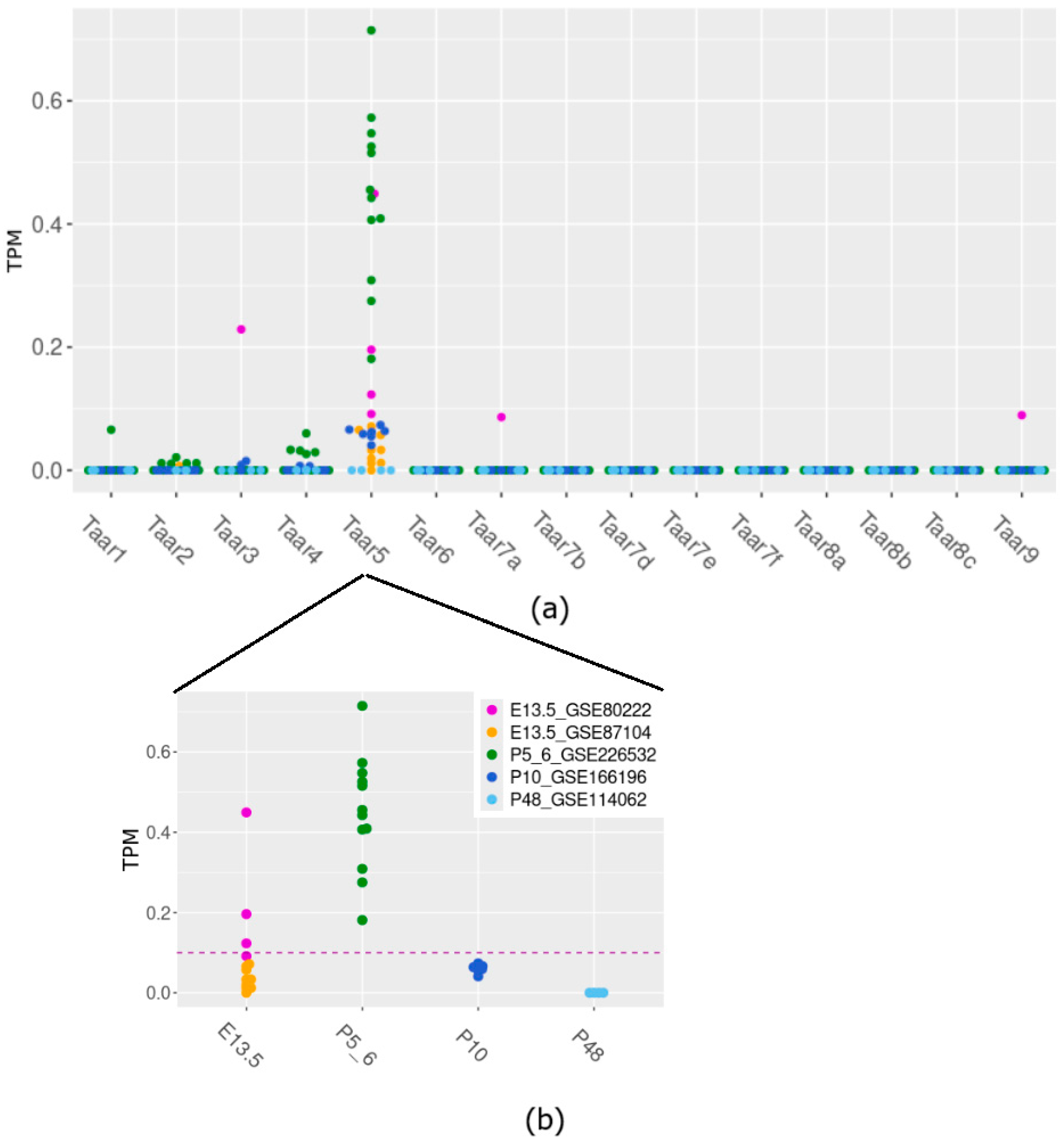
3.1. Taar5 mRNA Expression in Perinatal Mouse Cerebellum Samples in RNA-Seq-Generated GEO Data
3.2. Taar5 Functional Associations in Prenatal Cerebellum Samples
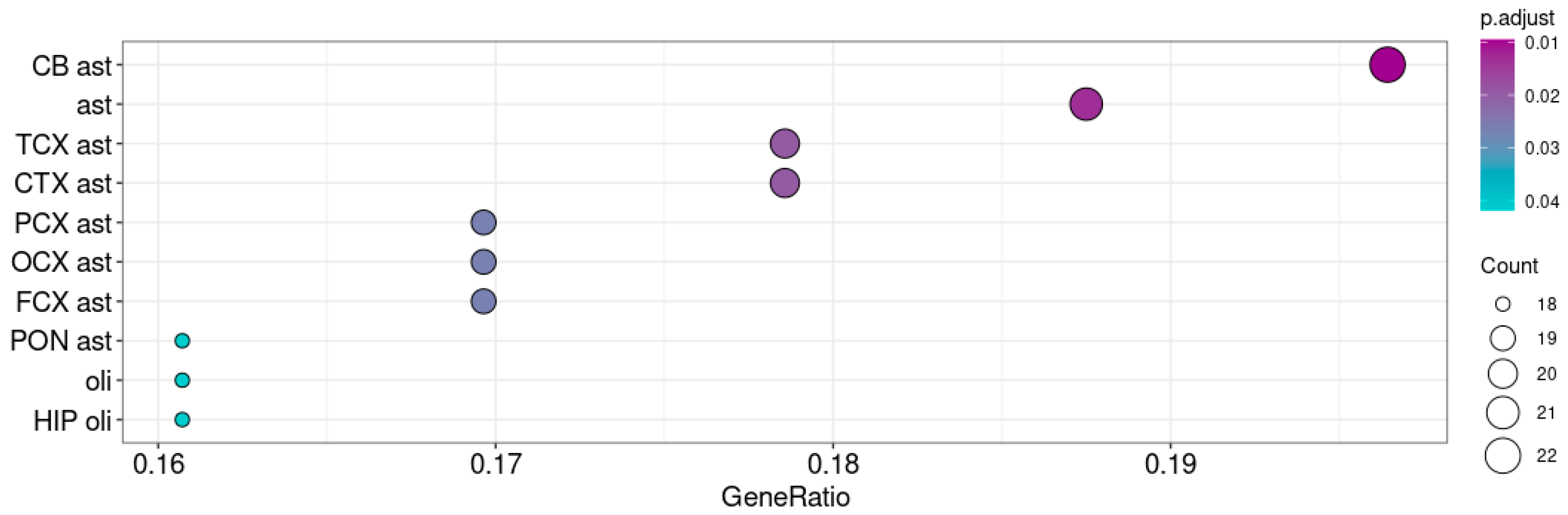
3.3. Taar5 Functional Associations in Postnatal P5–6 Cerebellum Samples
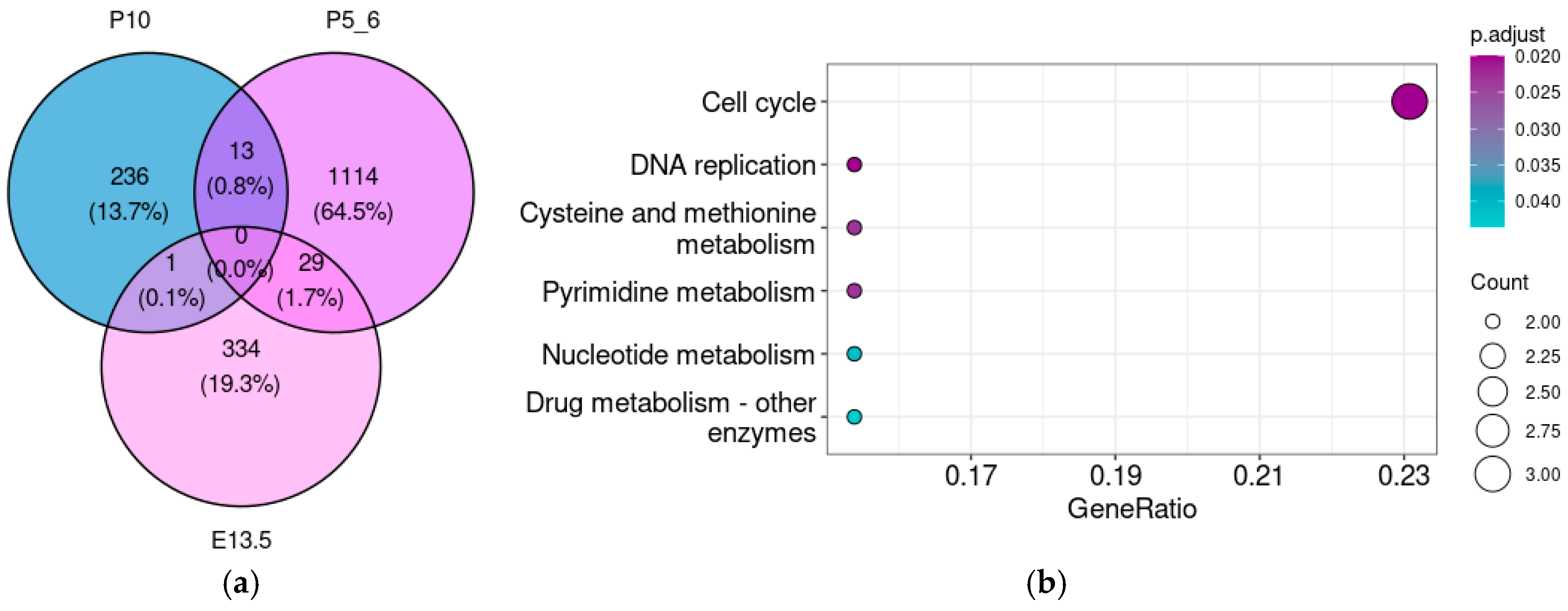
3.4. Taar5 Functional Associations in Postnatal P10 Cerebellum Samples
3.5. The Taar5 Co-Expressed Gene Pattern Gradually Changes from the Embryonal to Postnatal Stages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Efimova, E.V.; Kozlova, A.A.; Razenkova, V.; Katolikova, N.V.; Antonova, K.A.; Sotnikova, T.D.; Merkulyeva, N.S.; Veshchitskii, A.S.; Kalinina, D.S.; Korzhevskii, D.E.; et al. Increased Dopamine Transmission and Adult Neurogenesis in Trace Amine-Associated Receptor 5 (TAAR5) Knockout Mice. Neuropharmacology 2021, 182, 108373. [Google Scholar] [CrossRef] [PubMed]
- Efimova, E.V.; Kuvarzin, S.R.; Mor, M.S.; Katolikova, N.V.; Shemiakova, T.S.; Razenkova, V.; Ptukha, M.; Kozlova, A.A.; Murtazina, R.Z.; Smirnova, D.; et al. Trace Amine-Associated Receptor 2 Is Expressed in the Limbic Brain Areas and Is Involved in Dopamine Regulation and Adult Neurogenesis. Front. Behav. Neurosci. 2022, 16, 847410. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, S.; Ghisi, V.; Emanuele, M.; Leo, D.; Sukhanov, I.; Sotnikova, T.D.; Chieregatti, E.; Gainetdinov, R.R. Postsynaptic D2 Dopamine Receptor Supersensitivity in the Striatum of Mice Lacking TAAR1. Neuropharmacology 2015, 93, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, S.; Salahpour, A.; Masri, B.; Sotnikova, T.D.; Messa, M.; Barak, L.S.; Caron, M.G.; Gainetdinov, R.R. Functional Interaction between Trace Amine-Associated Receptor 1 and Dopamine D2 Receptor. Mol. Pharmacol. 2011, 80, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Borowsky, B.; Adham, N.; Jones, K.A.; Raddatz, R.; Artymyshyn, R.; Ogozalek, K.L.; Durkin, M.M.; Lakhlani, P.P.; Bonini, J.A.; Pathirana, S.; et al. Trace Amines: Identification of a Family of Mammalian G Protein-Coupled Receptors. Proc. Natl. Acad. Sci. USA 2001, 98, 8966–8971. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.V.; Sarkar, P.K. Nongenomic Roles of Thyroid Hormones and Their Derivatives in Adult Brain: Are These Compounds Putative Neurotransmitters? Front. Endocrinol. 2023, 14, 1210540. [Google Scholar] [CrossRef] [PubMed]
- Kalinina, D.S.; Ptukha, M.A.; Goriainova, A.V.; Merkulyeva, N.S.; Kozlova, A.A.; Murtazina, R.Z.; Shemiakova, T.S.; Kuvarzin, S.R.; Vaganova, A.N.; Volnova, A.B.; et al. Role of the Trace Amine Associated Receptor 5 (TAAR5) in the Sensorimotor Functions. Sci. Rep. 2021, 11, 23092. [Google Scholar] [CrossRef] [PubMed]
- Polini, B.; Ricardi, C.; Bertolini, A.; Carnicelli, V.; Rutigliano, G.; Saponaro, F.; Zucchi, R.; Chiellini, G. T1AM/TAAR1 System Reduces Inflammatory Response and β-Amyloid Toxicity in Human Microglial HMC3 Cell Line. Int. J. Mol. Sci. 2023, 24, 11569. [Google Scholar] [CrossRef] [PubMed]
- Bellusci, L.; Laurino, A.; Sabatini, M.; Sestito, S.; Lenzi, P.; Raimondi, L.; Rapposelli, S.; Biagioni, F.; Fornai, F.; Salvetti, A.; et al. New Insights into the Potential Roles of 3-Iodothyronamine (T1AM) and Newly Developed Thyronamine-Like TAAR1 Agonists in Neuroprotection. Front. Pharmacol. 2017, 8, 905. [Google Scholar] [CrossRef] [PubMed]
- Gainetdinov, R.R.; Hoener, M.C.; Berry, M.D. Trace Amines and Their Receptors. Pharmacol. Rev. 2018, 70, 549–620. [Google Scholar] [CrossRef] [PubMed]
- Cichero, E.; Espinoza, S.; Tonelli, M.; Franchini, S.; Gerasimov, A.S.; Sorbi, C.; Gainetdinov, R.R.; Brasili, L.; Fossa, P. A Homology Modelling-Driven Study Leading to the Discovery of the First Mouse Trace Amine-Associated Receptor 5 (TAAR5) Antagonists. MedChemComm 2016, 7, 353–364. [Google Scholar] [CrossRef]
- Dinter, J.; Mühlhaus, J.; Wienchol, C.L.; Yi, C.-X.; Nürnberg, D.; Morin, S.; Grüters, A.; Köhrle, J.; Schöneberg, T.; Tschöp, M.; et al. Inverse Agonistic Action of 3-Iodothyronamine at the Human Trace Amine-Associated Receptor 5. PLoS ONE 2015, 10, e0117774. [Google Scholar] [CrossRef] [PubMed]
- Vaganova, A.N.; Fesenko, Z.S.; Efimova, E.V.; Chekrygin, S.A.; Shafranskaya, D.D.; Prjibelski, A.D.; Katolikova, N.V.; Gainetdinov, R.R. Knocking Out TAAR5: A Pathway to Enhanced Neurogenesis and Dopamine Signaling in the Striatum. Cells 2024, 13, 1910. [Google Scholar] [CrossRef] [PubMed]
- Amore, G.; Spoto, G.; Ieni, A.; Vetri, L.; Quatrosi, G.; Di Rosa, G.; Nicotera, A.G. A Focus on the Cerebellum: From Embryogenesis to an Age-Related Clinical Perspective. Front. Syst. Neurosci. 2021, 15, 646052. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Heu, J.; Hoener, M.C.; Kilduff, T.S. Wakefulness Induced by TAAR1 Partial Agonism in Mice Is Mediated Through Dopaminergic Neurotransmission. Int. J. Mol. Sci. 2024, 25, 11351. [Google Scholar] [CrossRef] [PubMed]
- De Benedictis, A.; Rossi-Espagnet, M.C.; de Palma, L.; Carai, A.; Marras, C.E. Networking of the Human Cerebellum: From Anatomo-Functional Development to Neurosurgical Implications. Front. Neurol. 2022, 13, 806298. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; LeBel, A.; D’Mello, A.M. Ignoring the Cerebellum Is Hindering Progress in Neuroscience. Trends Cogn. Sci. 2025, 29, 318–330. [Google Scholar] [CrossRef] [PubMed]
- Pierce, J.E.; Péron, J. The Basal Ganglia and the Cerebellum in Human Emotion. Soc. Cogn. Affect. Neurosci. 2020, 15, 599–613. [Google Scholar] [CrossRef] [PubMed]
- Locke, T.M.; Soden, M.E.; Miller, S.M.; Hunker, A.; Knakal, C.; Licholai, J.A.; Dhillon, K.S.; Keene, C.D.; Zweifel, L.S.; Carlson, E.S. Dopamine D1 Receptor–Positive Neurons in the Lateral Nucleus of the Cerebellum Contribute to Cognitive Behavior. Biol. Psychiatry 2018, 84, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.J.; Luo, L. Neocortex–Cerebellum Circuits for Cognitive Processing. Trends Neurosci. 2020, 43, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Kruithof, E.S.; Klaus, J.; Schutter, D.J.L.G. The Cerebellum in Aggression: Extending the Cortico-Limbic Dual-Route Model of Motivation and Emotion. Motiv. Sci. 2022, 8, 150–160. [Google Scholar] [CrossRef]
- Olson, I.R.; Hoffman, L.J.; Jobson, K.R.; Popal, H.S.; Wang, Y. Little Brain, Little Minds: The Big Role of the Cerebellum in Social Development. Dev. Cogn. Neurosci. 2023, 60, 101238. [Google Scholar] [CrossRef] [PubMed]
- Martí-Clúa, J. Natural Apoptosis in Developing Mice Dopamine Midbrain Neurons and Vermal Purkinje Cells. Folia Neuropathol. 2016, 54, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Ciafrè, S.; Ferraguti, G.; Greco, A.; Polimeni, A.; Ralli, M.; Ceci, F.M.; Ceccanti, M.; Fiore, M. Alcohol as an Early Life Stressor: Epigenetics, Metabolic, Neuroendocrine and Neurobehavioral Implications. Neurosci. Biobehav. Rev. 2020, 118, 654–668. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Yu, D.; Li, X.; Huang, J.; Jing, S.; Bao, X.; Yang, T.; Fan, X. Propofol Exposure in Early Life Induced Developmental Impairments in the Mouse Cerebellum. Front. Cell. Neurosci. 2017, 11, 373. [Google Scholar] [CrossRef] [PubMed]
- Topper, L.A.; Baculis, B.C.; Valenzuela, C.F. Exposure of Neonatal Rats to Alcohol Has Differential Effects on Neuroinflammation and Neuronal Survival in the Cerebellum and Hippocampus. J. Neuroinflammation 2015, 12, 160. [Google Scholar] [CrossRef] [PubMed]
- Mapelli, L.; Soda, T.; D’Angelo, E.; Prestori, F. The Cerebellar Involvement in Autism Spectrum Disorders: From the Social Brain to Mouse Models. Int. J. Mol. Sci. 2022, 23, 3894. [Google Scholar] [CrossRef] [PubMed]
- van Essen, M.J.; Nayler, S.; Becker, E.B.E.; Jacob, J. Deconstructing Cerebellar Development Cell by Cell. PLOS Genet. 2020, 16, e1008630. [Google Scholar] [CrossRef] [PubMed]
- Carter, R.A.; Bihannic, L.; Rosencrance, C.; Hadley, J.L.; Tong, Y.; Phoenix, T.N.; Natarajan, S.; Easton, J.; Northcott, P.A.; Gawad, C. A Single-Cell Transcriptional Atlas of the Developing Murine Cerebellum. Curr. Biol. 2018, 28, 2910–2920.e2. [Google Scholar] [CrossRef] [PubMed]
- Locke, T.M.; Fujita, H.; Hunker, A.; Johanson, S.S.; Darvas, M.; du Lac, S.; Zweifel, L.S.; Carlson, E.S. Purkinje Cell-Specific Knockout of Tyrosine Hydroxylase Impairs Cognitive Behaviors. Front. Cell. Neurosci. 2020, 14, 228. [Google Scholar] [CrossRef] [PubMed]
- Flace, P.; Livrea, P.; Basile, G.A.; Galletta, D.; Bizzoca, A.; Gennarini, G.; Bertino, S.; Branca, J.J.V.; Gulisano, M.; Bianconi, S.; et al. The Cerebellar Dopaminergic System. Front. Syst. Neurosci. 2021, 15, 650614. [Google Scholar] [CrossRef] [PubMed]
- Leo, D.; Mus, L.; Espinoza, S.; Hoener, M.C.; Sotnikova, T.D.; Gainetdinov, R.R. Taar1-Mediated Modulation of Presynaptic Dopaminergic Neurotransmission: Role of D2 Dopamine Autoreceptors. Neuropharmacology 2014, 81, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Cutando, L.; Puighermanal, E.; Castell, L.; Tarot, P.; Belle, M.; Bertaso, F.; Arango-Lievano, M.; Ango, F.; Rubinstein, M.; Quintana, A.; et al. Cerebellar Dopamine D2 Receptors Regulate Social Behaviors. Nat. Neurosci. 2022, 25, 900–911. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Sakai, M.; Nagatsu, I. Immunohistochemical Demonstration of Expression of Tyrosine Hydroxylase in Cerebellar Purkinje Cells of the Human and Mouse. Neurosci. Lett. 1994, 165, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Gene Ontology Consortium; Aleksander, S.A.; Balhoff, J.; Carbon, S.; Cherry, J.M.; Drabkin, H.J.; Ebert, D.; Feuermann, M.; Gaudet, P.; Harris, N.L.; et al. The Gene Ontology Knowledgebase in 2023. Genetics 2023, 224, iyad031. [Google Scholar] [CrossRef] [PubMed]
- Vera Alvarez, R.; Pongor, L.S.; Mariño-Ramírez, L.; Landsman, D. TPMCalculator: One-Step Software to Quantify mRNA Abundance of Genomic Features. Bioinformatics 2019, 35, 1960–1962. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.A.; Krishnan, A. Robust Normalization and Transformation Techniques for Constructing Gene Coexpression Networks from RNA-Seq Data. Genome Biol. 2022, 23, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, M.-C.; Konaté, M.M.; Chen, L.; Das, B.; Karlovich, C.; Williams, P.M.; Evrard, Y.A.; Doroshow, J.H.; McShane, L.M. TPM, FPKM, or Normalized Counts? A Comparative Study of Quantification Measures for the Analysis of RNA-Seq Data from the NCI Patient-Derived Models Repository. J. Transl. Med. 2021, 19, 269. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Dai, J.; Lu, H.; Zhao, H. Comprehensive Analysis of Ubiquitously Expressed Genes in Humans from A Data-Driven Perspective. Genomics Proteomics Bioinformatics 2023, 21, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Wu, Y.; Shi, Q.; Wu, J.; Kong, D.; Wu, X.; He, X.; Liu, T.; Li, S. Systematic Characterization of Cancer Transcriptome at Transcript Resolution. Nat. Commun. 2022, 13, 6803. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the Unification of Biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A Universal Enrichment Tool for Interpreting Omics Data. Innov. Camb. Mass 2021, 2, 100141. [Google Scholar] [CrossRef] [PubMed]
- Tipney, H.; Hunter, L. An Introduction to Effective Use of Enrichment Analysis Software. Hum. Genomics 2010, 4, 202. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, A.T.; Wang, M.; Hauberg, M.E.; Fullard, J.F.; Kozlenkov, A.; Keenan, A.; Hurd, Y.L.; Dracheva, S.; Casaccia, P.; Roussos, P.; et al. Brain Cell Type Specific Gene Expression and Co-Expression Network Architectures. Sci. Rep. 2018, 8, 8868. [Google Scholar] [CrossRef] [PubMed]
- Liberles, S.D. Trace Amine-Associated Receptors Are Olfactory Receptors in Vertebrates. Ann. N. Y. Acad. Sci. 2009, 1170, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Susetyo, A.; Ishii, S.; Fujiwara, Y.; Amano, I.; Koibuchi, N. Histone Deacetylase 3 Inhibitor Alleviates Cerebellar Defects in Perinatal Hypothyroid Mice by Stimulating Histone Acetylation and Transcription at Thyroid Hormone-Responsive Gene Loci. Int. J. Mol. Sci. 2022, 23, 7869. [Google Scholar] [CrossRef] [PubMed]
- Tsukue, N.; Watanabe, M.; Kumamoto, T.; Takano, H.; Takeda, K. Perinatal Exposure to Diesel Exhaust Affects Gene Expression in Mouse Cerebrum. Arch. Toxicol. 2009, 83, 985–1000. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.A.; Tsai, L.; Roy, D.S.; Valenzuela, D.H.; Mosley, C.; Magklara, A.; Lomvardas, S.; Liberles, S.D.; Barnea, G. Neurons Expressing Trace Amine-Associated Receptors Project to Discrete Glomeruli and Constitute an Olfactory Subsystem. Proc. Natl. Acad. Sci. USA 2012, 109, 13410–13415. [Google Scholar] [CrossRef] [PubMed]
- Holloway, K.N.; Douglas, J.C.; Rafferty, T.M.; Majewska, A.K.; Kane, C.J.M.; Drew, P.D. Ethanol-Induced Cerebellar Transcriptomic Changes in a Postnatal Model of Fetal Alcohol Spectrum Disorders: Focus on Disease Onset. Front. Neurosci. 2023, 17, 1154637. [Google Scholar] [CrossRef] [PubMed]
- Katolikova, N.V.; Vaganova, A.N.; Shafranskaya, D.D.; Efimova, E.V.; Malashicheva, A.B.; Gainetdinov, R.R. Expression Pattern of Trace Amine-Associated Receptors during Differentiation of Human Pluripotent Stem Cells to Dopaminergic Neurons. Int. J. Mol. Sci. 2023, 24, 15313. [Google Scholar] [CrossRef] [PubMed]
- Vaganova, A.N.; Katolikova, N.V.; Murtazina, R.Z.; Kuvarzin, S.R.; Gainetdinov, R.R. Public Transcriptomic Data Meta-Analysis Demonstrates TAAR6 Expression in the Mental Disorder-Related Brain Areas in Human and Mouse Brain. Biomolecules 2022, 12, 1259. [Google Scholar] [CrossRef] [PubMed]
- Sepp, M.; Leiss, K.; Murat, F.; Okonechnikov, K.; Joshi, P.; Leushkin, E.; Spänig, L.; Mbengue, N.; Schneider, C.; Schmidt, J.; et al. Cellular Development and Evolution of the Mammalian Cerebellum. Nature 2024, 625, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Wizeman, J.W.; Guo, Q.; Wilion, E.M.; Li, J.Y. Specification of Diverse Cell Types during Early Neurogenesis of the Mouse Cerebellum. eLife 2019, 8, e42388. [Google Scholar] [CrossRef] [PubMed]
- Sotelo, C.; Rossi, F. Purkinje Cell Migration and Differentiation. In Handbook of the Cerebellum and Cerebellar Disorders; Springer: Dordrecht, The Netherlands, 2013. [Google Scholar]
- Schreglmann, S.R.; Riederer, F.; Galovic, M.; Ganos, C.; Kägi, G.; Waldvogel, D.; Jaunmuktane, Z.; Schaller, A.; Hidding, U.; Krasemann, E.; et al. Movement Disorders in Genetically Confirmed Mitochondrial Disease and the Putative Role of the Cerebellum. Mov. Disord. 2018, 33, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.E. Alcohol Withdrawal and Cerebellar Mitochondria. Cerebellum 2015, 14, 421–437. [Google Scholar] [CrossRef] [PubMed]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial Reactive Oxygen Species (ROS) and ROS-Induced ROS Release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef] [PubMed]
- Olguín-Albuerne, M.; Morán, J. Redox Signaling Mechanisms in Nervous System Development. Antioxid. Redox Signal. 2018, 28, 1603–1625. [Google Scholar] [CrossRef] [PubMed]
- Perez-Pouchoulen, M.; VanRyzin, J.W.; McCarthy, M.M. Morphological and Phagocytic Profile of Microglia in the Developing Rat Cerebellum. eNeuro 2015, 2, ENEURO.0036-15.2015. [Google Scholar] [CrossRef] [PubMed]
- Stoessel, M.B.; Majewska, A.K. Little Cells of the Little Brain: Microglia in Cerebellar Development and Function. Trends Neurosci. 2021, 44, 564–578. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Saliba, N.B.; Martin, H.; Losurdo, N.A.; Kolahdouzan, K.; Siddiqui, R.; Medeiros, D.; Li, W. Purkinje Cell Dopaminergic Inputs to Astrocytes Regulate Cerebellar-Dependent Behavior. Nat. Commun. 2023, 14, 1613. [Google Scholar] [CrossRef] [PubMed]
- Harsing, L.G.; Szénási, G.; Fehér, B.; Miklya, I. Regulation by Trace Amine-Associated Receptor 1 (TAAR1) of Dopaminergic-GABAergic Interaction in the Striatum: Effects of the Enhancer Drug (−)BPAP. Neurochem. Res. 2025, 50, 94. [Google Scholar] [CrossRef] [PubMed]
- Alfimova, M.V.; Plakunova, V.V.; Lezheiko, T.V.; Golimbet, V.E. [Polymorphic variants in the cluster of genes encoding trace amine receptors and cognitive functioning in patients with schizophrenia spectrum disorders and healthy controls]. Zh. Nevrol. Psikhiatr. Im. S. S. Korsakova 2024, 124, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Belov, D.R.; Efimova, E.V.; Fesenko, Z.S.; Antonova, K.A.; Kolodyazhny, S.F.; Lakstygal, A.M.; Gainetdinov, R.R. Putative Trace-Amine Associated Receptor 5 (TAAR5) Agonist α-NETA Increases Electrocorticogram Gamma-Rhythm in Freely Moving Rats. Cell. Mol. Neurobiol. 2020, 40, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Aleksandrov, A.A.; Dmitrieva, E.S.; Volnova, A.B.; Knyazeva, V.M.; Polyakova, N.V.; Ptukha, M.A.; Gainetdinov, R.R. Effect of Alpha-NETA on Auditory Event Related Potentials in Sensory Gating Study Paradigm in Mice. Neurosci. Lett. 2019, 712, 134470. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for Functional Genomics Data Sets—Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [PubMed]
- Cavaliere, A.F.; Parasiliti, M.; Franco, R.; Gallitelli, V.; Perelli, F.; Spanò, A.; Pallone, B.; Serafini, M.G.; Signore, F.; Eleftheriou, G.; et al. Successful Elimination of Endemic Rubella in the WHO European Region. Is It Proper to Remove the Recommendation for Preconceptional Immunization? Int. J. Environ. Res. Public Health 2024, 21, 957. [Google Scholar] [CrossRef] [PubMed]
- Raina, P.; Guinea, R.; Chatsirisupachai, K.; Lopes, I.; Farooq, Z.; Guinea, C.; Solyom, C.-A.; de Magalhães, J.P. GeneFriends: Gene Co-Expression Databases and Tools for Humans and Model Organisms. Nucleic Acids Res. 2023, 51, D145–D158. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GEO ID | Title | Mouse Strain | Age | n | Available Data |
|---|---|---|---|---|---|
| GSE80222 | Polycomb Ezh2 controls the fate of GABAergic neurons in the embryonic cerebellum | C57BL/6 | E13.5 | 4 * | RPKM |
| GSE87104 | RNA-seq Analysis of Wild Type and Ptpn11-deficient cerebellar Transcriptomes | CD1 | E13.5 | 9 * | Raw counts |
| GSE226532 | Ethanol Induced Cerebellar Transcriptomic Changes in a Postnatal Model of Fetal Alcohol Spectrum Disorders: Focus on Disease Onset | C57BL/6 | P5–6 | 12 * | Raw counts |
| GSE166196 | Divergent and overlapping hippocampal and cerebellar transcriptome responses following developmental ethanol exposure during the secondary neurogenic period. | C57BL/6 | P10 | 7 * | Raw counts |
| GSE114062 | Comparison between a mouse line selected for high voluntary wheel running and control line in two brain regions, cerebellum and striatum | Hsd:ICR | P48 | 4 | FPKM |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vaganova, A.N.; Murtazina, R.Z.; Volnova, A.B.; Tsytsarev, V.; Karaseva, A.B.; Efimova, E.V.; Gainetdinov, R.R. Transcriptomic Analysis of Taar5 Expression and Co-Expression Networks in the Cerebellum During Perinatal Development. Brain Sci. 2025, 15, 791. https://doi.org/10.3390/brainsci15080791
Vaganova AN, Murtazina RZ, Volnova AB, Tsytsarev V, Karaseva AB, Efimova EV, Gainetdinov RR. Transcriptomic Analysis of Taar5 Expression and Co-Expression Networks in the Cerebellum During Perinatal Development. Brain Sciences. 2025; 15(8):791. https://doi.org/10.3390/brainsci15080791
Chicago/Turabian StyleVaganova, Anastasia N., Ramilya Z. Murtazina, Anna B. Volnova, Vassiliy Tsytsarev, Alena B. Karaseva, Evgeniya V. Efimova, and Raul R. Gainetdinov. 2025. "Transcriptomic Analysis of Taar5 Expression and Co-Expression Networks in the Cerebellum During Perinatal Development" Brain Sciences 15, no. 8: 791. https://doi.org/10.3390/brainsci15080791
APA StyleVaganova, A. N., Murtazina, R. Z., Volnova, A. B., Tsytsarev, V., Karaseva, A. B., Efimova, E. V., & Gainetdinov, R. R. (2025). Transcriptomic Analysis of Taar5 Expression and Co-Expression Networks in the Cerebellum During Perinatal Development. Brain Sciences, 15(8), 791. https://doi.org/10.3390/brainsci15080791