3.1. Learning by Passive Exposure to Oriented Stimuli
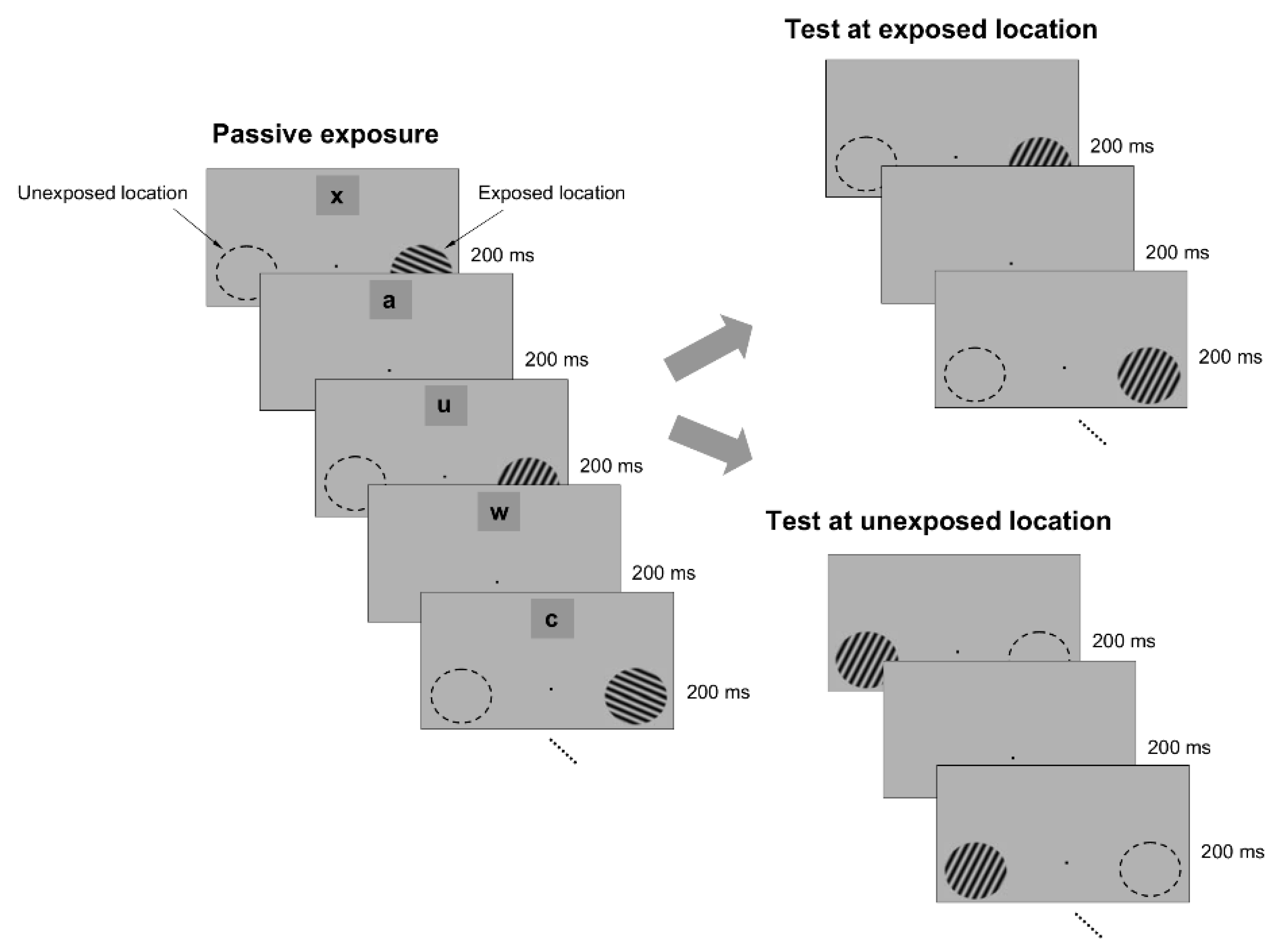
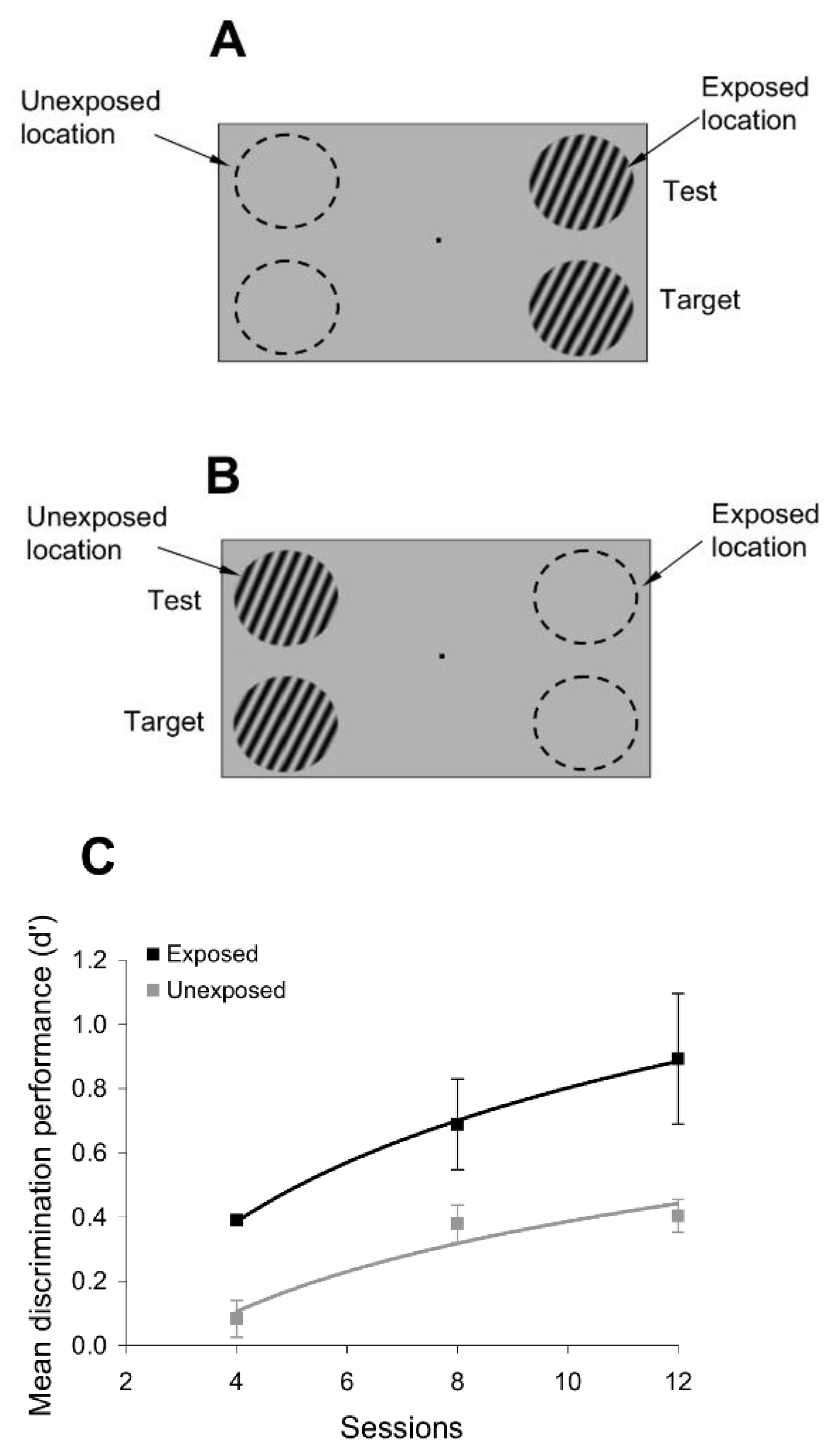
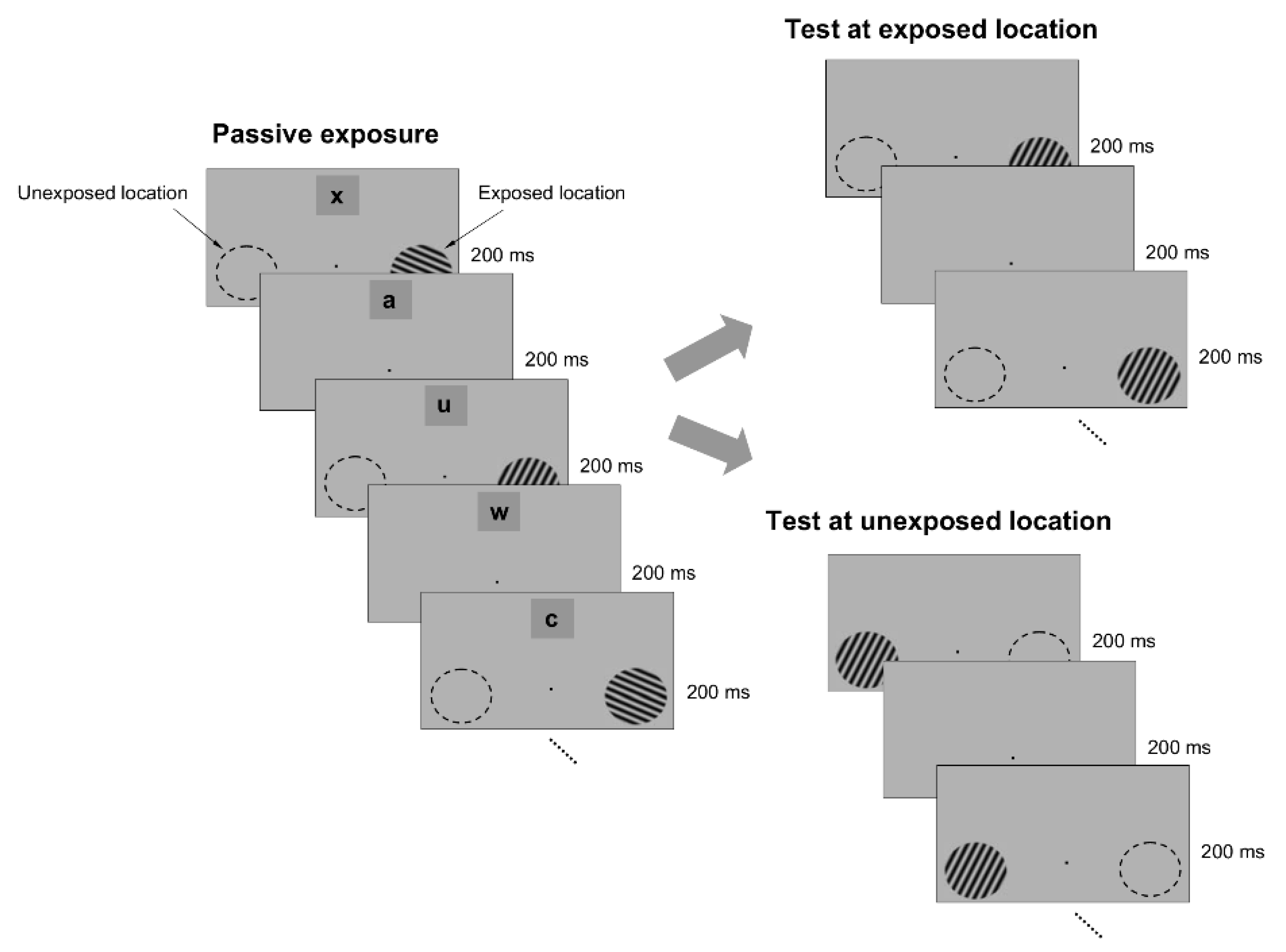
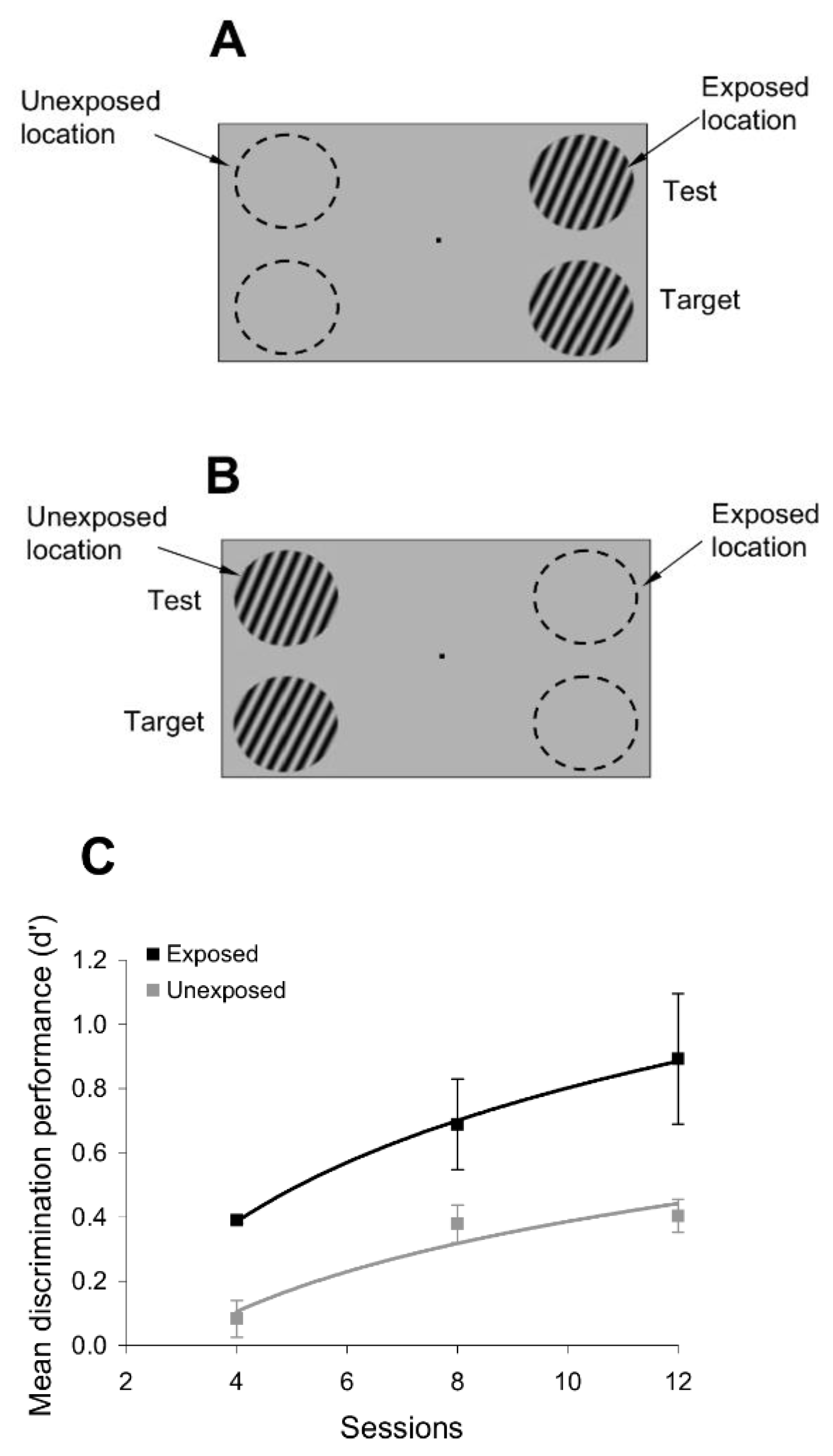
To test whether passive exposure to oriented stimuli induces learning, we asked subjects to fixate on a small dot in the center of a computer screen while oriented sine-wave gratings were repeatedly flashed for 200 ms each, and separated by a 200 ms blank interval (
Figure 1). The oriented stimuli were presented only at an ‘exposed’ location during a typical 3 min exposure session, while another location, symmetric with respect to the fixation point, was not stimulated (
Figure 1). Throughout the exposure stage, we controlled attention by asking subjects to complete a letter identification task (presented 7° above the fixation point), in which a new letter was randomly presented every 200 ms (
Figure 1). Subjects were required to count how many times a specific letter in the sequence was presented while they maintained fixation and were stimulated at the exposed location. We chose the location of the letter task based on pilot tests (
Supplementary Figure S1), conducted prior to the exposure experiments, such as to ensure that the exposed and unexposed locations were unattended when subjects performed the letter task at a performance level of 80% or more.
To examine whether passive exposure influences perceptual performance, we asked subjects to test their orientation discrimination performance around and orthogonal to the exposed orientation axes, both at the exposed and unexposed locations (the discrimination tests were performed after the completion of each exposure session). We measured orientation discrimination performance using a delayed match-to-sample task in which subjects were required to report whether two circular gratings, briefly flashed for 200 ms, and separated by a 200 ms blank, differed in orientation (
Figure 1). One of the gratings (target) had a fixed orientation, whereas the orientation of the other grating (test) was within 4° of the first (orientation was randomly varied in steps of ±1°). This allowed us to measure whether and how the orientation discrimination performance (quantified by the d’) changed at the exposed and unexposed locations after each exposure session.
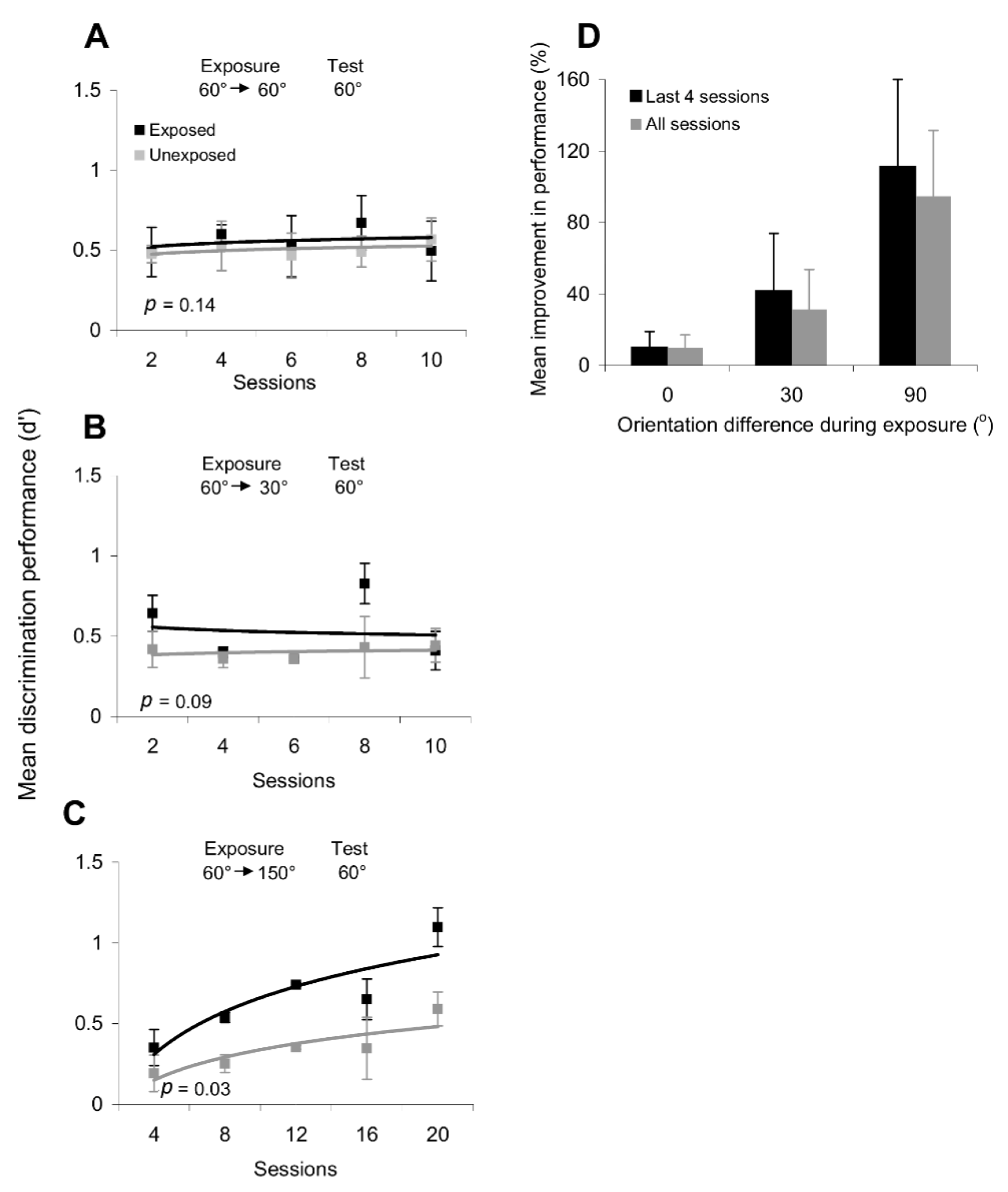
The effects of orientation exposure on the discrimination performance are summarized in
Figure 3. We found that repetitive passive exposure to one orientation did not improve or impair orientation discrimination performance along parallel or orthogonal axes. Indeed, as shown in
Figure 3A, after exposure to 60° orientations, there was no difference between the discrimination performance at the exposed and unexposed locations (
p > 0.1, Student’s
t-test). When the orientation discrimination performance was tested around the orthogonal orientation (
Figure 3B), we found that exposure caused an early increase in discrimination performance at the exposed location (exposure to 60° and test at 150°), but performance in the subsequent sessions did not differ significantly from that at the unexposed location (
p > 0.1, Student’s
t-test).
We further tested the possibility that repetitive exposure to sequences of different orientations could induce systematic changes in discrimination performance. Thus, we repeatedly exposed subjects to sequences of non-orthogonal (60°/30°) and orthogonal orientations (60°/150°). However, whereas repeated exposure to the 60°/30° sequence led to a non-significant improvement in orientation discrimination performance around 60° (
p = 0.09, Student’s
t-test;
Figure 3C), only exposure to an orthogonal orientation sequence led to a significant improvement in the discrimination of orientations along the exposed axes (for discrimination performance around 60°:
p = 0.03, Student’s
t-test;
Figure 3D—similar results were obtained when we tested orientation discrimination around 150°).
The session-by-session improvement in orientation discrimination performance after passive exposure to a series of orthogonal gratings was observed only at the exposed location, despite the fact that subjects had similar training in discriminating stimuli around the experienced orientations at both the exposed and unexposed locations. Indeed, after only four exposure sessions (
Figure 3D), subjects were able to correctly perceive increasingly smaller differences in orientation at the exposed location, which is equivalent to an increase in d’, whereas only a weak improvement in discrimination performance was observed at the unexposed location. These results are consistent with recent perceptual and physiological findings on orientation adaptation indicating that short-term adaptation to a grating of fixed orientation sharpens the orientation tuning curves of orthogonal V1 neurons and improves the perceptual discrimination performance around the orthogonal orientations [
23,
24,
29], while broadening the tuning curves of the neurons tuned to all other orientations and impairing the discrimination of intermediate orientations [
23,
24,
29]. This indicates that neuronal circuits in early and mid-level visual cortical areas are selectively altered by exposure to particular orientation sequences to modulate orientation discrimination performance in a way that induces learning.
The learning effects induced by repetitive exposure to orthogonal orientations are persistent. We found (data not shown) that, although the improvement in orientation discrimination performance gradually diminished when exposure was abolished, the discrimination performance remained significantly stronger at the exposed location even at 120 days after exposure was ceased (
p < 0.05, Student’s
t-test). These results are inconsistent with attentional learning theories [
11,
24,
30], which postulate that only attended stimuli induce long-lasting changes in sensory systems, and suggest that learning can arise through purely bottom-up mechanisms. Thus, repeated training on a specific visual task is not a necessary prerequisite of perceptual learning.
We further examined the degree of specificity of exposure learning by testing whether it depends on retinal position and orientation. Such dependency is a distinct feature of the perceptual learning of elementary features [
9,
10,
13,
14,
31,
32] and argues for the involvement of early and mid-level visual cortical areas in mediating perceptual learning as these areas are known to represent stimulus features with the finest resolution.
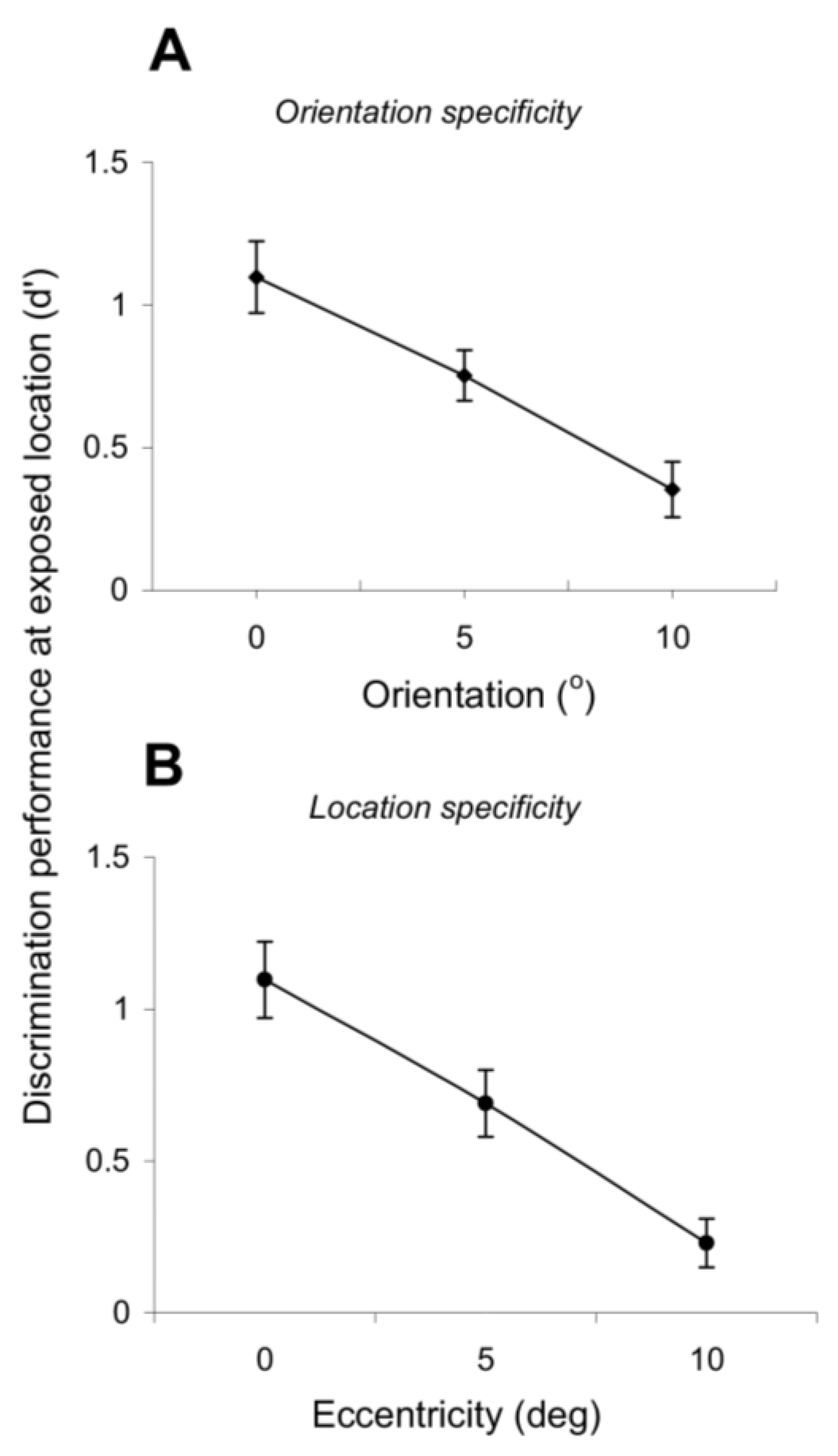
Figure 4A,B reveals a high degree of orientation and position specificity of exposure learning. Indeed, the orientation discrimination performance at the exposed location (d’) gradually decreases as a function of increasing difference between the test and exposed orientations (
Figure 4A;
p < 0.01, Student’s
t-test). Furthermore, the exposure-induced learning effects are spatially localized—
Figure 4B shows that the orientation discrimination performance decreases as the eccentricity at which the orientation discrimination is performed increases (
p < 0.01, Student’s
t-test). Thus, exposure learning retains the position and orientation specificity of task-related practice learning.
In principle, it could be argued that subjects may use orientation-insensitive mechanisms to distinguish between the target and test stimuli. Indeed, the timing of the stimuli in the test phase (200 ms) would be able to produce apparent motion. Hence, stimulus discriminations might be more easily performed by assessing the motion between the target and test rather than by comparing their orientation. Therefore, it is possible that the stimuli during the exposure stage may have altered the strength of the motion stimuli used in assessing the difference between the target and the test during the testing phase. To control for this possibility, we conducted exposure sessions similar to those described in
Figure 1, but, after each exposure session, presented the test and target stimuli simultaneously during the testing phase (
Figure 5A,B).
In these parallel orientation discrimination experiments, the test stimulus (of random orientation) was always presented at the exposed location, whereas the target stimulus was presented at a new, previously unexposed, location. If subjects rely exclusively on motion signals to distinguish between the target and the test, and if exposure alters the strength of those motion stimuli used in orientation discrimination, presenting the target and test stimuli in parallel should abolish the exposure-induced improvement in orientation discrimination performance. However, contrary to this prediction,
Figure 5C shows that exposure to orthogonal orientations improves orientation discrimination performance in a significant manner (
p < 0.05, Student’s
t-test), even when the target and test stimuli are presented together. This demonstrates that exposure to orientation sequences induces effects that rely primarily on orientation, not motion, mechanisms.
It is also possible that the large difference between the orientation discrimination performance at the exposed and unexposed locations simply reflects a lack of stimulation at the unexposed location rather than an effect of exposure to orthogonal orientation sequences at the exposed location. To control for this possibility, we repeatedly stimulated a new location using a mask composed of a superposition of eight orientations equally spanning 0–180° that was flashed for 200 ms and was followed by a 200 ms blank (the stimulation duration was 3 min). However, although the mask was repeatedly presented for several sessions, we found that the orientation discrimination performance measured after exposure to these control stimuli was not significantly different from that at the unexposed location (p > 0.05, Student’s t-test).
3.3. Time Course of Learning Effects
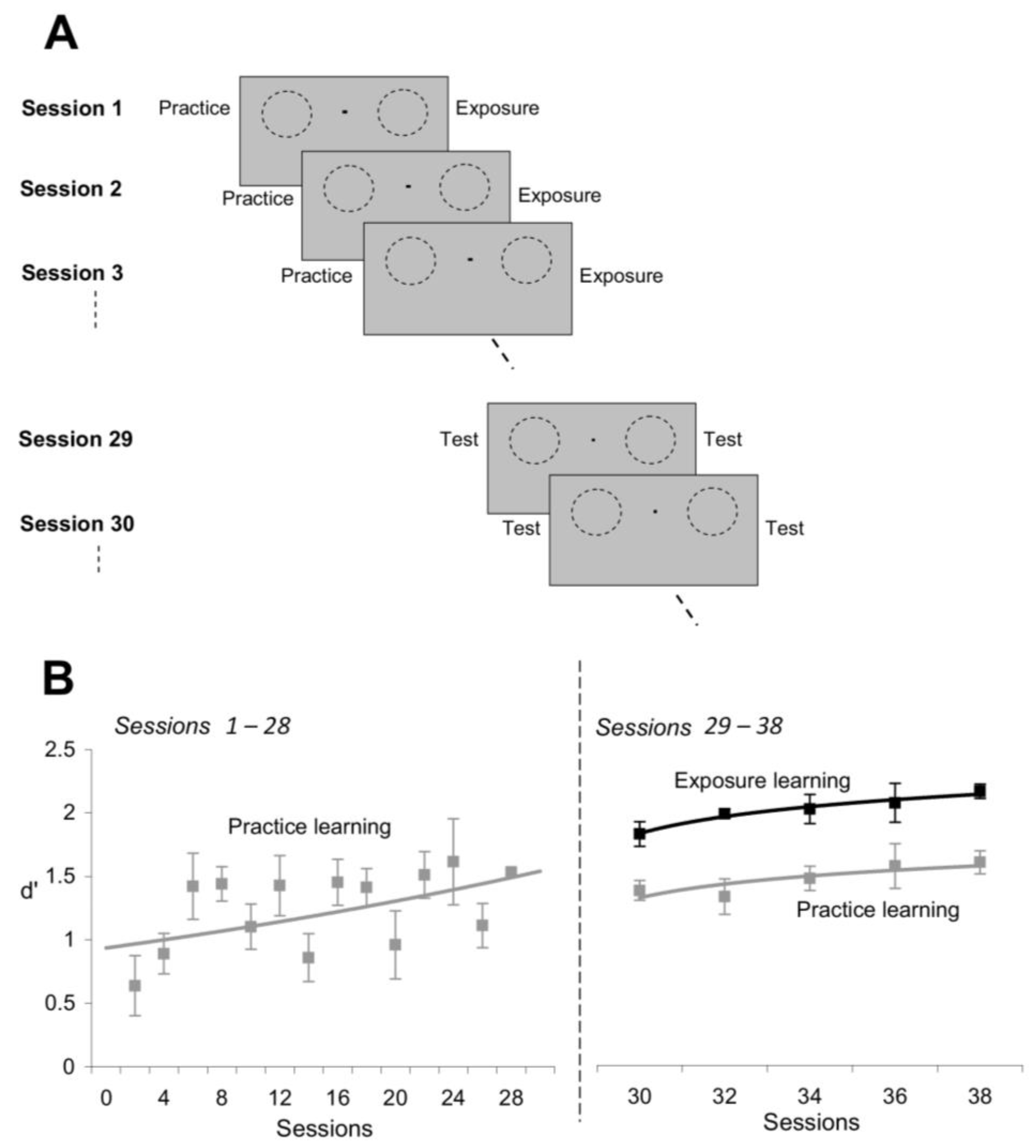
We asked subjects to practice orientation discriminations at one location and, in a separate session, perform an exposure session in conjunction with a letter identification task at another location (
Figure 6A). The two locations were symmetric with respect to the fixation point, and were stimulated in each session with the same number of stimuli, either nearby orientations (at the practice location) or orthogonal orientations (at the exposed location). The stimulus sequence for both the orientation discrimination practice sessions and the orientation exposure sessions was identical to the stimulus sequence from the experiments described in
Figure 1. These two sets of stimuli were chosen in order to maximize performance for practice learning and exposure learning. Indeed, the most effective stimuli for practice learning are the stimuli of nearby orientation, whereas the most effective stimuli for exposure learning are the stimuli of orthogonal orientation (e.g.,
Figure 3D).
In agreement with previous perceptual learning studies [
10,
14,
20], we confirmed that subjects gradually improved their orientation discrimination performance at the practice location during the time course of training (
Figure 6B, left). Indeed, the mean orientation discrimination performance, d’, at the end of training (Sessions 25–28) was significantly higher than the mean d’ at the beginning of training (Sessions 1–4;
p < 0.03, Student’s
t-test) in all the subjects. However, contrary to common belief that practice learning is stronger than exposure learning, we found that repetitive exposure to orthogonal orientations led to superior learning (
Figure 6B, right). That is, despite the fact that subjects did not perform any visual task at the exposed location for 28 sessions, they were nonetheless able to discriminate finer orientation differences at the exposed location compared to the practice location (Sessions 29–38). This demonstrates that passive exposure to orthogonal orientations induces stronger learning than task repetition (
p < 0.008, Student’s
t-test), and that exposure learning develops at a faster time course than practice learning.
3.4. Stimulus Generalization
Generalization is a key property of learning and is defined as a transfer of the improvement in perceptual performance achieved through training to new stimuli. Classical perceptual learning theories postulate the specificity of learning for the simple stimuli present during training, such as stimulus position and orientation, with little or no generalization to more complex stimuli. Indeed, there has been little evidence that practice learning leads to improvement in discrimination performance beyond the local stimulus configuration during task practice [
9,
13,
15,
18,
19,
20,
32]. Given the importance of the generalization issue, we examined whether the improvement in orientation discrimination performance at the exposed location leads to improvement in the discrimination of complex shapes composed of orientation signals around the experienced axes, and then compared the degree of transfer of exposure and practice learning. Based on the rapid rate of acquisition of exposure learning (
Figure 5), we expected that the discrimination of complex stimuli, such as shapes and natural scenes, would be improved more after repetitive exposure to orthogonal orientations, than after repetitive practice of orientation discriminations.
We tested the degree of stimulus generalization of exposure and practice learning by asking subjects to perform shape and image discrimination tasks after they had been exposed to orthogonal orientations and had practiced orientation discriminations for the same number of sessions (
Figure 7). Thus, we conducted daily orientation exposure sessions similar to those described in
Figure 1, in which subjects were exposed to two pairs of orthogonal orientations: 60°/150° and 30°/120° (in separate sessions; 12 sessions for each pair of orientations—
Figure 7A). In pilot experiments (
Suppl. Figure S2) we confirmed that exposure to two different pairs of orthogonal stimuli improved orientation discrimination along the four experienced axes (
p < 0.03, Student’s
t-test). The improvement in performance was restricted to the exposed location and did not impair or enhance performance around orientations other than the exposed ones. At another spatial location, we asked subjects to practice orientation discriminations around the four orientations used in the exposure experiment (12 sessions per orientation,
Figure 7B). After the orientation exposure and discrimination practice sessions were completed, subjects were required to discriminate shapes and natural scenes.
These discrimination tests were performed at both pairs of exposed/unexposed and trained/untrained locations (
Figure 7A,B) by successively flashing shapes or natural images that were slightly tilted relative to each other (a target stimulus was followed by a test stimulus that was either identical to the target or was rotated by ±2°, ±3°, ±4°, and ±5° with respect to the target). At the end of each trial, subjects were asked to judge whether the test was similar or different from the target (the number of match and non-match stimuli was identical). We reasoned that since complex stimuli have a rich representation in the orientation domain [
23,
33,
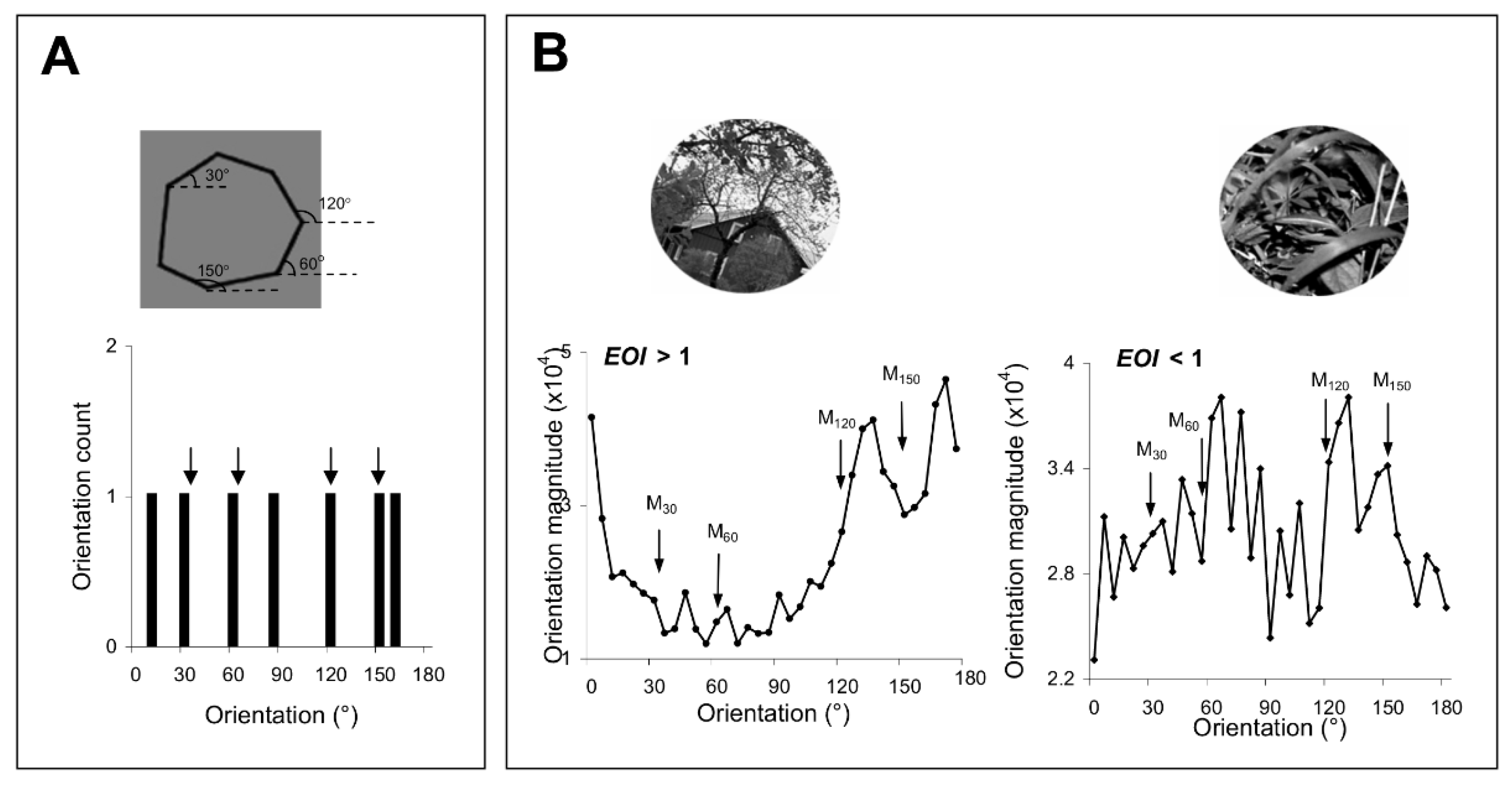
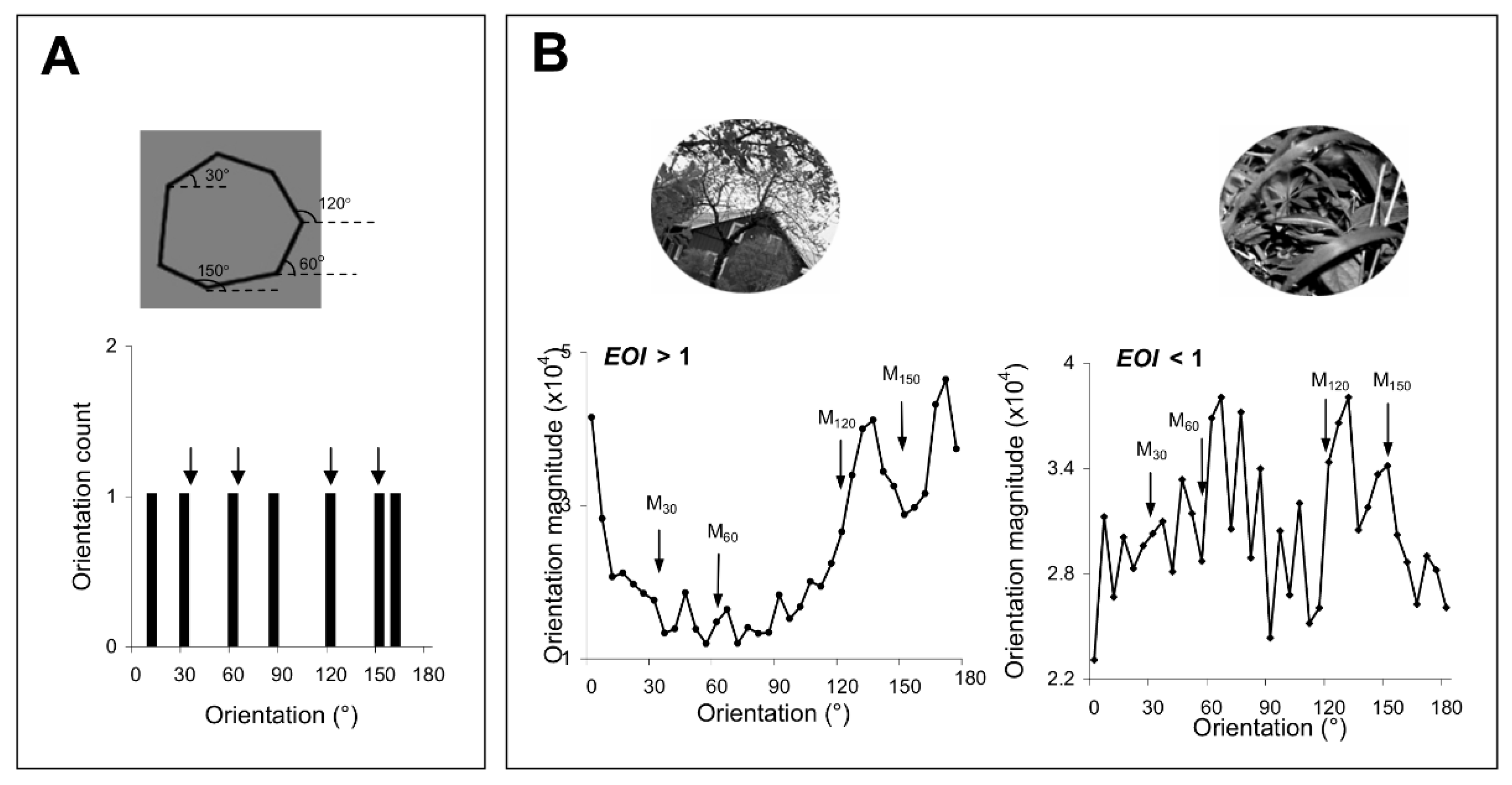
34], improving orientation discrimination through repetitive exposure or practice could lead to an improvement in the discrimination of those complex stimuli with strong orientation signals around the exposed or trained axes. We controlled the orientation content of the complex stimuli by generating polygons with four of the sides oriented along the exposed axes (
Figure 2A), and natural scenes of broad orientation spectrum, including orientation signals of various strengths along the experienced axes (
Figure 2B). We evaluated the orientation content of each image by calculating the orientation magnitude histogram, and, for each histogram, computed an exposure orientation index (EOI, see
Section 2) that quantifies the relative strength of orientation signals along the experienced axes.
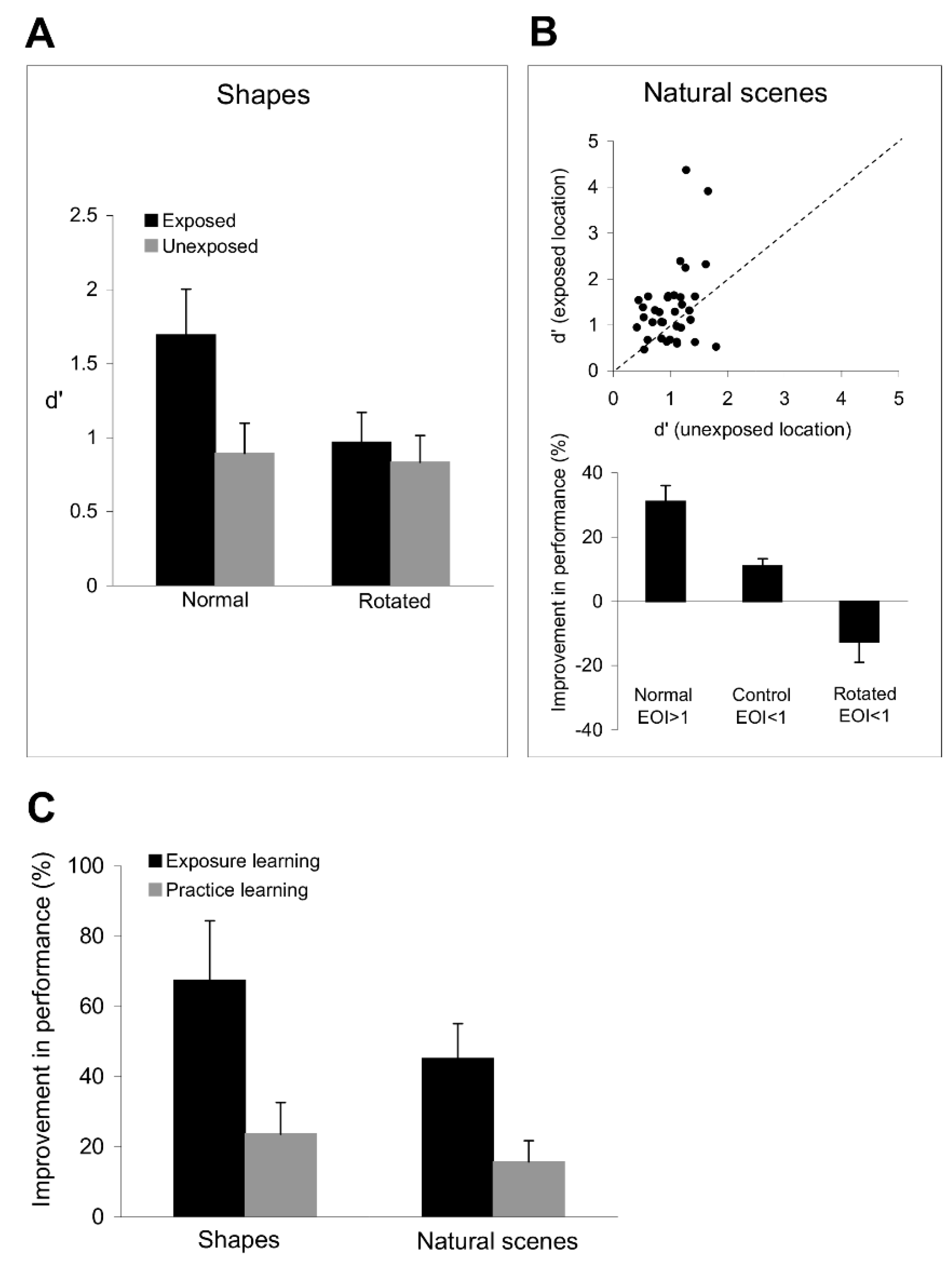
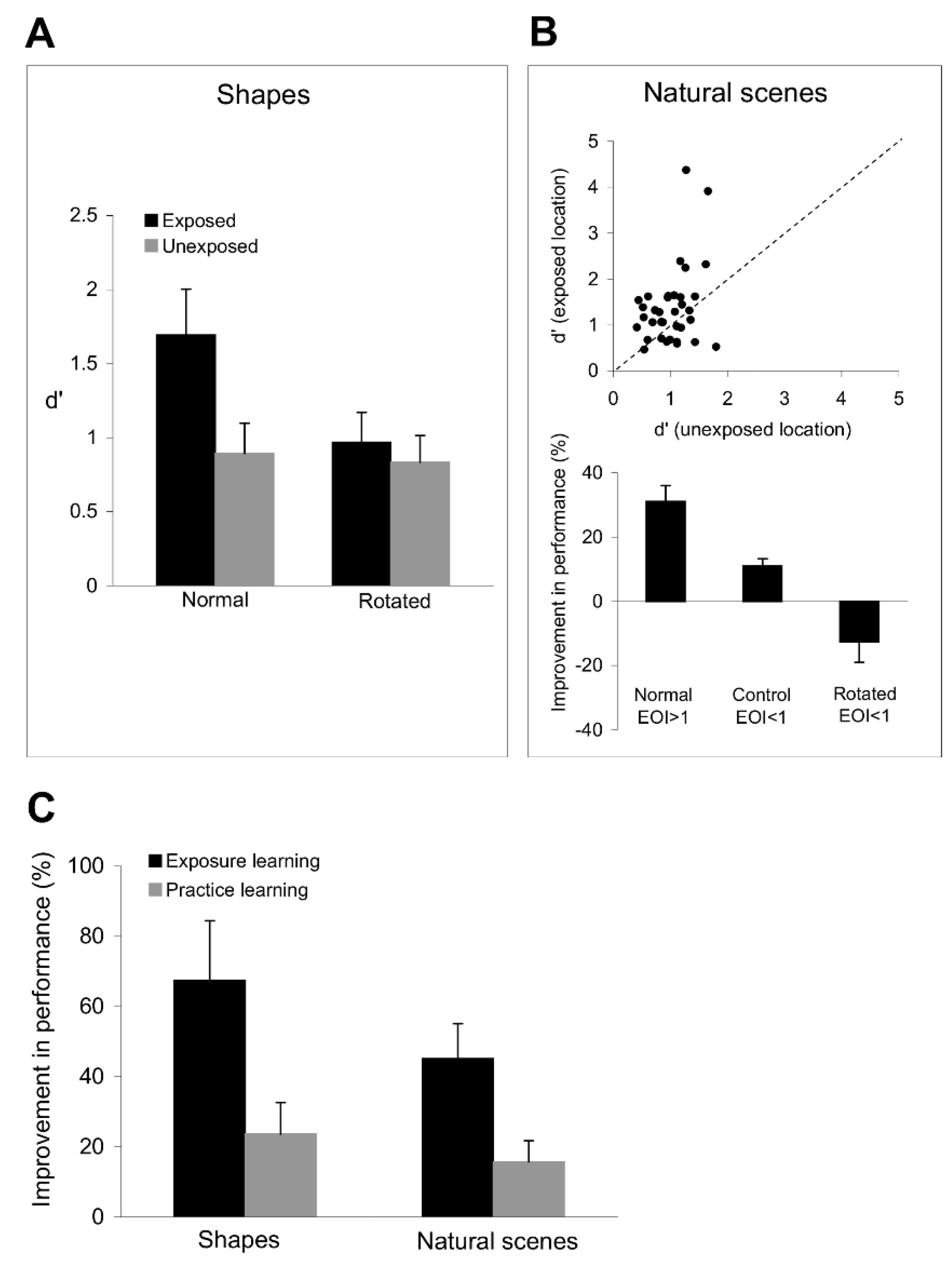
The results in
Figure 8A,B demonstrate that exposure to orthogonal orientations leads to a robust improvement in the discrimination of complex visual stimuli despite the fact that subjects had no prior experience with these stimuli. Indeed,
Figure 8A shows a significant improvement in shape discrimination performance at the exposed location (
p < 0.03, Student’s
t-test), likely reflecting an increase in neuronal orientation discrimination performance around the experienced axes after repeated exposure. To test whether there is a direct relationship between the improvement in orientation discrimination around the experienced axes and the shape discrimination performance, we conducted control experiments in which we rotated the shapes by 20°, such that all the sides of the polygons were now presented at orientations that differed by more than 10° from the experienced ones. Since orientation exposure improves orientation discrimination performance only around the experienced orientations (
Figure 3), we found that rotating the target and test shapes abolished the improvement in shape discrimination performance (
Figure 8A), which was similar at the exposed and unexposed locations (
p > 0.1, Student’s
t-test).
Importantly, exposure to orthogonal orientation sequences leads to a significant improvement not only in shape discrimination performance, but also in the ability of the visual system to discriminate natural images.
Figure 8B (top) shows that the image discrimination performance at the exposed location is significantly better than that at the unexposed location (
p < 0.01, Student’s
t-test). We controlled the orientation content of each image by dividing the images in our set into two groups depending on whether their exposure orientation index, EOI, was greater or smaller than 1.
Figure 8B (bottom) shows that for those images with strong orientation components along the experienced axes (EOI > 1), subjects were more likely to indicate that the test and target images were different at the exposed location than when they were presented at the unexposed location (
p < 0.03, Student’s
t-test). Importantly, exposure to orthogonal orientations did not improve discrimination performance (
p > 0.5, Student’s
t-test) for images with weak orientation signals (EOI < 1) along the experienced axes (
Figure 8B, bottom), possibly due to lack of exposure-induced plasticity in neurons tuned to those orientations. We further examined the link between the exposure-induced improvement in orientation and image discrimination performance by rotating the images with strong orientation signals along exposed axes by 20° (this procedure dropped EOI to values < 1 in each image) followed by image discrimination. Despite the fact that the images before rotation were associated with a significant improvement in discrimination performance at the exposed location, weakening the correlation between the orientation content of the image and the experienced orientations (by rotation) abolished the original improvement in discrimination performance (
p > 0.1, Student’s
t-test).
These data indicate that the improvement in the discrimination of simple stimuli after exposure generalizes to complex stimuli that include combinations of elementary stimuli. This raises the issue of whether practice learning exhibits a similar degree of stimulus generalization. We directly compared the degree of specificity of bottom-up and top-down plasticity by examining the subjects’ capacity to discriminate complex shapes after they completed 12 successive sessions of exposure and practice learning.
Figure 8C illustrates the mean improvement in shape and image discrimination performance at the exposed and trained locations (relative to the unexposed and untrained locations). Confirming our prediction, subjects exhibited a larger improvement in shape (
p < 0.01, Student’s
t-test) and image (
p < 0.03, Student’s
t-test) discrimination performance after exposure learning than after practice learning (comparing the improvement in discrimination performance in four sessions at the exposed and trained locations in each subject).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}