
Repetitive Anodal TDCS to the Frontal Cortex Increases the P300 during Working Memory Processing
 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Design and Working Memory Assessment
2.3. Transcranial Direct Current Stimulation
2.4. EEG Recording and Preprocessing
2.5. Data Analysis
2.5.1. Behavioral Data
2.5.2. Event-Related Potentials
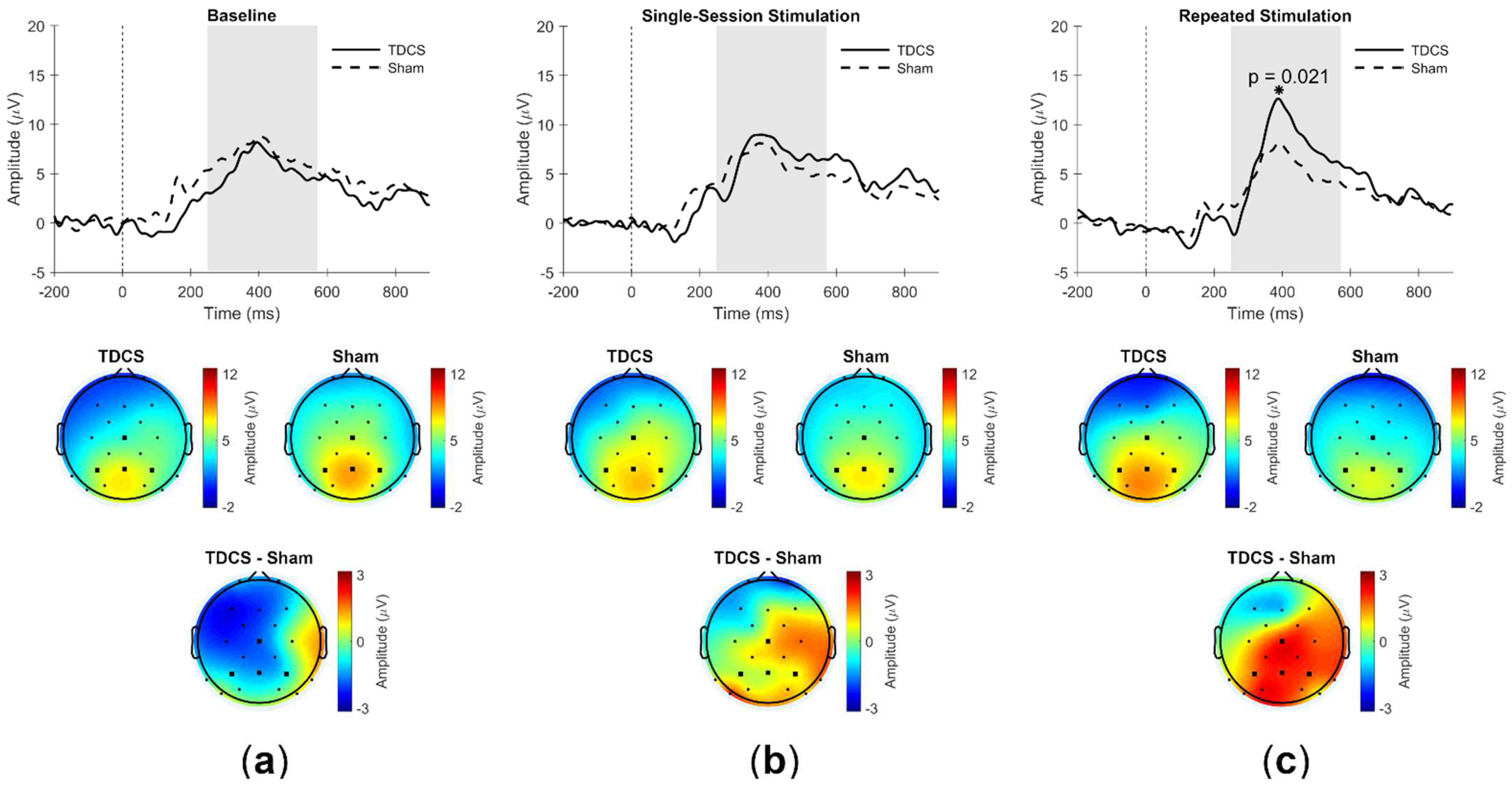
3. Results
3.1. Reaction Times
3.2. Accuracy
3.3. ERPs
3.4. Correlation between Working Memory Performance and P300 Amplitude
3.5. Side-Effects
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nitsche, M.A.; Cohen, L.G.; Wassermann, E.M.; Priori, A.; Lang, N.; Antal, A.; Paulus, W.; Hummel, F.; Boggio, P.S.; Fregni, F.; et al. Transcranial direct current stimulation: State of the art 2008. Brain Stimul. 2008, 1, 206–223. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Paulus, W. Excitability changes induced in the human motor cortex by weak transcranial direct current stimulation. J. Physiol. 2000, 527, 633–639. [Google Scholar] [CrossRef]
- Liebetanz, D.; Nitsche, M.A.; Tergau, F.; Paulus, W. Pharmacological approach to the mechanisms of transcranial DC-stimulation-induced after-effects of human motor cortex excitability. Brain 2002, 125, 2238–2247. [Google Scholar] [CrossRef]
- Filmer, H.L.; Dux, P.E.; Mattingley, J.B. Applications of transcranial direct current stimulation for understanding brain function. Trends Neurosci. 2014, 37, 742–753. [Google Scholar] [CrossRef]
- Bikson, M.; Brunoni, A.R.; Charvet, L.E.; Clark, V.P.; Cohen, L.G.; Deng, Z.-D.; Dmochowski, J.; Edwards, D.J.; Frohlich, F.; Kappenman, E.S.; et al. Rigor and reproducibility in research with transcranial electrical stimulation: An NIMH-sponsored workshop. Brain Stimul. 2017, 11, 465–480. [Google Scholar] [CrossRef]
- Liu, A.; Vöröslakos, M.; Kronberg, G.; Henin, S.; Krause, M.R.; Huang, Y.; Opitz, A.; Mehta, A.; Pack, C.C.; Krekelberg, B.; et al. Immediate neurophysiological effects of transcranial electrical stimulation. Nat. Commun. 2018, 9, 5092. [Google Scholar] [CrossRef]
- Jonides, J.; Reuter-Lorenz, P.A.; Smith, E.E.; Awh, E.; Barnes, L.L.; Drain, M.; Glass, J.; Lauber, E.J.; Patalano, A.L.; Schumacher, E.H. Verbal and Spatial Working Memory in Humans. Psychol. Learn. Motiv. 1996, 35, 43–88. [Google Scholar] [CrossRef]
- Baddeley, A. Working Memory: Theories, Models, and Controversies. Annu. Rev. Psychol. 2012, 63, 1–29. [Google Scholar] [CrossRef]
- Baddeley, A.D.; Hitch, G. Working Memory. In The Psychology of Learning and Motivation: Advances in Research and Theory; Bower, G.H., Ed.; Academic Press: Cambridge, MA, USA, 1974; pp. 47–89. [Google Scholar]
- Ferrucci, R.; Marceglia, S.; Vergari, M.; Cogiamanian, F.; Mrakic-Sposta, S.; Mameli, F.; Zago, S.; Barbieri, S.; Priori, A. Cerebellar Transcranial Direct Current Stimulation Impairs the Practice-dependent Proficiency Increase in Working Memory. J. Cogn. Neurosci. 2008, 20, 1687–1697. [Google Scholar] [CrossRef]
- Fregni, F.; Boggio, P.S.; Nitsche, M.; Bermpohl, F.; Antal, A.; Feredoes, E.; Marcolin, M.A.; Rigonatti, S.P.; Silva, M.T.; Paulus, W.; et al. Anodal transcranial direct current stimulation of prefrontal cortex enhances working memory. Exp. Brain Res. 2005, 166, 23–30. [Google Scholar] [CrossRef]
- Mulquiney, P.G.; Hoy, K.E.; Daskalakis, Z.J.; Fitzgerald, P.B. Improving working memory: Exploring the effect of transcranial random noise stimulation and transcranial direct current stimulation on the dorsolateral prefrontal cortex. Clin. Neurophysiol. 2011, 122, 2384–2389. [Google Scholar] [CrossRef] [PubMed]
- Zaehle, T.; Sandmann, P.; Thorne, J.D.; Jäncke, L.; Herrmann, C.S. Transcranial direct current stimulation of the prefrontal cortex modulates working memory performance: Combined behavioural and electrophysiological evidence. BMC Neurosci. 2011, 12, 2. [Google Scholar] [CrossRef] [PubMed]
- Ohn, S.H.; Park, C.-I.; Yoo, W.-K.; Ko, M.-H.; Choi, K.P.; Kim, G.-M.; Lee, Y.T.; Kim, Y.-H. Time-dependent effect of transcranial direct current stimulation on the enhancement of working memory. NeuroReport 2008, 19, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.A.; Fricke, K.; Henschke, U.; Schlitterlau, A.; Liebetanz, D.; Lang, N.; Henning, S.; Tergau, F.; Paulus, W. Pharmacological Modulation of Cortical Excitability Shifts Induced by Transcranial Direct Current Stimulation in Humans. J. Physiol. 2003, 553, 293–301. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Paulus, W. Sustained excitability elevations induced by transcranial DC motor cortex stimulation in humans. Neurology 2001, 57, 1899–1901. [Google Scholar] [CrossRef]
- Fregni, F.; Boggio, P.S.; Nitsche, M.A.; Rigonatti, S.P.; Pascual-Leone, A. Cognitive effects of repeated sessions of transcranial direct current stimulation in patients with depression. Depress. Anxiety 2006, 23, 482–484. [Google Scholar] [CrossRef]
- Martin, D.; Liu, R.; Alonzo, A.; Green, M.; Player, M.J.; Sachdev, P.; Loo, C.K. Can transcranial direct current stimulation enhance outcomes from cognitive training? A randomized controlled trial in healthy participants. Int. J. Neuropsychopharmacol. 2013, 16, 1927–1936. [Google Scholar] [CrossRef]
- Richmond, L.L.; Wolk, D.; Chein, J.; Olson, I.R. Transcranial Direct Current Stimulation Enhances Verbal Working Memory Training Performance over Time and Near Transfer Outcomes. J. Cogn. Neurosci. 2014, 26, 2443–2454. [Google Scholar] [CrossRef]
- Jones, K.T.; Stephens, J.A.; Alam, M.; Bikson, M.; Berryhill, M.E. Longitudinal Neurostimulation in Older Adults Improves Working Memory. PLoS ONE 2015, 10, e0121904. [Google Scholar] [CrossRef]
- Ruf, S.P.; Fallgatter, A.J.; Plewnia, C. Augmentation of working memory training by transcranial direct current stimulation (tDCS). Sci. Rep. 2017, 7, 876. [Google Scholar] [CrossRef]
- Jones, K.T.; Peterson, D.J.; Blacker, K.J.; Berryhill, M.E. Frontoparietal neurostimulation modulates working memory training benefits and oscillatory synchronization. Brain Res. 2017, 1667, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Mashal, N.; Metzuyanim-Gorelick, S. New information on the effects of transcranial direct current stimulation on n-back task performance. Exp. Brain Res. 2019, 237, 1315–1324. [Google Scholar] [CrossRef] [PubMed]
- Talsma, L.J.; Kroese, H.A.; Slagter, H.A. Boosting Cognition: Effects of Multiple-Session Transcranial Direct Current Stimulation on Working Memory. J. Cogn. Neurosci. 2017, 29, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Lally, N.; Nord, C.L.; Walsh, V.; Roiser, J.P. Does excitatory fronto-extracerebral tDCS lead to improved working memory perfor-mance? F1000Research 2013, 2, 219. [Google Scholar] [CrossRef]
- Owen, A.M.; McMillan, K.M.; Laird, A.R.; Bullmore, E. N-back working memory paradigm: A meta-analysis of normative functional neuroimaging studies. Hum. Brain Mapp. 2005, 25, 46–59. [Google Scholar] [CrossRef]
- Smith, E.E.; Jonides, J. Working Memory: A View from Neuroimaging. Cogn. Psychol. 1997, 33, 5–42. [Google Scholar] [CrossRef]
- Smith, E.E.; Jonides, J.; Koeppe, R.A. Dissociating Verbal and Spatial Working Memory Using PET. Cereb. Cortex 1996, 6, 11–20. [Google Scholar] [CrossRef]
- Mottaghy, F.M.; Döring, T.; Müller-Gärtner, H.-W.; Töpper, R.; Krause, B.J. Bilateral parieto-frontal network for verbal working memory: An interference approach using repetitive transcranial magnetic stimulation (rTMS). Eur. J. Neurosci. 2002, 16, 1627–1632. [Google Scholar] [CrossRef]
- Paulesu, E.; Frith, C.D.; Frackowiak, R.S.J. The neural correlates of the verbal component of working memory. Nature 1993, 362, 342–345. [Google Scholar] [CrossRef]
- Jones, K.T.; Berryhill, M.E. Parietal Contributions to Visual Working Memory Depend on Task Difficulty. Front. Psychiatry 2012, 3, 81. [Google Scholar] [CrossRef]
- Berryhill, M.E.; Wencil, E.B.; Coslett, H.B.; Olson, I.R. A selective working memory impairment after transcranial direct current stimulation to the right parietal lobe. Neurosci. Lett. 2010, 479, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Mottaghy, F. Interfering with working memory in humans. Neuroscience 2006, 139, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Lijffijt, M.; Lane, S.D.; Meier, S.L.; Boutros, N.N.; Burroughs, S.; Steinberg, J.L.; Moeller, F.G.; Swann, A.C. P50, N100, and P200 sensory gating: Relationships with behavioral inhibition, attention, and working memory. Psychophysiology 2009, 46, 1059–1068. [Google Scholar] [CrossRef]
- Lubitz, A.F.; Niedeggen, M.; Feser, M. Aging and working memory performance: Electrophysiological correlates of high and low performing elderly. Neuropsychologia 2017, 106, 42–51. [Google Scholar] [CrossRef]
- Picton, T.W. The P300 Wave of the Human Event-Related Potential. J. Clin. Neurophysiol. 1992, 9, 456–479. [Google Scholar] [CrossRef]
- Scharinger, C.; Soutschek, A.; Schubert, T.; Gerjets, P. Comparison of the Working Memory Load in N-Back and Working Memory Span Tasks by Means of EEG Frequency Band Power and P300 Amplitude. Front. Hum. Neurosci. 2017, 11, 6. [Google Scholar] [CrossRef] [PubMed]
- Watter, S.; Geffen, G.M.; Geffen, L.B. The n-back as a dual-task: P300 morphology under divided attention. Psychophysiology 2001, 38, 998–1003. [Google Scholar] [CrossRef] [PubMed]
- Lapenta, O.M.; Di Sierve, K.; de Macedo, E.C.; Fregni, F.; Boggio, P.S. Transcranial direct current stimulation modulates ERP-indexed inhibitory control and reduces food consumption. Appetite 2014, 83, 42–48. [Google Scholar] [CrossRef]
- Pfabigan, D.M.; Seidel, E.-M.; Sladky, R.; Hahn, A.; Paul, K.; Grahl, A.; Küblböck, M.; Kraus, C.; Hummer, A.; Kranz, G.S.; et al. P300 amplitude variation is related to ventral striatum BOLD response during gain and loss anticipation: An EEG and fMRI experiment. NeuroImage 2014, 96, 12–21. [Google Scholar] [CrossRef]
- Halgren, E.; Marinkovic, K.; Chauvel, P. Generators of the late cognitive potentials in auditory and visual oddball tasks. Electroencephalogr. Clin. Neurophysiol. 1998, 106, 156–164. [Google Scholar] [CrossRef]
- McCarthy, G.; Wood, C.C. Intracranial recordings of endogenous ERPs in humans. Electroencephalogr. Clin. Neurophysiol. Suppl. 1987, 39, 331–337. [Google Scholar] [CrossRef]
- Polich, J. Updating P300: An integrative theory of P3a and P3b. Clin. Neurophysiol. 2007, 118, 2128–2148. [Google Scholar] [CrossRef] [PubMed]
- Donchin, E.; Coles, M.G.H. Is the P300 component a manifestation of context updating? Behav. Brain Sci. 1988, 11, 357–374. [Google Scholar] [CrossRef]
- Näätänen, R. The role of attention in auditory information processing as revealed by event-related potentials and other brain measures of cognitive function. Behav. Brain Sci. 1990, 13, 201–233. [Google Scholar] [CrossRef]
- Pergher, V.; Wittevrongel, B.; Tournoy, J.; Schoenmakers, B.; Van Hulle, M.M. N-back training and transfer effects revealed by behavioral responses and EEG. Brain Behav. 2018, 8, e01136. [Google Scholar] [CrossRef]
- Pinal, D.; Zurrón, M.; Díaz, F. Effects of load and maintenance duration on the time course of information encoding and retrieval in working memory: From perceptual analysis to post-categorization processes. Front. Hum. Neurosci. 2014, 8, 165. [Google Scholar] [CrossRef]
- Chen, Y.-N.; Mitra, S.; Schlaghecken, F. Sub-processes of working memory in the N-back task: An investigation using ERPs. Clin. Neurophysiol. 2008, 119, 1546–1559. [Google Scholar] [CrossRef]
- Gevins, A.; Smith, M.E.; Le, J.; Leong, H.; Bennett, J.; Martin, N.; McEvoy, L.; Du, R.; Whitfield, S. High resolution evoked potential imaging of the cortical dynamics of human working memory. Electroencephalogr. Clin. Neurophysiol. 1996, 98, 327–348. [Google Scholar] [CrossRef]
- Scharinger, C.; Soutschek, A.; Schubert, T.; Gerjets, P. When flanker meets the n-back: What EEG and pupil dilation data reveal about the interplay between the two central-executive working memory functions inhibition and updating. Psychophysiology 2015, 52, 1293–1304. [Google Scholar] [CrossRef]
- Zhao, X.; Zhou, R.; Fu, L. Working Memory Updating Function Training Influenced Brain Activity. PLoS ONE 2013, 8, e71063. [Google Scholar] [CrossRef]
- Barré, J.; Michelet, D.; Truchot, J.; Jolivet, E.; Recanzone, T.; Stiti, S.; Tesnière, A.; Pourcher, G. Virtual Reality Single-Port Sleeve Gastrectomy Training Decreases Physical and Mental Workload in Novice Surgeons: An Exploratory Study. Obes. Surg. 2019, 29, 1309–1316. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.S.L.; Lu, J.; Tan, W.B.; Lomanto, D. Training improves laparoscopic tasks performance and decreases operator workload. Surg. Endosc. 2015, 30, 1742–1746. [Google Scholar] [CrossRef] [PubMed]
- Keeser, D.; Padberg, F.; Reisinger, E.; Pogarell, O.; Kirsch, V.; Palm, U.; Karch, S.; Möller, H.-J.; Nitsche, M.; Mulert, C. Prefrontal direct current stimulation modulates resting EEG and event-related potentials in healthy subjects: A standardized low resolution tomography (sLORETA) study. NeuroImage 2011, 55, 644–657. [Google Scholar] [CrossRef] [PubMed]
- Nikolin, S.; Martin, D.; Loo, C.K.; Boonstra, T.W. Effects of TDCS dosage on working memory in healthy participants. Brain Stimul. 2018, 11, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Ke, Y.; Liu, S.; Song, X.; Ming, D. Effects of HD-tDCS combined with working memory training on event-related potentials. In Proceedings of the 2020 42nd Annual International Conference of the IEEE Engineering in Medicine & Biology Society (EMBC), Montreal, QC, Canada, 20–24 July 2020; pp. 3553–3556. [Google Scholar]
- Pobric, G.; Taylor, J.R.; Ramalingam, H.M.; Pye, E.; Robinson, L.; Vassallo, G.; Jung, J.; Bhandary, M.; Szumanska-Ryt, K.; Theodosiou, L.; et al. Cognitive and Electrophysiological Correlates of Working Memory Impairments in Neurofibromatosis Type 1. J. Autism Dev. Disord. 2021, 52, 1478–1494. [Google Scholar] [CrossRef]
- Lehrl, S.; Gallwitz, A.; Blaha, L.; Fischer, B. Geistige Leistungsfähigkeit: Theorie und Messung der Biologischen Intelligenz mit dem Kurztest KAI; Vless: Ebersberg, Germany, 1991. [Google Scholar]
- Nitsche, A.M.; Liebetanz, D.; Lang, N.; Antal, A.; Tergau, F.; Paulus, W. Safety criteria for transcranial direct current stimulation (tDCS) in humans. Clin. Neurophysiol. 2003, 114, 2220–2222. [Google Scholar] [CrossRef]
- Brunoni, A.R.; Amadera, J.; Berbel, B.; Volz, M.S.; Rizzerio, B.G.; Fregni, F. A systematic review on reporting and assessment of adverse effects associated with transcranial direct current stimulation. Int. J. Neuropsychopharmacol. 2011, 14, 1133–1145. [Google Scholar] [CrossRef]
- Kirchner, W.K. Age differences in short-term retention of rapidly changing information. J. Exp. Psychol. 1958, 55, 352–358. [Google Scholar] [CrossRef]
- Iyer, M.B.; Mattu, U.; Grafman, J.H.; Lomarev, M.; Sato, S.; Wassermann, E.M. Safety and cognitive effect of frontal DC brain polarization in healthy individuals. Neurology 2005, 64, 872–875. [Google Scholar] [CrossRef]
- Priori, A.; Hallett, M.; Rothwell, J. Repetitive transcranial magnetic stimulation or transcranial direct current stimulation? Brain Stimul. 2009, 2, 241–245. [Google Scholar] [CrossRef]
- Gandiga, P.C.; Hummel, F.C.; Cohen, L.G. Transcranial DC stimulation (tDCS): A tool for double-blind sham-controlled clinical studies in brain stimulation. Clin. Neurophysiol. 2006, 117, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Delorme, A.; Makeig, S. EEGLAB: An Open Source Toolbox for Analysis of Single-Trial EEG Dynamics Including Independent Component Analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Macmillan, N.A.; Creelman, C.D. Response bias: Characteristics of detection theory, threshold theory, and "nonparametric" indexes. Psychol. Bull. 1990, 107, 401–413. [Google Scholar] [CrossRef]
- Haatveit, B.C.; Sundet, K.; Hugdahl, K.; Ueland, T.; Melle, I.; Andreassen, O.A. The validity of d prime as a working memory index: Results from the “Bergen n-back task”. J. Clin. Exp. Neuropsychol. 2010, 32, 871–880. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, N.; Opisso, E.; Pascual-Leone, Á.; Soler, M.D. Skin Lesions Induced by Transcranial Direct Current Stimulation (tDCS). Brain Stimul. 2014, 7, 765–767. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wei, Y.; Wen, J.; Li, X. Skin Burn After Single Session of Transcranial Direct Current Stimulation (tDCS). Brain Stimul. 2015, 8, 165–166. [Google Scholar] [CrossRef]
- McEvoy, L.K.; Pellouchoud, E.; Smith, M.E.; Gevins, A. Neurophysiological signals of working memory in normal aging. Cogn. Brain Res. 2001, 11, 363–376. [Google Scholar] [CrossRef]
- D’Esposito, M.; Postle, B.R. The Cognitive Neuroscience of Working Memory. Annu. Rev. Psychol. 2015, 66, 115–142. [Google Scholar] [CrossRef]
- Sarnthein, J.; Petsche, H.; Rappelsberger, P.; Shaw, G.L.; von Stein, A. Synchronization between prefrontal and posterior association cortex during human working memory. Proc. Natl. Acad. Sci. USA 1998, 95, 7092–7096. [Google Scholar] [CrossRef]
- Reinhart, R.M.G.; Nguyen, J.A. Working memory revived in older adults by synchronizing rhythmic brain circuits. Nat. Neurosci. 2019, 22, 820–827. [Google Scholar] [CrossRef]
- Jung, K.; Friston, K.J.; Pae, C.; Choi, H.H.; Tak, S.; Choi, Y.K.; Park, B.; Park, C.-A.; Cheong, C.; Park, H.-J. Effective connectivity during working memory and resting states: A DCM study. NeuroImage 2017, 169, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Polanía, R.; Nitsche, M.A.; Korman, C.; Batsikadze, G.; Paulus, W. The Importance of Timing in Segregated Theta Phase-Coupling for Cognitive Performance. Curr. Biol. 2012, 22, 1314–1318. [Google Scholar] [CrossRef] [PubMed]
- Violante, I.R.; Li, L.M.; Carmichael, D.W.; Lorenz, R.; Leech, R.; Hampshire, A.; Rothwell, J.C.; Sharp, D.J. Externally induced frontoparietal synchronization modulates network dynamics and enhances working memory performance. eLife 2017, 6, e22001. [Google Scholar] [CrossRef] [PubMed]
- Biel, A.L.; Sterner, E.; Röll, L.; Sauseng, P. Modulating verbal working memory with fronto-parietal transcranial electric stimulation at theta frequency: Does it work? Eur. J. Neurosci. 2022, 55, 405–425. [Google Scholar] [CrossRef] [PubMed]
- Röhner, F.; Breitling, C.; Rufener, K.S.; Heinze, H.-J.; Hinrichs, H.; Krauel, K.; Sweeney-Reed, C.M. Modulation of Working Memory Using Transcranial Electrical Stimulation: A Direct Comparison Between TACS and TDCS. Front. Neurosci. 2018, 12, 761. [Google Scholar] [CrossRef]
- Wild-Wall, N.; Falkenstein, M.; Gajewski, P.D. Age-Related Differences in Working Memory Performance in A 2-Back Task. Front. Psychol. 2011, 2, 186. [Google Scholar] [CrossRef]
- Dubreuil-Vall, L.; Chau, P.; Ruffini, G.; Widge, A.S.; Camprodon, J.A. tDCS to the left DLPFC modulates cognitive and physiological correlates of executive function in a state-dependent manner. Brain Stimul. 2019, 12, 1456–1463. [Google Scholar] [CrossRef]
- Andrews, S.C.; Hoy, K.E.; Enticott, P.G.; Daskalakis, Z.J.; Fitzgerald, P.B. Improving working memory: The effect of combining cognitive activity and anodal transcranial direct current stimulation to the left dorsolateral prefrontal cortex. Brain Stimul. 2011, 4, 84–89. [Google Scholar] [CrossRef]
- Boggio, P.S.; Ferrucci, R.; Rigonatti, S.P.; Covre, P.; Nitsche, M.; Pascual-Leone, A.; Fregni, F. Effects of transcranial direct current stimulation on working memory in patients with Parkinson’s disease. J. Neurol. Sci. 2006, 249, 31–38. [Google Scholar] [CrossRef]
- Hill, A.T.; Fitzgerald, P.B.; Hoy, K.E. Effects of Anodal Transcranial Direct Current Stimulation on Working Memory: A Systematic Review and Meta-Analysis of Findings from Healthy and Neuropsychiatric Populations. Brain Stimul. 2015, 9, 197–208. [Google Scholar] [CrossRef]
- Westwood, S.J.; Romani, C. Transcranial direct current stimulation (tDCS) modulation of picture naming and word reading: A meta-analysis of single session tDCS applied to healthy participants. Neuropsychologia 2017, 104, 234–249. [Google Scholar] [CrossRef] [PubMed]
- Miniussi, C.; Harris, J.A.; Ruzzoli, M. Modelling non-invasive brain stimulation in cognitive neuroscience. Neurosci. Biobehav. Rev. 2013, 37, 1702–1712. [Google Scholar] [CrossRef] [PubMed]
- de Aguiar, V.; Paolazzi, C.L.; Miceli, G. tDCS in post-stroke aphasia: The role of stimulation parameters, behavioral treatment and patient characteristics. Cortex 2015, 63, 296–316. [Google Scholar] [CrossRef]
- Tseng, P.; Hsu, T.-Y.; Chang, C.-F.; Tzeng, O.J.; Hung, D.L.; Muggleton, N.G.; Walsh, V.; Liang, W.-K.; Cheng, S.-K.; Juan, C.-H. Unleashing Potential: Transcranial Direct Current Stimulation over the Right Posterior Parietal Cortex Improves Change Detection in Low-Performing Individuals. J. Neurosci. 2012, 32, 10554–10561. [Google Scholar] [CrossRef]
- Hsu, T.-Y.; Tseng, P.; Liang, W.-K.; Cheng, S.-K.; Juan, C.-H. Transcranial direct current stimulation over right posterior parietal cortex changes prestimulus alpha oscillation in visual short-term memory task. NeuroImage 2014, 98, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Rogers, A.M.; Kasai, K.; Koji, M.; Fukuda, R.; Iwanami, A.; Nakagome, K.; Fukuda, M.; Kato, N. Executive and prefrontal dysfunction in unipolar depression: A review of neuropsychological and imaging evidence. Neurosci. Res. 2004, 50, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.-Y.; Choo, W.-C.; Fones, C.S.; Chee, M.W. fMRI Study of Maintenance and Manipulation Processes Within Working Memory in First-Episode Schizophrenia. Am. J. Psychiatry 2005, 162, 1849–1858. [Google Scholar] [CrossRef]
- Minzenberg, M.J.; Laird, A.; Thelen, S.; Carter, C.S.; Glahn, D.C. Meta-analysis of 41 Functional Neuroimaging Studies of Executive Function in Schizophrenia. Arch. Gen. Psychiatry 2009, 66, 811–822. [Google Scholar] [CrossRef]
- Potkin, S.G.; Turner, J.; Brown, G.G.; McCarthy, G.; Greve, D.N.; Glover, G.H.; Manoach, D.S.; Belger, A.; Diaz, M.; Wible, C.G.; et al. Working memory and DLPFC inefficiency in schizophrenia: The FBIRN study. Schizophr. Bull. 2008, 35, 19–31. [Google Scholar] [CrossRef]
- Jirsaraie, R.J.; Sheffield, J.M.; Barch, D.M. Neural correlates of global and specific cognitive deficits in schizophrenia. Schizophr. Res. 2018, 201, 237–242. [Google Scholar] [CrossRef]
- Cao, W.; Liao, H.; Cai, S.; Peng, W.; Liu, Z.; Zheng, K.; Liu, J.; Zhong, M.; Tan, C.; Yi, J. Increased functional interaction within frontoparietal network during working memory task in major depressive disorder. Hum. Brain Mapp. 2021, 42, 5217–5229. [Google Scholar] [CrossRef] [PubMed]
- Salehinejad, M.A.; Ghanavai, E.; Rostami, R.; Nejati, V. Cognitive control dysfunction in emotion dysregulation and psychopathology of major depression (MD): Evidence from transcranial brain stimulation of the dorsolateral prefrontal cortex (DLPFC). J. Affect. Disord. 2017, 210, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Fregni, F.; Boggio, P.S.; Nitsche, A.M.; Marcolin, A.M.; Rigonatti, S.P.; Pascual-Leone, A. Treatment of major depression with transcranial direct current stimulation. Bipolar Disord. 2006, 8, 203–204. [Google Scholar] [CrossRef] [PubMed]
- Brunoni, A.; Ferrucci, R.; Bortolomasi, M.; Vergari, M.; Tadini, L.; Boggio, P.; Giacopuzzi, M.; Barbieri, S.; Priori, A. Transcranial direct current stimulation (tDCS) in unipolar vs. bipolar depressive disorder. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 96–101. [Google Scholar] [CrossRef]
- Brunelin, J.; Mondino, M.; Gassab, L.; Haesebaert, F.; Gaha, L.; Suaud-Chagny, M.-F.; Saoud, M.; Mechri, A.; Poulet, E. Examining Transcranial Direct-Current Stimulation (tDCS) as a Treatment for Hallucinations in Schizophrenia. Am. J. Psychiatry 2012, 169, 719–724. [Google Scholar] [CrossRef]
- Smith, R.C.; Boules, S.; Mattiuz, S.; Youssef, M.; Tobe, R.H.; Sershen, H.; Lajtha, A.; Nolan, K.; Amiaz, R.; Davis, J.M. Effects of transcranial direct current stimulation (tDCS) on cognition, symptoms, and smoking in schizophrenia: A randomized controlled study. Schizophr. Res. 2015, 168, 260–266. [Google Scholar] [CrossRef]
- Zhu, R.; Luo, Y.; Wang, Z.; You, X. Within-session repeated transcranial direct current stimulation of the posterior parietal cortex enhances spatial working memory. Cogn. Neurosci. 2021, 13, 26–37. [Google Scholar] [CrossRef]
- Turner, C.; Jackson, C.; Learmonth, G. Is the “end-of-study guess” a valid measure of sham blinding during transcranial direct current stimulation? Eur. J. Neurosci. 2020, 53, 1592–1604. [Google Scholar] [CrossRef]
- Greinacher, R.; Buhôt, L.; Möller, L.; Learmonth, G. The time course of ineffective sham-blinding during low-intensity (1 mA) transcranial direct current stimulation. Eur. J. Neurosci. 2019, 50, 3380–3388. [Google Scholar] [CrossRef]
- Minarik, T.; Berger, B.; Althaus, L.; Bader, V.; Biebl, B.; Brotzeller, F.; Fusban, T.; Hegemann, J.; Jesteadt, L.; Kalweit, L.; et al. The Importance of Sample Size for Reproducibility of tDCS Effects. Front. Hum. Neurosci. 2016, 10, 453. [Google Scholar] [CrossRef]
- Medina, J.; Cason, S. No evidential value in samples of transcranial direct current stimulation (tDCS) studies of cognition and working memory in healthy populations. Cortex 2017, 94, 131–141. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voegtle, A.; Reichert, C.; Hinrichs, H.; Sweeney-Reed, C.M. Repetitive Anodal TDCS to the Frontal Cortex Increases the P300 during Working Memory Processing. Brain Sci. 2022, 12, 1545. https://doi.org/10.3390/brainsci12111545
Voegtle A, Reichert C, Hinrichs H, Sweeney-Reed CM. Repetitive Anodal TDCS to the Frontal Cortex Increases the P300 during Working Memory Processing. Brain Sciences. 2022; 12(11):1545. https://doi.org/10.3390/brainsci12111545
Chicago/Turabian StyleVoegtle, Angela, Christoph Reichert, Hermann Hinrichs, and Catherine M. Sweeney-Reed. 2022. "Repetitive Anodal TDCS to the Frontal Cortex Increases the P300 during Working Memory Processing" Brain Sciences 12, no. 11: 1545. https://doi.org/10.3390/brainsci12111545
APA StyleVoegtle, A., Reichert, C., Hinrichs, H., & Sweeney-Reed, C. M. (2022). Repetitive Anodal TDCS to the Frontal Cortex Increases the P300 during Working Memory Processing. Brain Sciences, 12(11), 1545. https://doi.org/10.3390/brainsci12111545