VDAC2 Mediates the Apoptosis of Cashmere Goat Hair Follicle Stem Cells Through the P53 Signaling Pathway


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Ethical Statement
2.2. Cell Isolation, Culture, and Transfection
2.3. Total RNA Extraction, cDNA Synthesis, Primer Design, and RT-qPCR
2.4. Overexpression Vector Construction and RNA Interference
2.5. Immunofluorescence
2.6. CCK-8 Assay
2.7. EdU Assay
2.8. Cell Cycle and Cell Apoptosis Detection
2.9. Total Protein Extraction and Western Blotting
2.10. Transcriptome Sequencing Analysis
2.11. Inhibition of P53
2.12. Statistical Analysis
2.13. Visualization and Software
3. Results
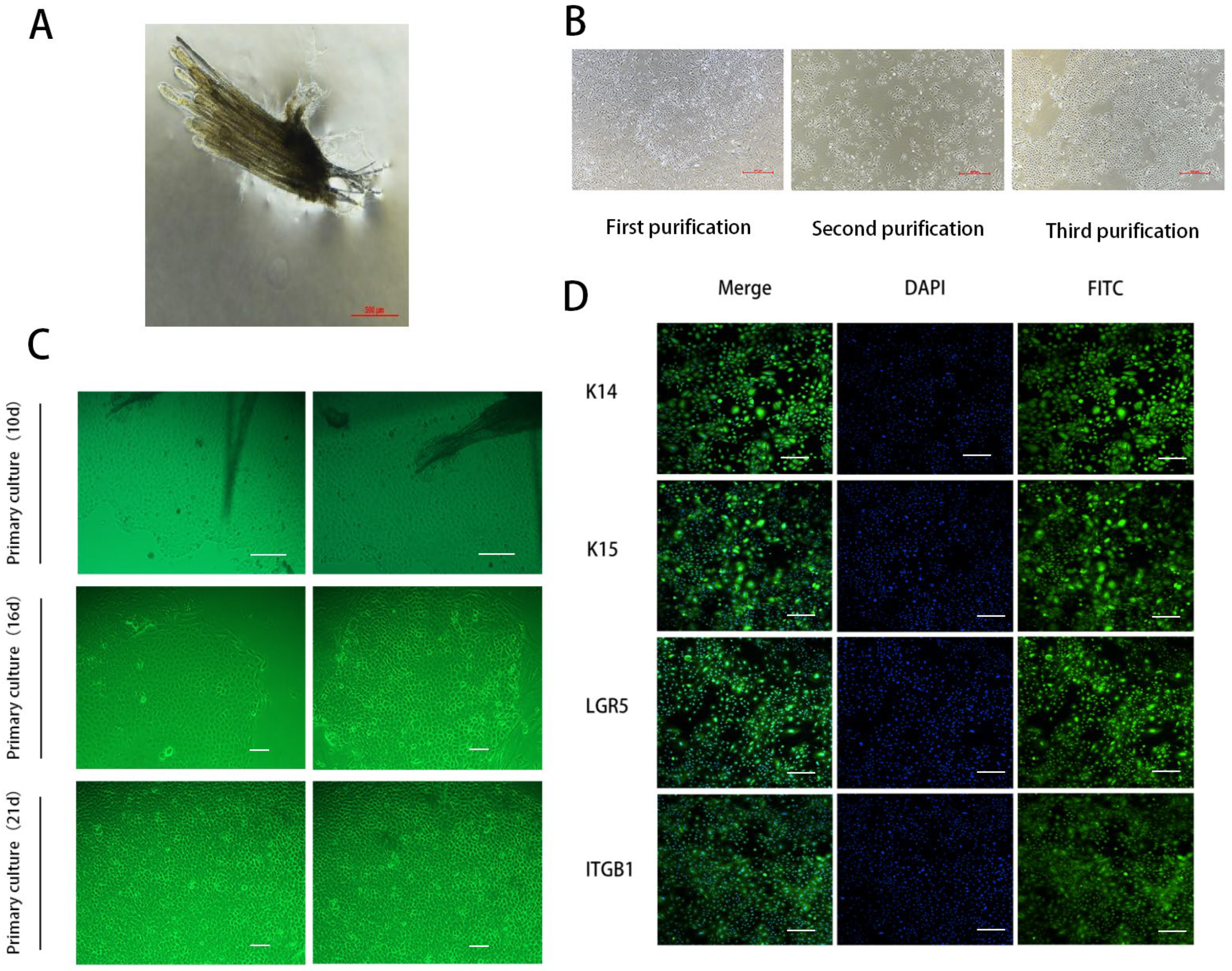
3.1. Isolation, Purification, and Identification of SHFSCs in Cashmere Goats
3.2. Impact of VDAC2 on the Growth of SHFSCs
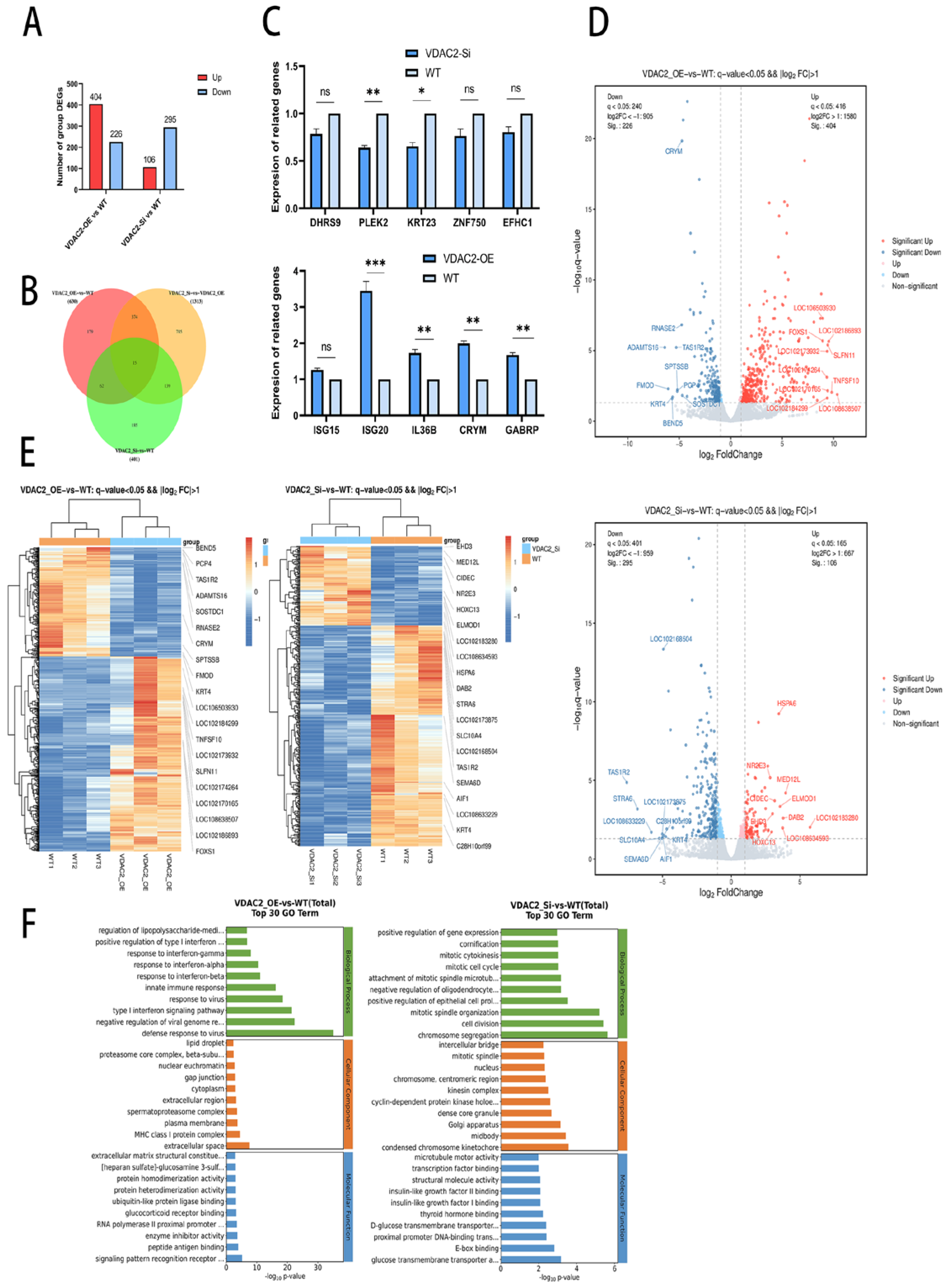
3.3. Impact of VDAC2 on Downstream Genes
3.4. Impact of VDAC2 on Signaling Pathways
3.5. Rescue of SHFSC Apoptosis Induced by VDAC2 Overexpression Using PFT
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| HF | Hair Follicle |
| HFSC | Hair Follicle Stem Cell |
| SHFSC | Secondary Hair Follicle Stem Cell |
| VDAC2 | Voltage Dependent Anion Channel 2 |
| GAPDH | Glyceraldehyde-3-phosphate |
| PFT | Pifithrin-μ |
| WT | Wild Type |
| NC | Negative Control |
| mRNA | Messenger RNA |
| PBS | Phosphate-Buffered Solution |
| PS | Penicillin-Streptomycin Solution |
| PCR | Polymerase Chain Reaction |
| RT-qPCR | Quantitative Real-time Polymerase Chain Reaction |
| EGF | Epidermal Growth Factor |
| Ins | Insulin |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequence (5′–3′) |
|---|---|
| GAPDH | F-GCAGCTACGTCTTTTACCGC |
| R-GGACGCGATCTTGGTGAAGTA | |
| VDAC2-Q | F-CAAGTCTTCTTACAAGAGGGAA |
| R-TTGGCACTGTCAAAGGTCAT | |
| Bcl2-Q | F-CACCTGACGCCCTTCACC |
| R-CCCCTCCGAACTCAAAGAA | |
| Bax-Q | F-CGGGTTGTCGCCCTTTTCTA |
| R-GAGAGGAGGCCGTCCCAAC | |
| P53-Q | F-CTCAGCACCTTATCCGGGTG |
| R-GACTCATAGGGCACCACCAC | |
| Casp3-Q | F-GGAGCAGTTTTATTTGCGTGC |
| R-TTCTGCAACAGTCCCCTCTG | |
| PUMA-Q | F-CGACGGTCCTCAGCCTTCA |
| R-CGTTGCCGCTCCTCTTGTC | |
| NOXA-Q | F-CCGGCAGATCCTGAAGTTGA |
| R-GCGGAGGAGTTTGGCTATCA |
| Sample | Clean Reads (M) | Clean Bases (G) | Valid Bases (%) | Q30 (%) | GC (%) |
|---|---|---|---|---|---|
| VDAC2-OE1 | 47.06 | 7.01 | 99.29 | 97.93 | 53.42 |
| VDAC2-OE2 | 48.3 | 7.02 | 96.72 | 98.46 | 48.05 |
| VDAC2-OE3 | 46.96 | 6.99 | 99.18 | 98.14 | 51.5 |
| VDAC2-Si1 | 46.94 | 7 | 99.42 | 97.63 | 52.69 |
| VDAC2-Si2 | 47.06 | 7 | 99.09 | 98.01 | 49.48 |
| VDAC2-Si3 | 47.38 | 7.03 | 98.92 | 98.11 | 48.94 |
| WT1 | 48.05 | 7.03 | 97.44 | 98.39 | 45.06 |
| WT2 | 46.96 | 6.9 | 97.89 | 98.02 | 48.89 |
| WT3 | 46.57 | 6.94 | 99.27 | 97.82 | 51.97 |
| Sample | Total Reads | Total Mapped | Multiple Mapped | Uniquely Mapped |
|---|---|---|---|---|
| VDAC2-OE1 | 47,055,692 | 46,369,795 (98.54%) | 2,669,412 (5.67%) | 43,700,383 (92.87%) |
| VDAC2-OE2 | 48,299,364 | 47,473,203 (98.29%) | 3,999,881 (8.28%) | 43,473,322 (90.01%) |
| VDAC2-OE3 | 46,960,902 | 46,311,246 (98.62%) | 3,563,553 (7.59%) | 42,747,693 (91.03%) |
| VDAC2-Si1 | 46,939,326 | 46,354,521 (98.75%) | 2,615,313 (5.57%) | 43,739,208 (93.18%) |
| VDAC2-Si2 | 47,055,312 | 46,392,258 (98.59%) | 3,585,096 (7.62%) | 42,807,162 (90.97%) |
| VDAC2-Si3 | 47,378,036 | 46,648,142 (98.46%) | 3,638,895 (7.68%) | 43,009,247 (90.78%) |
| WT1 | 48,050,916 | 46,931,162 (97.67%) | 6,469,682 (13.46%) | 40,461,480 (84.21%) |
| WT2 | 46,959,144 | 46,257,618 (98.51%) | 4,924,114 (10.49%) | 41,333,504 (88.02%) |
| WT3 | 46,573,670 | 45,954,427 (98.67%) | 3,742,117 (8.03%) | 42,212,310 (90.64%) |
| Interference Sites | Sequence (5′–3′) |
|---|---|
| siRNA-VDAC2-goat-568 | F:GGCGUGGAAUUCUCAACAUTT R:AUGUUGAGAAUUCCACGCCTT |
| siRNA-VDAC2-goat-699 | F:GGGAACAGAAAUCGCUAUUTT R:AAUAGCGAUUUCUGUUCCCTT |
| siRNA-VDAC2-goat-945 | F:GCUGACAAGGAAUAACUUUTT R:AAAGUUAUUCCUUGUCAGCTT |
| siRNA-VDAC2-NC | F:UUCUCCGAACGUGUCACGUTT R:ACGUGACACGUUCGGAGAATT |

References
- Ge, W.; Zhang, W.; Zhang, Y.; Zheng, Y.; Li, F.; Wang, S.; Liu, J.; Tan, S.; Yan, Z.; Wang, L.; et al. A single-cell transcriptome atlas of cashmere goat hair follicle morphogenesis. Genom. Proteom. Bioinform. 2021, 19, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Paus, R.; Foitzik, K. In search of the “hair cycle clock”: A guided tour. Differentiation 2004, 72, 489–511. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, Y.; He, J.; Wang, J.; Chen, X.; Yang, R. Regulation of signaling pathways in hair follicle stem cells. Burn. Trauma 2022, 10, tkac022. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-L.; Huang, W.-Y.; Wang, E.H.C.; Tai, K.-Y.; Lin, S.-J. Functional complexity of hair follicle stem cell niche and therapeutic targeting of niche dysfunction for hair regeneration. J. Biomed. Sci. 2020, 27, 43. [Google Scholar] [CrossRef]
- Li, K.N.; Tumbar, T. Hair follicle stem cells as a skin-organizing signaling center during adult homeostasis. EMBO J. 2021, 40, e107135. [Google Scholar] [CrossRef]
- Gentile, P.; Garcovich, S. Advances in regenerative stem cell therapy in androgenic alopecia and hair loss: Wnt pathway, growth-factor, and mesenchymal stem cell signaling impact analysis on cell growth and hair follicle development. Cells 2019, 8, 466. [Google Scholar] [CrossRef]
- Suen, W.-J.; Li, S.-T.; Yang, L.-T. Hes1 regulates anagen initiation and hair follicle regeneration through modulation of hedgehog signaling. Stem Cells 2020, 38, 301–314. [Google Scholar] [CrossRef]
- Yan, Q.; Qi, B.; Zhang, P.; Jin, Y.; Cao, K.; Liu, Y. Hair follicle stem cell proliferation and differentiation are achieved by miR-1285-3P through targeted regulation of NOTCH pathway. Prev. Med. 2023, 173, 107566. [Google Scholar] [CrossRef]
- Xiong, L.; Zhevlakova, I.; West, X.Z.; Gao, D.; Murtazina, R.; Horak, A.; Brown, J.M.; Molokotina, I.; Podrez, E.A.; Byzova, T.V. TLR2 regulates hair follicle cycle and regeneration via BMP signaling. Elife 2024, 12, RP89335. [Google Scholar] [CrossRef]
- Hu, X.M.; Li, Z.X.; Zhang, D.Y.; Yang, Y.C.; Fu, S.A.; Zhang, Z.Q.; Yang, R.H.; Xiong, K. A systematic summary of survival and death signalling during the life of hair follicle stem cells. Stem Cell Res. Ther. 2021, 12, 453. [Google Scholar] [CrossRef]
- Bertheloot, D.; Latz, E.; Franklin, B.S. Necroptosis, pyroptosis and apoptosis: An intricate game of cell death. Cell. Mol. Immunol. 2021, 18, 1106–1121. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.M.; Li, Z.X.; Lin, R.H.; Shan, J.Q.; Yu, Q.W.; Wang, R.X.; Liao, L.S.; Yan, W.T.; Wang, Z.; Shang, L.; et al. Guidelines for regulated cell death assays: A systematic summary, A categorical comparison, A prospective. Front. Cell Dev. Biol. 2021, 9, 634690. [Google Scholar] [CrossRef] [PubMed]
- Lindner, G.; Botchkarev, V.A.; Botchkareva, N.V.; Ling, G.; van der Veen, C.; Paus, R. Analysis of apoptosis during hair follicle regression (catagen). Am. J. Pathol. 1997, 151, 1601–1617. [Google Scholar]
- Kim, J.Y.; Ohn, J.; Yoon, J.-S.; Kang, B.M.; Park, M.; Kim, S.; Lee, W.; Hwang, S.; Kim, J.-I.; Kim, K.H.; et al. Priming mobilization of hair follicle stem cells triggers permanent loss of regeneration after alkylating chemotherapy. Nat. Commun. 2019, 10, 3694. [Google Scholar] [CrossRef]
- Zhao, B.; Li, J.; Zhang, X.; Dai, Y.; Yang, N.; Bao, Z.; Chen, Y.; Wu, X. Exosomal miRNA-181a-5p from the cells of the hair follicle dermal papilla promotes the hair follicle growth and development via the Wnt/β-catenin signaling pathway. Int. J. Biol. Macromol. 2022, 207, 110–120. [Google Scholar] [CrossRef]
- Li, J.; Zhao, B.; Yao, S.; Dai, Y.; Zhang, X.; Yang, N.; Bao, Z.; Cai, J.; Chen, Y.; Wu, X. Dermal PapillaCell-derived exosomes regulate hair follicle stem cell proliferation via LEF1. Int. J. Mol. Sci. 2023, 24, 3961. [Google Scholar] [CrossRef] [PubMed]
- Schein, S.J.; Colombini, M.; Finkelstein, A. Reconstitution in planar lipid bilayers of a voltage-dependent anion-selective channel obtained from paramecium mitochondria. J. Membr. Biol. 1976, 30, 99–120. [Google Scholar] [CrossRef]
- Baker, M.A.; Lane, D.J.R.; Ly, J.D.; De Pinto, V.; Lawen, A. VDAC1 is a transplasma membrane NADH-ferricyanide reductase. J. Biol. Chem. 2004, 279, 4811–4819. [Google Scholar] [CrossRef]
- Luo, P.; Zhang, Q.; Shen, S.; An, Y.; Yuan, L.; Wong, Y.K.; Huang, S.; Huang, S.; Huang, J.; Cheng, G.; et al. Mechanistic engineering of celastrol liposomes induces ferroptosis and apoptosis by directly targeting VDAC2 in hepatocellular carcinoma. Asian J. Pharm. Sci. 2023, 18, 100874. [Google Scholar] [CrossRef]
- Ren, D.; Kim, H.; Tu, H.-C.; Westergard, T.D.; Fisher, J.K.; Rubens, J.A.; Korsmeyer, S.J.; Hsieh, J.J.-D.; Cheng, E.H.-Y. The VDAC2-BAK rheostat controls thymocyte survival. Sci. Signal. 2009, 2, ra48. [Google Scholar] [CrossRef]
- Martin, L.J.; Adams, N.A.; Pan, Y.; Price, A.; Wong, M. The mitochondrial permeability transition pore regulates nitric oxide-mediated apoptosis of neurons induced by target deprivation. J. Neurosci. 2011, 31, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Yan, W. Study on the Mechanism of Hair Follicle Stem Cell Activation by Hair Papilla Cells in Albas Velvet Goats; Inner Mongolia University: Hohhot, China, 2023. [Google Scholar]
- Strom, E.; Sathe, S.; Komarov, P.G.; Chernova, O.B.; Pavlovska, I.; Shyshynova, I.; Bosykh, D.A.; Burdelya, L.G.; Macklis, R.M.; Skaliter, R.; et al. Small-molecule inhibitor of P53 binding to mitochondria protects mice from gamma radiation. Nat. Chem. Biol. 2006, 2, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Cotsarelis, G.; Sun, T.T.; Lavker, R.M. Label. Label-retaining cells reside in the bulge area of pilosebaceous unit: Implications for follicular stem cells, hair cycle, and skin carcinogenesis. Cell 1990, 61, 1329–1337. [Google Scholar] [CrossRef]
- Ohyama, M. Hair follicle bulge: A fascinating reservoir of epithelial stem cells. J. Dermatol. Sci. 2007, 46, 81–89. [Google Scholar] [CrossRef]
- Taylor, G.; Lehrer, M.S.; Jensen, P.J.; Sun, T.T.; Lavker, R.M. Involvement of follicular stem cells in forming not only the follicle but also the epidermis. Cell 2000, 102, 451–461. [Google Scholar] [CrossRef]
- Chovatiya, G.; Sunkara, R.R.; Roy, S. Context-dependent effect of sPLA-IIA induced proliferation on murine hair follicle stem cells and human epithelial cancer. EBioMedicine 2019, 48, 364–376. [Google Scholar] [CrossRef]
- López-Paniagua, M.; Nieto-Miguel, T.; de la Mata, A. Consecutive expansion of limbal epithelial stem cells from a single limbal biopsy. Curr. Eye Res. 2013, 38, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Joost, S.; Jacob, T.; Sun, X. Single-Cell Transcriptomics of Traced Epidermal and Hair Follicle Stem Cells Reveals Rapid Adaptations during Wound Healing. Cell Rep. 2018, 25, 585–597. [Google Scholar] [CrossRef]
- He, N.; Dong, Z.; Tao, L. Isolation and characterization of hair follicle stem cells from Arbas Cashmere goat. Cytotechnology 2016, 68, 2579–2588. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, P.; Gong, Y. Curcumin-zinc framework encapsulated microneedle patch for promoting hair growth. Theranostics 2023, 13, 3675–3688. [Google Scholar] [CrossRef]
- Glushakova, O.Y.; Glushakov, A.O.; Borlongan, C.V.; Valadka, A.B.; Hayes, R.L.; Glushakov, A.V. Role of caspase-3-mediated apoptosis in chronic caspase-3-cleaved tau accumulation and blood-brain barrier damage in the corpus callosum after traumatic brain injury in rats. J. Neurotrauma 2018, 35, 157–173. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Zhang, B.; Ma, S.; Gonzalez-Celeiro, M.; Stein, D.; Jin, X.; Kim, S.T.; Kang, Y.L.; Besnard, A.; Rezza, A.; et al. Corticosterone inhibits GAS6 to govern hair follicle stem-cell quiescence. Nature 2021, 592, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Greco, V.; Chen, T.; Rendl, M.; Schober, M.; Pasolli, H.A.; Stokes, N.; Dela Cruz-Racelis, J.; Fuchs, E. A two-step mechanism for stem cell activation during hair regeneration. Cell Stem Cell 2009, 4, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Tang, X.; Zhang, S.; Jin, M.; Wang, M.; Deng, Z.; Liu, Z.; Qian, M.; Shi, W.; Wang, Z.; et al. SIRT 7 activates quiescent hair follicle stem cells to ensure hair growth in mice. EMBO J. 2020, 39, e104365. [Google Scholar] [CrossRef]
- Anflous, K.; Armstrong, D.D.; Craigen, W.J. Altered mitochondrial sensitivity for ADP and maintenance of creatine-stimulated respiration in oxidative striated muscles from VDAC1-deficient mice. J. Biol. Chem. 2001, 276, 1954–1960. [Google Scholar] [CrossRef]
- Sampson, M.J.; Decker, W.K.; Beaudet, A.L.; Ruitenbeek, W.; Armstrong, D.; Hicks, M.J.; Craigen, W.J. Immotile sperm and infertility in mice lacking mitochondrial voltage-dependent anion channel type 3. J. Biol. Chem. 2001, 276, 39206–39212. [Google Scholar] [CrossRef]
- Weeber, E.J.; Levy, M.; Sampson, M.J.; Anflous, K.; Armstrong, D.L.; Brown, S.E.; Sweatt, J.D.; Craigen, W.J. The role of mitochondrial porins and the permeability transition pore in learning and synaptic plasticity. J. Biol. Chem. 2002, 277, 18891–18897. [Google Scholar] [CrossRef]
- Chin, H.S.; Li, M.X.; Tan, I.K.L.; Ninnis, R.L.; Reljic, B.; Scicluna, K.; Dagley, L.F.; Sandow, J.J.; Kelly, G.L.; Samson, A.L.; et al. VDAC2 enables BAX to mediate apoptosis and limit tumor development. Nat. Commun. 2018, 9, 4976. [Google Scholar] [CrossRef]
- Ma, S.B.; Nguyen, T.N.; Tan, I.; Ninnis, R.; Iyer, S.; Stroud, D.A.; Menard, M.; Kluck, R.M.; Ryan, M.T.; Dewson, G. Bax targets mitochondria by distinct mechanisms before or during apoptotic cell death: A requirement for VDAC2 or Bak for efficient Bax apoptotic function. Cell Death Differ. 2014, 21, 1925–1935. [Google Scholar] [CrossRef]
- Roy, S.S.; Ehrlich, A.M.; Craigen, W.J.; Hajnóczky, G. VDAC2 is required for truncated BID-induced mitochondrial apoptosis by recruiting BAK to the mitochondria. EMBO Rep. 2009, 10, 1341–1347. [Google Scholar] [CrossRef]
- Yamagata, H.; Shimizu, S.; Nishida, Y.; Watanabe, Y.; Craigen, W.J.; Tsujimoto, Y. Requirement of voltage-dependent anion channel 2 for pro-apoptotic activity of Bax. Oncogene 2009, 28, 3563–3572. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Luo, M.; Zhang, K.; Zhang, J.; Gao, T.; Connell, D.O.; Yao, F.; Mu, C.; Cai, B.; Shang, Y.; et al. Nedd4 ubiquitylates VDAC2/3 to suppress erastin-induced ferroptosis in melanoma. Nat. Commun. 2020, 11, 433. [Google Scholar] [CrossRef] [PubMed]
- Mihara, M.; Erster, S.; Zaika, A.; Petrenko, O.; Chittenden, T.; Pancoska, P.; Moll, U.M. P53 has a direct apoptogenic role at the mitochondria. Mol. Cell 2003, 11, 577–590. [Google Scholar] [CrossRef]
- Olie, R.A.; Hafner, C.; Küttel, R.; Sigrist, B.; Willers, J.; Dummer, R.; Hall, J.; Stahel, R.A.; Zangemeister-Wittke, U. Bcl-2 and bcl-xL antisense oligonucleotides induce apoptosis in melanoma cells of different clinical stages. J. Investig. Dermatol. 2002, 118, 505–512. [Google Scholar] [CrossRef]
- Adams, J.M.; Cory, S. The Bcl-2 protein family: Arbiters of cell survival. Science 1998, 281, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Kalpage, H.A.; Bazylianska, V.; Recanati, M.A.; Fite, A.; Liu, J.; Wan, J.; Mantena, N.; Malek, M.H.; Podgorski, I.; Heath, E.I.; et al. Tissue-specific regulation of cytochrome c by post-translational modifications: Respiration, the mitochondrial membrane potential, ROS, and apoptosis. FASEB J. 2019, 33, 1540–1553. [Google Scholar] [CrossRef]
- Tomita, Y.; Marchenko, N.; Erster, S.; Nemajerova, A.; Dehner, A.; Klein, C.; Pan, H.; Kessler, H.; Pancoska, P.; Moll, U.M. WT P53, but not tumor-derived mutants, bind to Bcl2 via the DNA binding domain and induce mitochondrial permeabilization. J. Biol. Chem. 2006, 281, 8600–8606. [Google Scholar] [CrossRef]
- Guerrache, A.; Micheau, O. TNF-related apoptosis-inducing ligand: Non-apoptotic signalling. Cells 2024, 13, 521. [Google Scholar] [CrossRef]
- Norbury, C.J.; Hickson, I.D. Cellular responses to DNA damage. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 367–401. [Google Scholar] [CrossRef]
- Hao, Q.; Chen, J.; Lu, H.; Zhou, X. The ARTS of P53-dependent mitochondrial apoptosis. J. Mol. Cell Biol. 2023, 14, mjac074. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, L.; Zhao, Y.; Zhou, M.; Guo, X.; Zhang, Y.; Liu, D.; Guo, X. VDAC2 Mediates the Apoptosis of Cashmere Goat Hair Follicle Stem Cells Through the P53 Signaling Pathway. Animals 2025, 15, 1671. https://doi.org/10.3390/ani15111671
Zhu L, Zhao Y, Zhou M, Guo X, Zhang Y, Liu D, Guo X. VDAC2 Mediates the Apoptosis of Cashmere Goat Hair Follicle Stem Cells Through the P53 Signaling Pathway. Animals. 2025; 15(11):1671. https://doi.org/10.3390/ani15111671
Chicago/Turabian StyleZhu, Long, Yueqi Zhao, Mei Zhou, Xiaotong Guo, Yinxian Zhang, Dongjun Liu, and Xudong Guo. 2025. "VDAC2 Mediates the Apoptosis of Cashmere Goat Hair Follicle Stem Cells Through the P53 Signaling Pathway" Animals 15, no. 11: 1671. https://doi.org/10.3390/ani15111671
APA StyleZhu, L., Zhao, Y., Zhou, M., Guo, X., Zhang, Y., Liu, D., & Guo, X. (2025). VDAC2 Mediates the Apoptosis of Cashmere Goat Hair Follicle Stem Cells Through the P53 Signaling Pathway. Animals, 15(11), 1671. https://doi.org/10.3390/ani15111671