
Description of a New Species of the Pareas hamptoni Complex from Yunnan, China, with Confirmation of P. hamptoni Sensu Stricto in China (Squamata, Pareidae) †

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
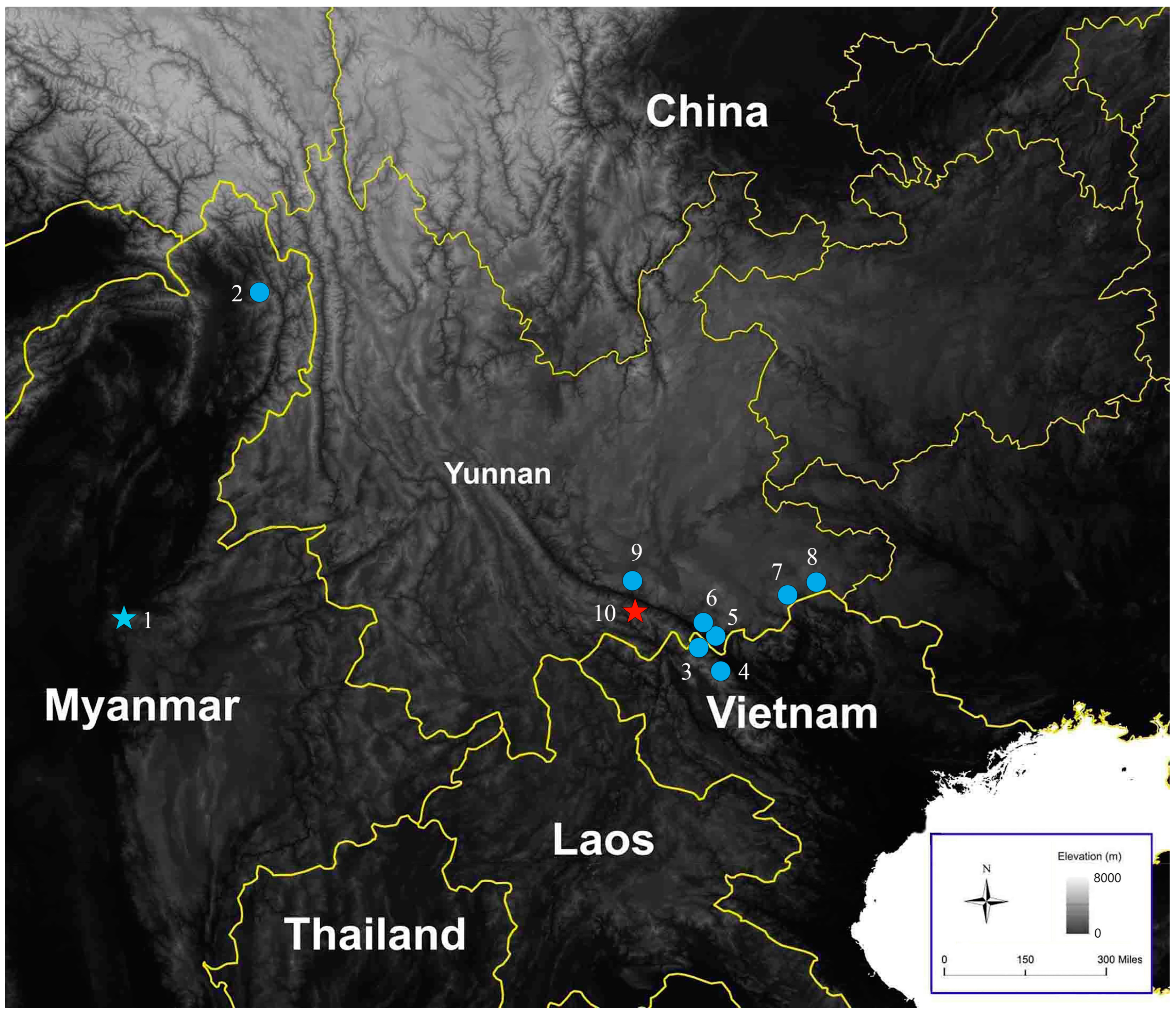
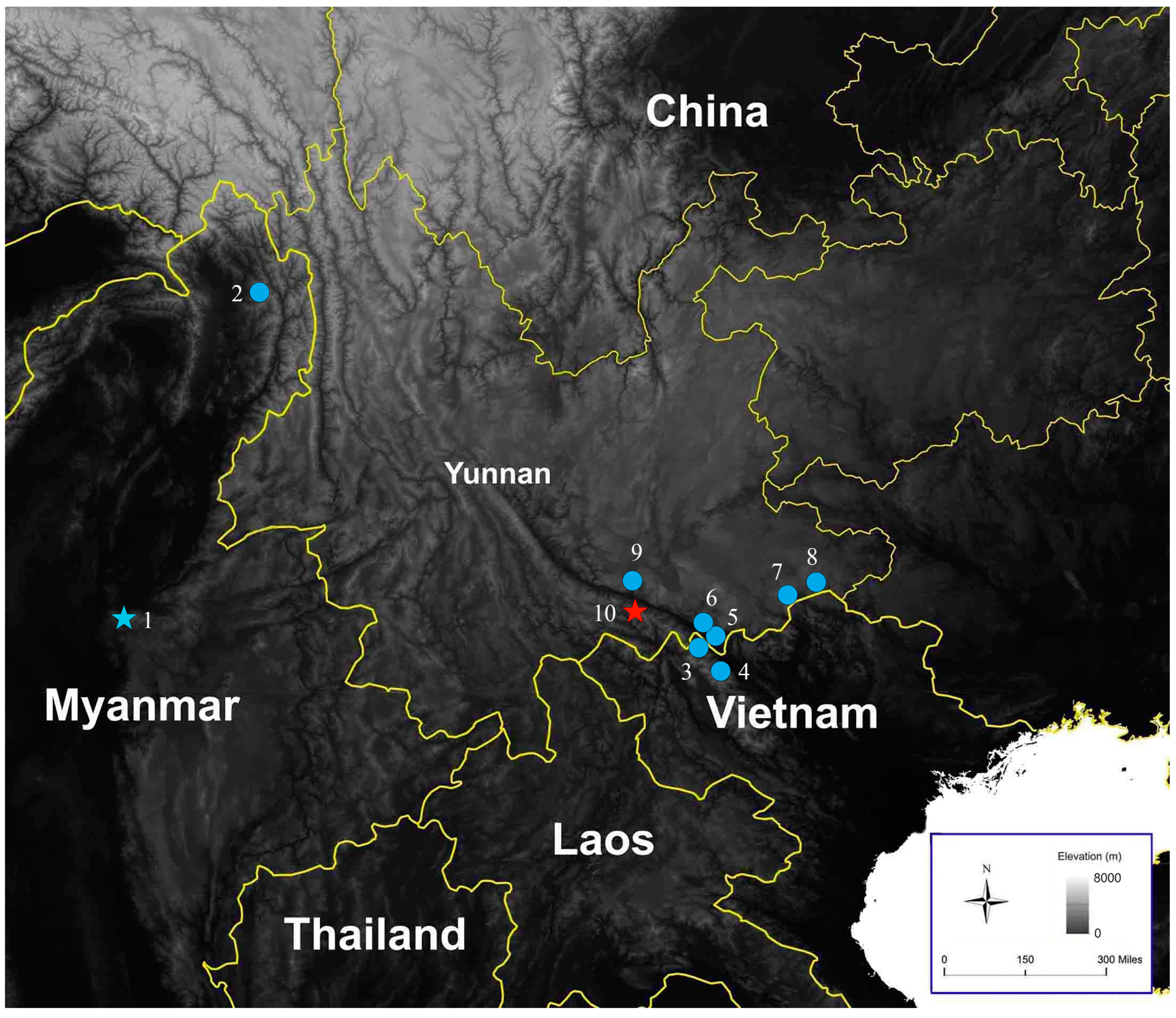
2.1. Sampling
2.2. Morphometrics
2.3. Molecular Analyses
3. Results
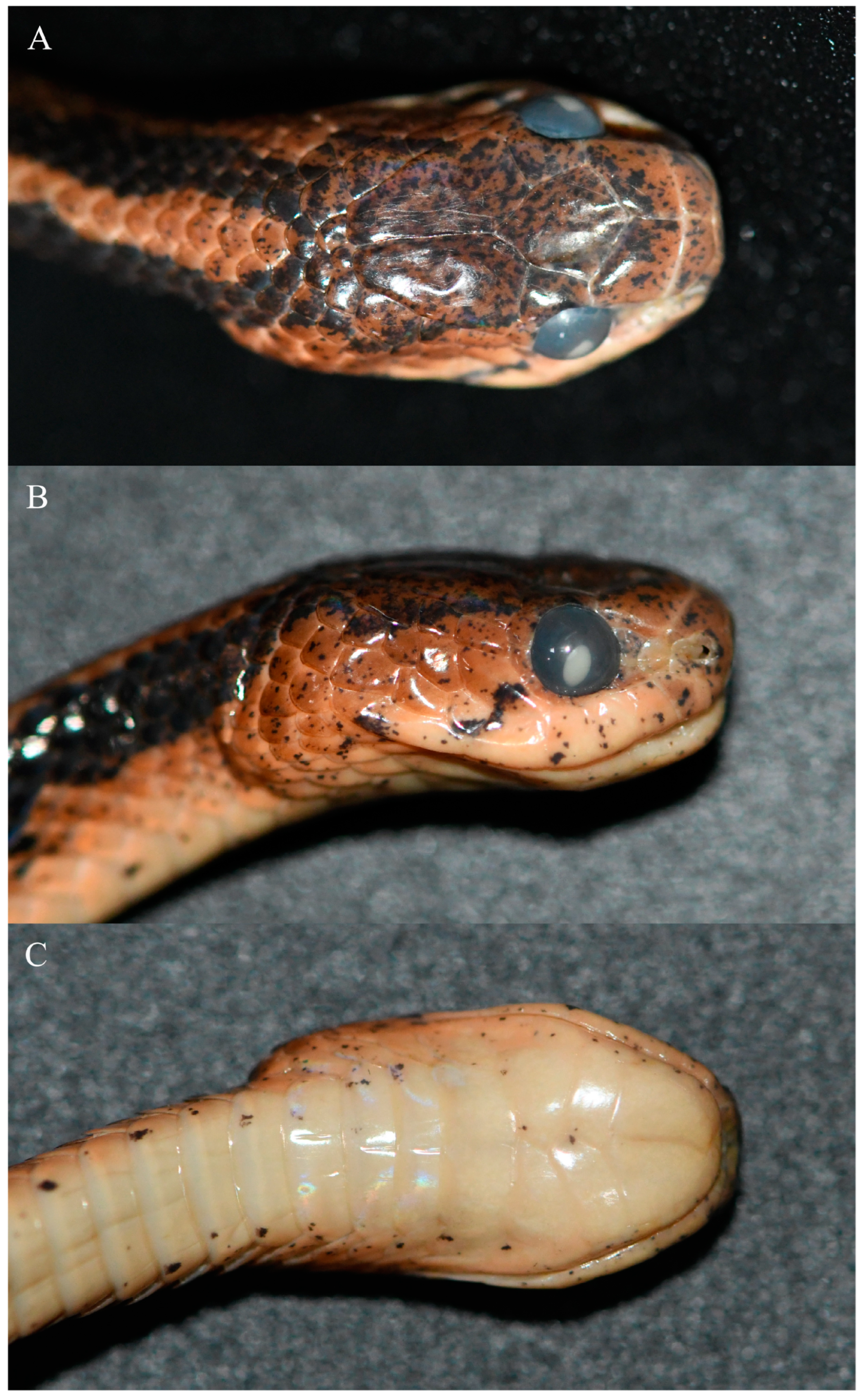
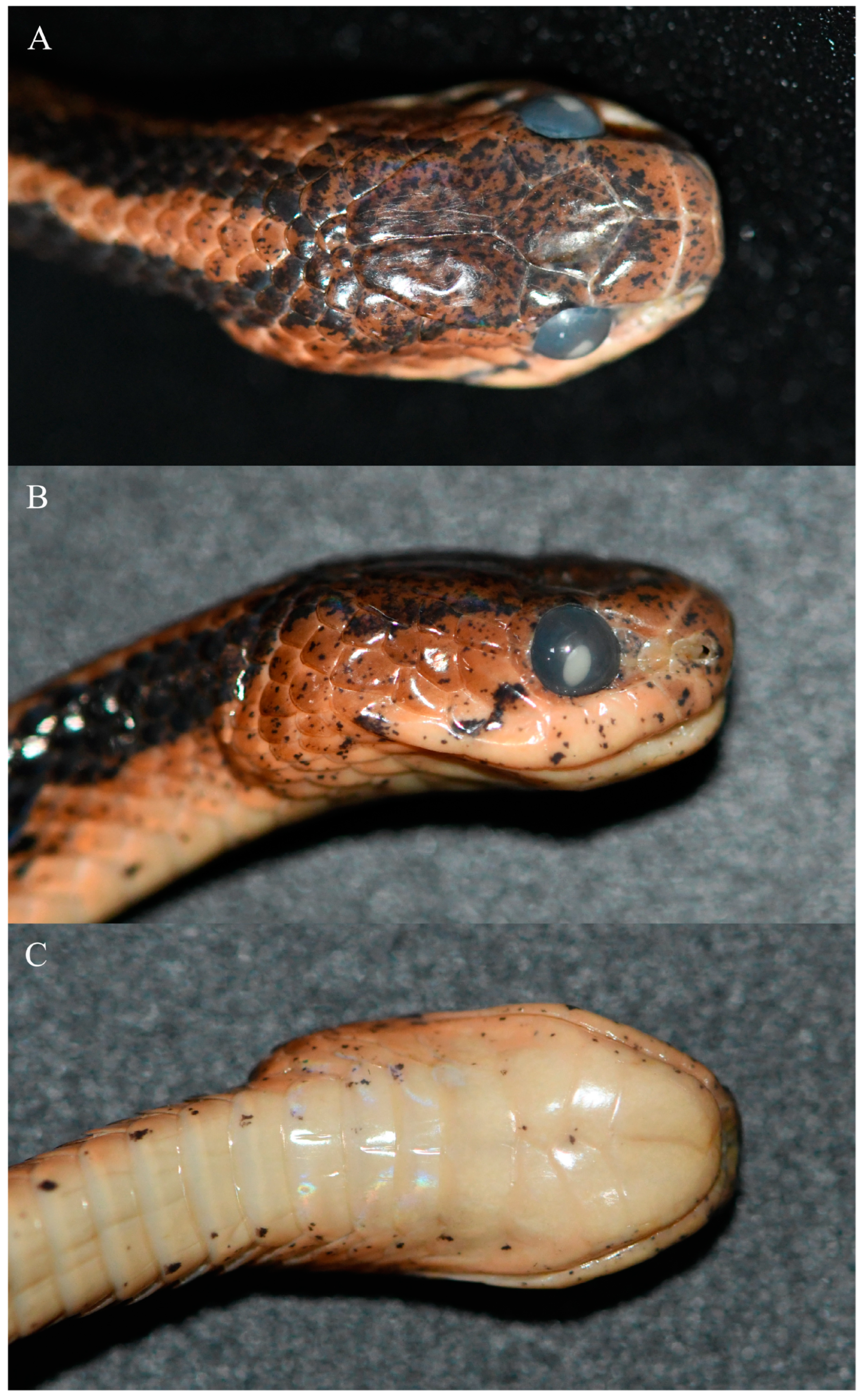
3.1. Morphological Results
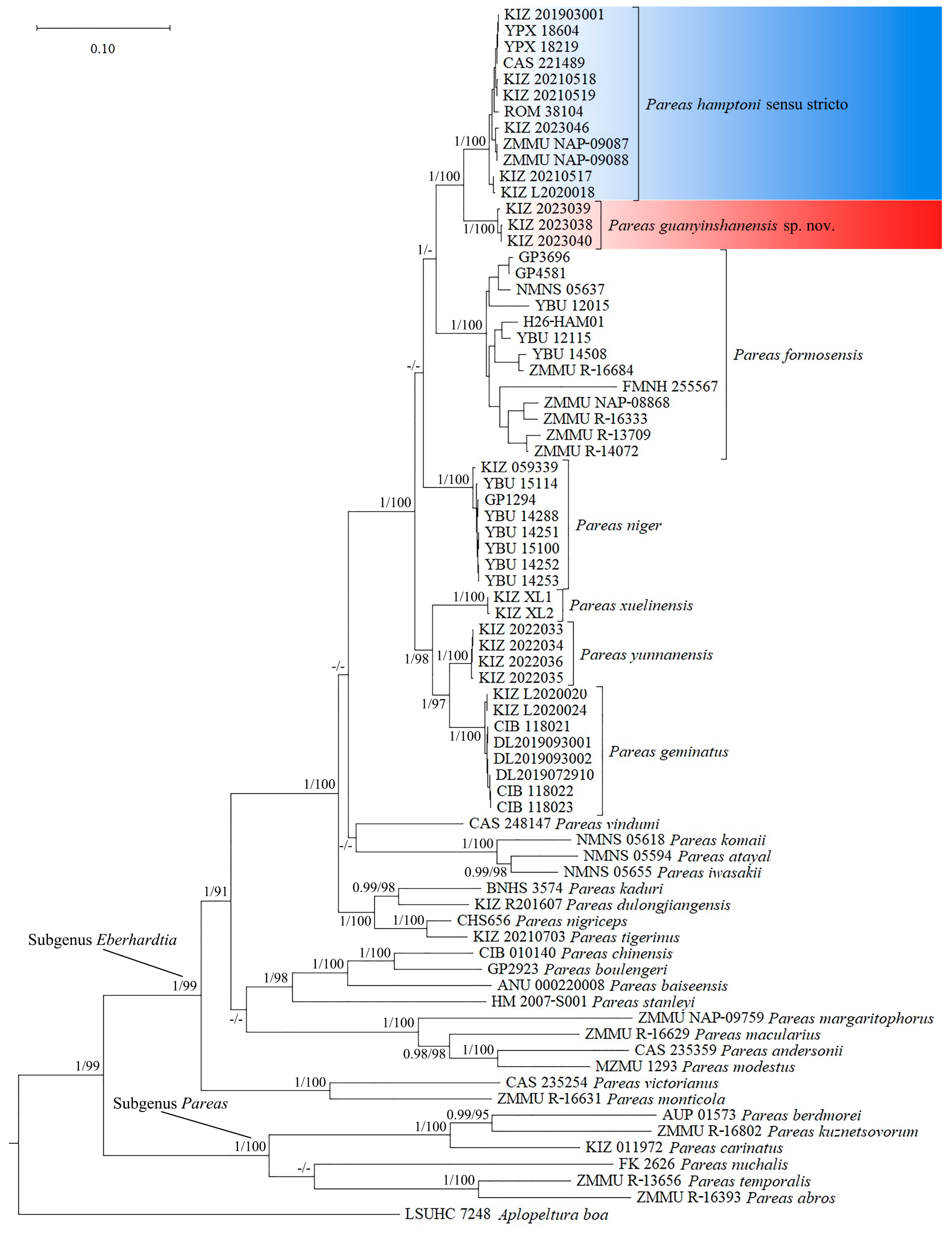
3.2. Molecular Results
3.3. Systematics
4. Discussion
5. Conclusions
| 1 | Dorsal head solid black........................................................................................................................2 |
| Dorsal head yellowish brown to reddish brown.............................................................................3 | |
| 2 | Dorsal body almost solid black........................................................................................Pareas niger |
| Dorsal body yellow to brownish red with black bars......................................Pareas yunnanensis | |
| 3 | Vertebral scales not enlarged..................................................................................Pareas xuelinensis |
| Three rows of vertebral scales enlarged...............................................................Pareas formosensis | |
| One row of vertebral scales enlarged................................................................................................4 | |
| 4 | Subcaudals more than 91....................................................................Pareas hamptoni sensu stricto |
| Subcaudals less than 91.......................................................................................................................5 | |
| 5 | Ventrals less than 188................................................................................................Pareas geminatus |
| Ventrals more than 189..................................................................Pareas guanyinshanensis sp. nov. |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, K.T.; Georgalis, G.L. The Diversity and Distribution of Palaeogene Snakes—A Review, with Comments on Vertebral Sufficiency. In The Origin and Early Evolution of Snakes; Gower, D., Zaher, H., Eds.; Cambridge University Press: Cambridge, UK, 2022; pp. 55–84. [Google Scholar]
- Zheng, Y.; Wiens, J.J. Combining phylogenomic and supermatrix approaches, and a time-calibrated phylogeny for squamate reptiles (lizards and snakes) based on 52 genes and 4162 species. Mol. Phylogenet. Evol. 2016, 94, 537–547. [Google Scholar] [CrossRef]
- Deepak, V.; Ruane, S.; Gower, D.J. A New Subfamily of Fossorial Colubroid Snakes from the Western Ghats of Peninsular India. J. Nat. Hist. 2019, 52, 2919–2934. [Google Scholar] [CrossRef]
- Zaher, H.; Murphy, R.W.; Arredondo, J.C.; Graboski, R.; Machado-Filho, P.R.; Mahlow, K.; Montingelli, G.G.; Quadros, A.B.; Orlov, N.L.; Wilkinson, M.; et al. Large-Scale Molecular Phylogeny, Morphology, Divergence-Time Estimation, and the Fossil Record of Advanced Caenophidian Snakes (Squamata: Serpentes). PLoS ONE 2019, 14, e0216148. [Google Scholar] [CrossRef]
- Poyarkov, N.A.; Nguyen, T.V.; Pawangkhanant, P.; Yushchenko, P.V.; Brakels, P.; Nguyen, L.H.; Nguyen, H.N.; Suwannapoom, C.; Orlov, N.; Vogel, G. An integrative taxonomic revision of slug-eating snakes (Squamata: Pareidae: Pareineae) reveals unprecedented diversity in Indochina. PeerJ 2022, 10, e12713. [Google Scholar] [CrossRef]
- Wang, P.; Che, J.; Liu, Q.; Li, K.; Jin, J.Q.; Jiang, K.; Shi, L.; Guo, P. A revised taxonomy of Asia snail-eating snakes Pareas (Squamata, Pareidae): Evidence from morphological comparison and molecular phylogeny. ZooKeys 2020, 939, 45–64. [Google Scholar] [CrossRef]
- Hoso, M.; Asami, T.; Hori, M. Right-handed snakes: Convergent evolution of asymmetry for functional specialization. Biol. Lett. 2007, 3, 169–172. [Google Scholar] [CrossRef]
- Wallach, V.; Williams, K.L.; Boundy, J. Snakes of the World. A Catalogue of Living and Extinct Species; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Vogel, G. A New Montane Species of the Genus Pareas Wagler, 1830 (Squamata: Pareatidae) from Northern Myanmar. Taprobanica 2015, 7, 1–7. [Google Scholar] [CrossRef]
- You, C.W.; Poyarkov, N.A.; Lin, S.M. Diversity of the snail-eating snakes Pareas (Serpentes, Pareatidae) from Taiwan. Zool. Scr. 2015, 44, 349–361. [Google Scholar] [CrossRef]
- Ding, L.; Chen, Z.; Suwannapoom, C.; Nguyen, T.V.; Poyarkov, N.A.; Vogel, G. A new species of the Pareas hamptoni complex (Squamata Serpentes: Pareidae) from the Golden Triangle. Taprobanica 2020, 9, 174–193. [Google Scholar] [CrossRef]
- Liu, S.; Rao, D.Q. A new species of the genus Pareas (Squamata, Pareidae) from Yunnan, China. ZooKeys 2021, 1011, 121–138. [Google Scholar] [CrossRef]
- Vogel, G.; Nguyen, T.V.; Zaw, T.; Poyarkov, N.A. A new species of the Pareas monticola complex (Squamata, Serpentes, Pareidae) from Chin Mountains with additions to the Pareas fauna of Myanmar. J. Nat. Hist. 2021, 54, 2577–2612. [Google Scholar] [CrossRef]
- Gong, Y.; Wu, J.; Huang, S.; Xu, Y.; Yang, D.; Liu, Y.; Liang, S.; Lee, P. A New Species of Pareas (Squamata, Pareidae) from Guangxi Province, China. Animals 2023, 13, 2233. [Google Scholar] [CrossRef] [PubMed]
- Uetz, P. The Reptile Database. Available online: http://www.reptile-database.org (accessed on 2 November 2023).
- Le, D.T.T.; Tran, T.G.; Hoang, H.D.; Stuart, B.L. A New Species of Pareas (Squamata, Pareidae) from Southern Vietnam. Vertebr. Zool. 2021, 71, 439–451. [Google Scholar] [CrossRef]
- Boulenger, G.A. Descriptions of Two New Snakes from Upper Burma. Bombay Nat. Hist. Soc. 1905, 16, 235–236. [Google Scholar] [CrossRef]
- Nguyen, S.V.; Ho, C.T.; Nguyen, T.Q. Herpetofauna of Vietnam; Edition Chimaira: Frankfurt, Germany, 2009. [Google Scholar]
- Zhao, E.M.; Huang, M.H.; Zong, Y.; Jiang, Y.M.; Huang, Q.Y.; Zhao, H.; Ma, J.F.; Zheng, J.; Huang, Z.J.; Wei, G.; et al. Fauna Sinica, Reptilia, Squamata. Serpentes; Science Press: Beijing, China, 1998. [Google Scholar]
- Zhao, E.M. Snakes of China; Anhui Science Technology Publishing House: Hefei, China, 2006. [Google Scholar]
- Yang, D.T.; Rao, D.Q. Amphibia and Reptilia of Yunnan; Yunnan Science and Technology Press: Kuming, China, 2008. [Google Scholar]
- Liu, S.; Zhang, D.R.; Poyarkov, N.A.; Hou, M.; Wu, L.; Rao, D.Q.; Nguyen, T.V.; Vogel, G. Resurrection of Pareas yunnanensis (Vogt, 1922) with Description of a New Species of Pareas from Yunnan Province, China (Squamata, Pareidae). Eur. J. Taxon. 2023, 860, 1–26. [Google Scholar] [CrossRef]
- Liu, S.; Yang, M.J.; Rao, J.Q.; Guo, Y.H.; Rao, D.Q. A New Species of Pareas Wagler, 1830 (Squamata, Pareidae) from Northwestern Yunnan, China. Taxonomy 2023, 3, 169–182. [Google Scholar] [CrossRef]
- Queiroz, A.D.; Lawson, R.; Lemos-Espinal, J.A. Phylogenetic relationships of North American garter snakes (Thamnophis) based on four mitochondrial genes: How much DNA is enough? Mol. Phylogenet. Evol. 2002, 22, 315–329. [Google Scholar] [CrossRef]
- Salvi, D.; Harris, D.J.; Kaliontzopoulou, A.; Carretero, M.A.; Pinho, C. Persistence across Pleistocene ice ages in Mediterranean and extra-Mediterranean refugia: Phylogeographic insights from the common wall lizard. BMC Evol. Biol. 2013, 13, 147. [Google Scholar] [CrossRef]
- Burland, T.G. DNASTAR’s Lasergene sequence analysis software. Methods Mol. Biol. 2000, 132, 71–91. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment Through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Voucher | Locality | Cyt b | ND4 |
|---|---|---|---|---|
| Pareas guanyinshanensis sp. nov. | KIZ 2023038 | Yuanyang, Yunnan, China | PP215390 | PP215399 |
| Pareas guanyinshanensis sp. nov. | KIZ 2023039 | Yuanyang, Yunnan, China | PP215389 | PP215398 |
| Pareas guanyinshanensis sp. nov. | KIZ 2023040 | Yuanyang, Yunnan, China | PP215388 | PP215397 |
| Pareas abros | ZMMU R-16393 | Song Thanh, Quang Nam, Vietnam | MZ712235 | MZ712262 |
| Pareas andersonii | CAS 235359 | Natmataung, Chin, Myanmar | MT968772 | MW287040 |
| Pareas atayal | NMNS 05594 | Beiheng, Taoyuan, Taiwan | KJ642124 | MW287041 |
| Pareas baiseensis | ANU 000220008 | Baise, Guangxi, China | OQ054328 | OQ054329 |
| Pareas berdmorei | AUP 01573 | Chiang Mai, Thailand | MZ712218 | MZ712244 |
| Pareas boulengeri | GP2923 | Jiangkou, Guizhou, China | MK135090 | MK805355 |
| Pareas carinatus | KIZ 011972 | Malaysia | MK135111 | MK805376 |
| Pareas chinensis | CIB 010140 | Tianquan, Sichuan, China | JF827691 | JF827669 |
| Pareas dulongjiangensis | KIZ R201607 | Gongshan, Yunnan, China | OQ718498 | — |
| Pareas formosensis | NMNS 05637 | Nantou, Taiwan | MW287060 | MW287042 |
| Pareas formosensis | H26-HAM01 | Guangdong, China | MW287061 | MW287043 |
| Pareas formosensis | YBU 12015 | Hainan, China | MK135068 | MK805333 |
| Pareas formosensis | GP4581 | Jingning, Zhejiang, China | MK135072 | MK805337 |
| Pareas formosensis | YBU 12115 | Rongjiang, Guizhou, China | MK135075 | MK805340 |
| Pareas formosensis | YBU 14508 | Guangxi, China | MK135076 | MK805341 |
| Pareas formosensis | GP3696 | Yanshan, Jiangxi, China | MH046857 | MK805382 |
| Pareas formosensis | ZMMU R-16333 | Kon Chu Rang, Gia Lai, Vietnam | MW287066 | MW287048 |
| Pareas formosensis | ZMMU NAP-08868 | Song Thanh, Quang Nam, Vietnam | MW287063 | MW287045 |
| Pareas formosensis | ZMMU R-16684 | Phia Oac, Cao Bang, Vietnam | MW287062 | MW287044 |
| Pareas formosensis | ZMMU R-13709 | Bidoup–Nui Ba, Lam Dong, Vietnam | MW287064 | MW287046 |
| Pareas formosensis | ZMMU R-14072 | Chu Yang Sin, Dak Lak, Vietnam | MW287065 | MW287047 |
| Pareas formosensis | FMNH 255567 | Pu Mat, Nghe An, Vietnam | AY425806 | — |
| Pareas geminatus | DL2019072910 | Jiangcheng, Yunnan, China | MW287067 | — |
| Pareas geminatus | DL2019093001 | Jiangcheng, Yunnan, China | MW287071 | — |
| Pareas geminatus | DL2019093002 | Jiangcheng, Yunnan, China | MW287072 | — |
| Pareas geminatus | CIB 118021 | Jiangcheng, Yunnan, China | MW287068 | — |
| Pareas geminatus | CIB 118022 | Jiangcheng, Yunnan, China | MW287069 | — |
| Pareas geminatus | CIB 118023 | Jiangcheng, Yunnan, China | MW287070 | — |
| Pareas geminatus | KIZ L2020020 | Jiangcheng, Yunnan, China | MW436707 | — |
| Pareas geminatus | KIZ L2020024 | Jiangcheng, Yunnan, China | MW436708 | — |
| Pareas hamptoni sensu stricto | YPX 18604 | Kachin, Myanmar | MK135078 | MK805343 |
| Pareas hamptoni sensu stricto | YPX 18219 | Kachin, Myanmar | MK135077 | MK805342 |
| Pareas hamptoni sensu stricto | CAS 221489 | Putao, Kachin, Myanmar | MW287077 | — |
| Pareas hamptoni sensu stricto | ZMMU NAP-09087 | Bat Xat, Lao Cai, Vietnam | MW287078 | MW287054 |
| Pareas hamptoni sensu stricto | ZMMU NAP-09088 | Bat Xat, Lao Cai, Vietnam | MW287079 | MW287053 |
| Pareas hamptoni sensu stricto | ROM 38104 | Sa Pa, Lao Cai, Vietnam | KX694896 | — |
| Pareas hamptoni sensu stricto | KIZ 20210517 | Malipo, Yunnan, China | PP215386 | PP215395 |
| Pareas hamptoni sensu stricto | KIZ L2020018 | Malipo, Yunnan, China | PP215382 | PP215391 |
| Pareas hamptoni sensu stricto | KIZ 201903001 | Hekou, Yunnan, China | PP215383 | PP215392 |
| Pareas hamptoni sensu stricto | KIZ 20210518 | Hekou, Yunnan, China | PP215385 | PP215394 |
| Pareas hamptoni sensu stricto | KIZ 20210519 | Hekou, Yunnan, China | PP215384 | PP215393 |
| Pareas hamptoni sensu stricto | KIZ 2023046 | Jianshui, Yunnan, China | PP215387 | PP215396 |
| Pareas iwasakii | NMNS 05655 | Ishigaki, Okinawa, Japan | KJ642160 | — |
| Pareas kaduri | BNHS 3574 | Lohit, Arunachal Pradesh, India | MW026190 | — |
| Pareas komaii | NMNS 05618 | Lijia, Taitung, Taiwan | KJ642185 | MW287056 |
| Pareas kuznetsovorum | ZMMU R-16802 | Song Hinh, Phu Yen, Vietnam | MZ712232 | MZ712258 |
| Pareas macularius | ZMMU R-16629 | Ban Mauk, Sagaing, Myanmar | MT968771 | MW287057 |
| Pareas margaritophorus | ZMMU NAP-09759 | Suan Phueng, Ratchaburi, Thailand | MZ712217 | MZ712243 |
| Pareas modestus | MZMU 1293 | Aizawl, Mizoram, India | MT968773 | — |
| Pareas monticola | ZMMU R-16631 | Ban Mauk, Sagaing, Myanmar | MW438296 | MW438301 |
| Pareas niger | KIZ 059339 | Kunming, Yunnan, China | MW436706 | — |
| Pareas niger | GP1294 | Mengzi, Yunnan, China | MK135079 | MK805344 |
| Pareas niger | YBU 14251 | Mengzi, Yunnan, China | MK135080 | MK805345 |
| Pareas niger | YBU 14252 | Mengzi, Yunnan, China | MK135081 | MK805346 |
| Pareas niger | YBU 14253 | Mengzi, Yunnan, China | MK135082 | MK805347 |
| Pareas niger | YBU 14288 | Mengzi, Yunnan, China | MK135083 | MK805348 |
| Pareas niger | YBU 15100 | Kaiyuan, Yunnan, China | MK135084 | MK805349 |
| Pareas niger | YBU 15114 | Kaiyuan, Yunnan, China | MK135085 | MK805350 |
| Pareas nigriceps | CHS656 | Tengchong, Yunnan, China | MK201455 | — |
| Pareas nuchalis | FK 2626 | Belait, Brunei Darussalam, Brunei | MZ603794 | U49311 |
| Pareas stanleyi | HM 2007-S001 | Guilin, Guangxi, China | JN230704 | JN230705 |
| Pareas temporalis | ZMMU R-13656 | Cat Loc, Lam Dong, Vietnam | MZ712238 | MZ712265 |
| Pareas tigerinus | KIZ 20210703 | Menghai, Yunnan, China | OP752143 | — |
| Pareas victorianus | CAS 235254 | Natmataung, Chin, Myanmar | MW438300 | MW438302 |
| Pareas vindumi | CAS 248147 | Lukpwi, Kachin, Myanmar | MW287080 | MW287059 |
| Pareas xuelinensis | KIZ XL1 | Lancang, Yunnan, China | MW436709 | — |
| Pareas xuelinensis | KIZ XL2 | Lancang, Yunnan, China | MW436710 | — |
| Pareas yunnanensis | KIZ 2022033 | Dali, Yunnan, China | OP752146 | — |
| Pareas yunnanensis | KIZ 2022034 | Dali, Yunnan, China | OP752147 | — |
| Pareas yunnanensis | KIZ 2022035 | Dali, Yunnan, China | OP752148 | — |
| Pareas yunnanensis | KIZ 2022036 | Dali, Yunnan, China | OP752149 | — |
| Aplopeltura boa | LSUHC 7248 | Sepilok, Sabah, Malaysia | KC916746 | U49312 |
| KIZ 2023038 Holotype ♀ | KIZ 2023039 Paratype ♀ | KIZ 2023040 Paratype ♀ | |
|---|---|---|---|
| SVL | 488 | 482 | 540 |
| TL | 146 | 126 | 152 |
| PrFBO | Yes | Yes | Yes |
| PreO | 1/1 | 1/1 | 1/1 |
| PosO | 1/1 | Fused/1 | 1/Fused |
| SubO | 1/1 | Fused/1 | 1/Fused |
| SPOF | No/No | Yes/No | No/Yes |
| ATem | 2/2 | 2/2 | 2/2 |
| PTem | 3/3 | 4/4 | 2/2 |
| SupL | 7/7 | 8/8 | 7/7 |
| InfL | 7/8 | 7/6 | 7/7 |
| LoBO | No | No | No |
| Vs | 190 | 192 | 189 |
| Prec | Undivided | Undivided | Undivided |
| Sc | 89 | 72 | 80 |
| DS | 15-15-15 | 15-15-15 | 15-15-15 |
| NED | 1 | 1 | 1 |
| NKD | 0-5-5 | 0-5-7 | 0-5-5 |
| Max | 5/4 | 5/5 | 5/5 |
| KIZ 20210517 ♂ | KIZ L2020018 Juvenile | KIZ 201903001 ♀ | KIZ 20210518 Juvenile | KIZ 20210519 ♂ | KIZ 2023046 ♂ | |
|---|---|---|---|---|---|---|
| SVL | 458 | 276 | 360 | 306 | 459 | 532 |
| TL | 157 | 91 | 114 | 97 | 151 | Incomplete |
| PrFBO | Yes | Yes | Yes | Yes | Yes | Yes |
| PreO | 1/1 | 1/1 | 1/1 | 1/1 | 1/1 | 1/1 |
| PosO | Fused/Fused | Fused/Fused | Fused/Fused | 1/1 | 1/1 | 1/1 |
| SubO | Fused/Fused | Fused/Fused | Fused/Fused | 1/1 | 1/1 | 1/1 |
| SPOF | Yes/yes | Yes/yes | Yes/yes | No/No | No/No | No/No |
| ATem | 2/2 | 2/2 | 1/1 | 2/2 | 3/3 | 2/2 |
| PTem | 3/2 | 3/3 | 3/2 | 2/3 | 3/3 | 3/3 |
| SupL | 7/8 | 7/7 | 7/7 | 7/7 | 8/7 | 7/7 |
| InfL | 6/6 | 7/7 | 7/7 | 7/7 | 8/8 | 7/7 |
| LoBO | No | No | No | No | No | No |
| Vs | 188 | 188 | 187 | 193 | 191 | 186 |
| Prec | Undivided | Undivided | Undivided | Undivided | Undivided | Undivided |
| Sc | 92 | 97 | 94 | 95 | 91 | Incomplete |
| DS | 15-15-15 | 15-15-15 | 15-15-15 | 15-15-15 | 15-15-15 | 15-15-15 |
| NED | 1 | 1 | 1 | 1 | 1 | 1 |
| NKD | 0-3-5 | 0-3-3 | 0-3-5 | 0-3-3 | 0-3-5 | 0-5-7 |
| Max | 4/5 | 5/5 | 5/5 | 4/4 | 5/5 | 4/5 |
| (1) | (2) | (3) | (4) | (5) | (6) | (7) | (8) | (9) | (10) | (11) | (12) | (13) | (14) | (15) | (16) | (17) | (18) | (19) | (20) | (21) | (22) | (23) | (24) | (25) | (26) | (27) | (28) | (29) | (30) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (1) | ||||||||||||||||||||||||||||||
| (2) | 23.5 | |||||||||||||||||||||||||||||
| (3) | 21.5 | 23.5 | ||||||||||||||||||||||||||||
| (4) | 13.6 | 22.8 | 20.3 | |||||||||||||||||||||||||||
| (5) | 19.2 | 23.7 | 20.6 | 20.6 | ||||||||||||||||||||||||||
| (6) | 23.1 | 21.0 | 23.8 | 23.5 | 22.5 | |||||||||||||||||||||||||
| (7) | 17.2 | 23.2 | 19.7 | 18.4 | 14.2 | 23.6 | ||||||||||||||||||||||||
| (8) | 23.1 | 21.8 | 22.9 | 23.2 | 22.4 | 13.9 | 22.6 | |||||||||||||||||||||||
| (9) | 18.0 | 23.8 | 19.2 | 18.8 | 14.6 | 24.9 | 9.0 | 22.8 | ||||||||||||||||||||||
| (10) | 12.8 | 24.0 | 19.6 | 13.9 | 19.6 | 24.2 | 18.0 | 23.7 | 17.8 | |||||||||||||||||||||
| (11) | 8.0 | 23.9 | 21.6 | 15.0 | 19.2 | 24.3 | 17.1 | 23.9 | 17.8 | 13.4 | ||||||||||||||||||||
| (12) | 7.5 | 22.9 | 22.2 | 14.1 | 19.0 | 22.9 | 17.4 | 23.2 | 19.0 | 12.8 | 9.5 | |||||||||||||||||||
| (13) | 4.2 | 23.5 | 21.7 | 14.0 | 18.9 | 23.3 | 17.2 | 23.5 | 18.3 | 12.6 | 8.0 | 7.4 | ||||||||||||||||||
| (14) | 13.1 | 23.4 | 20.5 | 7.0 | 19.3 | 24.4 | 17.4 | 24.3 | 18.3 | 14.0 | 14.5 | 14.5 | 13.7 | |||||||||||||||||
| (15) | 12.3 | 24.5 | 20.2 | 14.7 | 19.8 | 24.5 | 19.4 | 22.2 | 18.8 | 9.1 | 13.4 | 13.4 | 12.7 | 14.8 | ||||||||||||||||
| (16) | 14.3 | 23.2 | 19.5 | 8.6 | 19.5 | 23.9 | 18.0 | 23.9 | 18.2 | 13.7 | 14.7 | 14.9 | 14.4 | 8.1 | 15.5 | |||||||||||||||
| (17) | 22.7 | 21.2 | 23.8 | 23.2 | 23.3 | 13.2 | 22.6 | 13.2 | 23.0 | 24.2 | 23.3 | 23.1 | 23.2 | 24.4 | 22.9 | 24.3 | ||||||||||||||
| (18) | 18.7 | 23.9 | 14.5 | 18.9 | 18.5 | 22.7 | 18.4 | 21.7 | 17.6 | 18.3 | 19.8 | 20.3 | 18.9 | 19.5 | 19.6 | 19.2 | 22.6 | |||||||||||||
| (19) | 19.9 | 24.7 | 16.2 | 18.9 | 20.0 | 24.2 | 19.2 | 22.8 | 18.7 | 17.9 | 19.6 | 20.5 | 19.4 | 19.0 | 20.1 | 18.4 | 23.1 | 14.9 | ||||||||||||
| (20) | 20.4 | 23.5 | 12.0 | 18.4 | 19.1 | 24.2 | 19.2 | 24.1 | 18.8 | 18.6 | 20.3 | 20.1 | 19.7 | 19.8 | 19.2 | 17.9 | 24.4 | 12.0 | 14.8 | |||||||||||
| (21) | 18.6 | 22.7 | 18.7 | 17.7 | 20.3 | 21.7 | 18.3 | 22.9 | 18.5 | 17.6 | 19.5 | 19.5 | 18.6 | 17.7 | 18.7 | 18.1 | 21.9 | 17.1 | 18.9 | 18.7 | ||||||||||
| (22) | 5.7 | 22.6 | 20.6 | 14.1 | 19.0 | 23.5 | 17.6 | 23.3 | 17.9 | 12.2 | 8.3 | 6.9 | 6.0 | 13.6 | 12.1 | 14.6 | 22.7 | 18.6 | 19.9 | 19.0 | 18.9 | |||||||||
| (23) | 12.4 | 23.6 | 18.8 | 15.7 | 16.9 | 22.9 | 16.9 | 22.9 | 16.2 | 10.3 | 13.3 | 13.5 | 12.6 | 16.1 | 9.7 | 16.2 | 23.9 | 19.3 | 17.8 | 16.4 | 18.8 | 12.6 | ||||||||
| (24) | 25.0 | 21.1 | 24.3 | 23.4 | 23.4 | 21.2 | 24.3 | 21.2 | 24.1 | 24.6 | 24.6 | 25.1 | 24.8 | 24.3 | 25.0 | 23.5 | 20.4 | 23.3 | 24.6 | 24.5 | 22.5 | 25.2 | 23.8 | |||||||
| (25) | 19.2 | 25.7 | 20.4 | 19.2 | 16.3 | 25.2 | 15.7 | 24.7 | 15.4 | 19.2 | 19.5 | 19.5 | 18.7 | 18.9 | 20.3 | 17.3 | 24.9 | 18.6 | 19.5 | 19.4 | 18.8 | 19.6 | 19.0 | 24.0 | ||||||
| (26) | 23.8 | 13.3 | 23.8 | 23.6 | 23.3 | 20.6 | 22.5 | 20.1 | 21.9 | 25.1 | 24.6 | 23.9 | 23.7 | 23.6 | 24.8 | 24.2 | 20.1 | 23.4 | 24.8 | 23.6 | 23.0 | 23.6 | 24.1 | 20.2 | 24.1 | |||||
| (27) | 12.1 | 23.0 | 19.3 | 14.3 | 19.7 | 24.1 | 19.0 | 23.3 | 18.7 | 10.3 | 12.9 | 12.4 | 11.8 | 14.2 | 10.7 | 14.0 | 24.3 | 18.6 | 19.5 | 18.2 | 18.7 | 11.4 | 4.3 | 25.0 | 19.3 | 24.8 | ||||
| (28) | 19.1 | 24.3 | 20.6 | 19.5 | 19.2 | 22.8 | 19.1 | 22.9 | 17.5 | 18.0 | 18.1 | 18.6 | 18.6 | 19.9 | 18.4 | 19.3 | 22.9 | 19.0 | 20.6 | 19.3 | 14.3 | 17.8 | 19.1 | 24.7 | 19.0 | 24.5 | 17.9 | |||
| (29) | 11.0 | 24.4 | 20.8 | 14.7 | 19.5 | 24.4 | 18.4 | 24.2 | 17.7 | 12.6 | 12.5 | 12.4 | 11.5 | 14.5 | 12.3 | 15.0 | 23.8 | 19.0 | 19.9 | 19.9 | 17.9 | 11.0 | 12.3 | 24.7 | 19.4 | 25.3 | 11.9 | 17.8 | ||
| (30) | 8.3 | 23.1 | 21.3 | 13.8 | 20.5 | 24.7 | 16.9 | 24.3 | 18.7 | 13.3 | 8.9 | 6.0 | 8.3 | 13.9 | 13.2 | 14.8 | 24.6 | 19.7 | 20.0 | 20.2 | 19.6 | 7.3 | 12.5 | 25.9 | 19.5 | 24.7 | 12.3 | 18.8 | 12.6 | |
| (31) | 7.1 | 23.2 | 22.1 | 14.4 | 19.1 | 24.1 | 16.7 | 23.5 | 18.2 | 12.9 | 8.6 | 4.1 | 6.2 | 13.9 | 12.4 | 14.7 | 23.7 | 20.1 | 20.1 | 20.6 | 19.6 | 6.4 | 12.8 | 24.9 | 19.5 | 23.9 | 11.7 | 18.7 | 11.5 | 6.2 |
| (1) | (2) | (3) | (4) | (5) | (6) | (7) | (8) | (9) | (10) | (11) | (12) | (13) | (14) | (15) | (16) | (17) | (18) | (19) | (20) | (21) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (1) | |||||||||||||||||||||
| (2) | 18.6 | ||||||||||||||||||||
| (3) | 19.0 | 20.4 | |||||||||||||||||||
| (4) | 16.7 | 22.3 | 19.1 | ||||||||||||||||||
| (5) | 18.1 | 21.8 | 18.0 | 20.2 | |||||||||||||||||
| (6) | 21.0 | 19.5 | 21.5 | 22.0 | 25.5 | ||||||||||||||||
| (7) | 18.7 | 21.1 | 17.8 | 18.4 | 12.1 | 22.8 | |||||||||||||||
| (8) | 21.5 | 18.9 | 23.0 | 21.1 | 22.4 | 14.3 | 21.0 | ||||||||||||||
| (9) | 18.3 | 21.1 | 18.3 | 20.1 | 12.2 | 22.8 | 10.1 | 22.6 | |||||||||||||
| (10) | 9.1 | 18.8 | 19.6 | 16.2 | 17.1 | 20.7 | 19.3 | 21.3 | 17.5 | ||||||||||||
| (11) | 5.0 | 19.4 | 19.0 | 16.4 | 17.5 | 21.2 | 17.9 | 20.9 | 16.5 | 8.7 | |||||||||||
| (12) | 17.8 | 21.9 | 19.7 | 7.5 | 20.8 | 22.3 | 19.0 | 21.6 | 19.2 | 16.2 | 17.5 | ||||||||||
| (13) | 19.1 | 19.9 | 21.3 | 21.1 | 22.8 | 14.1 | 19.8 | 15.0 | 21.7 | 19.7 | 18.9 | 21.3 | |||||||||
| (14) | 20.3 | 22.5 | 11.8 | 19.8 | 18.6 | 22.9 | 19.5 | 23.2 | 20.2 | 20.9 | 20.4 | 20.8 | 23.7 | ||||||||
| (15) | 19.9 | 20.8 | 14.0 | 19.6 | 18.9 | 21.3 | 18.6 | 21.0 | 20.8 | 19.6 | 20.1 | 21.0 | 20.2 | 14.7 | |||||||
| (16) | 18.7 | 21.7 | 19.6 | 18.9 | 19.3 | 21.6 | 18.2 | 20.7 | 18.4 | 20.0 | 18.5 | 20.5 | 19.5 | 22.0 | 21.4 | ||||||
| (17) | 9.5 | 19.8 | 18.8 | 16.0 | 18.0 | 22.2 | 17.6 | 21.4 | 17.1 | 8.8 | 9.0 | 16.2 | 21.3 | 20.4 | 18.6 | 19.7 | |||||
| (18) | 21.3 | 17.4 | 19.9 | 23.4 | 22.8 | 18.2 | 22.3 | 18.8 | 22.5 | 20.3 | 21.1 | 23.7 | 19.5 | 23.4 | 21.1 | 21.3 | 21.4 | ||||
| (19) | 20.3 | 21.6 | 18.0 | 19.5 | 16.2 | 23.9 | 14.8 | 22.5 | 16.6 | 19.3 | 20.4 | 18.6 | 22.4 | 18.0 | 19.5 | 19.2 | 19.1 | 23.0 | |||
| (20) | 19.0 | 9.4 | 20.0 | 19.9 | 20.8 | 19.3 | 20.2 | 19.2 | 18.7 | 18.9 | 19.4 | 19.6 | 18.9 | 21.9 | 19.6 | 19.5 | 18.0 | 17.4 | 20.1 | ||
| (21) | 17.6 | 20.4 | 18.5 | 18.4 | 18.4 | 22.9 | 18.3 | 20.8 | 18.6 | 18.0 | 17.4 | 19.0 | 21.7 | 20.5 | 20.2 | 12.8 | 18.6 | 23.5 | 19.5 | 18.3 | |
| (22) | 12.5 | 19.0 | 16.3 | 13.9 | 14.8 | 21.1 | 15.6 | 20.4 | 15.7 | 12.8 | 13.0 | 14.2 | 19.2 | 18.0 | 18.6 | 17.4 | 13.6 | 20.1 | 16.5 | 18.5 | 15.6 |
| Pareas guanyinshanensis sp. nov. | Pareas hamptoni Sensu Stricto | |
|---|---|---|
| SVL (adult) | 482–540 | 360–532 |
| TL (adult) | 126–152 | 114–157 |
| TL/SVL | 0.26–0.30 | 0.32–0.37 |
| PrFBO | Yes | Yes |
| PreO | 1 | 1 |
| PosO | Fused or 1 | Fused or 1 |
| SubO | Fused or 1 | Fused or 1 |
| SPOF | Yes or No | Yes or No |
| ATem | 2 | 1–3 |
| PTem | 2–4 | 2–3 |
| SupL | 7–8 | 7–8 |
| InfL | 6–8 | 6–9 |
| LoBO | No | No |
| Vs | 189–192 | 185–195 |
| Prec | Undivided | Undivided |
| Sc | 72–89 | 91–99 |
| DS | 15-15-15 | 15-15-15 |
| NED | 1 | 1 |
| NKD | 0–5 | 0–9 |
| Max | 4–5 | 4–5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Mo, M.; Li, M.; Li, B.; Luo, X.; Rao, D.; Li, S. Description of a New Species of the Pareas hamptoni Complex from Yunnan, China, with Confirmation of P. hamptoni Sensu Stricto in China (Squamata, Pareidae). Animals 2024, 14, 421. https://doi.org/10.3390/ani14030421
Liu S, Mo M, Li M, Li B, Luo X, Rao D, Li S. Description of a New Species of the Pareas hamptoni Complex from Yunnan, China, with Confirmation of P. hamptoni Sensu Stricto in China (Squamata, Pareidae). Animals. 2024; 14(3):421. https://doi.org/10.3390/ani14030421
Chicago/Turabian StyleLiu, Shuo, Mingzhong Mo, Mei Li, Biao Li, Xiong Luo, Dingqi Rao, and Song Li. 2024. "Description of a New Species of the Pareas hamptoni Complex from Yunnan, China, with Confirmation of P. hamptoni Sensu Stricto in China (Squamata, Pareidae)" Animals 14, no. 3: 421. https://doi.org/10.3390/ani14030421
APA StyleLiu, S., Mo, M., Li, M., Li, B., Luo, X., Rao, D., & Li, S. (2024). Description of a New Species of the Pareas hamptoni Complex from Yunnan, China, with Confirmation of P. hamptoni Sensu Stricto in China (Squamata, Pareidae). Animals, 14(3), 421. https://doi.org/10.3390/ani14030421