
Short- and Long-Term Effects of Split-Suckling in Pigs According to Birth Weight

 , , , and
, , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Farrowing, Split-Suckling Treatment, and Lactation
- At the end of farrowing (when the sow expelled the placenta), normal-BW piglets were separated for 1 h to allow access for the low-BW piglets. Piglets removed from the litter were calculated according to the number of functional teats.
- After the first hour, the piglets that had been removed in step 1 were reintroduced among the piglets remaining with the sow, and another set of heavy piglets different from the first selection was removed for 1 h.
- After the second hour, the procedure was repeated by removing those piglets that were selected the first time (step 1).
- Finally, after the third hour, the last piglets removed were reintroduced so that the litter was complete for the rest of the lactation.
2.3. Weaning, Growing, and Fattening
2.4. Carcass and Meat Quality
2.5. Statistical Analysis
3. Results
3.1. Mortality
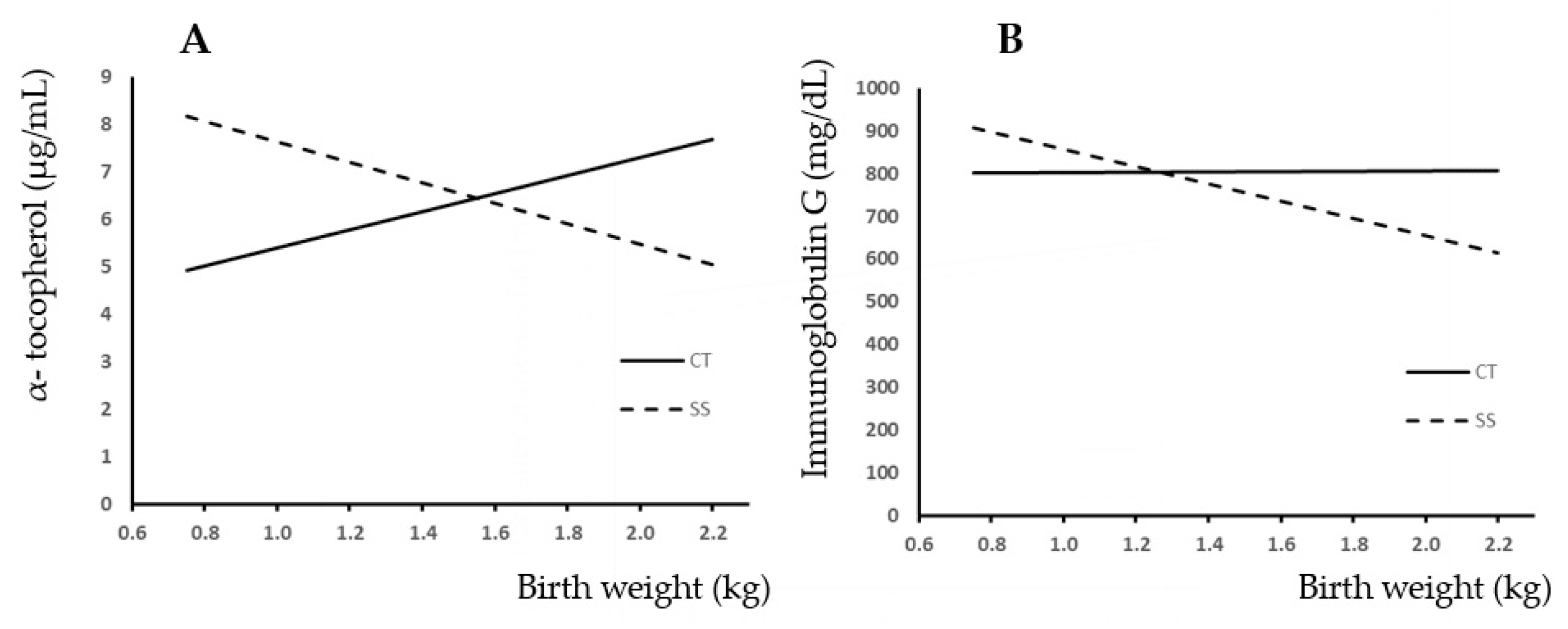
3.2. Colostrum Intake
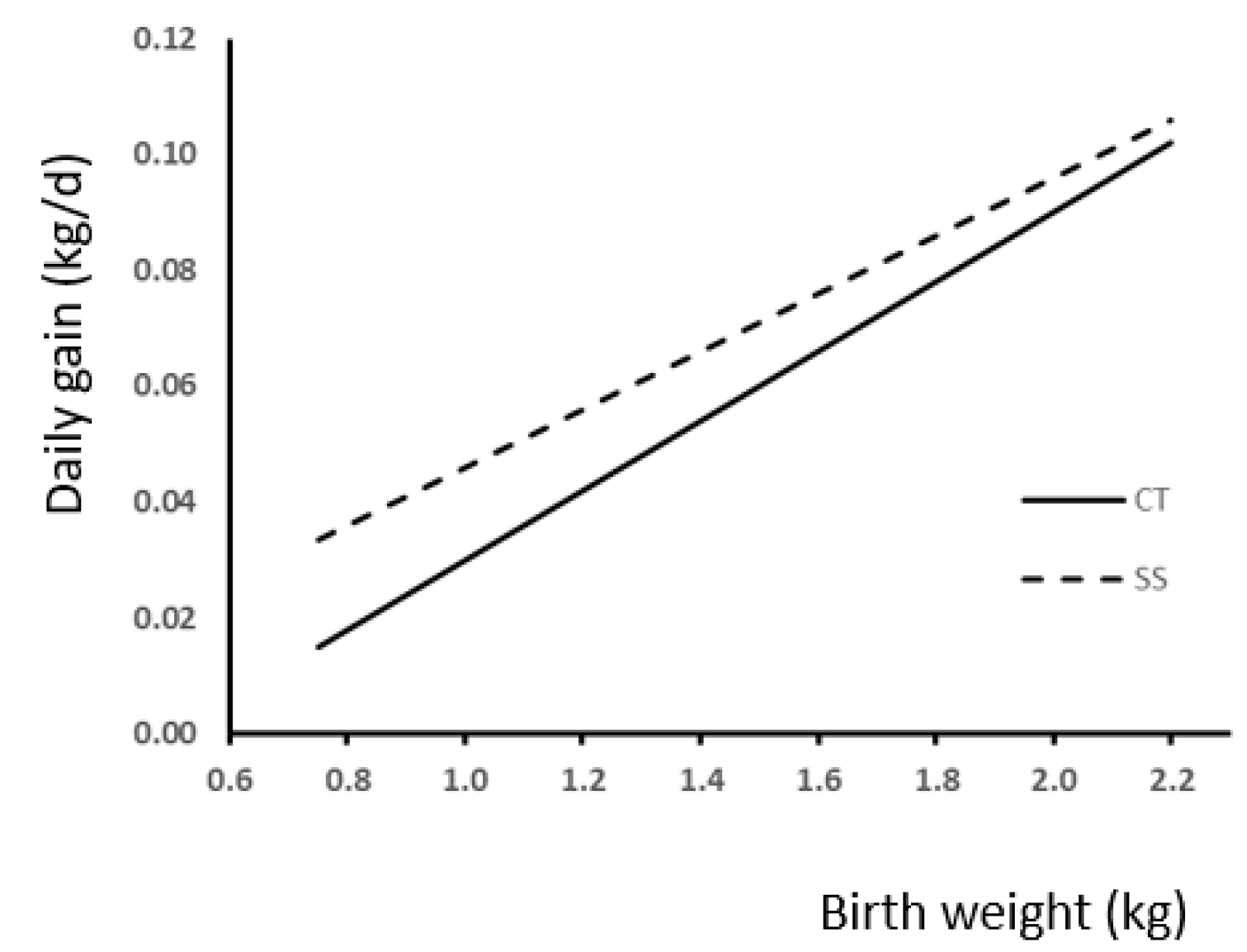
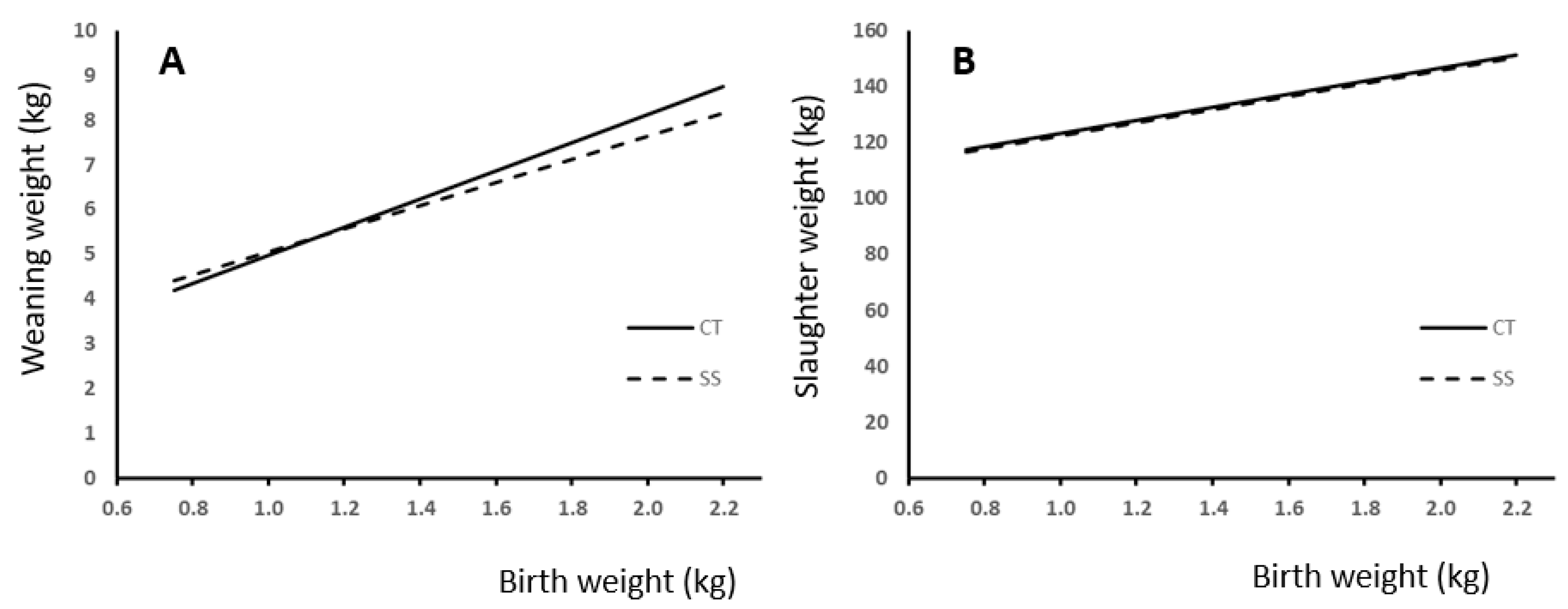
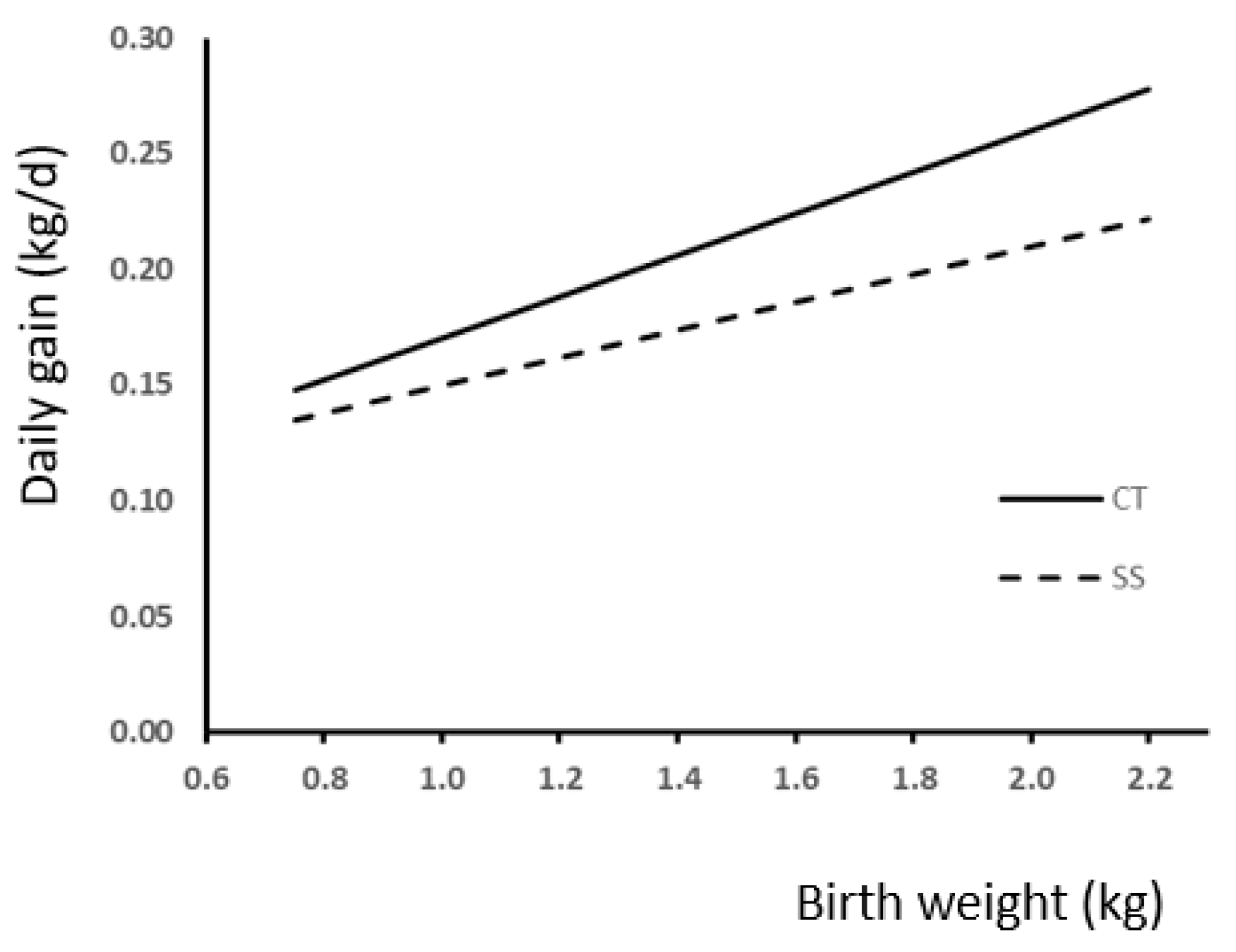
3.3. Weight and Gains
3.4. Carcass and Meat Composition and Quality Characteristics
4. Discussion
4.1. Mortality
4.2. Colostrum Intake
4.3. Weight and Gains
4.4. Carcass and Meat Composition and Quality Characteristics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hansen, C.F.; Hales, J.; Amdi, C.; Moustsen, V.A. Intrauterine growth-restricted piglets defined by their head shape have impaired survival and growth during the suckling period. Anim. Prod. Sci. 2019, 59, 1056–1062. [Google Scholar] [CrossRef]
- Kemp, B.; Da Silva, C.L.; Soede, N.M. Recent advances in pig reproduction: Focus on impact of genetic selection for female fertility. Reprod. Domest. Anim. 2018, 53, 28–36. [Google Scholar] [CrossRef]
- Rutherford, K.M.D.; Baxter, E.M.; D’Eath, R.B.; Turner, S.P.; Arnott, G.; Roehe, R.; Ask, B.; Sandoe, P.; Moustsen, V.A.; Thorup, F.; et al. The welfare implications of large litter size in the domestic pig I: Biological factors. Anim. Welf. 2013, 22, 199–218. [Google Scholar] [CrossRef]
- Le Dividich, J.; Rooke, J.A.; Herpin, P. Nutritional and immunological importance of colostrum for the new-born pig. J. Agric. Sci. 2005, 143, 469–485. [Google Scholar] [CrossRef]
- Devillers, N.; van Milgen, J.; Prunier, A.; Le Dividich, J. Estimation of colostrum intake in the neonatal pig. Anim. Sci. 2004, 78, 305–313. [Google Scholar] [CrossRef]
- Bartol, F.F.; Wiley, A.A.; Miller, D.J.; Silva, A.J.; Roberts, K.E.; Davolt, M.L.P.; Chen, J.C.; Frankshun, A.L.; Camp, M.E.; Rahman, K.M.; et al. Lactation biology symposium: Lactocrine signaling and developmental programming. J. Anim. Sci. 2013, 91, 696–705. [Google Scholar] [CrossRef]
- Morton, J.M.; Langemeier, A.J.; Rathbun, T.J.; Davis, D.L. Immunocrit, colostrum intake, and preweaning body weight gain in piglets after split suckling based on birth weight or birth order. Transl. Anim. Sci. 2019, 3, 1460–1465. [Google Scholar] [CrossRef] [PubMed]
- Michiels, J.; Vandenberghe, H.; Vangansbeke, S.; Degroote, J.; Fremaut, D.; Ampe, B. Handling of supernumerary piglets in flanders, a survey among pig breeders. In Proceedings of the 64th Annual Meeting of the European Federation of Animal Science (EAAP), Nantes, France, 20 August 2013; Wageningen Academic Publishers: Wageningen, The Netherlands, 2013; Volume 19, p. 399. [Google Scholar]
- Vandaele, M.; Van Kerschaver, C.; Degroote, J.; Van Ginneken, C.; Michiels, J. Piglet performance and colostrum intake in litters either or not split-suckled during the first day or during the first three days of life. Livest. Sci. 2020, 241, 104265. [Google Scholar] [CrossRef]
- Kyriazakis, I.; Edwards, S.A. The effect of split-suckling on behavior and performance of piglets. Appl. Anim. Behav. Sci. 1986, 16, 92. [Google Scholar] [CrossRef]
- Alonso, V.; Rocco, V.V.B.; Galdeano, J.V.B.; Santana, R.S.; Molero, A.H.T.; de Oliveira, M.C. Split suckling in relation to the parturition order of sows. Rev. Bras. De Zootec.-Braz. J. Anim. Sci. 2012, 41, 221–224. [Google Scholar] [CrossRef]
- Rosvold, E.M.; Kielland, C.; Ocepek, M.; Framstad, T.; Fredriksen, B.; Andersen-Ranberg, I.; Naess, G.; Andersen, I.L. Management routines influencing piglet survival in loose-housed sow herds. Livest. Sci. 2017, 196, 1–6. [Google Scholar] [CrossRef][Green Version]
- Huser, J.; Plush, K.; Pitchford, W.; Kennett, T.; Lines, D. Neonatal split suckling improves survival of small piglets. Anim. Prod. Sci. 2015, 55, 1477. [Google Scholar] [CrossRef]
- De Blas, C.; Gasa, J.; Mateos, G. Necesidades Nutricionales para Ganado Porcino; Fundacion Española para el Desarrollo de la Nutrición Animal: Madrid, Spain, 2006. [Google Scholar]
- Branscheid, W.; Dobrowolski, A.; Sack, E. Simplification of the ec-reference method for the full dissection of pig carcasses. Fleischwirtschaft 1990, 70, 550–553. [Google Scholar]
- Segura, J.; Lopez-Bote, C.J. A laboratory efficient method for intramuscular fat analysis. Food Chem. 2014, 145, 821–825. [Google Scholar] [CrossRef] [PubMed]
- Rey, A.I.; Daza, A.; Lopez-Carrasci, C.; Lopez-Bote, C.J. Quantitative study of the alpha- and gamma-tocopherols accumulation in muscle and backfat from Iberian pigs kept free-range as affected by time of free-range feeding or weight gain. Anim. Sci. 2006, 82, 901–908. [Google Scholar] [CrossRef]
- Muns, R.; Nuntapaitoon, M.; Tummaruk, P. Non-infectious causes of pre-weaning mortality in piglets. Livest. Sci. 2016, 184, 46–57. [Google Scholar] [CrossRef]
- Vanden Hole, C.; Ayuso, M.; Aerts, P.; Prims, S.; Van Cruchten, S.; Van Ginneken, C. Glucose and glycogen levels in piglets that differ in birth weight and vitality. Heliyon 2019, 5, e02510. [Google Scholar] [CrossRef]
- Rekiel, A.; Wiecek, J.; Batorska, M.; Kulisiewicz, J. Effect of piglet birth weight on carcass muscle and fat content and pork quality-a review. Ann. Anim. Sci. 2015, 15, 271–287. [Google Scholar] [CrossRef]
- Donovan, T.; Dritz, S.S. Effects of split-nursing management on growth performance in nursing pigs. In Kansas Agricultural Experiment Station Research Reports; Kansas State University: Manhattan, KS, USA, 1996; Volume 0. [Google Scholar] [CrossRef]
- Holyoake, P.K.; Dial, G.D.; Trigg, T.; King, V.L. Reducing pig mortality through supervision during the perinatal period. J. Anim. Sci. 1995, 73, 3543–3551. [Google Scholar] [CrossRef]
- Cabrera, R.A.; Boyd, R.D.; Jungst, S.B.; Wilson, E.R.; Johnston, M.E.; Vignes, J.L.; Odle, J. Impact of lactation length and piglet weaning weight on long-term growth and viability of progeny. J. Anim. Sci. 2010, 88, 2265–2276. [Google Scholar] [CrossRef]
- Jensen, P.T.; Pedersen, K. Studies on immunoglobulins and trypsin inhibitor in colostrum and milk from sows and in serum of their piglets. Acta Vet. Scand. 1979, 20, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Hidiroglou, M.; Batra, T.; Farnworth, E.; Markham, F. Effect of vitamin E supplementation on immune status and α-tocopherol in plasma of piglets. Reprod. Nutr. Dev. 1995, 35, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Surek, D.; Barrilli, L.; Bueno, I.; Krabbe, E.; Alberton, G.; Maiorka, A. Growth of suckling piglets in litters standardized by weight. J. Anim. Sci. 2014, 92, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, A.D.; Aalhus, J.L.; Williams, N.H.; Patience, J.F. Impact of piglet birth weight, birth order, and litter size on subsequent growth performance, carcass quality, muscle composition, and eating quality of pork. J. Anim. Sci. 2010, 88, 2767–2778. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.; Calvo, L.; Morales, J.I.; Rodriguez, A.I.; Escudero, R.M.; Olivares, A.; Lopez-Bote, C. Short- and long-term effects of birth weight and neonatal care in pigs. Animals 2022, 12, 2936. [Google Scholar] [CrossRef]
- Gondret, F.; Lefaucheur, L.; Juin, H.; Louveau, I.; Lebret, B. Low birth weight is associated with enlarged muscle fiber area and impaired meat tenderness of the longissimus muscle in pigs. J. Anim. Sci. 2006, 84, 93–103. [Google Scholar] [CrossRef]
- Arnaud, E.A.; Gardiner, G.E.; Halpin, K.M.; Ribas, C.; O’Doherty, J.V.; Sweeney, T.; Lawlor, P.G. Post-partum meloxicam administration to sows but not split-suckling increases piglet growth and reduces clinical incidence of disease in suckling piglets. J. Anim. Sci. 2023, 101, skad275. [Google Scholar] [CrossRef]
- Latorre, M.A.; Olivares, A.; Callejo, A.; Rey, A.I.; Perez-Ciria, L.; Lopez Bote, C.J.; Daza, A. A comparison of female and castrate pigs slaughtered at weights above and below 120 kg on carcass traits, intramuscular fat and fatty acid composition of carcasses intended for dry-cured ham and shoulder production. Anim. Prod. Sci. 2019, 59, 1923–1930. [Google Scholar] [CrossRef]
- Rehfeldt, C.; Kuhn, G. Consequences of birth weight for postnatal growth performance and carcass quality in pigs as related to myogenesis. J. Anim. Sci. 2006, 84 (Suppl. S13), E113–E123. [Google Scholar] [CrossRef]
- George, A.D.; Burugupalli, S.; Paul, S.; Mansell, T.; Burgner, D.; Meikle, P.J. The role of human milk lipids and lipid metabolites in protecting the infant against non-communicable disease. Int. J. Mol. Sci. 2022, 23, 7490. [Google Scholar] [CrossRef]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Estany, J.; Ros-Freixedes, R.; Tor, M.; Pena, R.N. Triennial growth and development symposium: Genetics and breeding for intramuscular fat and oleic acid content in pigs. J. Anim. Sci. 2017, 95, 2261–2271. [Google Scholar] [CrossRef] [PubMed]
- Isabel, B.; Lopez-Bote, C.J.; de la Hoz, L.; Timon, M.; Garcia, C.; Ruiz, J. Effects of feeding elevated concentrations of monounsaturated fatty acids and vitamin E to swine on characteristics of dry cured hams. Meat Sci. 2003, 64, 475–482. [Google Scholar] [CrossRef]
- Kim, J.M.; Lee, S.N.; Ryu, Y.C. Comparisons of meat quality and muscle fibre characteristics on multiple pig breeds and sexes using principal component analysis. Anim. Prod. Sci. 2018, 58, 2091–2099. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Birth Weight (BW) | Treatment (T) | Parity (P) | Sex (S) | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Low | Normal | CT | SS | PP | MP | M | F | BW | T | P | S | |
| 6d | 0.304 | 0.047 | 0.08 | 0.086 | 0.067 | 0.090 | 0.075 | 0.092 | 0.0001 | 0.805 | 0.356 | 0.427 |
| Weaning | 0.554 | 0.157 | 0.171 | 0.25 | 0.247 | 0.201 | 0.212 | 0.215 | 0.0001 | 0.016 | 0.204 | 0.928 |
| T | P | S | SD | P < F | T | P | S | TxP | TxS | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CT | SS | PP | MP | CM | F | ||||||||
| Gain 0–24 h (kg) | 0.05 | 0.07 | 0.07 | 0.05 | 0.05 | 0.06 | 0.074 | 0.000 | 0.015 | 0.011 | 0.154 | 0.007 | 0.405 |
| Plasma α-toc (µg/mL) | 5.68 | 6.84 | 5.82 | 6.70 | 6.77 | 6.18 | 3.391 | 0.405 | 0.126 | 0.264 | 0.906 | 0.105 | 0.251 |
| Plasma IgG (mg/dL) | 811 | 790 | 825 | 776 | 812 | 780 | 371.4 | 0.503 | 0.763 | 0.673 | 0.941 | 0.687 | 0.457 |
| T | P | S | SD | P < F | T | P | S | TxP | TxS | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CT | SS | PP | MP | CM | F | ||||||||
| Weight (kg) | |||||||||||||
| Birth | 1.27 | 1.31 | 1.26 | 1.31 | 1.34 | 1.27 | 0.1103 | 0.0001 | 0.4841 | 0.4913 | 0.4361 | 0.4215 | 0.1435 |
| Weaning | 6.06 | 6.01 | 5.99 | 6.08 | 6.11 | 5.99 | 1.4191 | 0.0001 | 0.1817 | 0.6591 | 0.3587 | 0.501 | 0.9011 |
| Slaughter | 129 | 129 | 125 | 133 | 137 | 126 | 14.379 | 0.0001 | 0.4076 | 0.0119 | 0.0005 | 0.175 | 0.6027 |
| Daily gain (kg/d) | |||||||||||||
| Lactation | 0.20 | 0.18 | 0.20 | 0.19 | 0.19 | 0.19 | 0.0603 | 0.0001 | 0.0012 | 0.0866 | 0.2356 | 0.1072 | 0.8515 |
| Fattening | 0.74 | 0.74 | 0.71 | 0.76 | 0.78 | 0.72 | 0.0842 | 0.0001 | 0.5548 | 0.0059 | 0.0004 | 0.1654 | 0.5622 |
| T | P | S | SD | P < F | T | P | S | TxP | TxS | Cov | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CT | SS | PP | MP | CM | F | |||||||||
| Carcass | ||||||||||||||
| SC fat (P2, mm) | 19.2 | 17.9 | 18.5 | 18.5 | 19.7 | 17.33 | 3.12 | 0.0001 | 0.0028 | 0.9580 | 0.0001 | 0.0006 | 0.8558 | 0.0001 |
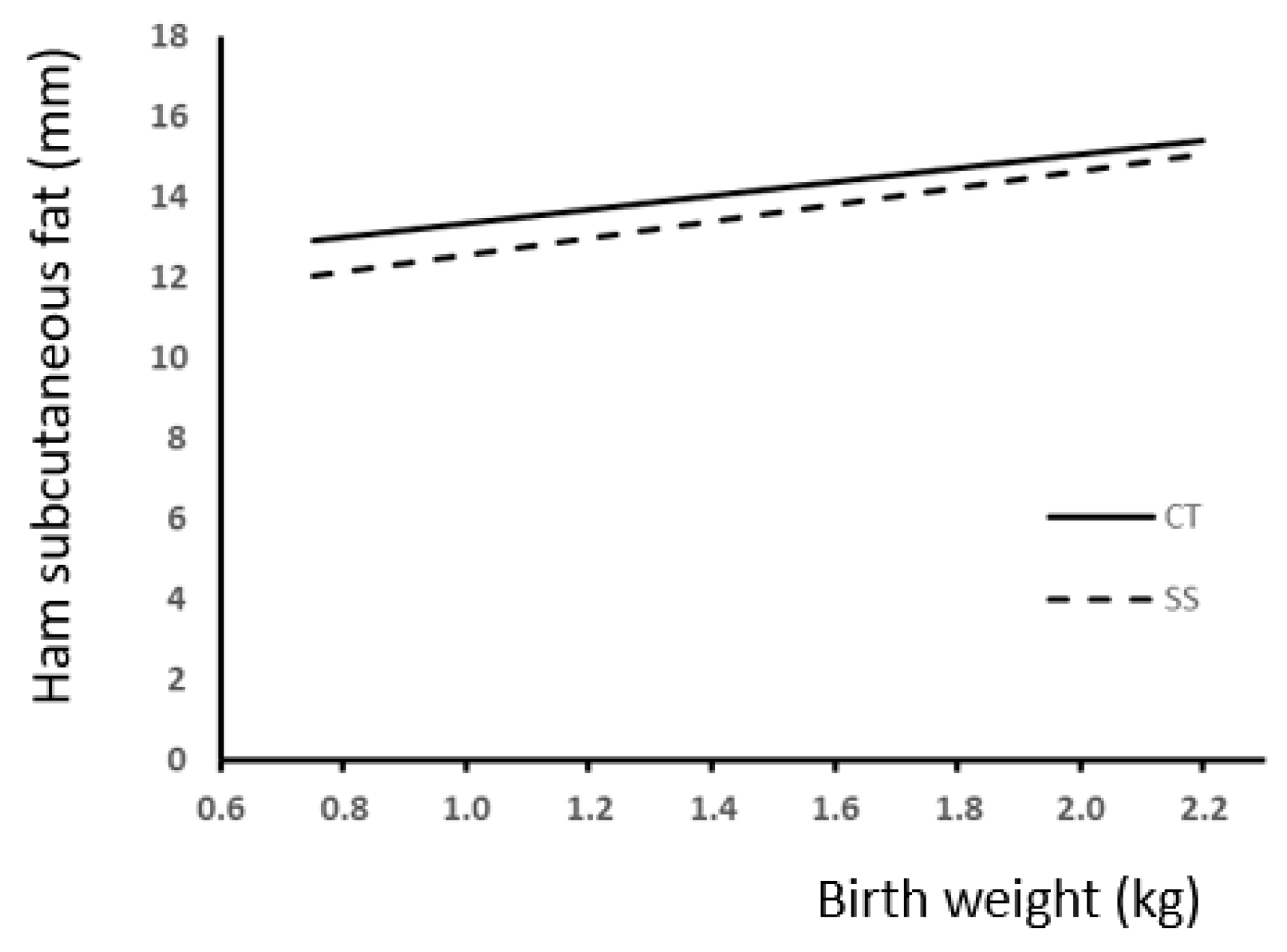
| SC fat (Ham, mm) | 14.2 | 13.3 | 13.7 | 13.8 | 14.625 | 12.85 | 2.19 | 0.0001 | 0.0013 | 0.6143 | 0.0001 | 0.0034 | 0.7065 | 0.0001 |
| % Lean | 57.8 | 58.8 | 58.3 | 58.3 | 57.15 | 59.43 | 2.73 | 0.0001 | 0.0027 | 0.8051 | 0.0001 | 0.0002 | 0.9764 | 0.0001 |
| Splitting yield (kg/kg carcass) | ||||||||||||||
| Loin | 3.18 | 3.17 | 3.16 | 3.19 | 3.115 | 3.238 | 0.22 | 0.0001 | 0.8986 | 0.3809 | 0.0011 | 0.5923 | 0.8051 | 0.0001 |
| Ham | 13.3 | 13.4 | 13.3 | 13.4 | 13.25 | 13.4 | 0.403 | 0.0001 | 0.4831 | 0.1644 | 0.0801 | 0.0009 | 0.6811 | 0.0001 |
| Ham muscles | 7.38 | 7.41 | 7.26 | 7.53 | 7.2225 | 7.565 | 0.409 | 0.0001 | 0.7653 | 0.0044 | 0.0004 | 0.0042 | 0.2905 | 0.0001 |
| BW | T | P | S | SD | P < F | BW | T | P | S | TxBW | TxP | TxS | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L | H | CT | SS | PP | MP | CM | F | ||||||||||
| pH and sacoplasmic protein solubility | |||||||||||||||||
| pH25 (loin) | 6.7 | 6.7 | 6.7 | 6.7 | 6.7 | 6.7 | 6.6 | 6.7 | 0.19 | 0.252 | 0.755 | 0.410 | 0.458 | 0.612 | 0.946 | 0.525 | 0.215 |
| pHu (loin) | 5.4 | 5.3 | 5.4 | 5.4 | 5.4 | 5.4 | 5.39 | 5.35 | 0.09 | 0.019 | 0.010 | 0.816 | 0.345 | 0.007 | 0.562 | 0.742 | 0.668 |
| Sarc. Pr.Sol. | 12.6 | 14.8 | 14.2 | 14.4 | 13.2 | 15.5 | 14.3 | 14.7 | 8.38 | 0.003 | 0.126 | 0.084 | 0.114 | 0.082 | 0.094 | 0.439 | 0.002 |
| Intramuscular fat | |||||||||||||||||
| LD | 2.9 | 2.6 | 2.9 | 2.6 | 2.8 | 2.8 | 3.0 | 2.5 | 0.84 | 0.078 | 0.055 | 0.622 | 0.712 | 0.028 | 0.200 | 0.646 | 0.591 |
| BF | 2.9 | 2.9 | 3.1 | 2.8 | 3.1 | 2.9 | 3.2 | 2.7 | 0.58 | 0.037 | 0.233 | 0.435 | 0.339 | 0.074 | 0.378 | 0.715 | 0.091 |
| SM | 3.9 | 3.6 | 4.1 | 3.7 | 4.0 | 3.7 | 4.2 | 3.6 | 0.92 | 0.045 | 0.139 | 0.494 | 0.217 | 0.264 | 0.521 | 0.737 | 0.056 |
| a value | |||||||||||||||||
| LD | 1.9 | 2.3 | 2.1 | 2.0 | 2.0 | 2.1 | 2.1 | 2.0 | 0.84 | 0.057 | 0.090 | 0.409 | 0.420 | 0.466 | 0.699 | 0.380 | 0.452 |
| BF | 1.9 | 13.9 | 14.1 | 13.8 | 13.8 | 14.0 | 13.9 | 13.9 | 1.23 | 0.524 | 0.677 | 0.169 | 0.800 | 0.726 | 0.552 | 0.281 | 0.566 |
| SM | 13.5 | 13.7 | 13.7 | 13.2 | 13.6 | 13.4 | 13.4 | 13.5 | 1.26 | 0.791 | 0.838 | 0.044 | 0.504 | 0.993 | 0.258 | 0.258 | 0.894 |
| b value | |||||||||||||||||
| LD | 13.1 | 13.3 | 13.2 | 13.1 | 13.2 | 13.1 | 13.1 | 13.1 | 1.01 | 0.014 | 0.058 | 0.134 | 0.141 | 0.950 | 0.059 | 0.177 | 0.886 |
| BF | 13.4 | 13.3 | 13.6 | 13.1 | 13.4 | 13.3 | 13.4 | 13.3 | 1.12 | 0.788 | 0.617 | 0.137 | 0.108 | 0.837 | 0.752 | 0.778 | 0.655 |
| SM | 14.1 | 14.0 | 14.4 | 13.6 | 14.1 | 14.0 | 14.0 | 14.1 | 0.99 | 0.231 | 0.662 | 0.086 | 0.118 | 0.788 | 0.902 | 0.196 | 0.557 |
| L value | |||||||||||||||||
| LD | 70.4 | 70.0 | 70.5 | 70.1 | 70.7 | 69.9 | 70.3 | 70.3 | 2.27 | 0.288 | 0.118 | 0.143 | 0.010 | 0.622 | 0.689 | 0.491 | 0.773 |
| BF | 51.8 | 51.4 | 52.0 | 51.1 | 51.9 | 51.2 | 51.5 | 51.5 | 2.65 | 0.930 | 0.459 | 0.191 | 0.073 | 0.842 | 0.785 | 0.777 | 0.951 |
| SM | 54.7 | 54.4 | 54.8 | 54.2 | 54.7 | 54.2 | 54.5 | 54.5 | 2.28 | 0.655 | 0.542 | 0.641 | 0.045 | 0.589 | 0.932 | 0.491 | 0.680 |
| Intramuscular fatty acids (loin) | |||||||||||||||||
| C16:0 | 24.0 | 23.7 | 24.1 | 23.6 | 23.9 | 23.8 | 24.4 | 23.4 | 1.02 | 0.001 | 0.655 | 0.053 | 0.488 | 0.000 | 0.350 | 0.894 | 0.758 |
| C18:0 | 12.5 | 12.6 | 12.7 | 12.4 | 12.6 | 12.6 | 12.8 | 12.3 | 0.92 | 0.017 | 0.192 | 0.219 | 0.189 | 0.010 | 0.048 | 0.145 | 0.216 |
| C18:1 n-9 | 42.4 | 41.6 | 42.3 | 41.9 | 42.4 | 41.8 | 42.1 | 42.0 | 2.03 | 0.361 | 0.153 | 0.777 | 0.220 | 0.938 | 0.371 | 0.872 | 0.920 |
| C18:2 n-6 | 8.0 | 8.8 | 7.9 | 8.8 | 8.2 | 8.5 | 7.7 | 8.9 | 1.83 | 0.041 | 0.290 | 0.107 | 0.325 | 0.008 | 0.923 | 0.578 | 0.778 |
| C18:3 n-3 | 0.4 | 0.4 | 0.36 | 0.38 | 0.4 | 0.4 | 0.35 | 0.39 | 0.06 | 0.010 | 0.135 | 0.036 | 0.287 | 0.010 | 0.936 | 0.199 | 0.685 |
| SFA | 38.1 | 38.0 | 38.5 | 37.7 | 38.1 | 38.0 | 38.9 | 37.3 | 1.71 | 0.000 | 0.697 | 0.064 | 0.802 | 0.010 | 0.063 | 0.395 | 0.672 |
| MUFA | 50.4 | 49.4 | 50.2 | 49.7 | 50.2 | 49.7 | 50.1 | 49.8 | 2.28 | 0.322 | 0.128 | 0.623 | 0.144 | 0.990 | 0.470 | 0.964 | 0.913 |
| PUFA | 11.4 | 12.6 | 11.3 | 12.6 | 11.7 | 12.2 | 11.0 | 12.9 | 2.92 | 0.044 | 0.303 | 0.141 | 0.319 | 0.010 | 0.889 | 0.641 | 0.738 |
| Δ9-Desat | 55.6 | 55.1 | 55.3 | 55.5 | 55.5 | 55.3 | 54.9 | 55.8 | 1.6 | 0.004 | 0.281 | 0.166 | 0.339 | 0.000 | 0.036 | 0.628 | 0.626 |
| UI | 79.2 | 81.5 | 78.8 | 81.8 | 79.7 | 80.9 | 77.8 | 82.6 | 6.36 | 0.012 | 0.427 | 0.113 | 0.440 | 0.010 | 0.635 | 0.604 | 0.665 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romero, M.; Calvo, L.; Morales, J.I.; Magro, A.; Rodríguez, A.I.; Segura, J.; Escudero, R.; López-Bote, C.; Olivares, Á. Short- and Long-Term Effects of Split-Suckling in Pigs According to Birth Weight. Animals 2023, 13, 3521. https://doi.org/10.3390/ani13223521
Romero M, Calvo L, Morales JI, Magro A, Rodríguez AI, Segura J, Escudero R, López-Bote C, Olivares Á. Short- and Long-Term Effects of Split-Suckling in Pigs According to Birth Weight. Animals. 2023; 13(22):3521. https://doi.org/10.3390/ani13223521
Chicago/Turabian StyleRomero, María, Luis Calvo, José Ignacio Morales, Antonio Magro, Ana Isabel Rodríguez, José Segura, Rosa Escudero, Clemente López-Bote, and Álvaro Olivares. 2023. "Short- and Long-Term Effects of Split-Suckling in Pigs According to Birth Weight" Animals 13, no. 22: 3521. https://doi.org/10.3390/ani13223521
APA StyleRomero, M., Calvo, L., Morales, J. I., Magro, A., Rodríguez, A. I., Segura, J., Escudero, R., López-Bote, C., & Olivares, Á. (2023). Short- and Long-Term Effects of Split-Suckling in Pigs According to Birth Weight. Animals, 13(22), 3521. https://doi.org/10.3390/ani13223521