Simple Summary
The sea urchin (Heliocidaris crassispina) is an ecologically important invertebrate in structuring marine benthic communities. Most dwell within intertidal regions, rendering them highly susceptible to elevated artificial light at night. However, their potential acclimation to artificial light exposure remains largely unexplored. This study investigates the changes of prolonged artificial light pollution on the fitness-related traits of sea urchins. Following a six-week exposure to artificial light at night, survival remained unaffected, while behavioral responses exhibited slower reactions. Concurrently, growth inhibition was observed in sea urchins, which might be attributed to reduced mouthparts weight and decreased food consumption. This study also revealed that sea urchin gonads are more susceptible to artificial light than guts. Pax6 gene expression may serve as a sensitive indicator to assess the impact on the photosensitive system of sea urchins. These results increase our understanding of the effects of artificial light at night on sea urchins and provide valuable information about coastal animals’ safety.
Abstract
Limited data are available regarding the effects of elevated coastal artificial light at night (ALAN) on intertidal echinoderms. In this study, we investigated the behavioral, morphological, and physiological responses of the sea urchin (Heliocidaris crassispina) after continuous exposure to ALAN at light intensities of 0.1, 300, and 600 Lux for 6 weeks. Our findings revealed that ALAN at 300 Lux substantially reduced food consumption, Lantern weight, and gonadosomatic index (GSI). On the other hand, ALAN at 600 Lux notably prolonged the righting and covering response times and elevated the 5-HIAA/5-HT ratio, while concurrently decreasing food consumption, body weight, Lantern weight, GSI, and Pax6 gene expression. These results indicated that continuous exposure to ALAN could cause an adverse effect on fitness-related traits, including behavioral responses, growth, reproductive performance, and photoreception of sea urchins. The present study provides new insights on the impact of light pollution on echinoderms.
1. Introduction
Artificial light at night (ALAN) changes the natural state of a dark environment from dark to bright by increasing the distribution and intensity of light in space [1,2,3]. As coastal population densities surge, the proliferation of ALAN is expanding at an annual growth rate of approximately 6% [2,4]. While ALAN offers substantial convenience for human activities, it concurrently appears to present a potential threat to coastal ecosystems [2,5,6,7,8,9]. For example, in shallow water coral reef ecosystems, recent studies reported the gametogenesis cycle of Acropora millepora was delayed or masked by exposure to ALAN [10]. In sandy beach ecosystems, sea turtle orientation was negatively affected by ALAN, impairing the ability of hatchlings to respond to natural orientation cues [11,12]. In seabird communities, increasing ALAN levels in and around nesting colonies could impact sea mew breeding behavior and, consequently, chick provisioning [13]. However, scant attention has been directed towards investigating the implications of ALAN in intertidal ecosystems, particularly concerning invertebrates such as slow-moving echinoderms. Therefore, a comprehensive grasp of how intertidal invertebrates’ fitness reacts to ALAN exposure is imperative as it provides essential insights into potential ecological risks in coastal environments.
Sea urchin Heliocidaris crassispina (previously Anthocidaris crassispina) is an echinoderm species found across a wide range, from the intertidal zone to shallow waters in the Western Pacific region. Its habitat spans from the Japan Sea (at 38 °N) to Hainan Island (at 19 °N) [14,15,16,17]. This species holds a crucial role in intertidal ecosystems, acting both as an herbivorous grazer on macroalgal species such as Ulva Lactuca and as prey for crabs and starfish [18,19]. Echinoderm behaviors (such as righting, covering, and feeding) are important for their fitness but also affect intertidal ecosystems through effects on their predators, prey, and competitors [20]. Righting is the behavior of inverted sea urchins placed on their aboral surface to resume the posture with the aboral side up [21,22]. Covering behavior refers to sea urchins using their tubular feet and spines to move objects such as algae fragments, stones, and shells to the dorsal surface [23,24,25]. These two behaviors are used to help sea urchins defend against abiotic disturbances and predation [24]. Health-related attributes, such as food consumption, body weight, and gut-weight, alongside reproductive traits like gonad weight, constitute foundational elements influencing the fitness of sea urchins [20,21]. Serotonin serves as one of the monoamine neurotransmitters integral to regulating animal behavior [26]. It can be used to reflect the physiological state of sea urchins when their behavioral expression changes. Pax6 is a pivotal opsin transcription factor gene within the animal photosensitive system. Deviations in Pax6 gene expression could indicate a disruption of the light defense system in sea urchins [27].
The primary objective of this study is to examine the effects of prolonged artificial lighting on the righting and covering behaviors, health-related and reproductive traits, coelomic monoaminergic activity, and Pax6 gene expression in H. crassispina. This investigation aims to offer novel insights into the repercussions of ALAN on intertidal creatures, alongside potential underlying mechanisms.
2. Materials and Methods
Before commencing the study, we meticulously reviewed animal experiments and welfare regulations, ensuring strict adherence. All procedures undertaken in this study received approval from the Institutional Animal Care and Use Committee of Zhejiang Ocean University.
2.1. Sea Urchin Materials
Sea urchins H. crassispina were collected from the waters near Zhoushan Islands (122°41’27.040” E, 30°12’41.625” N) by scuba diving and delivered to a 5000 L PVC tank in the Culture Enhancement Laboratory of Zhejiang Ocean University for acclimation. The PVC tank was supplied with filtered seawater at constant aeration and a flow rate of 10 L/min. The seawater was changed every 5 days and its temperature was set at 24.5–26.5 °C. Daytime lighting was provided by low-pressure mercury discharge fluorescent lamps (626–1023 Lux, 12 h light-12 h dark). Sea urchins were fed ad libitum with chopped U. Lactuca and their droppings were cleaned every day [21,28].
2.2. Experimental Design
After 3 weeks of acclimation, seventy-two sea urchins were selected from the PVC tank and weighed for this experiment (body weight ± SD =54.402 ± 6.240 g, n = 72). The weighed sea urchins were then randomly divided evenly into nine 150 L glass tanks (long × wide × deep = 60 cm × 50 cm × 50 cm) placed in the laboratory (n = 8 for each tank; three tanks for each group; n = 24 for each group). The glass tanks’ condition during the experiment was similar to the PVC tank except for artificial light illumination at night. Nocturnal illumination stimulation was provided by metal halide streetlights (HC-ZY521, OSRAM, Munich, Germany) mounted on the glass tanks. Neutral density filters (248565, Grand Unified Optics, Beijing, China) were utilized to attenuate the intensity of the streetlights, thereby creating varying levels of nocturnal light intensities on the water surface, while ensuring no alteration to the light spectrum. Three groups were thus set up which, respectively, represent the simulated living environment in three situations (Figure 1a) [29,30,31,32,33]. The first group was the control group, simulating a moon-transparent night (ALAN = ~0.1 Lux) [29,33]. The second group was stimulated continuously with low artificial light to simulate the marine environment under the influence of city lights (ALAN = ~300 Lux) [29,30,31]. The third group was given continuous intense artificial light stimulation to simulate the marine environment of direct irradiation near industrial port facilities (ALAN = ~600 Lux) [32,33].
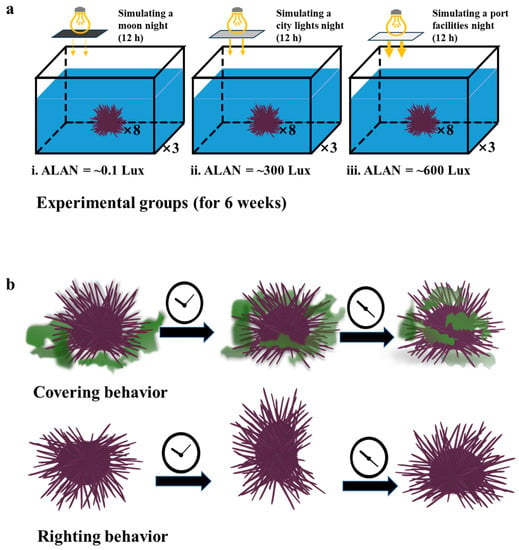
Figure 1.
Schematic drawing of the three ALAN experimental groups (a) and the representative fitness behaviors (b) of sea urchins.
During the experiment, dead sea urchins (if any) were removed and recorded every day. Water quality assessments were performed at weekly intervals utilizing a portable water quality monitoring meter (YSI, Yellow Springs, OH, USA) (Supplementary Materials: Table S1). Six weeks later, the three groups of sea urchins that had been subjected to different long-term artificial light treatments were used for subsequent behavioral and physiological analysis.
2.3. Behavioral Analysis
Righting behavior, covering behavior, and food consumption of H. crassispina were measured in glass tanks (long × wide × deep = 40 × 30 × 30 cm; 20 cm of aeration water depth). Righting behavior [16,20,21,22,23]: A sea urchin was placed at the tank’s center with its aboral side facing downward, and the time taken for it to reorient itself to the oral side facing downward was measured as the righting response time. If the sea urchin failed to reorient within 10 min, the righting response time was recorded as 600 s. This experimental protocol aligns with previous studies on covering behavior [23,24,25,27]: A sea urchin was carefully positioned at the central point of the tank, with U. Lactuca available ad libitum at the bottom. The time required for the sea urchin to cover its body with U. Lactuca was recorded as the covering response time. If the individual failed to engage in covering behavior within a 30-min timeframe, its covering response time was recorded as 1800 s (Figure 1b). Food consumption [16,20,21]: Four sea urchins were delicately placed at the center of the tank, with weighted U. Lactuca available ad libitum at the bottom. The sea urchins began to feed about two hours later. Food consumption was quantified by subtracting the remaining uneaten quantity of U. Lactuca in the tank after a 24-h period from the initially supplied amount. To mitigate observer bias, a double-blinded approach was employed for the recording and analysis of all behavioral data.
2.4. Sampling Protocol
The sampling procedure of H. crassispina was conducted by the method described in previous papers [34]. Test diameter, test height, and Aristotle’s lantern length were assessed with a vernier caliper, with measurements recorded to the nearest 0.1 mm (CD-AX/C, MITUTOYO, Kawasaki city, Japan). Test weight, Aristotle’s lantern weight, gonads weight, and gut-weight were measured to the nearest 0.01 g by an electronic balance (C3-6553, AS ONE, Osaka, Japan). Subsequently, coelomic fluid was extracted and subjected to centrifugation at 10,000 rpm for 10 min in a refrigerated centrifuge (4 °C). The resulting supernatant was meticulously collected and preserved at −80 °C for subsequent analysis of monoaminergic activity. Tubular feet were cut off and washed thoroughly with ice-cold artificial coelomic fluid (10 mM CaCl2; 14 mM KCl; 50 mM MgCl2; 398 mM NaCl; 1.7 mM Na2HCO3; 25 mM Na2SO4) [35], and frozen quickly in liquid nitrogen for gene expression analysis. Test height/test diameter [25] and gonads weight/test weight (GSI) [20] were subsequently calculated.
2.5. Physiological Analysis
The concentrations of 5-hydroxytryptamine (5-HT) and 5-hydroxyindoleacetic acid (5-HIAA) in body cavity liquid samples were measured by commercial enzyme-linked immunosorbent assay (ELISA) Assay Kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) according to the manufacturer’s instructions. These ELISA Assay Kits have been validated in marine life previously [36,37,38,39]. Pax6 gene expression levels of tubular feet samples were measured using quantitative real-time polymerase chain reaction (qPCR) [40,41]. Total RNA was extracted using Trizol Reagent (Invitrogen, Carlsbad, CA, USA) containing bromochloropropane for phase separation. Afterward, RNA integrity was confirmed via electrophoresis on a 1.0% agarose gel, and quantification was conducted using a NanoDrop 2000 spectrophotometer (Thermo Scientific, Waltham, WA, USA). Subsequently, double-strand cDNA was synthesized from the total RNA template, and the construction of a cDNA library was achieved using the SMART cDNA construction kit (Clontech, Palo Alto, CA, USA). Primers for the study were designed utilizing Primer Premier 6 software (PREMIER Biosoft International, Palo Alto, CA, USA) and obtained commercially from Sangon Biotech (Shanghai, China). As a reference gene in sea urchins [42] (Table 1), the internal control gene 18S rRNA was employed. Each qPCR reaction involved a 20 μL volume, including 10 μL of the 2× UltraSYBR Mixture, 0.2 μM specific forward and reverse primers, and 1.0 μL of the diluted cDNA template (10 ng/μL). The thermal cycling process was carried out on the ABI Prism 7900HT Sequence Detection System (PE Applied Bio-systems, Foster City, CA, USA), encompassing an initial denaturation step at 95 °C for 30 s, followed by 40 cycles of denaturation at 94 °C for 5 s, annealing at 60 °C for 30 s, and extension at 72 °C for 10 s. To confirm the specificity of amplification, a melting curve analysis of the PCR products was executed. Relative mRNA levels were determined using the 2−ΔΔCt method [43]. The procedure of Pax6 gene full-length conventional cloning of H. crassispina was fully described in the Supplementary Materials: Procedure (Sp).

Table 1.
Nucleotide sequences of the primers for qPCR.
2.6. Data Statistics
Data throughout the text, tables, and figures are expressed as means ± standard error (S.E.). Statistical significance was established at p < 0.05. Data conforming to a normal distribution with equal variances underwent one-way analysis of variance (ANOVA) followed by Duncan’s multiple range post hoc test for statistical examination. When necessary, data were subjected to appropriate transformations, such as square root, exponential, or logarithm, to meet the assumptions of normal distribution and homogeneity of variance. Statistical analyses were conducted utilizing SPSS 17.0 for Windows (SPSS, IBM, Armonk, NY, USA, 2007).
3. Results
3.1. Survival
We found that ALAN at 0.1, 300, and 600 Lux could not cause sea urchin death. There was no significant difference in the number of surviving sea urchins among the individuals exposed to 0.1, 300, and 600 Lux of ALAN. However, sea urchins exposed to ALAN at 600 Lux showed severe spine loss during the experiment.
3.2. Righting and Covering Behavior
We found that ALAN at 600 Lux significantly increased the righting response time of H. crassispina compared to those exposed to ALAN at 0.1 and 300 Lux (p < 0.005; p < 0.005). However, there was no significant difference between the individuals exposed to 0.1 and 300 Lux of ALAN (p = 0.313; Figure 2a). Similarly, ALAN at 600 Lux significantly increased the covering response time of H. crassispina compared to those exposed to ALAN at 0.1 and 300 Lux (p < 0.005; p < 0.005). However, there was no significant difference between the individuals exposed to 0.1 and 300 Lux of ALAN (p = 0.442; Figure 2b).
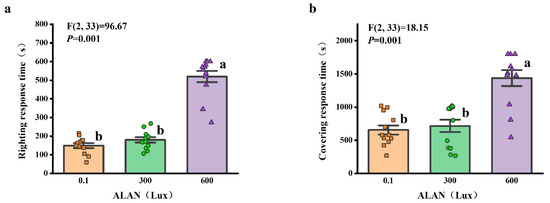
Figure 2.
Effects of ALAN at different intensities for 6 weeks on righting response time (a) and covering response time (b) of sea urchins. Different letters indicate significant differences. Values are shown in data points and mean ± S.E. (n = 12).
3.3. Food Consumption
We found that the food consumption of H. crassispina significantly decreased after 6 weeks of ALAN exposure. Sea urchins exposed to ALAN at 300 Lux and 600 Lux resulted in a significant decrease in food consumption compared with the control (0.1 Lux) (p < 0.05; p < 0.05). However, there was no significant difference between the individuals exposed to 300 and 600 Lux of ALAN (p = 0.558; Figure 3).
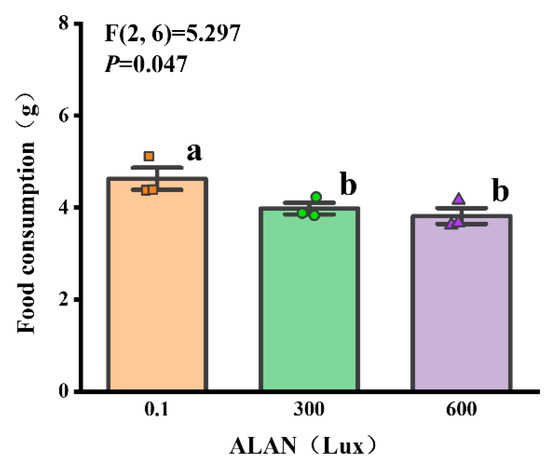
Figure 3.
Effects of ALAN at different intensities for 6 weeks on food consumption of sea urchins. Different letters indicate significant differences. Values are shown in data points and mean ± S.E. (n = 3).
3.4. Body Size
We found that ALAN at 600 Lux significantly decreased the body weight of H. crassispina compared to those exposed to ALAN at 0.1 Lux (p < 0.05; Figure 4a). However, there was no significant difference between the individuals exposed to 0.1 and 300 Lux or 300 and 600 Lux of ALAN (p = 0.133; p = 0.245; Figure 4a). In addition, there was no significant difference in test height, test diameter, and test height/test diameter among the individuals exposed to 0.1, 300, and 600 Lux of ALAN (Figure 4b–d).
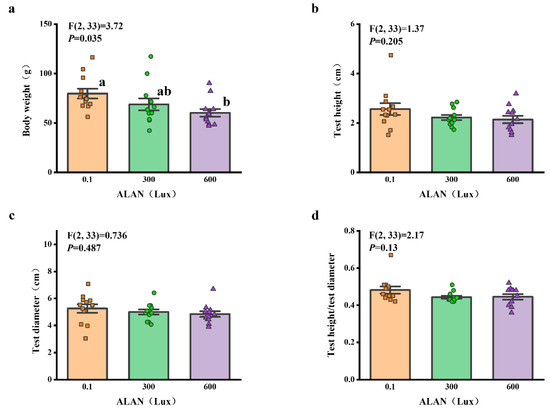
Figure 4.
Effects of ALAN at different intensities for 6 weeks on body weight (a), test height (b), test diameter (c), and test height/test diameter (d) of sea urchins. Different letters indicate significant differences. Values are shown in data points and mean ± S.E. (n = 12).
3.5. Lantern Weight and Length
We found that Aristotle’s lantern weight of H. crassispina significantly decreased exposure to ALAN at 300 Lux and 600 Lux compared to those exposed to ALAN at 0.1 (p < 0.05; p < 0.05). However, there was no significant difference between the individuals exposed to 0.1 and 300 Lux (p = 0.906; Figure 5a). In addition, Aristotle’s lantern length among the individuals exposed to 0.1, 300, and 600 Lux of ALAN showed no significant difference after 6 weeks of the experiment (Figure 5b).
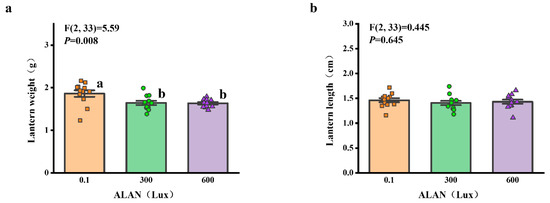
Figure 5.
Effects of ALAN at different intensities for 6 weeks on Aristotle’s lantern weight (a) and Aristotle’s lantern length (b) of sea urchins. Different letters indicate significant differences. Values are shown in data points and mean ± S.E. (n = 12).
3.6. Gonad and Gut-Weight
We found that there was a significant difference in GSI (gonads weight/test weight) among the individuals exposed to 0.1, 300, and 600 Lux of ALAN (all p < 0.05; Figure 6a). There was no significant difference in gut-weight among the individuals exposed to 0.1, 300, and 600 Lux of ALAN (Figure 6b).
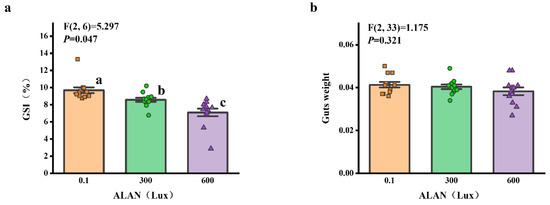
Figure 6.
Effects of ALAN at different intensities for 6 weeks on GSI (a) and gut-weight (b) of sea urchins. Different letters indicate significant differences. Values are shown in data points and mean ± S.E. (n = 12).
3.7. 5-HIAA/5-HT Ratio
We used the 5-HIAA/5-HT ratio as an indicator of serotonin system activity, and we found that ALAN at 600 Lux significantly increased the 5-HIAA/5-HT ratio of H. crassispina compared to those exposed to ALAN at 0.1 and 300 Lux of ALAN (p < 0.005; p < 0.005). However, there was no significant difference between the individuals exposed to 0.1 and 300 Lux of ALAN (p = 0.054; Figure 7).
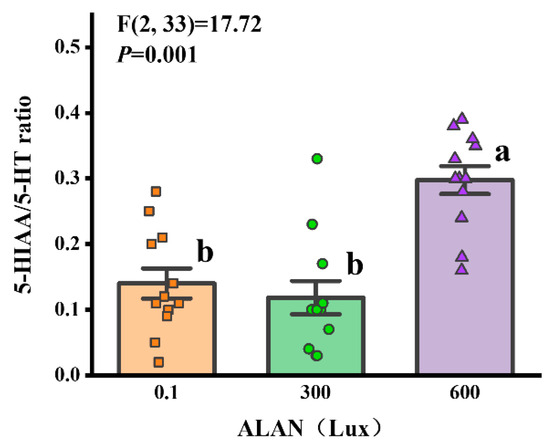
Figure 7.
Effects of ALAN at different intensities for 6 weeks on coelomic fluid 5-HIAA/5-HT ratio of sea urchins. Different letters indicate significant differences. Values are shown in data points and mean ± S.E. (n = 12).
3.8. Pax6 Gene Expression
We found that Pax6 gene expression was significantly affected by ALAN at 600 Lux in H. crassispina compared with the control (0.1 Lux) (p < 0.005; p < 0.005). No significant difference in Pax6 gene expression was found between the individuals exposed to 0.1 and 300 Lux of ALAN (p = 0.192; Figure 8).
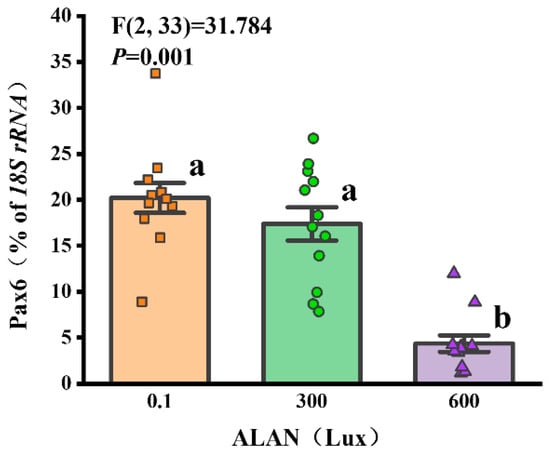
Figure 8.
Effects of ALAN at different intensities for 6 weeks on Pax6 gene expression of sea urchins. Different letters indicate significant differences. Values are shown in data points and mean ± S.E. (n = 12).
4. Discussion
In this work, we constructed small-scale systems to investigate the effects of artificial light at night (ALAN) on fitness-related traits of H. crassispina. The main results were as follows: (1) ALAN at 600 Lux significantly increased the righting response time, the covering response time, and the 5-HIAA/5-HT ratio, and decreased food consumption, body weight, Lantern weight, GSI, and Pax6 gene expression; (2) ALAN at 300 Lux significantly decreased food consumption, Lantern weight, and GSI.
4.1. Effects of ALAN on Behavioral Responses
Complex marine benthic environments shape a number of ecologically important behaviors in sea urchins. Covering and righting are two representative behaviors with fitness importance in antipredation [21,22,23,24,25]. The diminished covering and righting capacities of H. crassispina observed in this study suggest that extended exposure to ALAN might unfavorably impact fitness-related behaviors. This behavioral shift was linked to the decrease in sea urchin populations, potentially attributed to heightened predation pressure within the coastal ecosystem [28]. Similar outcomes highlighting the influence of ALAN on fitness-related behaviors have been documented in diverse coastal organisms. For instance, sea turtles (Caretta caretta and Chelonia mydas) may fail to navigate toward the ocean post-hatching due to ALAN-induced disruptions in their behavioral orientation [44]. Seabird migratory patterns also exhibit alteration, as birds are drawn to and linger around artificial light sources [45,46,47]. The composition of shallow water fish species underwent modification under ALAN influence: large predatory and small shoaling fish congregated around illuminated zones during the night, leading to reduced prey availability [30]. Notably, a study involving mud snails (Hydrobia ulvae) on French mudflats unveiled a decline in the crawling activity (and growth) of snails in response to light exposure [48].
The formation of metabolites like 5-HIAA arises subsequent to the re-uptake of 5-HT from the synaptic cleft, and their accumulation in neural tissue likely exhibits a time-dependent pattern [36]. Consequently, evaluating tissue concentrations of neurotransmitter metabolites fails to provide an immediate depiction of neural activity. As such, the ratio of metabolite to monoamine was adopted as a marker indicating heightened monoamine utilization. In this study, the 5-HIAA/5-HT ratio in H. crassispina exhibited an increase following prolonged ALAN exposure. This observation suggests that the serotonin system was stimulated in response to light pollution, causing a progressive augmentation in metabolite levels over time. Numerous reports have delved into serotonin’s influence on animal behavior, consistently revealing its inhibitory effects on traits such as aggressive behavior, feeding patterns, and motor activity [36,49]. Therefore, alterations in the concentration of the monoamine transmitter serotonin may underlie the internal mechanism driving the behavioral shifts observed in sea urchins within this study.
4.2. Effects of ALAN on Growth
H. crassispina subjected to prolonged artificial light exposure displayed a pronounced reduction in food consumption compared to those shielded from ALAN. This correlation is substantiated by our finding that ALAN significantly suppressed Aristotle’s lantern reflex, a pivotal factor in sea urchin food intake. This diminished food consumption provides a plausible explanation for the subsequent reduction in body size. These findings strongly indicate that light pollution could exert an impact on the growth of intertidal organisms. Correspondingly, studies by Luarte et al. involving both laboratory and field investigations demonstrated that the foraging behavior and growth rate of the amphipod Orchestoidea tuberculate were significantly affected by ALAN [50]. Field observations indicated that lower light levels (60 Lux) curtailed amphipod feeding and growth rates. The growth rates of amphipods under ambient conditions (darkness) were nearly three times higher compared to those exposed to ecological light pollution. Similarly, under ambient conditions, the food consumption rates were nearly two times higher than under exposure to ecological light pollution. The potential influence of light pollution on growth and development also extends to mollusks. Research unveiled a correlation between daily light exposure duration and the growth pace and stored energy levels in the pulmonate Lymnaea stagnalis [51]. These outcomes find alignment with investigations on rodents, specifically Myotis emarginatus, showcasing that light pollution diminishes consumption rates and impedes the growth of juvenile and suckling bats [52]. Thus, the decreased food consumption stemming from light pollution is likely to curtail the fitness of littoral organisms by reducing energy reserves.
4.3. Effects of ALAN on Reproductive Performance
Among the pivotal factors influencing sea urchin reproduction, food consumption holds paramount significance [20,21]. To gauge reproductive performance, we employed the Gonad-Somatic Index (GSI) and identified a GSI reduction in H. crassispina exposed to ALAN, which correlated with diminished food intake. This could suggest that sea urchins possibly allocate nutrients and energy to combat environmental stress, consequently compromising gonad tissue development. Furthermore, ALAN exhibited no significant influence on gut-weight in H. crassispina, a finding consistent with previous research [21]. This implies that sea urchin gonads are potentially more susceptible to environmental stressors than their gastrointestinal counterparts. Thus, ALAN exposure might detrimentally impact the reproductive capacity of sea urchins.
The development of gonad tissue follows a hormonal cascade, with gonadotropin-releasing hormone (GnRH) triggering the release of luteinizing hormone (LH) and follicle-stimulating hormone (FSH) from the pituitary gland, leading to the production of sex steroids in vertebrates [53]. The regulatory mechanism governing gonadal development is less defined in invertebrate sea urchins. As a result, we did not investigate relevant hormones to probe the mechanism through which light pollution affects sea urchin reproduction. Nonetheless, analogous studies have been conducted in marine fish. For instance, in female Lateolabrax maculates, mRNA expression of luteinizing hormone and follicle-stimulating hormone was suppressed even at white light levels as low as 1 Lux at night [54]. Similarly, a study in a natural setting found diminished mRNA expression of gonadotropins (luteinizing hormone and follicle-stimulating hormone) and circulating sex hormones (17β-oestradiol and 11-ketotestosterone) in European perch under street lighting conditions (13.3–16.5 Lux at the surface) [55]. Consequently, exploring the internal mechanism of light’s impact on gonad development in invertebrates such as sea urchins should be a prospective avenue of research.
4.4. Effects of ALAN on Pax6 Gene Expression
In a pioneering effort, we successfully isolated and sequenced a complete cDNA encoding a 440-amino-acid Pax6 from the tubular feet of H. crassispina. We conducted an in-depth analysis of Pax6’s similarity and evolutionary relationships across various species (Supplementary Materials: Table S2, Figure S1–S3), thereby furnishing fundamental biological insights for subsequent investigations into the Pax6 gene within sea urchins. Notably, Pax6 stands as a transcription factor gene with established involvement in eye development and photoreception in ocular-forming organisms [56]. The presence of Pax6 expression signifies the preservation of photoreceptor cell specification processes, functioning as a canonical eye gene even in creatures devoid of specialized eyes [57]. Consequently, the Pax6 gene emerges as a central focus for probing the photosensitive systems responding to the light milieu in sea urchins. For instance, the relative Pax6 expression in Strongylocentrotus intermedius displayed a negative correlation with natural light intensity. However, Pax6 expression patterns in S. intermedius underwent alterations following UV-B radiation exposure [58]. In line with this, our present experiment reveals that prolonged ALAN exposure also disrupts the typical expression of the pax6 gene in H. crassispina. This suggests that persistent light stimulation might perturb molecules inherent to the photosensitive systems of sea urchins, ultimately yielding anomalous gene expression. On the whole, Pax6 gene expression is poised to function as a sensitive gauge for assessing potential damage to the photosensitive system of sea urchins.
4.5. Sea Urchins in the Wild Light Environment
The profound impact of nocturnal light on the fitness traits of sea urchins can be attributed to their inherent nocturnal activity patterns [2]. Continuous exposure to light inhibits sea urchin activity, consequently slowing down behaviors such as covering and righting, as well as impairing their feeding efficiency. Additionally, this light exposure also leads to changes in hormone levels and gene expression within their bodies. This strong correlation between nocturnal light and the behavioral responses of sea urchins highlights the significant role that light plays in regulating their activity levels and overall ecological dynamics [31]. Despite this, considering the complexity of intertidal habitats, the impact of nocturnal light on sea urchins in the wild may not be consistent, especially in rocky coastal areas where cracks, rocks, and shade-providing seaweeds are present. Sea urchins may opt to inhabit shaded areas or regions with suitable light intensity. For instance, a study conducted on a drumfish species found that their activity range shifted in response to nighttime light exposure of 70 Lux [59]. However, the narrowing of the activity range was determined by the encroachment of intensifying nocturnal light into dark spaces. Therefore, it is crucial to adequately understand the complexity of natural settings. Further research conducted in the wild, including the use of large mesocosms, or in more natural laboratory settings, will be necessary.
5. Conclusions
These findings illuminate the potential detrimental impact of extended (6-week) exposure to ALAN on the behavioral responses, growth, reproductive performance, and photoreception of sea urchins. Importantly, the study findings reveal that the sea urchins’ responses to light are more pronounced when exposed to higher light intensities, emphasizing the increased threat posed by intensified artificial light at night (ALAN) to their overall fitness. This highlights the potential risks that intertidal coastal ecosystems face in areas characterized by prominent luminous installations, such as stadiums, ports, and wharves. These three locations, due to their excessive lighting, emerge as significant sources of disturbance and risk for the delicate balance of these ecosystems. Consequently, it is crucial to consider the conservation and management of these areas to mitigate the adverse effects of ALAN and preserve the ecological health and biodiversity of intertidal coastal environments. This study significantly advances our comprehension of the fitness-related attributes of sea urchins under ALAN exposure, furnishing crucial insights into the ecological risks of coastal regions.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ani13193035/s1, Figure S1: The deduced amino acid sequences of Pax6 from H. crassispina, Figure S2: Alignment of deduced amino acid sequences of Pax6 from H. crassispina and select species, Figure S3: Phylogenetic tree illustrating the relationship of Pax6 from H. crassispina and select species, Table S1: Water quality conditions during the experiment; Procedure (Sp): Cloning and analysis of full-length Pax6 gene, Table S2: Nucleotide sequences of the primers for PCR.
Author Contributions
Conceptualization, T.W.; investigation, X.X. and Z.W.; data curation, X.X. and K.D.; validation, J.Y.; format analysis, X.X. and Z.W.; resources, X.X. and X.J.; writing—original draft preparation, X.X. and T.W.; writing—review and editing, X.X., T.W. and J.Y.; project administration, T.W; funding acquisition, X.X. and T.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the “Pioneer” and “Leading Goose” R&D Program of Zhejiang (2022C02040) and the Science and Technology Program of Zhoushan (2022C41016).
Institutional Review Board Statement
Ethical review and approval were waived for this study due to no authorization being required for experimental studies on invertebrates such as sea urchins. The experiment with local laws and regulations, adheres to basic ethical principles, and ensures respect and responsible utilization of the animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available upon reasonable request from the corresponding author.
Acknowledgments
The authors also thank the Fisheries Research Institute of Zhoushan Zhejiang for their support and help in providing data and sample collection throughout the study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gaston, K.J.; Bennie, J.; Davies, T.W.; Hopkins, J. The ecological impacts of nighttime light pollution: A mechanistic appraisal. Biol. Rev. 2013, 88, 912–927. [Google Scholar] [CrossRef] [PubMed]
- Zapata, M.J.; Sullivan, S.M.P.; Sullivan, S.M. Artificial lighting at night in estuaries implications from individuals to ecosystems. Estuar. Coast. 2019, 42, 309–330. [Google Scholar] [CrossRef]
- Bassi, A.; Love, O.P.; Cooke, S.J.; Warriner, T.R.; Harris, C.M.; Madliger, C.L. Effects of artificial light at night on fishes: A synthesis with future research priorities. Fish Fish. 2022, 23, 631–647. [Google Scholar] [CrossRef]
- Davies, T.W.; Duffy, J.P.; Bennie, J.; Gaston, K.J. Stemming the tide of light pollution encroaching into marine protected areas. Conserv. Lett. 2016, 9, 164–171. [Google Scholar] [CrossRef]
- Davies, T.W.; Smyth, T. Why artificial light at night should be a focus for global change research in the 21st century. Global Change Biol. 2018, 24, 872–882. [Google Scholar] [CrossRef]
- Marangoni, L.F.B.; Davies, T.; Smyth, T.; Rodríguez, A.; Hamann, M.; Duarte, C.; Pendoley, K.; Berge, J.; Maggi, E.; Levy, O. Impacts of artificial light at night in marine ecosystems-A review. Global Change Biol. 2022, 28, 5346–5367. [Google Scholar] [CrossRef]
- Hussein, A.A.A.; Bloem, E.; Fodor, I.; El-Sayed, B.; Tadros, M.M.; Soliman, M.F.M.; El-Shenawy, N.S.; Koene, J.M. Slowly seeing the light: An integrative review on ecological light pollution as a potential threat for mollusks. Environ. Sci. Pollut. Res. Int. 2021, 28, 5036–5048. [Google Scholar] [CrossRef]
- Davies, T.W.; Duffy, J.P.; Bennie, J.; Gaston, K.J. The nature, extent, and ecological implications of marine light pollution. Front. Ecol. Environ. 2014, 12, 347–355. [Google Scholar] [CrossRef]
- Russart, K.L.G.; Nelson, R.J. Artificial light at night alters behavior in laboratory and wild animals. J. Exp. Zool. Part A 2018, 329, 401–408. [Google Scholar] [CrossRef]
- Ayalon, I.; Rosenberg, Y.; Benichou, J.I.C.; Campos, C.L.D.; Sayco, S.L.G.; Nada, M.A.L.; Baquiran, J.I.P.; Ligson, C.A.; Avisar, D.; Conaco, C.; et al. Coral gametogenesis collapse under artificial light pollution. Curr. Biol. 2021, 31, 413–419. [Google Scholar] [CrossRef]
- Colman, L.P.; Lara, P.H.; Bennie, J.; Broderick, A.C.; de Freitas, J.R.; Marcondes, A.; Witt, M.J.; Godley, B.J. Assessing coastal artificial light and potential exposure of wildlife at a national scale: The case of marine turtles in Brazil. Biodivers. Conserv. 2020, 29, 1135–1152. [Google Scholar] [CrossRef]
- Shimada, T.; Meekan, M.G.; Baldwin, R.; Al-Suwailem, A.M.; Clarke, C.; Santillan, A.S.; Duarte, C.M. Distribution and temporal trends in the abundance of nesting sea turtles in the Red Sea. Biol. Conserv. 2021, 261, 109235. [Google Scholar] [CrossRef]
- Ronconi, R.A.; Allard, K.A.; Taylor, P.D. Bird interactions with offshore oil and gas platforms: A review of impacts and monitoring techniques. J. Environ. Manag. 2015, 147, 34–45. [Google Scholar] [CrossRef]
- Maboloc, E.A.; Fang, J.K.H. Tissue regeneration of the purple sea urchin Heliocidaris crassispina. Bull. Mar. Sci. 2023, 99, 19–20. [Google Scholar] [CrossRef]
- Kanamoto, Y.; Nakamura, K. Age and growth analyses of the purple sea urchin Heliocidaris crassispina inhabiting different feeding environments in the Shimane Peninsula, Japan. Reg. Stud. Mar. Sci. 2023, 65, 103096. [Google Scholar] [CrossRef]
- Kadota, T.; Kiyomoto, S.; Masuda, Y.; Miyano, T.; Yoshimura, T. Restoration of a small-sized macroalgal bed through the removal of sea urchins in Kashiyama, Nagasaki Prefecture. Nippon. Suisan Gakk. 2022, 88, 49–57. [Google Scholar] [CrossRef]
- Feng, W.P.; Nakabayashi, N.; Inomata, E.; Inomata, E.; Aoki, M.N.; Agatsuma, Y. Sexually unbalanced gonad development and nutrition of the newly range-extended sea urchin Heliocidaris crassispina in the northeastern Honshu, Japan. Estuar. Coast. Shelf S. 2021, 249, 107120. [Google Scholar] [CrossRef]
- Ishii, M.; Unuma, T.; Masadate, A.; Hoshikawa, H.; Takahashi, K.; Kosaka, S.; Masuda, A.; Murakami, K. Accelerated photoperiod promotes gonadal maturation in the sea urchin Strongylocentrotus intermedius. Fish. Sci. 2022, 88, 299–310. [Google Scholar] [CrossRef]
- Feng, W.P.; Nakabayashi, N.; Inomata, E.; Aoki, M.N.; Agatsuma, Y. Impacts of water temperature on the physiology and behaviours of the sea urchins Heliocidaris crassispina and Mesocentrotus nudus that reflect their range extension and disappearance in the Oga Peninsula, northern Honshu, Japan. Can. J. Fish. Aquat. Sci. 2020, 78, 580–588. [Google Scholar] [CrossRef]
- Wang, X.B.; Li, X.S.; Xiong, D.Q.; Chen, H.S.; Ren, H. Effects of stranded heavy fuel oil subacute exposure on the fitness-related traits of sea urchin Strongylocentrotus intermedius. Mar. Freshwater Res. 2022, 73, 754–761. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, L.L.; Shi, D.T.; Chi, X.M.; Yin, D.H.; Sun, J.N.; Ding, J.Y.; Yang, M.F.; Chang, Y.Q. Carryover effects of short-term UV-B radiation on fitness related traits of the sea urchin Strongylocentrotus intermedius. Ecotox. Environ. Safe. 2018, 164, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Zhang, L.S.; Zhao, C.; Feng, W.P.; Sun, P.; Chang, Y.Q. Correlation analyses of covering and righting behaviors to fitness related traits of the sea urchin Glyptocidaris crenularis in different environmental conditions. Chin. J. Oceanol. Limn. 2016, 34, 1183–1190. [Google Scholar] [CrossRef]
- Verling, E.; Crook, A.; Barnes, D. Covering behaviour in Paracentrotus lividus: Is light important? Mar. Biol. 2002, 140, 391–396. [Google Scholar] [CrossRef]
- Chi, X.; Shi, D.; Ma, Z.; Hu, F.Y.; Sun, J.N.; Huang, X.Y.; Zhang, L.S.; Chang, Y.Q.; Zhao, C. Carryover effects of long-term high water temperatures on fitness-related traits of the offspring of the sea urchin Strongylocentrotus intermedius. Mar. Environ. Res. 2021, 169, 105371. [Google Scholar] [CrossRef] [PubMed]
- Pawson, D.L.; Pawson, D.J. Bathyal sea urchins of the Bahamas, with notes on covering behavior in deep sea echinoids (Echinodermata: Echinoidea). Deep Sea Res. Part II 2013, 92, 207–213. [Google Scholar] [CrossRef]
- Backström, T.; Winberg, S. Serotonin coordinates responses to social stress-what we can learn from fish. Front. Neurosci. 2017, 11, 595. [Google Scholar] [CrossRef]
- Zhao, C.; Ji, N.J.; Tian, X.F.; Feng, W.P.; Sun, P.; Wei, J. Opsin4, Opsin5, and Pax6 significantly increase their expression in recently settled juveniles of the sea urchin Strongylocentrotus intermedius (Echinodermata: Echinoidea). Inver. Reprod. Dev. 2015, 5, 119–123. [Google Scholar] [CrossRef]
- Belleza, D.F.C.; Kawabata, Y.; Toda, T.; Gregory, N.N. Effects of dead conspecifics, hunger states, and seasons on the foraging behavior of the purple urchin Heliocidaris crassispina. Mar. Ecol. Prog. Ser. 2020, 664, 133–148. [Google Scholar] [CrossRef]
- Vowles, A.S.; Kemp, P.S. Artificial light at night (ALAN) affects the downstream movement behaviour of the critically endangered European eel, Anguilla anguilla. Environ. Pollut. 2021, 274, 116585. [Google Scholar] [CrossRef]
- Pulgar, J.; Manríquez, P.H.; Widdicombe, S.; García-Huidobro, R.; Quijón, P.A.; Carter, M.; Aldana, M.; Quintanilla-Ahumada, D.; Duarte, C. Artificial Light at Night (ALAN) causes size-dependent effects on intertidal fish decision-making. Mar. Pollut. Bull. 2023, 193, 115190. [Google Scholar] [CrossRef]
- Fobert, E.K.; Burke da Silva, K.; Swearer, S.E. Artificial light at night causes reproductive failure in clownfish. Biol. Lett. 2019, 15, 20190272. [Google Scholar] [CrossRef] [PubMed]
- Kupprat, F.; Kloas, W.; Krüger, A.; Schmalsch, C.; Hölker, F. Misbalance of thyroid hormones after two weeks of exposure to artificial light at night in Eurasian perch Perca fluviatilis. Conserv. Physiol. 2021, 9, coaa124. [Google Scholar] [CrossRef] [PubMed]
- Newman, R.C.; Ellis, T.; Davison, P.I.; Ives, M.J.; Thomas, R.J.; Griffiths, S.W.; Riley, W.D. Using novel methodologies to examine the impact of artificial light at night on the cortisol stress response in dispersing Atlantic salmon (Salmo salar L.) fry. Conserv. Physiol. 2015, 3, cov051. [Google Scholar] [CrossRef][Green Version]
- Wang, H.; Ding, J.; Ding, S.; Chang, Y. Transcriptome analysis to characterize the genes related to gonad growth and fatty acid metabolism in the sea urchin Strongylocentrotus intermedius. Genes Genom. 2019, 41, 1397–1415. [Google Scholar] [CrossRef] [PubMed]
- Terwilliger, D.P.; Buckley, K.M.; Brockton, V.; Ritter, N.J.; Smith, L.C. Distinctive expression patterns of 185/333 genes in the purple sea urchin, Strongylocentrotus purpuratus: An unexpectedly diverse family of transcripts in response to LPS, beta-1,3-glucan, and dsRNA. BMC Mol. Biol. 2007, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhang, Z.; Guo, H.; Qin, J.; Zhang, X. Changes in aggressive behavior, cortisol and brain monoamines during the formation of social hierarchy in black rockfish (Sebastes schlegelii). Animals 2020, 10, 2357. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Sun, X.; Bai, Q.; Zhang, Y.; Qin, J.; Zhang, X. Molecular identification of an androgen receptor and the influence of long-term aggressive interaction on hypothalamic genes expression in black rockfish (Sebastes schlegelii). J. Comp. Physiol. A 2021, 207, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Guo, H.; Zhang, Z.; Wang, Y.; Qin, J.; Zhang, X. Impact of pre-aggressive experience on behavior and physiology of black rockfish (Sebastes schlegelii). Aquaculture 2021, 536, 736416. [Google Scholar] [CrossRef]
- Zhang, Z.; Bai, Q.; Xu, X.; Zhang, X. Effects of the dominance hierarchy on social interactions, cortisol level, HPG-axis activities and reproductive success in the golden cuttlefish Sepia esculent. Aquaculture 2020, 533, 736059. [Google Scholar] [CrossRef]
- Yang, M.; Yu, Y.; Ding, J.; Sun, J.; Hu, F.; Chang, Y.; Zhao, C. Effects of light intensity on Opsin4, Opsin5, and Pax6 expressions of the sea urchin Strongylocentrotus intermedius. Mar. Ecol. 2020, 41, e12593. [Google Scholar] [CrossRef]
- Parra, M.; Rubilar, T.; Latorre, M.P.; Parra, M.; Rubilar, T.; Latorre, M.P.; Ephedra, L.; Gil, D.G.; Díaz de Vivar, M.E. Nutrient allocation in the gonads of the sea urchin Arbacia dufresnii in different stages of gonadal development. Invertebr. Reprod. Dev. 2015, 59, 26–36. [Google Scholar] [CrossRef]
- Machado, A.M.; Fernández-Boo, S.; Nande, M.; Pinto, R.; Costas, B.; Castro, L.F. The male and female gonad transcriptome of the edible sea urchin, Paracentrotus lividus: Identification of sex-related and lipid biosynthesis genes. Aquacult. Rep. 2022, 22, 100936. [Google Scholar] [CrossRef]
- Zhang, B.; Yang, J.W.; Han, T.; Huang, D.X.; Zhao, Z.H.; Feng, J.Q.; Zhou, N.M.; Xie, H.Q.; Wang, T.M. Identification and characterization of a novel 5-hydroxytryptamine receptor in the sea cucumber Apostichopus japonicus (Selenka). J. Exp. Zool. Part A 2021, 335, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Tuxbury, S.M.; Salmon, M. Competitive interactions between artificial lighting and natural cues during seafinding by hatchling marine turtles. Biol. Conserv. 2005, 121, 311–316. [Google Scholar] [CrossRef]
- La Sorte, F.A.; Fink, D.; Butler, J.J.; Farnsworth, A.; Cabrera-Cruz, S.A. Seasonal associations with urban light pollution for nocturnally migrating bird populations. Glob. Change Biol. 2017, 23, 4609–4619. [Google Scholar] [CrossRef]
- Van Doren, B.M.; Horton, K.G.; Dokter, A.M.; Klinck, H.; Elbin, S.B.; Farnsworth, A. High-intensity urban light installation dramatically alters nocturnal bird migration. Proc. Natl. Acad. Sci. USA 2017, 114, 11175–11180. [Google Scholar] [CrossRef]
- Burt, C.S.; Kelly, J.F.; Fox, A.S.; Jenkins-Smith, H.C.; Leon-Corwin, M.; Khalighifar, A.; Trankina, G.E.; Silva, C.L.; Horton, K.G. Can ecological forecasting lead to convergence on sustainable lighting policies? Conserv. Sci. Pract. 2023, 5, e12920. [Google Scholar] [CrossRef]
- Orvain, O.; Sauriau, P.G. Environmental and behavioural factors affecting activity in the intertidal gastropod Hydrobia ulvae. J. Exp. Mar. Biol. Ecol. 2002, 272, 191–216. [Google Scholar] [CrossRef]
- Morandini, L.; Ramallo, M.R.; Scaia, M.F.; Höcht, C.; Somoza, G.M.; Pandolfi, M. Dietary L-tryptophan modulates agonistic behavior and brain serotonin in male dyadic contests of a cichlid fish. J. Comp. Physiol. A 2019, 205, 867–880. [Google Scholar] [CrossRef]
- Luarte, T.; Bonta, C.C.; Silva-Rodriguez, E.A.; Quijón, P.A.; Miranda, C.; Fariasm, A.A.; Duarte, C. Light pollution reduces activity, food consumption and growth rates in a sandy beach invertebrate. Environ. Pollut. 2016, 218, 1147–1153. [Google Scholar] [CrossRef]
- Hussein, A.A.A.; Baz, E.S.; Mariën, J.; Tadros, M.M.; El-Shenawy, N.S.; Koene, J.M. Effect of photoperiod and light intensity on learning ability and memory formation of the pond snail Lymnaea stagnalis. Invert. Neurosci. 2020, 20, 18. [Google Scholar] [CrossRef]
- Boldogh, S.; Dobrosi, D.; Samu, P. The effects of the illumination of buildings on house-dwelling bats and its conservation consequences. Acta. Chiropterol. 2007, 9, 527–534. [Google Scholar] [CrossRef]
- Silvia, L.; Manuel, T.S. Dissecting the roles of gonadotropin-inhibitory hormone in mammals: Studies using pharmacological tools and genetically modified mouse models. Front. Endocrinol. 2015, 6, 189. [Google Scholar] [CrossRef]
- Brüning, A.; Hölker, F.; Franke, S.; Kleiner, W.; Kloas, W. Impact of different colours of artificial light at night on melatonin rhythm and gene expression of gonadotropins in European perch. Sci. Total. Environ. 2016, 543, 214–222. [Google Scholar] [CrossRef]
- Brüning, A.; Kloas, W.; Preuer, T.; Hölker, F. Influence of artificially induced light pollution on the hormone system of two common fish species, perch and roach, in a rural habitat. Conserv. Physiol. 2018, 6, coy016. [Google Scholar] [CrossRef]
- Raible, F.; Tessmar-Raible, K.; Arboleda, E.; Kaller, T.; Bork, P.; Arendt, D.; Arnone, M.I. Opsins and clusters of sensory G-protein-coupled receptors in the sea urchin genome. Dev. Biol. 2006, 300, 461–475. [Google Scholar] [CrossRef] [PubMed]
- Ullrich-Luter, E.M.; Dupont, S.; Arboleda, E.; Hausen, H.; Arnone, M.I. Unique system of photoreceptors in sea urchin tube feet. Proc. Natl. Acad. Sci. USA 2011, 108, 8367–8372. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Ji, N.; Sun, P.; Feng, W.; Wei, J.; Chang, Y. Effects of light and covering behavior on PAX6 expression in the sea urchin Strongylocentrotus intermedius. PLoS ONE 2014, 9, e110895. [Google Scholar] [CrossRef] [PubMed]
- Pulgar, J.; Zeballos, D.; Vargas, J.; Aldana, M.; Manriquez, P.H.; Manriquez, K.; Quijón, P.A.; Widdicombe, S.; Anguita, C.; Quintanilla, D.; et al. Endogenous cycles, activity patterns and energy expenditure of an intertidal fish is modified by artificial light pollution at night (ALAN). Environ. Pollut. 2019, 244, 361–366. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).