
Genetic Relationship between Salmonella Isolates Recovered from Calves and Broilers Chickens in Kafr El-Sheikh City Using ERIC PCR
 and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples Collection
2.2. Biochemical Identification
2.3. Serological Identification
2.4. Antimicrobial Sensitivity Testing (Phenotypic Resistance)
2.5. Molecular Identification of B-Lactamase Encoding Genes, Class 1 Integron, and ERIC PCR of Isolated Salmonella
3. Results
3.1. The Prevalence of Salmonella Isolated from Different Spp.
3.2. Serological Identification of Salmonella Isolates
3.3. The Antimicrobial Resistance
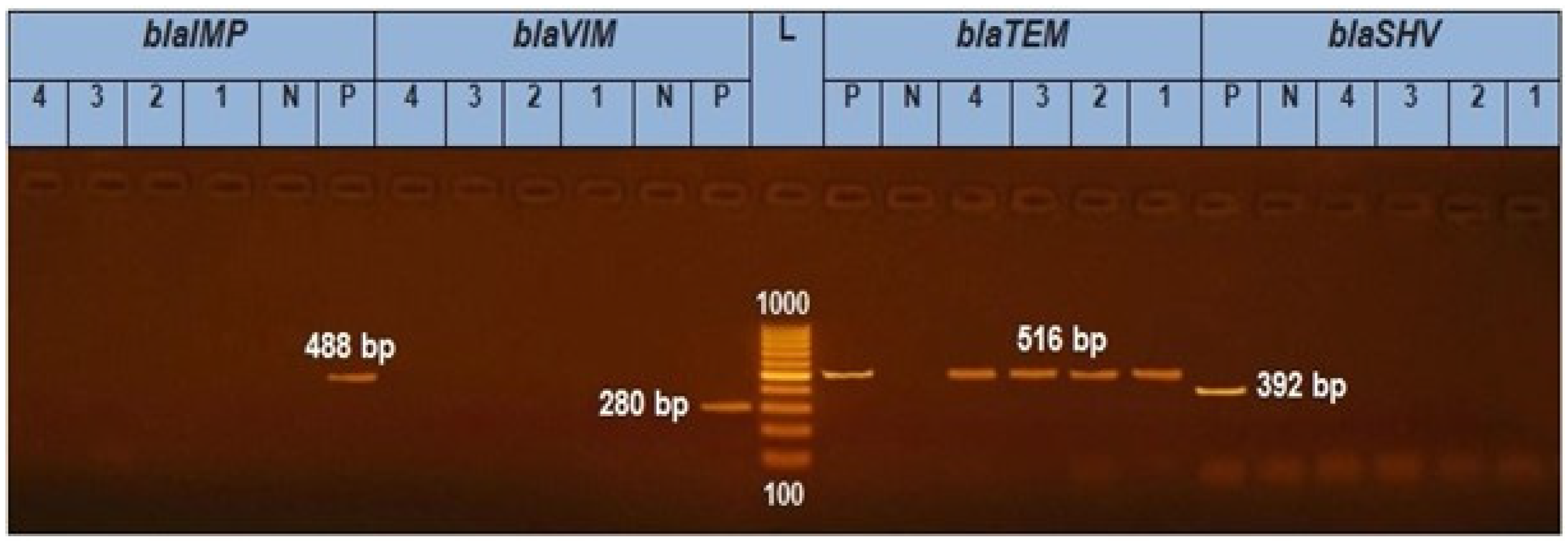
3.4. Prevalence of β-Lactamase-Encoding Genes in Salmonella Isolation
3.5. Prevalence of Class 1 Integrons for MDR Salmonella
3.6. ERIC PCR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodriguez-Rivera, L.; Wright, E.; Siler, J.; Elton, M.; Cummings, K.; Warnick, L.; Wiedmann, M. Subtype analysis of Salmonella isolated from subclinically infected dairy cattle and dairy farm environments reveals the presence of both human-and bovine-associated subtypes. Vet. Microbiol. 2014, 170, 307–316. [Google Scholar] [CrossRef]
- Kebede, A.; Kemal, J.; Alemayehu, H.; Habte Mariam, S. Isolation, identification, and antibiotic susceptibility testing of Salmonella from slaughtered bovines and ovines in Addis Ababa Abattoir Enterprise, Ethiopia: A cross-sectional study. Int. J. Bacteriol. 2016, 2016, 714785. [Google Scholar] [CrossRef]
- Vandeplas, S.; Dauphin, R.D.; Beckers, Y.; Thonart, P.; Thewis, A. Salmonella in chicken: Current and developing strategies to reduce contamination at farm level. J. Food Prot. 2010, 73, 774–785. [Google Scholar] [CrossRef] [PubMed]
- Habing, G.G.; Lombard, J.E.; Kopral, C.A.; Dargatz, D.A.; Kaneene, J.B. Farm-level associations with the shedding of Salmonella and antimicrobial-resistant Salmonella in US dairy cattle. Foodborne Pathog. Dis. 2012, 9, 815–821. [Google Scholar] [CrossRef]
- Huston, C.L.; Wittum, T.E.; Love, B.C.; Keen, J.E. Prevalence of fecal shedding of Salmonella spp in dairy herds. J. Am. Vet. Med. Assoc. 2002, 220, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Cliver, D.O.; Riemann, H.P. Foodborne Infections and Intoxications; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Ahmed, A.M.; Motoi, Y.; Sato, M.; Maruyama, A.; Watanabe, H.; Fukumoto, Y.; Shimamoto, T. Zoo animals as reservoirs of gram-negative bacteria harboring integrons and antimicrobial resistance genes. Appl. Environ. Microbiol. 2007, 73, 6686–6690. [Google Scholar] [CrossRef]
- Roy Chowdhury, P.; Ingold, A.; Vanegas, N.; Martínez, E.; Merlino, J.; Merkier, A.K.; Castro, M.; González Rocha, G.; Borthagaray, G.; Centrón, D. Dissemination of multiple drug resistance genes by class 1 integrons in Klebsiella pneumoniae isolates from four countries: A comparative study. Antimicrob. Agents Chemother. 2011, 55, 3140–3149. [Google Scholar] [CrossRef] [PubMed]
- Betteridge, T.; Partridge, S.R.; Iredell, J.R.; Stokes, H. Genetic context and structural diversity of class 1 integrons from human commensal bacteria in a hospital intensive care unit. Antimicrob. Agents Chemother. 2011, 55, 3939–3943. [Google Scholar] [CrossRef] [PubMed]
- Stalder, T.; Barraud, O.; Casellas, M.; Dagot, C.; Ploy, M.-C. Integron involvement in environmental spread of antibiotic resistance. Front. Microbiol. 2012, 3, 119. [Google Scholar] [CrossRef]
- Bonnet, R. Growing group of extended-spectrum β-lactamases: The CTX-M enzymes. Antimicrob. Agents Chemother. 2004, 48, 1–14. [Google Scholar] [CrossRef]
- Jacoby, G.A.; Sutton, L. Properties of plasmids responsible for production of extended-spectrum beta-lactamases. Antimicrob. Agents Chemother. 1991, 35, 164–169. [Google Scholar] [CrossRef]
- Villa, L.; Pezzella, C.; Tosini, F.; Visca, P.; Petrucca, A.; Carattoli, A. Multiple-antibiotic resistance mediated by structurally related IncL/M plasmids carrying an extended-spectrum β-lactamase gene and a class 1 integron. Antimicrob. Agents Chemother. 2000, 44, 2911–2914. [Google Scholar] [CrossRef]
- Cantón, R.; Coque, T.M.; Baquero, F. Multi-resistant Gram-negative bacilli: From epidemics to endemics. Curr. Opin. Infect. Dis. 2003, 16, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Fagbamila, I.O.; Barco, L.; Mancin, M.; Kwaga, J.; Ngulukun, S.S.; Zavagnin, P.; Lettini, A.A.; Lorenzetto, M.; Abdu, P.A.; Kabir, J. Salmonella serovars and their distribution in Nigerian commercial chicken layer farms. PLoS ONE 2017, 12, e0173097. [Google Scholar] [CrossRef] [PubMed]
- Quinn, P.; Markey, B.K.; Carter, M.; Donnelly, W.; Leonard, F. Veterinary Microbiology and Microbial Disease; Blackwell Science: Oxford, UK, 2002. [Google Scholar]
- Krieg, N.; Holt, J.; Murray, R. Bergey’s Manual of Systematic Bacteriology; Williams and Wilkins: Baltimore, MD, USA, 1984; Volume 1, pp. 331–371. [Google Scholar]
- Topley, W.W.; Wilson, G.S.; Coller, L.; Sussman, M. Topley & Wilson’s Microbiology and Microbial Infections; John Wiley & Sons, Ltd.: New York, NY, USA, 1998. [Google Scholar]
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie Van Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef]
- Kauffmann, F. Das Kauffmann-White-Schema. In Ergebnisse der Mikrobiologie, Immunitätsforschung und Experimentellen Therapie; Springer: Berlin, Germany, 1957; pp. 160–216. [Google Scholar]
- Versalovic, J.; Koeuth, T.; and Lupski, R. Distribution of repetitive DNA sequences in eubacteria and application to finerpriting of bacterial enomes. Nucleic Acids Res. 1991, 19, 6823–6831. [Google Scholar] [CrossRef] [PubMed]
- White, P.A.; McIver, C.J.; Deng, Y.; Rawlinson, W.D. Characterisation of two new gene cassettes, aadA5 and dfrA17. FEMS Microbiol. Lett. 2000, 182, 265–269. [Google Scholar] [CrossRef]
- Gassama Sow, A.; Aïdara-Kane, A.; Barraud, O.; Gatet, M.; Denis, F.; Ploy, M.C. High prevalence of trimethoprim-resistance cassettes in class 1 and 2 integrons in Senegalese Shigella spp isolates. J. Infect. Dev. Ctries. 2010, 4, 207–212. [Google Scholar] [CrossRef]
- Xia, Y.; Liang, Z.; Su, X.; Xiong, Y. Characterization of carbapenemase genes in Enterobacteriaceae species exhibiting decreased susceptibility to carbapenems in a university hospital in Chongqing, China. Ann. Lab. Med. 2012, 32, 270–275. [Google Scholar] [CrossRef]
- Colom, K.; Pérez, J.; Alonso, R.; Fernández-Aranguiz, A.; Lariño, E.; Cisterna, R. Simple and reliable multiplex PCR assay for detection of bla TEM, bla SHV and bla OXA–1 genes in Enterobacteriaceae. FEMS Microbiol. Lett. 2003, 223, 147–151. [Google Scholar] [CrossRef]
- Carpenter, K.E.; Smith-Vaniz, W.F.; de Bruyne, G.; de Morais, L. Dicentrarchus punctatus. IUCN Red List of Threatened Species. 2015. Available online: https://www.iucnredlist.org/species/198671/21913001 (accessed on 21 May 2022).
- Beyene, T. Identification and antimicrobial susceptibility profile of Salmonella isolated from selected dairy farms, abattoir and humans at Asella town, Ethiopia. J. Vet. Sci. Technol. 2016, 7, 320. [Google Scholar] [CrossRef]
- Saroj, S.; Shashidhar, R.; Bandekar, J. Gamma radiation used as hygienization technique for foods does not induce viable but non-culturable state (VBNC) in Salmonella enterica subsp. enterica serovar Typhimurium. Curr. Microbiol. 2009, 59, 420–424. [Google Scholar] [CrossRef]
- Kagambèga, A.; Lienemann, T.; Aulu, L.; Traoré, A.; Barro, N.; Siitonen, A.; Haukka, K. Prevalence and characterization of Salmonella enterica from the feces of cattle, poultry, swine and hedgehogs in Burkina Faso and their comparison to human Salmonellaisolates. BMC Microbiol. 2013, 13, 1–9. [Google Scholar] [CrossRef]
- Le Bouquin, S.; Allain, V.; Rouxel, S.; Petetin, I.; Picherot, M.; Michel, V.; Chemaly, M. Prevalence and risk factors for Salmonella spp. contamination in French broiler-chicken flocks at the end of the rearing period. Prev. Vet. Med. 2010, 97, 245–251. [Google Scholar] [CrossRef]
- Aragaw, K.; Terefe, L.; Abera, M. Prevalence of Salmonella infection in intensive poultry farms in Hawassa and isolation of Salmonella species from sick and dead chickens. Ethiop. Vet. J. 2010, 14, 115–124. [Google Scholar] [CrossRef]
- Awad, A.; Gwida, M.; Khalifa, E.; Sadat, A. Phenotypes, antibacterial-resistant profile, and virulence-associated genes of Salmonella serovars isolated from retail chicken meat in Egypt. Vet. World 2020, 13, 440. [Google Scholar] [CrossRef]
- Ball, T.; Monte, D.; Aidara-Kane, A.; Matheu, J.; Ru, H.; Thakur, S.; Ejobi, F.; Fedorka-Cray, P. International lineages of Salmonella enterica serovars isolated from chicken farms, Wakiso District, Uganda. PLoS ONE 2020, 15, e0220484. [Google Scholar] [CrossRef] [PubMed]
- Gu, D.; Wang, Z.; Tian, Y.; Kang, X.; Meng, C.; Chen, X.; Pan, Z.; Jiao, X. Prevalence of Salmonella isolates and their distribution based on whole-genome sequence in a chicken slaughterhouse in Jiangsu, China. Front. Vet. Sci. 2020, 7, 29. [Google Scholar] [CrossRef]
- Cetin, E.; Temelli, S.; Eyigor, A. Salmonella prevalence and serovar distribution in healthy slaughter sheep and cattle determined by ISO 6579 and VIDAS UP Salmonella methods. J. Food Sci. Technol. 2019, 56, 5317–5325. [Google Scholar] [CrossRef] [PubMed]
- Halawa, M.; Moawad, A.; Eldesouky, I.; Ramadan, H. Detection of antimicrobial phenotypes, ß-lactamase encoding genes and class L INTEGRONS in Salmonella serovars isolated from broilers. Int. J. Poult. Sci. 2016, 15, 1–7. [Google Scholar] [CrossRef][Green Version]
- Yang, X.; Huang, J.; Zhang, Y.; Liu, S.; Chen, L.; Xiao, C.; Zeng, H.; Wei, X.; Gu, Q.; Li, Y.; et al. Prevalence, abundance, serovars and antimicrobial resistance of Salmonella isolated from retail raw poultry meat in China. Sci. Total Environ. 2020, 713, 136385. [Google Scholar] [CrossRef]
- Ahmed, A.M.; Shimamoto, T. Genetic analysis of multiple antimicrobial resistance in Salmonella isolated from diseased broilers in Egypt. Microbiol. Immunol. 2012, 56, 254–261. [Google Scholar] [CrossRef]
- Mihaiu, L.; Lapusan, A.; Tanasuica, R.; Sobolu, R.; Mihaiu, R.; Oniga, O.; Mihaiu, M. First study of Salmonella in meat in Romania. J. Infect. Dev. Ctries. 2014, 8, 050–058. [Google Scholar] [CrossRef]
- Zhang, L.; Fu, Y.; Xiong, Z.; Ma, Y.; Wei, Y.; Qu, X.; Zhang, H.; Zhang, J.; Liao, M. Highly prevalent multidrug-resistant Salmonella from chicken and pork meat at retail markets in Guangdong, China. Front. Microbiol. 2018, 9, 2104. [Google Scholar] [CrossRef]
- Bhatta, D.; Bangtrakulnonth, A.; Tishyadhigama, P.; Saroj, S.; Bandekar, J.; Hendriksen, R.; Kapadnis., B. Serotyping, PCR, phage-typing and antibiotic sensitivity testing of Salmonella serovars isolated from urban drinking water supply systems of Nepal. Lett. Appl. Microbiol. 2007, 44, 588–594. [Google Scholar] [CrossRef]
- Carattoli, A. Plasmid-mediated antimicrobial resistance in Salmonella enterica. Curr. Issues Mol. Biol. 2003, 5, 113–122. [Google Scholar]
- Domingues, S.; da Silva, G.; Nielsen, K. Integrons: Vehicles and pathways for horizontal dissemination in bacteria. Mob. Genet. Elem. 2012, 2, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, S.; Fatima, J.; Shakil, S.; Rizvi, S.; Kamal, M. Antibiotic resistance and extended spectrum beta-lactamases: Types, epidemiology and treatment. Saudi J. Biol. Sci. 2015, 22, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Bradford, P. Extended-spectrum β-lactamases in the 21st century: Characterization, epidemiology, and detection of this important resistance threat. Clin. Microbiol. Rev. 2001, 14, 933–951. [Google Scholar] [CrossRef]
- Wu, H.; Wang, Y.; Wu, Y.; Qiao, J.; Li, H.; Zheng, S.; Xia, X.; Cui, S.; Wang, X.; Xi, M.; et al. Emergence of β-lactamases and extended-spectrum β-lactamases (ESBLs) producing Salmonella in retail raw chicken in China. Foodborne Pathog. Dis. 2015, 12, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.; Hungerford, L.; Fedorka-Cray, P.; Headrick, M. Extended-spectrum-cephalosporin resistance in Salmonella enterica isolates of animal origin. Antimicrob. Agents Chemother. 2004, 48, 3179–3181. [Google Scholar] [CrossRef] [PubMed]
- Seiffert, S.; Perreten, V.; Johannes, S.; Droz, S.; Bodmer, T.; Endimiani, A. OXA-48 carbapenemase-producing Salmonella enterica serovar Kentucky isolate of sequence type 198 in a patient transferred from Libya to Switzerland. Antimicrob. Agents Chemother. 2014, 58, 2446–2449. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, Q.; Zhao, C.; Zhang, Y.; Li, L.; Qi, J.; Luo, Y.; Zhou, D.; Liu, Y. Prevalence and antimicrobial resistance of Salmonella isolated from broilers in Shandong, China. Res. Sq. 2020. [Google Scholar] [CrossRef]
- Moawad, A.A.; Hotzel, H.; Awad, O.; Tomaso, H.; Neubauer, H.; Hafez, H.M.; El-Adawy, H. Occurrence of Salmonella enterica and Escherichia coli in raw chicken and beef meat in northern Egypt and dissemination of their antibiotic resistance markers. Gut Pathog. 2017, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Nath, G.; Maurya, P.; Gulati, A. ERIC PCR and RAPD based fingerprinting of Salmonella Typhi strains isolated over a period of two decades. Infect. Genet. Evol. 2010, 10, 530–536. [Google Scholar] [CrossRef]
- de Souza, A.; de Freitas Neto, O.; Batista, D.; Estupinan, A.; de Almeida, A.; Barrow, P.; Berchieri, A. ERIC-PCR genotyping of field isolates of Salmonella enterica subsp. enterica serovar Gallinarum biovars Gallinarum and Pullorum. Avian Pathol. 2015, 44, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Ali-Shiroodi, A.; Jamshidian, M.; Salehi, T.; Boroujeni, G.; Amini, K. Genotyping of Salmonella enterica subsp. enterica serovar Entritidis, isolated from poultry, cattle and human in Iran by ERIC-PCR. Int. J. Biosci. 2014, 5, 147–153. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Spp. | Type of Sample | Age | Number |
|---|---|---|---|
| Calves | Rectal swabs and feces | First month | 40 |
| 1–2 month | 30 | ||
| 2–3 month | 30 | ||
| Chickens | Native breeds | One day until 45 days old | 40 |
| Cross breeds | 30 | ||
| Foreign breeds | 30 | ||
| Total | 200 | ||
| Target Gene | Primers Sequences | Amplified Segment (bp) | Primary Denaturation | Amplification (35 Cycles) | Final Extension | Reference | ||
|---|---|---|---|---|---|---|---|---|
| Secondary Denaturation | Annealing | Extension | ||||||
| ERIC | ATGTAAGCT CCTGGGGAT TCAC | Variable | 94 °C 5 min | 94 °C 30 s | 52 °C 40 s | 72 °C 1 min | 72 °C 12 min | [21] |
| AAGTAAGTG ACTGGGGTG AGCG | ||||||||
| Integrase gene (hep 35 and hep 36 primers) | TGCGGGTYAARGATBTKGATTT | 491 | 55 °C 40 s | 72 °C 45 s | 72 °C 10 min | [22] | ||
| CARCACATGCGTRTARAT | ||||||||
| class 1 integron cassettes(5’-CS and 3’-CS) | GGCATCCAA GCAGCAAG | Variable | 55 °C 40 s | 72 °C 45 s | 72 °C 10 min | [23] | ||
| AAGCAGACT TGACCTGA | ||||||||
| BlaIMP | CATGGTTTGGTGGTTCTTGT | 488 | 53 °C 40 s | 72 °C 45 s | 72 °C 10 min | [24] | ||
| ATAATTTGGCGGACTTTGGC | ||||||||
| blaVIM | AGTGGTGAGTATCCGACA | 280 | 53 °C 30 s | 72 °C 30 s | 72 °C 10 min | |||
| ATGAAAGTGCGTGGAGAC | ||||||||
| BlaTEM | ATCAGCAATAAACCAGC | 516 | 54 °C 40 s | 72 °C 45 s | 72 °C 10 min | [25] | ||
| CCCCGAAGAACGTTTTC | ||||||||
| blaSHV | AGGATTGACTGCCTTTTTG | 392 | 54 °C 40 s | 72 °C 40 s | 72 °C 10 min | |||
| ATTTGCTGATTTCGCTCG | ||||||||
| Animal Spp. | Type of Sample | Age | Number of Samples | Positive % |
|---|---|---|---|---|
| Calves | Rectal swabs and feces | First month | 40 | 4 |
| 1–2 month | 30 | 3 | ||
| 2–3 month | 30 | 1 | ||
| Total | 100 | 8% | ||
| Chickens | Native | One day until 45 days old | 40 | 1 |
| Cross breed | 30 | 0 | ||
| Foreign | 30 | 1 | ||
| Total | 100 | 2% | ||
| Serotype | Number | % | O Antigen | H Antigen | |
|---|---|---|---|---|---|
| Phase 1 | Phase 11 | ||||
| Salmonella Kentucky | 3 | 30% | 8, 20 | I | Z6 |
| Salmonella Typhimurium | 3 | 30% | 1, 4, [26], 12 | I | 1, 2 |
| Salmonella Enteritidis | 4 | 40% | 1, 9, 12 | g, m | - |
| Types of Flock | Salmonella Positive | The Isolated Serotypes | |||
|---|---|---|---|---|---|
| Salmonella Serotypes | Number of Serotypes | Proportion/Source | Total Proportion | ||
| Diseased poultry | 8 | Salmonella Kentucky | 2 | 25% | 20% |
| SalmonellaTyphimurium | 3 | 37.50% | 30% | ||
| Salmonella Enteritidis | 3 | 37.50% | 30% | ||
| Diseased calves | 2 | Salmonella Kentucky | 1 | 50% | 10% |
| Salmonella Enteritidis | 1 | 50% | 10% | ||
| Antimicrobial Class | Antimicrobial Agent(s) Tested | Salmonella Isolates | ||
|---|---|---|---|---|
| Resistant No. (%) | Intermediate No. (%) | Sensitive No. (%) | ||
| Quinolones | Nalidixic acid (NAL) | 9 (90) | 1 (10) | 0 (0) |
| Aminoglycosides | Streptomycin (STR) | 6 (60) | 4 (40) | 0 (0) |
| Kanamycin (KAN) | 4 (40) | 3 (30) | 3 (30) | |
| Cephalosporins | Ceftriaxone (CRO) | 0 (0) | 4 (40) | 6 (60) |
| Cefotaxime (CTX) | 1 (10) | 6 (60) | 3 (30) | |
| Penicillins | Amoxicillin (AMC) | 8 (80) | 2 (20) | 0 (0) |
| Ampicillin (AMP) | 9 (90) | 1 (10) | 0 (0) | |
| Phenicols | Chloramphenicol (C) | 1 (10) | 3 (30) | 6 (60) |
| Macrolides | Erthyromycin (E) | 10 (100) | 0 (00) | 0 (0) |
| Vancomycin (VA) | 9 (90) | 1 (10) | 0 (0) | |
| Tetracycline | Tetracycline (TE) | 4 (40) | 4 (40) | 2(20) |
| Fluoroquinolones | Ciprofloxacin (CIP) | 1 (10) | 4 (40) | 5 (50) |
| Enrofloxacin (ENR) | 2 (20) | 4 (40) | 4 (40) | |
| Sulphonamides | Trimethoprim/sulfamethoxazole (SXT) | 4 (40) | 5 (50) | 1 (10) |
| Number of Intersecting Elements | Jaccard/Tanimoto Coefficient | |
|---|---|---|
| Samples 3, 4 | 2 | 1 |
| Samples 1, 2 | 3 | 0.33 |
| Samples 1, 3 | 1 | 0.17 |
| Samples 1, 4 | 1 | 0.17 |
| Samples 2, 3 | 1 | 0.13 |
| Samples 2, 4 | 1 | 0.13 |
| Stage | Cluster Combined | Coefficients | Stage Cluster First Appears | Next Stage | ||
|---|---|---|---|---|---|---|
| Cluster 1 | Cluster 2 | Cluster 1 | Cluster 2 | |||
| 1 | 3 | 4 | 0.000 | 0 | 0 | 3 |
| 2 | 1 | 2 | 3.000 | 0 | 0 | 3 |
| 3 | 1 | 3 | 7.500 | 2 | 1 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tawfik, R.G.; Gawish, M.F.; Abotaleb, M.M.; Nada, H.S.; Morsy, K.; Abumandour, M.M.A.; Torky, H. Genetic Relationship between Salmonella Isolates Recovered from Calves and Broilers Chickens in Kafr El-Sheikh City Using ERIC PCR. Animals 2022, 12, 3428. https://doi.org/10.3390/ani12233428
Tawfik RG, Gawish MF, Abotaleb MM, Nada HS, Morsy K, Abumandour MMA, Torky H. Genetic Relationship between Salmonella Isolates Recovered from Calves and Broilers Chickens in Kafr El-Sheikh City Using ERIC PCR. Animals. 2022; 12(23):3428. https://doi.org/10.3390/ani12233428
Chicago/Turabian StyleTawfik, Rasha Gomaa, Mahmoud F. Gawish, Mahmoud M. Abotaleb, Hassan S. Nada, Kareem Morsy, Mohamed M. A. Abumandour, and Helmy Torky. 2022. "Genetic Relationship between Salmonella Isolates Recovered from Calves and Broilers Chickens in Kafr El-Sheikh City Using ERIC PCR" Animals 12, no. 23: 3428. https://doi.org/10.3390/ani12233428
APA StyleTawfik, R. G., Gawish, M. F., Abotaleb, M. M., Nada, H. S., Morsy, K., Abumandour, M. M. A., & Torky, H. (2022). Genetic Relationship between Salmonella Isolates Recovered from Calves and Broilers Chickens in Kafr El-Sheikh City Using ERIC PCR. Animals, 12(23), 3428. https://doi.org/10.3390/ani12233428