
Fishmeal Dietary Replacement Up to 50%: A Comparative Study of Two Insect Meals for Rainbow Trout (Oncorhynchus mykiss)

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Experimental Animals and Rearing Conditions
2.3. Growth Trial and Samples Collection
2.4. Histomorphology
2.4.1. Samples Processing
2.4.2. Distal Intestine and Pyloric Caeca Histomorphology Analyses
2.4.3. Liver Histomorphology Analysis
2.5. Analytical Determinations
2.5.1. Chemical Analyses
2.5.2. Digestive Enzymes Determination
2.5.3. Liver Intermediary Metabolism
2.5.4. Non-Specific Immune Status
Plasma Immunological Determinations
TNF-α Detection in Distal Intestine and Skin Mucus
2.5.5. Liver Antioxidant Status and Fish Welfare Indicators
2.6. Statistical Analysis
3. Results and Discussion
3.1. Growth Performance
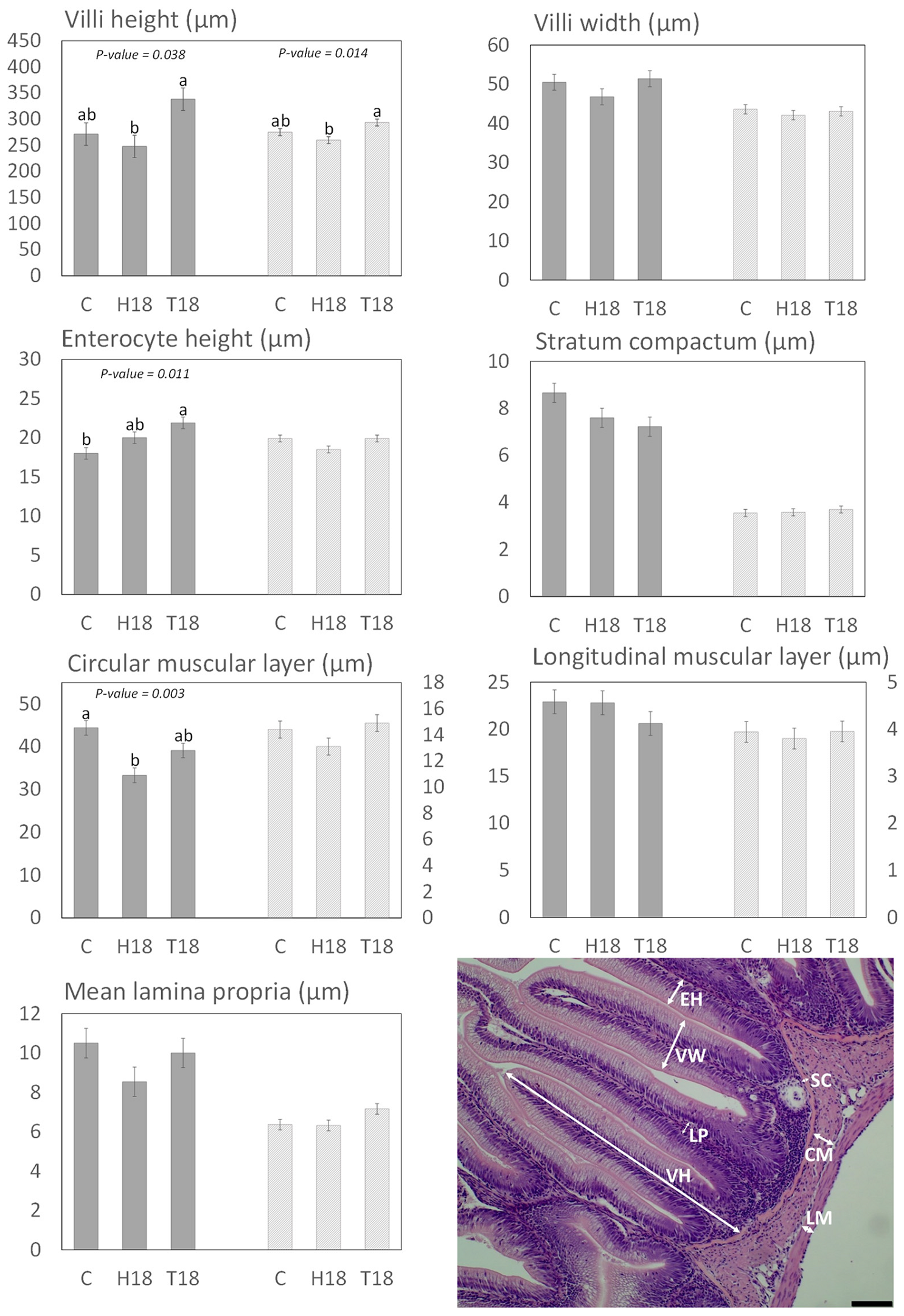
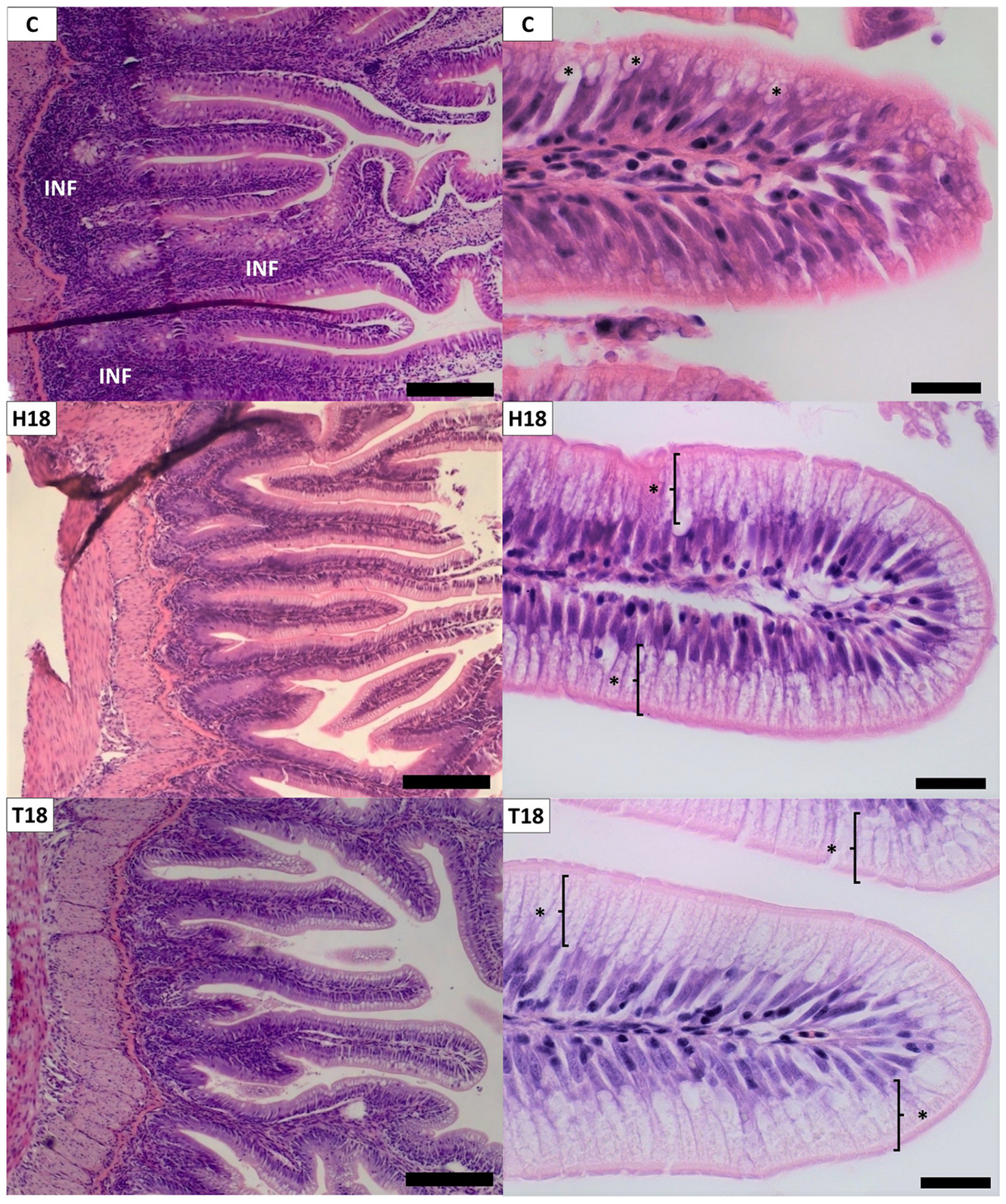
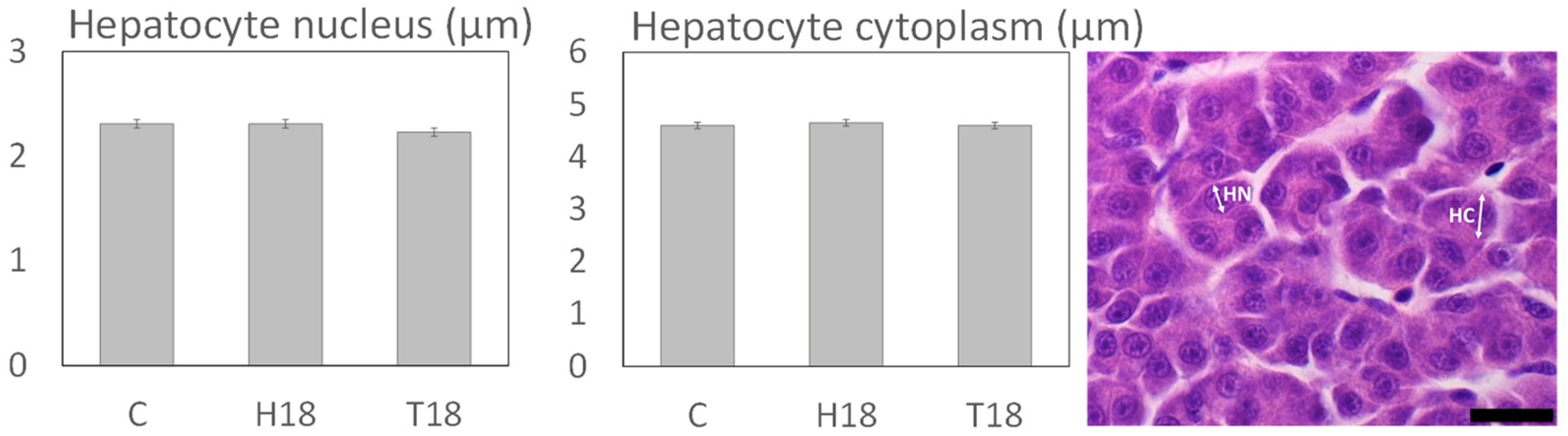
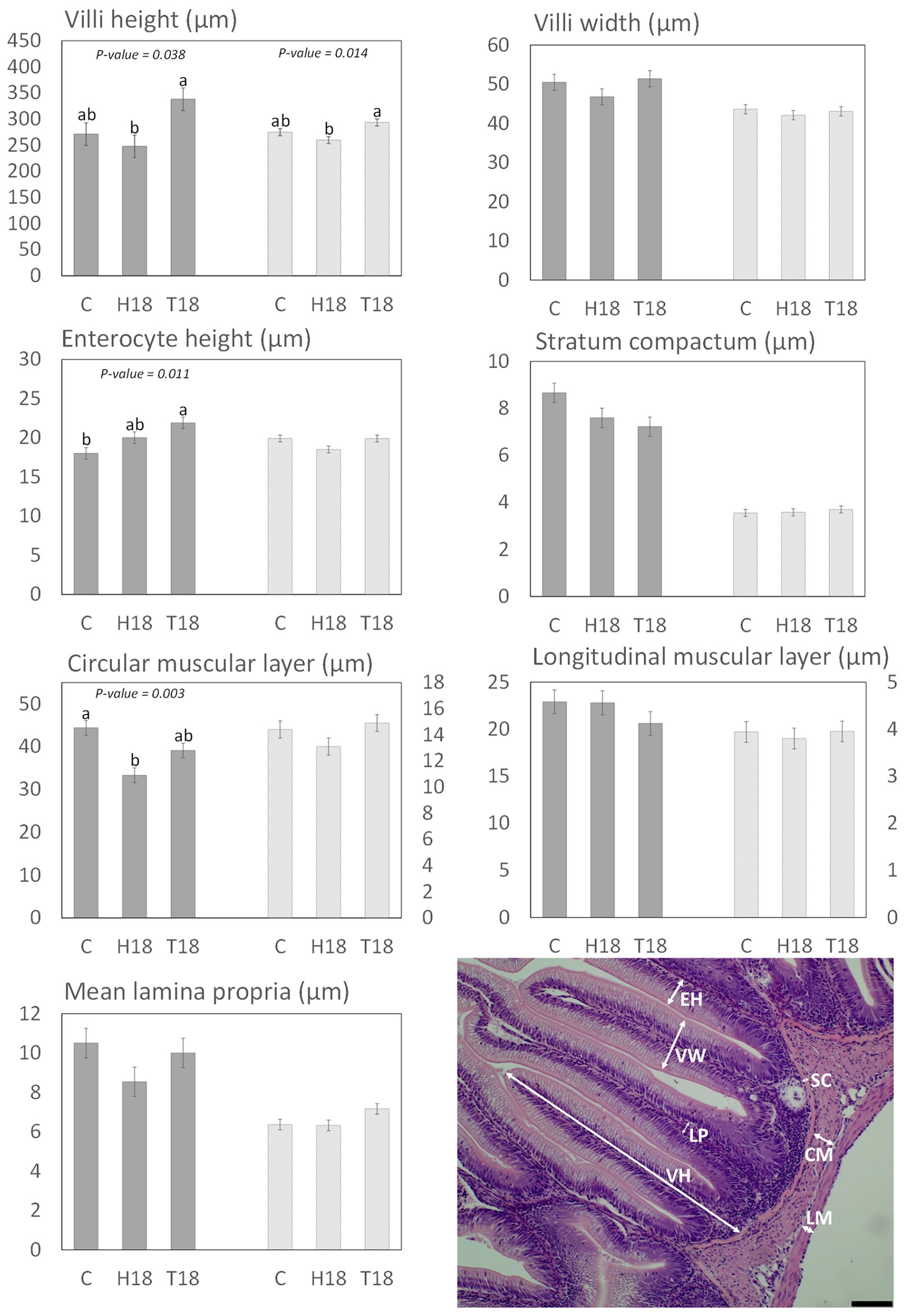
3.2. Histomorphology
3.2.1. Distal Intestine and Pyloric Caeca
3.2.2. Liver
3.3. Digestive Enzymes
3.4. Liver Intermediary Metabolism
3.5. Non-Specific Immune Status
3.6. Liver Antioxidant Status and Fish Welfare Indicators
3.7. Proximate Composition of the Fillet
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations, Department of Economic and Social Affairs, Population Division. World Population Prospects 2019: Highlights; United Nations: New York, NY, USA, 2019; p. 40. [Google Scholar]
- Fry, J.P.; A Mailloux, N.; Love, D.C.; Milli, M.C.; Cao, L. Feed conversion efficiency in aquaculture: Do we measure it correctly? Environ. Res. Lett. 2018, 13, 024017. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2020. In Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Tacon, A.G.J.; Metian, M. Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects. Aquaculture 2008, 285, 146–158. [Google Scholar] [CrossRef]
- Tomás-Almenar, C.; Toledo-Solís, F.J.; Larrán, A.M.; De Mercado, E.; Alarcón, F.J.; Rico, D.; Martín-Diana, A.B.; Fernández, I. Effects and Safe Inclusion of Narbonne Vetch (Vicia narbonensis) in Rainbow Trout (Oncorhynchus mykiss) Diets: Towards a More Sustainable Aquaculture. Animals 2020, 10, 2175. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Guo, X.; Tan, B.; Dong, X.; Yang, Q.; Liu, H.; Zhang, S.; Chi, S. Replacing fish meal with fermented rice protein in diets for hybrid groupers (Epinephelus fuscoguttatus × Epinephelus lanceolatus): Effects on growth, digestive and absorption capacities, inflammatory-related gene expression, and intestinal microbiota. Aquaculture Rep. 2021, 19, 100603. [Google Scholar] [CrossRef]
- Øverland, M.; Skrede, A. Yeast derived from lignocellulosic biomass as a sustainable feed resource for use in aquaculture. J. Sci. Food Agric. 2017, 97, 733–742. [Google Scholar] [CrossRef]
- Agboola, J.O.; Schiavone, M.; Øverland, M.; Morales-Lange, B.; Lagos, L.; Arntzen, M.Ø.; Lapeña, D.; Eijsink, V.G.H.; Horn, S.J.; Mydland, L.T.; et al. Author Correction: Impact of down-stream processing on functional properties of yeasts and the implications on gut health of Atlantic salmon (Salmo salar). Sci. Rep. 2021, 11, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Alagawany, M.; Taha, A.E.; Noreldin, A.; El-Tarabily, K.A.; El-Hack, M.E.A. Nutritional applications of species of Spirulina and Chlorella in farmed fish: A review. Aquaculture 2021, 542, 736841. [Google Scholar] [CrossRef]
- Serrano, E.; Simpfendorfer, R.; Medina, A.; Sandoval, C.; Martínez, A.; Morales, R.; Davies, S.J. Partially replacing fish oil with microalgae (Schizochytrium limacinum and Nannochloropsis oceanica) in diets for rainbow trout (Oncorhynchus mykiss) reared in saltwater with reference to growth performance, muscle fatty acid composition and liver ultrastructure. Aquac. Res. 2021, 52, 4401–4413. [Google Scholar] [CrossRef]
- Tran, G.; Heuzé, V.; Makkar, H.P.S. Insects in Fish Diets. Animals Front. 2015, 5, 37–44. [Google Scholar] [CrossRef]
- Gasco, L.; Acuti, G.; Bani, P.; Dalle Zotte, A.; Danieli, P.P.; De Angelis, A.; Fortina, R.; Marino, R.; Parisi, G.; Piccolo, G.; et al. Insect and fish by-products as sustainable alternatives to conventional animal proteins in animal nutrition. Ital. J. Animals Sci. 2020, 19, 360–372. [Google Scholar] [CrossRef] [Green Version]
- Mousavi, S.; Zahedinezhad, S.; Loh, J.Y. A review on insect meals in aquaculture: The immunomodulatory and physiological effects. Int. Aquat. Res 2020, 12, 100–115. [Google Scholar]
- Shafique, L.; Abdel-Latif, H.; Hassan, F.-U.; Alagawany, M.; Naiel, M.; Dawood, M.; Yilmaz, S.; Liu, Q. The Feasibility of Using Yellow Mealworms (Tenebrio molitor): Towards a Sustainable Aquafeed Industry. Animals 2021, 11, 811. [Google Scholar] [CrossRef]
- Rodjaroen, S.; Thongprajukaew, K.; Khongmuang, P.; Malawa, S.; Tuntikawinwong, K.; Saekhow, S. Ontogenic Development of Digestive Enzymes in Mealworm Larvae (Tenebrio molitor) and Their Suitable Harvesting Time for Use as Fish Feed. Insects 2020, 11, 393. [Google Scholar] [CrossRef]
- Barroso, F.G.; de Haro, C.; Sánchez-Muros, M.-J.; Venegas, E.; Martínez-Sánchez, A.; Pérez-Bañón, C. The potential of various insect species for use as food for fish. Aquaculture 2014, 422-423, 193–201. [Google Scholar] [CrossRef]
- Makkar, H.P.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Animals Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Van Huis, A. Potential of Insects as Food and Feed in Assuring Food Security. Annu. Rev. Èntomol. 2013, 58, 563–583. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Factsheets on the 21 SDG Indicators under FAO Custodianship—A Highlight of the Main Indicators with the Greatest Gaps in Country Reporting; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar]
- Moutinho, S.; Pedrosa, R.; Magalhães, R.; Oliva-Teles, A.; Parisi, G.; Peres, H. Black soldier fly (Hermetia illucens) pre-pupae larvae meal in diets for European seabass (Dicentrarchus labrax) juveniles: Effects on liver oxidative status and fillet quality traits during shelf-life. Aquaculture 2021, 533, 736080. [Google Scholar] [CrossRef]
- Li, S.; Ji, H.; Zhang, B.; Zhou, J.; Yu, H. Defatted black soldier fly (Hermetia illucens) larvae meal in diets for juvenile Jian carp (Cyprinus carpio var. Jian): Growth performance, antioxidant enzyme activities, digestive enzyme activities, intestine and hepatopancreas histological structure. Aquaculture 2017, 477, 62–70. [Google Scholar] [CrossRef]
- Guerreiro, I.; Castro, C.; Antunes, B.; Coutinho, F.; Rangel, F.; Couto, A.; Serra, C.; Peres, H.; Pousão-Ferreira, P.; Matos, E.; et al. Catching black soldier fly for meagre: Growth, whole-body fatty acid profile and metabolic responses. Aquaculture 2020, 516, 734613. [Google Scholar] [CrossRef]
- Coutinho, F.; Castro, C.; Guerreiro, I.; Rangel, F.; Couto, A.; Serra, C.R.; Peres, H.; Pousão-Ferreira, P.; Rawski, M.; Oliva-Teles, A.; et al. Mealworm larvae meal in diets for meagre juveniles: Growth, nutrient digestibility and digestive enzymes activity. Aquaculture 2021, 535, 736362. [Google Scholar] [CrossRef]
- Tilami, S.K.; Turek, J.; Červený, D.; Lepič, P.; Kozák, P.; Burkina, V.; Sakalli, S.; Tomčala, A.; Sampels, S.; Mráz, J. Insect Meal as a Partial Replacement for Fish Meal in a Formulated Diet for Perch Perca fluviatilis. Turk. J. Fish. Aquat. Sci. 2020, 20, 867–878. [Google Scholar] [CrossRef]
- Terova, G.; Rimoldi, S.; Ascione, C.; Gini, E.; Ceccotti, C.; Gasco, L. Rainbow trout (Oncorhynchus mykiss) gut microbiota is modulated by insect meal from Hermetia illucens prepupae in the diet. Rev. Fish Biol. Fish. 2019, 29, 465–486. [Google Scholar] [CrossRef]
- Belghit, I.; Liland, N.S.; Gjesdal, P.; Biancarosa, I.; Menchetti, E.; Li, Y.; Waagbo, R.; Krogdahl, Å.; Lock, E.-J. Black soldier fly larvae meal can replace fish meal in diets of sea-water phase Atlantic salmon (Salmo salar). Aquaculture 2019, 503, 609–619. [Google Scholar] [CrossRef]
- Mikołajczak, Z.; Rawski, M.; Mazurkiewicz, J.; Kierończyk, B.; Józefiak, D. The Effect of Hydrolyzed Insect Meals in Sea Trout Fingerling (Salmo trutta m. trutta) Diets on Growth Performance, Microbiota and Biochemical Blood Parameters. Animals 2020, 10, 1031. [Google Scholar] [CrossRef] [PubMed]
- Chemello, G.; Renna, M.; Caimi, C.; Guerreiro, I.; Oliva-Teles, A.; Enes, P.; Biasato, I.; Schiavone, A.; Gai, F.; Gasco, L. Partially Defatted Tenebrio molitor Larva Meal in Diets for Grow-Out Rainbow Trout, Oncorhynchus mykiss (Walbaum): Effects on Growth Performance, Diet Digestibility and Metabolic Responses. Animals 2020, 10, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngo, D.-H.; Kim, S.-K. Chapter Two–Antioxidant Effects of Chitin, Chitosan, and Their Derivatives. Adv. Food Nutr. Res. 2014, 73, 15–31. [Google Scholar] [PubMed]
- Henry, M.; Gai, F.; Enes, P.; Peréz-Jiménez, A.; Gasco, L. Effect of partial dietary replacement of fishmeal by yellow mealworm (Tenebrio molitor) larvae meal on the innate immune response and intestinal antioxidant enzymes of rainbow trout (Oncorhynchus mykiss). Fish Shellfish. Immunol. 2018, 83, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Ji, H.; Yu, H.; Zhou, J. Influence of dietary black soldier fly (Hermetia illucens Linnaeus) pulp on growth performance, antioxidant capacity and intestinal health of juvenile mirror carp (Cyprinus carpio var. specularis). Aquac. Nutr. 2019, 26, 432–443. [Google Scholar] [CrossRef]
- Melenchón, F.; Larrán, A.; de Mercado, E.; Hidalgo, M.; Cardenete, G.; Barroso, F.; Fabrikov, D.; Lourenço, H.; Pessoa, M.; Tomás-Almenar, C. Potential use of black soldier fly (Hermetia illucens) and mealworm (Tenebrio molitor) insectmeals in diets for rainbow trout (Oncorhynchus mykiss). Aquac. Nutr. 2021, 27, 491–505. [Google Scholar] [CrossRef]
- Weththasinghe, P.; Lagos, L.; Cortés, M.; Hansen, J. Øvrum; Øverland, M. Dietary Inclusion of Black Soldier Fly (Hermetia Illucens) Larvae Meal and Paste Improved Gut Health but Had Minor Effects on Skin Mucus Proteome and Immune Response in Atlantic Salmon (Salmo salar). Front. Immunol. 2021, 12, 599530. [Google Scholar] [CrossRef]
- European Parliament. Commission Regulation 2017/893/EU of 24 May 2017 Amending Annexes I and IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council and Annexes X, XIV and XV to Commission Regulation (EU) No 142/2011 as Regards the Provisions on Processed Animal Protein; European Parliament: Strasbourg, France, 2017; pp. 92–116. [Google Scholar]
- Blanco Cachafeiro, M.C. La Trucha: Cría Industrial, 2nd ed.; Mundi-Prensa: Madrid, Spain, 2005. [Google Scholar]
- National Research Council. Nutrient Requirements of Fish; National Academies Press: Washington, DC, USA, 1993. [Google Scholar]
- European Parliament. Book Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes; European Parliament: Strasbourg, France, 2010; pp. 33–78. [Google Scholar]
- Real Decreto 53/2013, de 1 de Febrero, Por el Que se Establecen las Normas Básicas Aplicables para la Protección de Los Animales Utilizados en Experimentación y Otros Fines Científicos, Incluyendo la Docencia. 2013. Available online: https://www.boe.es/eli/es/rd/2013/02/01/53 (accessed on 16 December 2021).
- Cho, C.; Slinger, S.; Bayley, H. Bioenergetics of salmonid fishes: Energy intake, expenditure and productivity. Comp. Biochem. Physiol. Part B Comp. Biochem. 1982, 73, 25–41. [Google Scholar] [CrossRef]
- De Mercado, E.; Larrán, A.M.; Pinedo, J.; Tomás-Almenar, C. Skin mucous: A new approach to assess stress in rainbow trout. Aquaculture 2018, 484, 90–97. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Communities International, 18th ed.; AOAC: Rockville, MD, USA, 2005. [Google Scholar]
- Saint-Denis, T.; Goupy, J. Optimization of a nitrogen analyser based on the Dumas method. Anal. Chim. Acta 2004, 515, 191–198. [Google Scholar] [CrossRef]
- Janssen, R.H.; Vincken, J.-P.; van den Broek, L.A.M.; Fogliano, V.; Lakemond, C.M.M. Nitrogen-to-Protein Conversion Factors for Three Edible Insects: Tenebrio molitor, Alphitobius diaperinus, and Hermetia illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef]
- Atkinson, J.L.; Hilton, J.W.; Slinger, S.J. Evaluation of Acid-Insoluble Ash as an Indicator of Feed Digestibility in Rainbow Trout (Salmo gairdneri). Can. J. Fish. Aquat. Sci. 1984, 41, 1384–1386. [Google Scholar] [CrossRef]
- ISO 13730. Meat and Meat Products—Determination of Total Phosphorus Content—Spectrometric Method: ISO Technical Committee TC 34/SC 6; ISO: Geneva, Switzerland, 1996. [Google Scholar]
- Pessoa, M.; Campos, P.S.; Pais, I.; Feteiro, A.; Canuto, D.; Simões, M.; Pelica, J.; Pataco, I.; Ribeiro, V.S.; Ramalho, J.C.; et al. Nutritional profile of the Portuguese cabbage (Brassica oleracea L var. costata) and its relationship with the elemental soil analysis. Emir. J. Food Agric. 2016, 28, 381–388. [Google Scholar] [CrossRef] [Green Version]
- Gamage, A.; Shahidi, F. Use of chitosan for the removal of metal ion contaminants and proteins from water. Food Chem. 2007, 104, 989–996. [Google Scholar] [CrossRef] [Green Version]
- Somogyi, M. Notes on Sugar Determination. J. Biol. Chem. 1952, 195, 19–23. [Google Scholar] [CrossRef]
- Walter, H.E. Proteinases: Methods with Hemoglobin, Casein and Azocoll as Substrates. In Methods of Enzymatic Analysis; Bergmeyer, H.J., Ed.; Elsevier: Amsterdam, The Netherlands, 1984; Volume 5, pp. 270–277. [Google Scholar]
- Anson, M.L. The Estimation of Pepsin, Trypsin, Papain, And Cathepsin with Hemoglobin. J. Gen. Physiol. 1938, 22, 79–89. [Google Scholar] [CrossRef]
- Furné, M.; Morales, A.E.; Trenzado, C.E.; García-Gallego, M.; Hidalgo, M.C.; Domezain, A.; Rus, A.S. The metabolic effects of prolonged starvation and refeeding in sturgeon and rainbow trout. J. Comp. Physiol. B 2011, 182, 63–76. [Google Scholar] [CrossRef]
- Swain, P.; Dash, S.; Sahoo, P.K.; Routray, P.; Sahoo, S.; Gupta, S.D.; Meher, P.K.; Sarangi, N. Non-specific immune parameters of brood Indian major carp Labeo rohita and their seasonal variations. Fish Shellfish Immunol. 2007, 22, 38–43. [Google Scholar] [CrossRef]
- Mashiter, K.E.; Morgan, M. Carbonic anhydrase levels in the tissues of flounders adapted to sea water and fresh water. Comp. Biochem. Physiol. Part A Physiol. 1975, 52, 713–717. [Google Scholar] [CrossRef]
- Thompson, I.; Choubert, G.; Houlihan, D.; Secombes, C. The effect of dietary vitamin A and astaxanthin on the immunocompetence of rainbow trout. Aquaculture 1995, 133, 91–102. [Google Scholar] [CrossRef]
- Huang, Z.-H.; Ma, A.-J.; Wang, X.-A. The immune response of turbot, Scophthalmus maximus (L.), skin to high water temperature. J. Fish Dis. 2011, 34, 619–627. [Google Scholar] [CrossRef]
- Mohanty, B.; Sahoo, P. Immune responses and expression profiles of some immune-related genes in Indian major carp, Labeo rohita to Edwardsiella tarda infection. Fish Shellfish. Immunol. 2010, 28, 613–621. [Google Scholar] [CrossRef]
- Panigrahi, A.; Kiron, V.; Puangkaew, J.; Kobayashi, T.; Satoh, S.; Sugita, H. The viability of probiotic bacteria as a factor influencing the immune response in rainbow trout Oncorhynchus mykiss. Aquaculture 2005, 243, 241–254. [Google Scholar] [CrossRef]
- Morales, B.; Bethke, J.; Schmitt, P.; Mercado, L. Phenotypical parameters as a tool to evaluate the immunostimulatory effects of laminarin in Oncorhynchus mykiss. Aquac. Res. 2014, 46, 2707–2715. [Google Scholar] [CrossRef]
- Pérez-Jiménez, A.; Hidalgo, M.C.; Morales, A.E.; Arizcun, M.; Abellán, E.; Cardenete, G. Antioxidant enzymatic defenses and oxidative damage in Dentex dentex fed on different dietary macronutrient levels. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2009, 150, 537–545. [Google Scholar] [CrossRef]
- Sealey, W.M.; Gaylord, T.G.; Barrows, F.T.; Tomberlin, J.K.; McGuire, M.A.; Ross, C.; St-Hilaire, S. Sensory Analysis of Rainbow Trout, Oncorhynchus mykiss, Fed Enriched Black Soldier Fly Prepupae, Hermetia illucens. J. World Aquaculture Soc. 2011, 42, 34–45. [Google Scholar] [CrossRef]
- Borgogno, M.; Dinnella, C.; Iaconisi, V.; Fusi, R.; Scarpaleggia, C.; Schiavone, A.; Monteleone, E.; Gasco, L.; Parisi, G. Inclusion of Hermetia illucens larvae meal on rainbow trout (Oncorhynchus mykiss) feed: Effect on sensory profile according to static and dynamic evaluations. J. Sci. Food Agric. 2017, 97, 3402–3411. [Google Scholar] [CrossRef] [PubMed]
- Jozefiak, A.; Nogales-Mérida, S.; Mikołajczak, Z.; Rawski, M.; Kierończyk, B.; Mazurkiewicz, J. The Utilization of Full-Fat Insect Meal in Rainbow Trout (Oncorhynchus mykiss) Nutrition: The Effects on Growth Performance, Intestinal Microbiota and Gastrointestinal Tract Histomorphology. Ann. Animals Sci. 2019, 19, 747–765. [Google Scholar] [CrossRef] [Green Version]
- Caimi, C.; Renna, M.; Lussiana, C.; Bonaldo, A.; Gariglio, M.; Meneguz, M.; Dabbou, S.; Schiavone, A.; Gai, F.; Elia, A.C.; et al. First insights on Black Soldier Fly (Hermetia illucens L.) larvae meal dietary administration in Siberian sturgeon (Acipenser baerii Brandt) juveniles. Aquaculture 2020, 515, 734539. [Google Scholar] [CrossRef]
- Belforti, M.; Gai, F.; Lussiana, C.; Renna, M.; Malfatto, V.; Rotolo, L.; De Marco, M.; Dabbou, S.; Schiavone, A.; Zoccarato, I.; et al. Tenebrio molitor meal in rainbow trout (Oncorhynchus mykiss) diets: Effects on animal performance, nutrient digestibility and chemical composition of fillets. Ital. J. Animals Sci. 2015, 14, 4170. [Google Scholar] [CrossRef] [Green Version]
- Iaconisi, V.; Bonelli, A.; Pupino, R.; Gai, F.; Parisi, G. Mealworm as dietary protein source for rainbow trout: Body and fillet quality traits. Aquaculture 2018, 484, 197–204. [Google Scholar] [CrossRef]
- Dumas, A.; Raggi, T.; Barkhouse, J.; Lewis, E.; Weltzien, E. The oil fraction and partially defatted meal of black soldier fly larvae (Hermetia illucens) affect differently growth performance, feed efficiency, nutrient deposition, blood glucose and lipid digestibility of rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 492, 24–34. [Google Scholar] [CrossRef]
- Kumar, R.; Kaur, N.; Kamilya, D. Chitin modulates immunity and resistance of Labeo rohita (Hamilton, 1822) against gill monogeneans. Aquaculture 2019, 498, 522–527. [Google Scholar] [CrossRef]
- Kroeckel, S.; Harjes, A.-G.; Roth, I.; Katz, H.; Wuertz, S.; Susenbeth, A.; Schulz, C. When a turbot catches a fly: Evaluation of a pre-pupae meal of the Black Soldier Fly (Hermetia illucens) as fish meal substitute—Growth performance and chitin degradation in juvenile turbot (Psetta maxima). Aquaculture 2012, 364–365, 345–352. [Google Scholar] [CrossRef]
- Marono, S.; Piccolo, G.; Loponte, R.; Di Meo, C.; Attia, Y.A.; Nizza, A.; Bovera, F. In Vitro Crude Protein Digestibility of Tenebrio Molitor and Hermetia Illucens Insect Meals and its Correlation with Chemical Composition Traits. Ital. J. Animals Sci. 2015, 14, 3889. [Google Scholar] [CrossRef] [Green Version]
- Cardinaletti, G.; Randazzo, B.; Messina, M.; Zarantoniello, M.; Giorgini, E.; Zimbelli, A.; Bruni, L.; Parisi, G.; Olivotto, I.; Tulli, F. Effects of Graded Dietary Inclusion Level of Full-Fat Hermetia illucens Prepupae Meal in Practical Diets for Rainbow Trout (Oncorhynchus mykiss). Animals 2019, 9, 251. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K.; Fotedar, R.; Foysal, J.; Priyam, M.; Siddik, M.A.B.; Chaklader, R.; Dao, T.T.T.; Howieson, J. Impact of varied combinatorial mixture of non-fishmeal ingredients on growth, metabolism, immunity and gut microbiota of Lates calcarifer (Bloch, 1790) fry. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Basto, A.; Calduch-Giner, J.; Oliveira, B.; Petit, L.; Sá, T.; Maia, M.R.G.; Fonseca, S.C.; Matos, E.; Pérez-Sánchez, J.; Valente, L.M.P. The Use of Defatted Tenebrio molitor Larvae Meal as a Main Protein Source Is Supported in European Sea Bass (Dicentrarchus labrax) by Data on Growth Performance, Lipid Metabolism, and Flesh Quality. Front. Physiol. 2021, 12, 659567. [Google Scholar] [CrossRef]
- Chaklader, R.; Howieson, J.; Siddik, M.A.B.; Foysal, J.; Fotedar, R. Supplementation of tuna hydrolysate and insect larvae improves fishmeal replacement efficacy of poultry by-product in Lates calcarifer (Bloch, 1790) juveniles. Sci. Rep. 2021, 11, 1–20. [Google Scholar] [CrossRef]
- Vargas-Abúndez, A.J.; Randazzo, B.; Foddai, M.; Sanchini, L.; Truzzi, C.; Giorgini, E.; Gasco, L.; Olivotto, I. Insect meal based diets for clownfish: Biometric, histological, spectroscopic, biochemical and molecular implications. Aquaculture 2019, 498, 1–11. [Google Scholar] [CrossRef]
- Lu, R.; Chen, Y.; Yu, W.; Lin, M.; Yang, G.; Qin, C.; Meng, X.; Zhang, Y.; Ji, H.; Nie, G. Defatted black soldier fly (Hermetia illucens) larvae meal can replace soybean meal in juvenile grass carp (Ctenopharyngodon idellus) diets. Aquac. Rep. 2020, 18, 100520. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Randazzo, B.; Nozzi, V.; Truzzi, C.; Giorgini, E.; Cardinaletti, G.; Freddi, L.; Ratti, S.; Girolametti, F.; Osimani, A.; et al. Physiological responses of Siberian sturgeon (Acipenser baerii) juveniles fed on full-fat insect-based diet in an aquaponic system. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Randazzo, B.; Gioacchini, G.; Truzzi, C.; Giorgini, E.; Riolo, P.; Gioia, G.; Bertolucci, C.; Osimani, A.; Cardinaletti, G.; et al. Zebrafish (Danio rerio) physiological and behavioural responses to insect-based diets: A multidisciplinary approach. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef]
- Hender, A.; Siddik, M.; Howieson, J.; Fotedar, R. Black Soldier Fly, Hermetia illucens as an Alternative to Fishmeal Protein and Fish Oil: Impact on Growth, Immune Response, Mucosal Barrier Status, and Flesh Quality of Juvenile Barramundi, Lates calcarifer (Bloch, 1790). Biology 2021, 10, 505. [Google Scholar] [CrossRef]
- Józefiak, A.; Nogales-Mérida, S.; Rawski, M.; Kierończyk, B.; Mazurkiewicz, J. Effects of insect diets on the gastrointestinal tract health and growth performance of Siberian sturgeon (Acipenser baerii Brandt, 1869). BMC Veter. Res. 2019, 15, 348. [Google Scholar] [CrossRef] [Green Version]
- Rawski, M.; Mazurkiewicz, J.; Kierończyk, B.; Józefiak, D. Black Soldier Fly Full-Fat Larvae Meal Is More Profitable Than Fish Meal and Fish Oil in Siberian Sturgeon Farming: The Effects on Aquaculture Sustainability, Economy and Fish GIT Development. Animals 2021, 11, 604. [Google Scholar] [CrossRef]
- Romarheim, O.H.; Skrede, A.; Gao, Y.; Krogdahl, Å.; Denstadli, V.; Lilleeng, E.; Storebakken, T. Comparison of white flakes and toasted soybean meal partly replacing fish meal as protein source in extruded feed for rainbow trout (Oncorhynchus mykiss). Aquaculture 2006, 256, 354–364. [Google Scholar] [CrossRef]
- Penn, M.H.; Bendiksen, E.Å.; Campbell, P.; Krogdahl, Å. High level of dietary pea protein concentrate induces enteropathy in Atlantic salmon (Salmo salar L.). Aquaculture 2011, 310, 267–273. [Google Scholar] [CrossRef]
- Zhang, C.; Rahimnejad, S.; Wang, Y.-R.; Lu, K.; Song, K.; Wang, L.; Mai, K. Substituting fish meal with soybean meal in diets for Japanese seabass (Lateolabrax japonicus): Effects on growth, digestive enzymes activity, gut histology, and expression of gut inflammatory and transporter genes. Aquaculture 2018, 483, 173–182. [Google Scholar] [CrossRef]
- Wiles, T.; Jemielita, M.; Baker, R.P.; Schlomann, B.H.; Logan, S.L.; Ganz, J.; Melancon, E.; Eisen, J.S.; Guillemin, K.; Parthasarathy, R. Host Gut Motility Promotes Competitive Exclusion within a Model Intestinal Microbiota. PLoS Biol. 2016, 14, e1002517. [Google Scholar] [CrossRef]
- Lignot, J.-H.; Helmstetter, C.; Secor, S.M. Postprandial morphological response of the intestinal epithelium of the Burmese python (Python molurus). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2005, 141, 280–291. [Google Scholar] [CrossRef] [Green Version]
- Starck, J.M.; Cruz-Neto, A.; Abe, A.S. Physiological and morphological responses to feeding in broad-nosed caiman (Caiman latirostris). J. Exp. Biol. 2007, 210, 2033–2045. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Fawole, F.J.; Romano, N.; Hossain, S.; Labh, S.N.; Overturf, K.; Small, B.C. Insect (black soldier fly, Hermetia illucens) meal supplementation prevents the soybean meal-induced intestinal enteritis in rainbow trout and health benefits of using insect oil. Fish Shellfish Immunol. 2020, 109, 116–124. [Google Scholar] [CrossRef]
- Baeverfjord, G.; Krogdahl, A. Development and regression of soybean meal induced enteritis in Atlantic salmon, Salmo salar L., distal intestine: A comparison with the intestines of fasted fish. J. Fish Dis. 1996, 19, 375–387. [Google Scholar] [CrossRef]
- Lindsay, G.J.; Walton, M.; Adron, J.; Fletcher, T.C.; Cho, C.; Cowey, C. The growth of rainbow trout (Salmo gairdneri) given diets containing chitin and its relationship to chitinolytic enzymes and chitin digestibility. Aquaculture 1984, 37, 315–334. [Google Scholar] [CrossRef]
- German, D.P.; Horn, M.H.; Gawlicka, A. Digestive Enzyme Activities in Herbivorous and Carnivorous Prickleback Fishes (Teleostei: Stichaeidae): Ontogenetic, Dietary, and Phylogenetic Effects. Physiol. Biochem. Zoöl. 2004, 77, 789–804. [Google Scholar] [CrossRef] [Green Version]
- Rapatsa, M.; Moyo, N.A. Evaluation of Imbrasia belina meal as a fishmeal substitute in Oreochromis mossambicus diets: Growth performance, histological analysis and enzyme activity. Aquaculture Rep. 2017, 5, 18–26. [Google Scholar] [CrossRef]
- Rapatsa, M.M.; Moyo, N.A.G. Enzyme activity and histological analysis of Clarias gariepinus fed on Imbrasia belina meal used for partial replacement of fishmeal. Fish Physiol. Biochem. 2019, 45, 1309–1320. [Google Scholar] [CrossRef] [PubMed]
- Mastoraki, M.; Ferrándiz, P.M.; Vardali, S.C.; Kontodimas, D.C.; Kotzamanis, Y.P.; Gasco, L.; Chatzifotis, S.; Antonopoulou, E. A comparative study on the effect of fish meal substitution with three different insect meals on growth, body composition and metabolism of European sea bass (Dicentrarchus labrax L.). Aquaculture 2020, 528, 735511. [Google Scholar] [CrossRef]
- Mastoraki, M.; Vlahos, N.; Patsea, E.; Chatzifotis, S.; Mente, E.; Antonopoulou, E. The effect of insect meal as a feed ingredient on survival, growth, and metabolic and antioxidant response of juvenile prawn Palaemon adspersus (Rathke, 1837). Aquaculture Res. 2020, 51, 3551–3562. [Google Scholar] [CrossRef]
- Alejo, A.; Tafalla, C. Chemokines in teleost fish species. Dev. Comp. Immunol. 2011, 35, 1215–1222. [Google Scholar] [CrossRef] [PubMed]
- Grosell, M.; Farrell, A.P.; Brauner, C.J. Fish Physiology: The Multifunctional Gut of Fish; Academic Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Subramanian, S.; Ross, N.W.; MacKinnon, S.L. Comparison of antimicrobial activity in the epidermal mucus extracts of fish. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 150, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Molina, R.; Moreno, I.; Pichardo, S.; Jos, A.; Moyano, R.; Monterde, J.; Cameán, A. Acid and alkaline phosphatase activities and pathological changes induced in Tilapia fish (Oreochromis sp.) exposed subchronically to microcystins from toxic cyanobacterial blooms under laboratory conditions. Toxicon 2005, 46, 725–735. [Google Scholar] [CrossRef]
- Moreno, I.M.; Mate, A.; Repetto, G.; Vázquez, C.M.; Cameán, A.M. Influence of Microcystin-LR on the activity of membrane enzymes in rat intestinal mucosa. J. Physiol. Biochem. 2003, 59, 293–299. [Google Scholar] [CrossRef]
- Alves, A.P.D.C.; Paulino, R.R.; Pereira, R.T.; Da Costa, D.V.; E Rosa, P.V. Nile tilapia fed insect meal: Growth and innate immune response in different times under lipopolysaccharide challenge. Aquaculture Res. 2021, 52, 529–540. [Google Scholar] [CrossRef]
- Tippayadara, N.; Dawood, M.; Krutmuang, P.; Hoseinifar, S.; Doan, H.; Paolucci, M. Replacement of Fish Meal by Black Soldier Fly (Hermetia illucens) Larvae Meal: Effects on Growth, Haematology, and Skin Mucus Immunity of Nile Tilapia, Oreochromis niloticus. Animals 2021, 11, 193. [Google Scholar] [CrossRef]
- Vogel, H.; Müller, A.; Heckel, D.; Gutzeit, H.; Vilcinskas, A. Nutritional immunology: Diversification and diet-dependent expression of antimicrobial peptides in the black soldier fly Hermetia illucens. Dev. Comp. Immunol. 2018, 78, 141–148. [Google Scholar] [CrossRef]
- Xu, X.; Ji, H.; Belghit, I.; Liland, N.S.; Wu, W.; Li, X. Effects of black soldier fly oil rich in n-3 HUFA on growth performance, metabolism and health response of juvenile mirror carp (Cyprinus carpio var. specularis). Aquaculture 2021, 533, 736144. [Google Scholar] [CrossRef]
- Grotto, D.; Maria, L.S.; Valentini, J.; Paniz, C.; Schmitt, G.; Garcia, S.; Pomblum, V.J.; da Rocha, J.B.T.; Farina, M. Importance of the lipid peroxidation biomarkers and methodological aspects FOR malondialdehyde quantification. Química Nova 2009, 32, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Muros, M.J.; De Haro, C.; Sanz, A.; Trenzado, C.E.; Villareces, S.; Barroso, F.G. Nutritional evaluation ofTenebrio molitormeal as fishmeal substitute for tilapia (Oreochromis niloticus) diet. Aquac. Nutr. 2016, 22, 943–955. [Google Scholar] [CrossRef]
- Acerete, L.; Reig, L.; Alvarez, D.; Flos, R.; Tort, L. Comparison of two stunning/slaughtering methods on stress response and quality indicators of European sea bass (Dicentrarchus labrax). Aquaculture 2009, 287, 139–144. [Google Scholar] [CrossRef]
- Naderi, M.; Keyvanshokooh, S.; Salati, A.P.; Ghaedi, A. Effects of dietary vitamin E and selenium nanoparticles supplementation on acute stress responses in rainbow trout (Oncorhynchus mykiss) previously subjected to chronic stress. Aquaculture 2017, 473, 215–222. [Google Scholar] [CrossRef]
- Gesto, M. Consistent individual competitive ability in rainbow trout as a proxy for coping style and its lack of correlation with cortisol responsiveness upon acute stress. Physiol. Behav. 2019, 208, 112576. [Google Scholar] [CrossRef]
- Tran, H.Q.; Van Doan, H.; Stejskal, V. Does dietary Tenebrio molitor affect swimming capacity, energy use, and physiological responses of European perch Perca fluviatilis? Aquaculture 2021, 539, 736610. [Google Scholar] [CrossRef]
- Jeong, S.; Khosravi, S.; Mauliasari, I.R.; Lee, B.; You, S.; Lee, S. Nutritional evaluation of cricket, Gryllus bimaculatus, meal as fish meal substitute for olive flounder, Paralichthys olivaceus, juveniles. J. World Aquaculture Soc. 2021, 52, 859–880. [Google Scholar] [CrossRef]
- Mancini, S.; Medina, I.; Iaconisi, V.; Gai, F.; Basto, A.; Parisi, G. Impact of black soldier fly larvae meal on the chemical and nutritional characteristics of rainbow trout fillets. Animals 2018, 12, 1672–1681. [Google Scholar] [CrossRef]
- Renna, M.; Schiavone, A.; Gai, F.; Dabbou, S.; Lussiana, C.; Malfatto, V.; Prearo, M.; Capucchio, M.T.; Biasato, I.; Biasibetti, E.; et al. Evaluation of the suitability of a partially defatted black soldier fly (Hermetia illucens L.) larvae meal as ingredient for rainbow trout (Oncorhynchus mykiss Walbaum) diets. J. Animals Sci. Biotechnol. 2017, 8, 57. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Proximate Composition | Hermetia ilucens (HI) | Tenebrio molitor (TM) |
|---|---|---|
| Crude protein (%) | 28.5 | 39.1 |
| Crude fat (%) | 25.6 | 27.0 |
| Crude ash (%) | 9.75 | 3.42 |
| Moisture (%) | 8.00 | 5.00 |
| Calcium (g/Kg) | 35.2 | 0.93 |
| Phosphorus (g/Kg) | 7.00 | 7.50 |
| Calcium–phosphorus ratio | 5.03 | 0.12 |
| Chitin (%) | 7.50 | 5.90 |
| Amino acid composition (g/100 g IM) | ||
| Asp (aspartate) | 2.92 | 3.71 |
| Thr (threonine) | 0.95 | 1.44 |
| Ser (serine) | 1.43 | 2.49 |
| Glu (glutamate) | 3.19 | 4.98 |
| Pro (proline) | 1.58 | 3.04 |
| Gly (glycine) | 1.84 | 2.87 |
| Ala (alanine) | 2.37 | 3.92 |
| Cys (cysteine) | 0.13 | 0.24 |
| Val (valine) | 1.42 | 2.32 |
| Met (methionine) | 0.47 | 0.57 |
| Ile (isoleucine) | 0.91 | 1.31 |
| Leu (leucine) | 1.86 | 2.96 |
| Tyr (tyrosine) | 2.23 | 4.47 |
| Phe (phenylalanine) | 2.16 | 3.07 |
| His (histidine) | 1.07 | 1.77 |
| Lys (lysine) | 1.94 | 2.49 |
| Arg (arginine) | 1.24 | 1.81 |
| Ingredients (%; on Wet Basis) | C | H18 | T18 |
|---|---|---|---|
| Fishmeal | 35.9 | 18.0 | 18.0 |
| HI meal | 0.00 | 18.0 | 0.00 |
| TM meal | 0.00 | 0.00 | 18.0 |
| Wheat gluten | 10.5 | 15.4 | 11.9 |
| Soy protein concentrate | 15.5 | 18.3 | 17.0 |
| Wheat meal | 16.4 | 11.5 | 17.0 |
| Soy lecithin | 1.30 | 0.50 | 0.50 |
| Fish oil | 12.2 | 9.50 | 9.00 |
| Vitamins and minerals | 2.00 | 2.00 | 2.00 |
| Goma guar | 2.00 | 2.00 | 2.00 |
| Blood meal | 4.00 | 4.00 | 4.00 |
| Methionine | 0.20 | 0.50 | 0.50 |
| Lysine | 0.00 | 0.40 | 0.10 |
| Total | 100 | 100 | 100 |
| Proximate composition (%; on wet basis) | |||
| Moisture | 7.43 | 7.74 | 7.21 |
| Total crude protein | 43.9 | 42.8 | 43.1 |
| Total crude fat | 17.2 | 17.1 | 17.9 |
| Ash | 7.41 | 6.45 | 6.11 |
| Calcium | 0.51 | 0.26 | 0.45 |
| Phosphorus | 0.33 | 0.24 | 0.24 |
| Calcium–phosphorus ratio | 1.6 | 1.1 | 1.9 |
| Amino acid composition * (g/100 g feed) | |||
| Asp (aspartate) | 2.89 | 1.86 | 2.29 |
| Thr (threonine) | 1.65 | 1.25 | 1.38 |
| Ser (serine) | 1.59 | 1.22 | 1.46 |
| Glu (glutamate) | 6.55 | 5.96 | 5.96 |
| Pro (proline) | 2.15 | 2.11 | 1.84 |
| Gly (glycine) | 1.78 | 1.16 | 1.53 |
| Ala (alanine) | 1.73 | 1.05 | 1.58 |
| Cys (cysteine) | 0.68 | 0.67 | 0.64 |
| Val (valine) | 2.07 | 1.65 | 1.87 |
| Met (methionine) | 1.22 | 1.13 | 1.26 |
| Ile (isoleucine) | 1.93 | 1.55 | 1.63 |
| Leu (leucine) | 3.15 | 2.56 | 2.79 |
| Tyr (tyrosine) | 1.11 | 0.87 | 1.42 |
| Phe (phenylalanine) | 1.89 | 1.63 | 1.93 |
| His (histidine) | 1.03 | 0.84 | 1.03 |
| Lys (lysine) | 2.66 | 1.88 | 2.18 |
| Arg (arginine) | 2.50 | 1.86 | 2.04 |
| Growth Performance | C | H18 | T18 | SEM | p-Value |
|---|---|---|---|---|---|
| IBW (g) | 14.3 | 14.8 | 14.7 | 0.22 | 0.325 |
| IBL (cm) | 11.1 | 11.2 | 11.2 | 0.05 | 0.377 |
| FBW (g) | 76.4 a,b | 69.4 b | 81.9 a | 2.41 | 0.016 |
| FBL (cm) | 18.0 a,b | 17.6 b | 18.5 a | 0.16 | 0.010 |
| SGR (%/day) | 2.17 a | 2.00 b | 2.23 a | 0.04 | 0.011 |
| DFI (g/100 g fish·day) | 1.57 | 1.62 | 1.57 | 0.02 | 0.267 |
| FCR | 0.90 a | 0.98 b | 0.88 a | 0.02 | 0.006 |
| Protein utilization | |||||
| PER | 2.49 a,b | 2.34 b | 2.58 a | 0.05 | 0.015 |
| PPV (%) | 49.2 | 48.6 | 51.8 | 1.23 | 0.196 |
| ADCprot (%) | 92.6 a | 81.0 b | 91.2 a | 1.05 | <0.0001 |
| Biometric indices | |||||
| CF (g/cm3) | 1.31 | 1.28 | 1.3 | 0.01 | 0.461 |
| HSI (%) | 1.26 | 1.44 | 1.29 | 0.07 | 0.244 |
| VSI (%) | 14.3 | 15.9 | 14.5 | 0.42 | 0.051 |
| Digestive Enzymes (U/mg Protein) | C | H18 | T18 | SEM | p-Value |
|---|---|---|---|---|---|
| Acidic proteases | 400.9 | 314.9 | 383.4 | 47.6 | 0.470 |
| Alkaline proteases | 82.2 b | 117.0 b | 263.5 a | 23.4 | 0.001 |
| Acidic–alkaline ratio | 6.36 a | 2.60 a,b | 1.49 b | 0.90 | 0.009 |
| Amylase | 81.5 b | 85.1 a,b | 139.5 a | 15.2 | 0.043 |
| Enzymes (mU/mg Protein) | C | H18 | T18 | SEM | p-Value |
|---|---|---|---|---|---|
| Fructose 1,6- biphosphatase (FBPase) | 26.6 | 22.8 | 27.7 | 4.49 | 0.724 |
| Pyruvate kinase (PK) | 55.1 | 50.2 | 64.9 | 6.84 | 0.346 |
| Glutamate pyruvate transaminase (GPT) | 329.3 | 371.5 | 333.2 | 30.1 | 0.568 |
| Glutamate oxaloacetate transaminase (GOT) | 270.3 | 169.3 | 326.3 | 40.1 | 0.059 |
| Glutamate dehydrogenase (GDH) | 574.6 | 379.4 | 523.0 | 65.9 | 0.151 |
| Immune Parameters | C | H18 | T18 | SEM | p-Value |
|---|---|---|---|---|---|
| TNF-α DI | 0.43 | 0.36 | 0.59 | 0.08 | 0.189 |
| TNF-α SM | 0.75 | 0.37 | 0.52 | 0.11 | 0.105 |
| Lysozyme | 4.76 | 5.03 | 3.95 | 0.47 | 0.288 |
| Esterase | 1014.0 | 1114.9 | 1451.5 | 228.9 | 0.405 |
| Acid phosphatase | 882.1 a | 791.2 a,b | 581.9 b | 53.9 | 0.009 |
| Alkaline phosphatase | 2350.4 | 2215.5 | 1780.8 | 291.6 | 0.392 |
| Anti-protease activity | 91.8 | 101.2 | 103.7 | 7.56 | 0.525 |
| Peroxidase | 0.58 | 0.69 | 0.47 | 0.08 | 0.223 |
| Total Immunoglobulins | 12.2 | 14.5 | 16.3 | 1.51 | 0.205 |
| Antioxidant Enzymes and Lipid Peroxidation | C | H18 | T18 | SEM | p-Value |
|---|---|---|---|---|---|
| Superoxide Dismutase (SOD) | 208.1 b | 209.7 b | 273.3 a | 15.3 | 0.023 |
| Catalase (CAT) | 186.5 a,b | 167.7 b | 216.7 a | 11.6 | 0.043 |
| Glutathione Peroxidase (GPx) | 12.2 a | 10.1 b | 9.85 b | 0.32 | 0.001 |
| Glutathione Reductase (GR) | 7.02 | 7.82 | 6.36 | 0.46 | 0.136 |
| Glucose-6-phosphate dehydrogenase (G6PDH) | 43.2 | 41.5 | 41.6 | 2.71 | 0.887 |
| Malondialdehyde (MDA) | 21.3 a | 15.7 a,b | 11.1 b | 2.40 | 0.043 |
| Fish welfare indicators | |||||
| Glucose | 3.33 | 3.69 | 3.23 | 0.19 | 0.319 |
| Lactate | 1.85 | 2.38 | 2.53 | 0.32 | 0.540 |
| Proximate Composition of the Fillet (Wet Basis) | C | H18 | T18 | SEM | p-Value |
|---|---|---|---|---|---|
| Moisture (%) | 77.6 a | 77.2 a,b | 76.4 b | 0.24 | 0.017 |
| Protein (%) | 18.2 b | 19.6 a | 19.5 a | 0.24 | 0.006 |
| Fat (%) | 1.09 | 1.87 | 1.52 | 0.21 | 0.070 |
| Ash (%) | 1.29 b | 1.42 a | 1.38 a | 0.01 | 0.0001 |
| Calcium (mg/Kg) | 325.4 | 418.0 | 400.0 | 46.2 | 0.365 |
| Phosphorus (g/Kg) | 2.98 | 2.92 | 2.96 | 0.06 | 0.813 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melenchón, F.; de Mercado, E.; Pula, H.J.; Cardenete, G.; Barroso, F.G.; Fabrikov, D.; Lourenço, H.M.; Pessoa, M.-F.; Lagos, L.; Weththasinghe, P.; et al. Fishmeal Dietary Replacement Up to 50%: A Comparative Study of Two Insect Meals for Rainbow Trout (Oncorhynchus mykiss). Animals 2022, 12, 179. https://doi.org/10.3390/ani12020179
Melenchón F, de Mercado E, Pula HJ, Cardenete G, Barroso FG, Fabrikov D, Lourenço HM, Pessoa M-F, Lagos L, Weththasinghe P, et al. Fishmeal Dietary Replacement Up to 50%: A Comparative Study of Two Insect Meals for Rainbow Trout (Oncorhynchus mykiss). Animals. 2022; 12(2):179. https://doi.org/10.3390/ani12020179
Chicago/Turabian StyleMelenchón, Federico, Eduardo de Mercado, Héctor J. Pula, Gabriel Cardenete, Fernando G. Barroso, Dmitri Fabrikov, Helena M. Lourenço, María-Fernanda Pessoa, Leidy Lagos, Pabodha Weththasinghe, and et al. 2022. "Fishmeal Dietary Replacement Up to 50%: A Comparative Study of Two Insect Meals for Rainbow Trout (Oncorhynchus mykiss)" Animals 12, no. 2: 179. https://doi.org/10.3390/ani12020179
APA StyleMelenchón, F., de Mercado, E., Pula, H. J., Cardenete, G., Barroso, F. G., Fabrikov, D., Lourenço, H. M., Pessoa, M.-F., Lagos, L., Weththasinghe, P., Cortés, M., & Tomás-Almenar, C. (2022). Fishmeal Dietary Replacement Up to 50%: A Comparative Study of Two Insect Meals for Rainbow Trout (Oncorhynchus mykiss). Animals, 12(2), 179. https://doi.org/10.3390/ani12020179