
Sperm Repository for a Breeding Program of the Eastern Oyster Crassostrea virginica: Sample Collection, Processing, Cryopreservation, and Data Management Plan

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
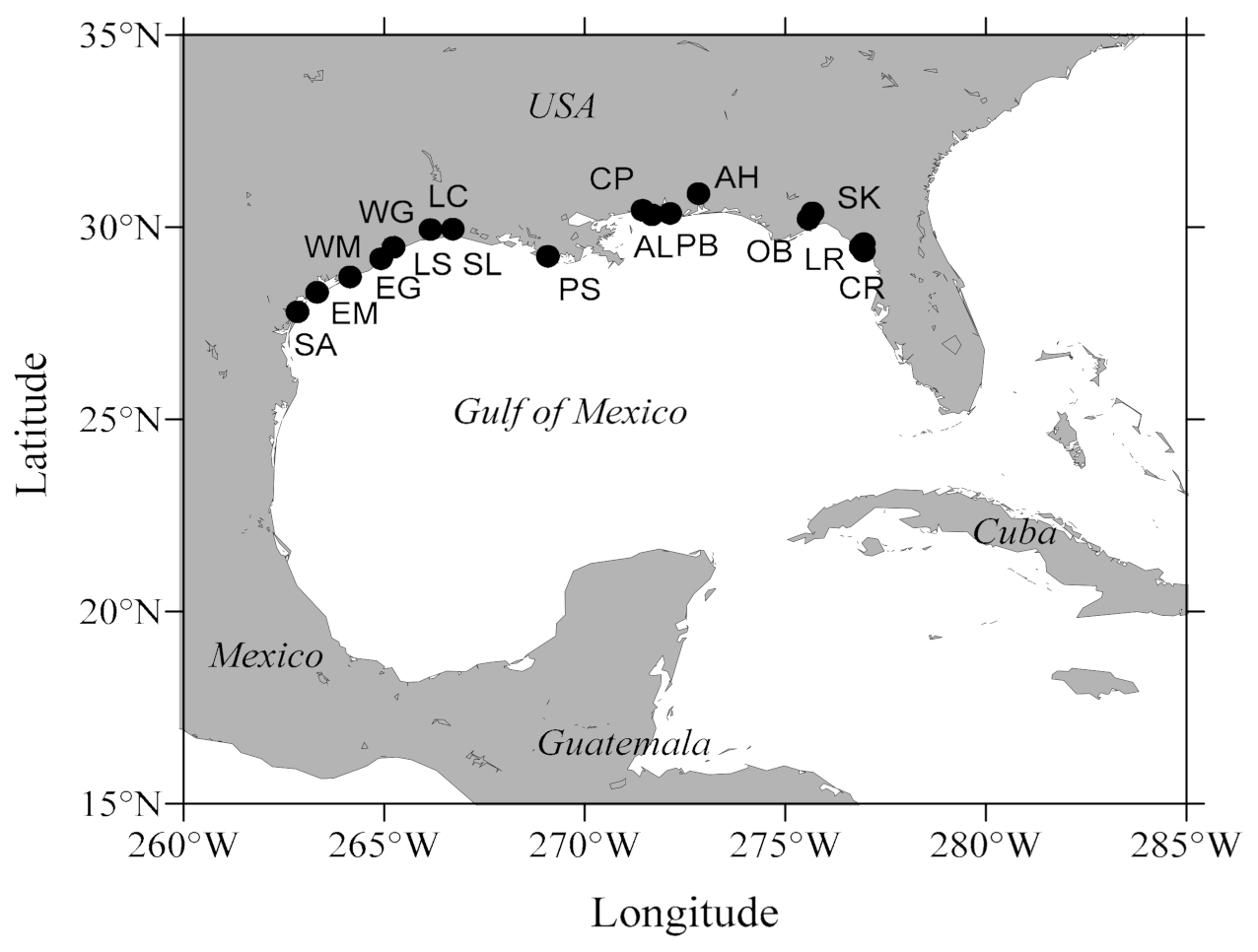
2.1. Broodstock Collection
2.2. Breeding Strategy
2.3. Sperm Collection
2.4. Determination of Sperm Concentration
2.5. Estimation of Fresh Sperm Motility and Fertility
2.6. Sperm Cryopreservation Process
2.7. Post-Thaw Sperm Quality Analysis
2.8. Data Analysis
3. Results
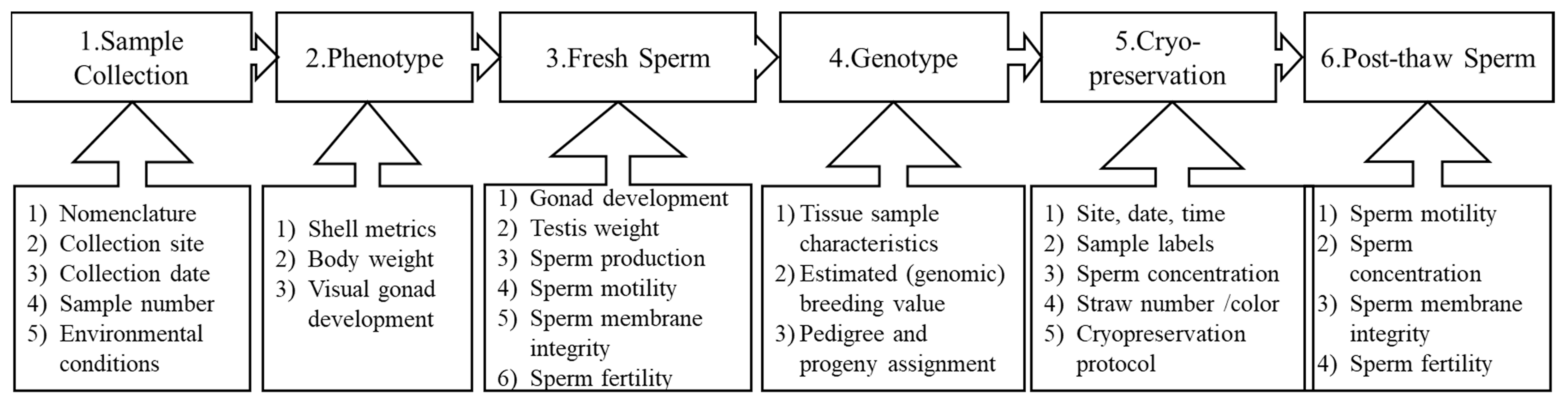
3.1. Data Management Plan in the Sperm Repository of Base Populations
- (1)
- Sample collection metadata
- (2)
- Phenotypic characteristics
- (3)
- Fresh sperm information
- (4)
- Linking to genetic and genomic information
- (5)
- Sperm cryopreservation
- (6)
- Post-thaw sperm information
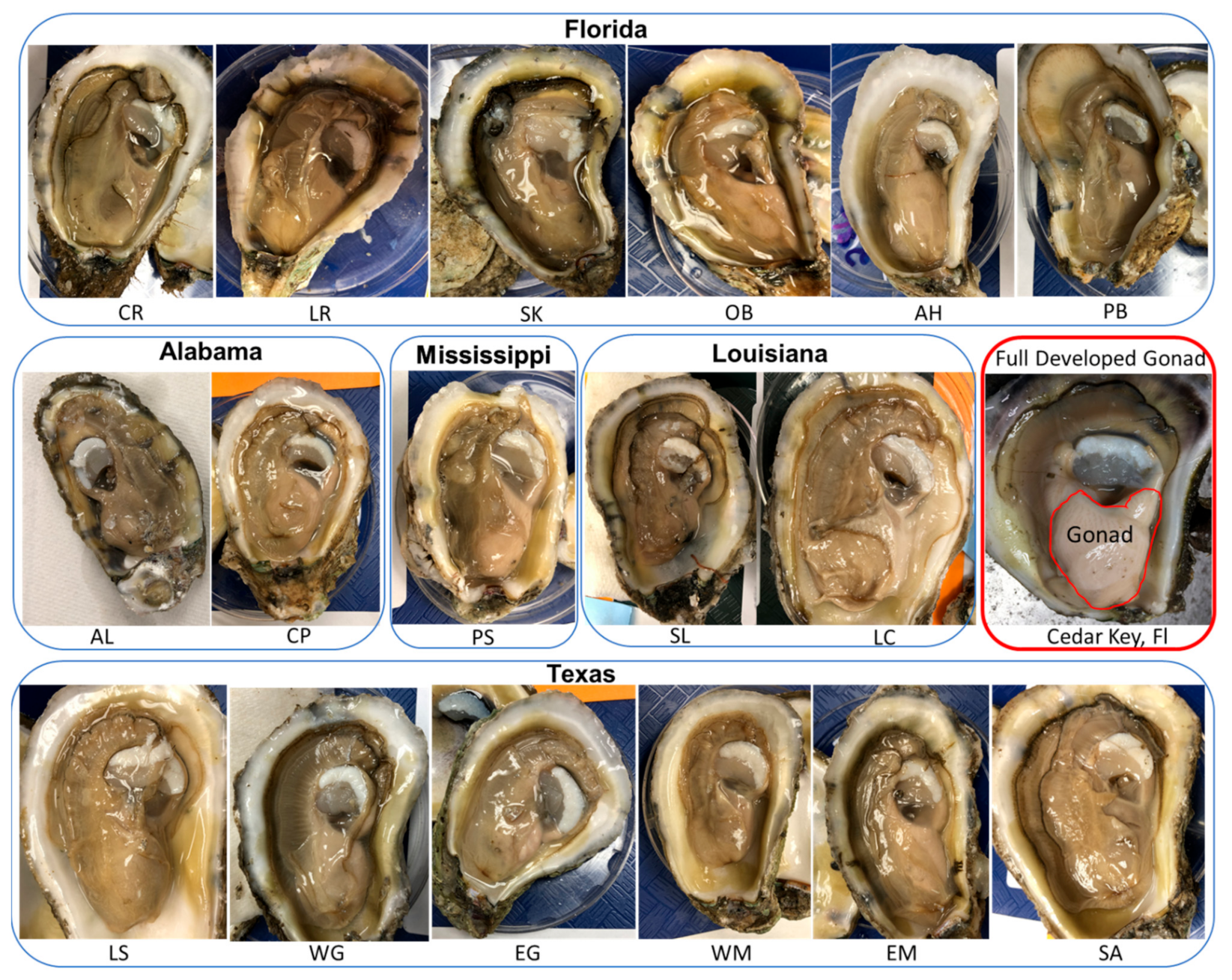
3.2. Phenotypic Characteristics
3.3. Fresh Sperm Production and Motility
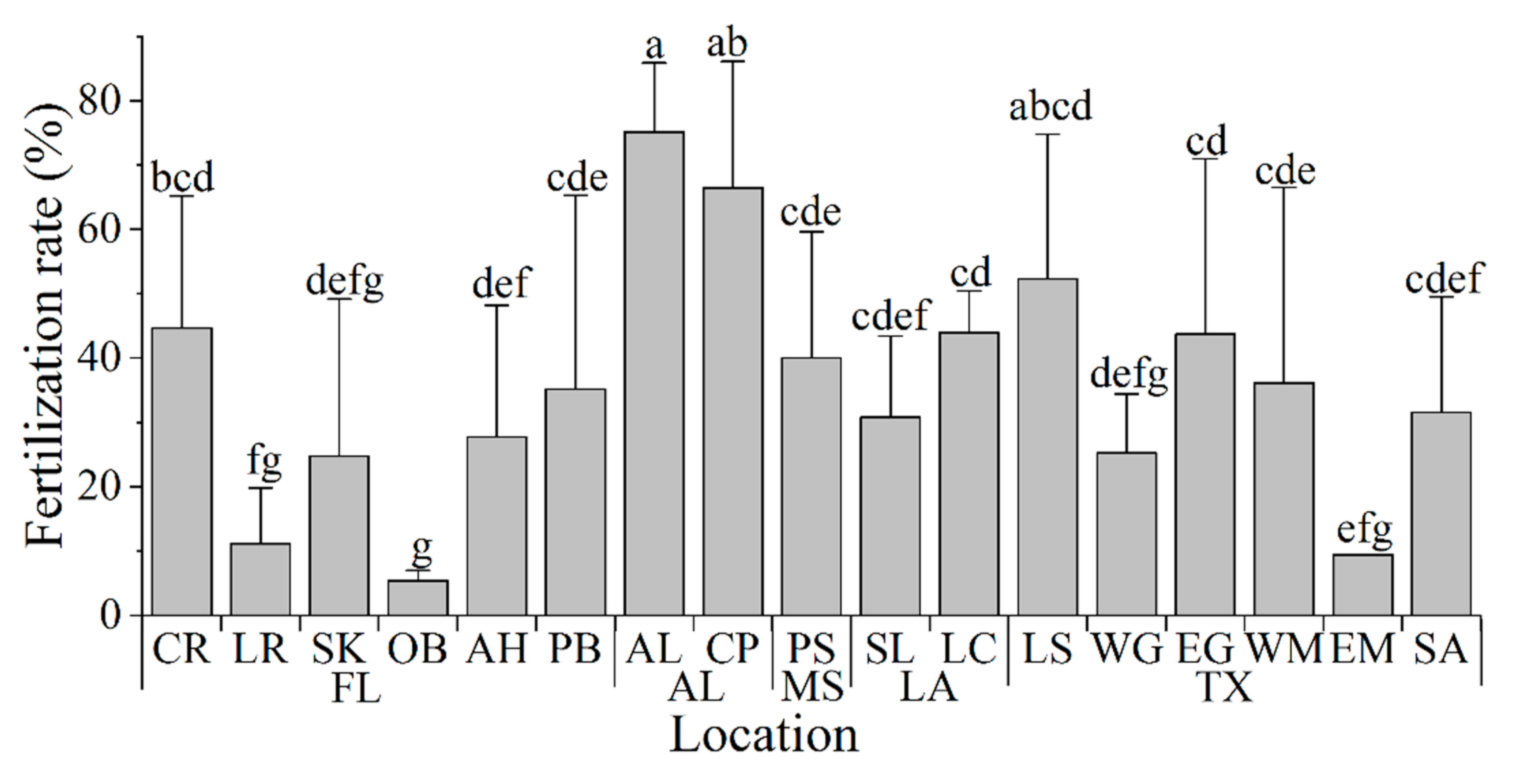
3.4. Fertility Test of Fresh Sperm
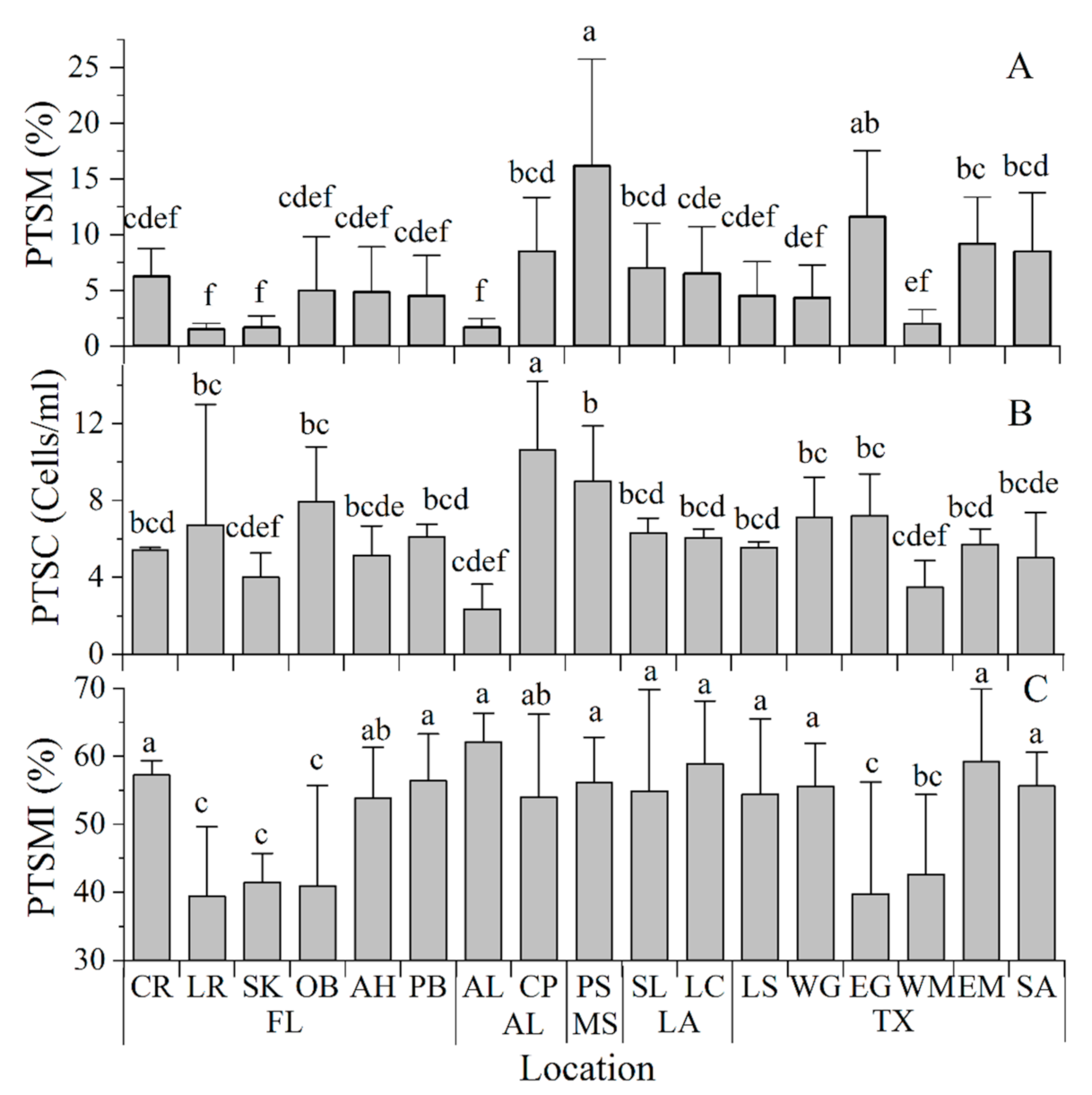
3.5. Post-Thaw Sperm Viabilities
3.6. Correlation between Parameters
4. Discussion
4.1. Strategies for Germplasm Collection for Germplasm Repositories
4.2. Data Management Plan for Oyster Sperm Repository
4.3. The Season for Oyster Sperm (Gamete) Collection and Cryopreservation
4.4. The Method to Collect Oyster Sperm (Gametes) in Oysters for Germplasm Repository
4.5. Streamlined Procedure for Sperm Sample Collection, Processing, and Cryopreservation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olivier, A.V.; Jones, L.; Le Vay, L.; Christie, M.; Wilson, J.; Malham, S.K. A global review of the ecosystem services provided by bivalve aquaculture. Rev. Aquac. 2020, 12, 3–25. [Google Scholar] [CrossRef] [Green Version]
- Galtsoff, P.S. The American Oyster, Crassostrea Virginica Gmelin, Fishery Bulletin Volume 64; United States Government Printing Office: Washington, DC, USA, 1964; p. 458.
- MacKenzie, C.L.J.; Burrell, V.G.J.; Rosenfield, A.; Hobart, W.L. The history, present condition, and future of the molluscan fisheries of North and Central America and Europe, Volume 1, Atlantic and Gulf Coasts. U.S. Dep. Commer. NOAA Tech. Rep. 1997, 127, 234. [Google Scholar]
- Schulte, D.M. History of the Virginia oyster fishery, Chesapeake Bay, USA. Front. Mar. Sci. 2017, 4, 127. [Google Scholar] [CrossRef] [Green Version]
- Pine, W.E.; Walters, C.J.; Camp, E.V.; Bouchillon, R.; Ahrens, R.; Sturmer, L.; Berrigan, M.E. The curious case of eastern oyster Crassostrea virginica stock status in Apalachicola Bay, Florida. Ecol. Soc. 2015, 20, 46. [Google Scholar] [CrossRef] [Green Version]
- Florida Fish and Wildlife Conservation Commission (FWRI). Florida’s Inshore and Nearshore Species: 2020 Status and Trends Report—Eastern Oyster. Available online: https://myfwc.com/media/26265/status-trends-oyster.pdf (accessed on 21 September 2021).
- Loosanoff, V.L.; Davis, H.C. Rearing of bivalve mollusks. In Advances in Marine Biology; Russell, F.S., Ed.; Academic Press: Cambridge, MA, USA, 1963; Volume 1, pp. 1–136. [Google Scholar]
- Davis, H.C.; Guillard, R.R. Relative value of ten genera of micro-organisms as food for oyster and clam larvae. Fish. Bull. 136 Fish. Bull. Fish Wildl. Serv. 1958, 58, 293–304. [Google Scholar]
- Guillard, R.R. Some factors in the use of nannoplankton cultures as food for larval and juvenile bivalves. Proc. Natl. Shellfish Assoc. 1957, 48, 134–142. [Google Scholar]
- Quayle, D.B.; Newkirk, G.F. Farming Bivalve Molluscs: Methods for Study and Development. In Advances in World Aquaculture; Sandifer, P.A., Ed.; The World Aquaculture Society: Baton Rouge, LA, USA, 1989; Volume 1, p. 294. [Google Scholar]
- Guo, X. Use and exchange of genetic resources in molluscan aquaculture. Rev. Aquac. 2009, 1, 251–259. [Google Scholar] [CrossRef]
- Guo, X.; Wang, Y.; DeBrosse, G.A.; Bushek, D.; Ford, S. Building a superior oyster for aquauclture. Jersey Shorel. 2008, 28, 7–9. [Google Scholar]
- Ragone Calvo, L.M.; Calvo, G.W.; Burreson, E.M. Dual disease resistance in a selectively bred eastern oyster, Crassostrea virginica, strain tested in Chesapeake Bay. Aquaculture 2003, 220, 69–87. [Google Scholar] [CrossRef]
- Jenderek, M.M.; Reed, B.M. Cryopreserved storage of clonal germplasm in the USDA National Plant Germplasm System. Vitr. Cell. Dev. Biol. Plant 2017, 53, 299–308. [Google Scholar] [CrossRef]
- Blackburn, H. The National Animal Germplasm Program: Challenges and opportunities for poultry genetic resources. Poult. Sci. 2006, 85, 210–215. [Google Scholar] [CrossRef]
- Lannan, J.E. Experimental self-fertilization of Pacific oysters, Crassostrea gigas, utilizing cryopreserved sperm. Genetics 1971, 68, 599–601. [Google Scholar] [CrossRef]
- Yang, H. Application of germplasm preservation in breeding programs for molluscan shellfish aquaculture and restoration. Bull. Jpn. Fish. Res. Educ. Agency 2017, 45, 15–20. [Google Scholar]
- Hassan, M.M.; Qin, J.; Li, X. Sperm cryopreservation in oysters: A review of its current status and potentials for future application in aquaculture. Aquaculture 2015, 438, 24–32. [Google Scholar] [CrossRef]
- Liu, Y.; Li, X.; Robinson, N.; Qin, J. Sperm cryopreservation in marine mollusk: A review. Aquac. Int. 2015, 23, 1505–1524. [Google Scholar] [CrossRef]
- Yang, H.; Huo, Y. Review of molluscan larval cryopreservation and application to germplasm cryobanking and commercial seed production. Aquaculture 2021, 547, 737491. [Google Scholar] [CrossRef]
- Hughes, J.B. An examination of eggs challenged with cryopreserved spermatozoa of American oyster Crassostrea virginica. Cryobiology 1973, 10, 342–344. [Google Scholar] [CrossRef]
- Zell, S.R.; Bamford, M.H.; Hidu, H. Cryopreservation of spermatozoa of the American oyster Crassostrea virginica Gmelin. Cryobiology 1979, 16, 448–460. [Google Scholar] [CrossRef]
- Paniagua-Chavez, C.G.; Tiersch, T.R. Laboratory studies of cryopreservation of sperm and trochophore larvae of the eastern oyster. Cryobiology 2001, 43, 211–223. [Google Scholar] [CrossRef]
- Yang, H.; Hu, E.; Cuevas-Uribe, R.; Supan, J.; Guo, X.; Tiersch, T.R. High-throughput sperm cryopreservation of eastern oyster Crassostrea virginica. Aquaculture 2012, 344, 223–230. [Google Scholar] [CrossRef]
- Paniagua-Chavez, C.G.; Buchanan, J.T.; Supan, J.E.; Tiersch, T.R. Settlement and growth of eastern oysters produced from cryopreserved larvae. Cryo Lett. 1998, 19, 283–292. [Google Scholar]
- Yang, H.; Wang, Y.; Guo, X.; Tiersch, T.R. Production of inbred larvae through self-fertilization using oocytes and cryopreserved sperm from the same individuals after sex reversal in eastern oyster Crassostrea virginica. Aquac. Res. 2015, 46, 2153–2165. [Google Scholar] [CrossRef]
- Brown, A.; Marshall, D. A basic sampling strategy: Theory and practice. In Collecting Plant Genetic Diversity: Technical Guidelines; CAB Int.: Wallingford, UK, 1995; Volume 75, p. 91. [Google Scholar]
- Franco-Duran, J.; Crossa, J.; Chen, J.F.; Hearne, S.J. The impact of sample selection strategies on genetic diversity and representativeness in germplasm bank collections. BMC Plant Biol. 2019, 19, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Hanks, J.H. Hanks’ balanced salt solution and pH control. Methods Cell Sci. 1975, 1, 1–4. [Google Scholar] [CrossRef]
- FAO. The State of the World’s Aquatic Genetic Resources for Food and Agriculture; FAO: Rome, Italy, 2019; p. 251. [Google Scholar]
- Witzenberger, K.A.; Hochkirch, A. Ex situ conservation genetics: A review of molecular studies on the genetic consequences of captive breeding programmes for endangered animal species. Biodivers. Conserv. 2011, 20, 1843–1861. [Google Scholar] [CrossRef]
- Kasso, M.; Balakrishnan, M. Ex situ conservation of biodiversity with particular emphasis to Ethiopia. Int. Sch. Res. Not. 2013, 2013, 985037. [Google Scholar] [CrossRef] [Green Version]
- Reed, B.M.; Engelmann, F.; Dulloo, E.; Engels, J.M.M. Technical Guidelines for the Management of Field and In Vitro Germplasm Collections; Bioversity International: Rome, Italy, 2004; p. 95. [Google Scholar]
- Wilkinson, M.D.; Dumontier, M.; Aalbersberg, I.J.; Appleton, G.; Axton, M.; Baak, A.; Blomberg, N.; Boiten, J.-W.; da Silva Santos, L.B.; Bourne, P.E.; et al. The FAIR Guiding Principles for scientific data management and stewardship. Sci. Data 2016, 3, 160018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. Commission on Genetic Resources for Food and Agriculture: Innovations in Cryoconservation of Animal Genetic Resources—Draft Technical Guidelines. 2021, p. 290. Available online: http://www.fao.org/3/nf388/nf388.pdf (accessed on 21 September 2021).
- Imarhiagbe, O.; Osazee, J.; Aiwansoba, R.; Shittu, O. In vitro germplasm collection and storage: A review. Int. J. Biol. Res. 2016, 1, 9–14. [Google Scholar]
- Cabrita, E.; Martínez-Páramo, S.; Gavaia, P.J.; Riesco, M.F.; Valcarce, D.G.; Sarasquete, C.; Herráez, M.P.; Robles, V. Factors enhancing fish sperm quality and emerging tools for sperm analysis. Aquaculture 2014, 432, 389–401. [Google Scholar] [CrossRef]
- Sandoval-Vargas, L.; Jimenez, M.S.; Gonzalez, J.R.; Villalobos, E.F.; Cabrita, E.; Isler, I.V. Oxidative stress and use of antioxidants in fish semen cryopreservation. Rev. Aquac. 2021, 13, 365–387. [Google Scholar] [CrossRef]
- Martinez-Montero, M.E.; Harding, K. Cryobionomics: Evaluating the concept in plant cryopreservation. In PlantOmics: The Omics of Plant Science; Barh, D., Khan, M.S., Davies, E., Eds.; Springer India: New Delhi, India, 2015; pp. 655–682. [Google Scholar]
- Riesco, M.F.; Robles, V. Cryopreservation causes genetic and epigenetic changes in zebrafish genital ridges. PLoS ONE 2013, 8, e67614. [Google Scholar] [CrossRef] [PubMed]
- Gosling, E. Reproduction, Settlement and Recruitment. In Bivalve Molluscs Biology, Ecology and Culture; Fishing News Books: Malden, MA, USA, 2003; pp. 131–168. [Google Scholar]
- Mackie, G.L. Chapter 5—Bivalves. In The Mollusca, Volume 7 Reproduction; Tompa, A.S., Verdonk, N.H., Van Den Biggelaar, J.A.M., Eds.; Academic Press Inc.: Orlando, FL, USA, 1984; pp. 351–418. [Google Scholar]
- Loosanoff, V.L.; Davis, H.C. Temperature requirements for maturation of gonads of northern oysters. Biol. Bull. 1952, 103, 80–96. [Google Scholar] [CrossRef]
- Mathieu, M.; Lubet, P. Storage tissue metabolism and reproduction in marine bivalves—A brief review. Invertebr. Reprod. Dev. 1993, 23, 123–129. [Google Scholar] [CrossRef]
- Galtsoff, P.S. Physiology of reproduction in molluscs. Am. Zool. 1961, 1, 273–289. [Google Scholar] [CrossRef]
- Loosanoff, V.L. Seasonal gonadal changes in the adult oysters, Ostrea virginica, of long island sound. Biol. Bull. 1942, 82, 195–206. [Google Scholar] [CrossRef]
- Kennedy, A.V.; Battle, H.I. Cyclic changes in the gonad of the american oyster, Crassostrea virginica (Gmelin). Can. J. Zool. 1964, 42, 305–321. [Google Scholar] [CrossRef]
- Yang, H.; Supan, J.; Guo, X.; Tiersch, T.R. Nonlethal sperm collection and cryopreservation in the eastern oyster Crassostrea virginica. J. Shellfish Res. 2013, 32, 429–437. [Google Scholar] [CrossRef]
- Aji, L.P. Review: Spawning induction in bivalve. J. Penelit. Sains 2011, 14, 33–36. [Google Scholar]
- Galtsoff, P.S. Physiology of reproduction of Ostrea virginica II. Stimulation of spawning in the female oyster. Biol. Bull. 1938, 75, 286–307. [Google Scholar] [CrossRef]
- Galtsoff, P.S. Physiology of reproduction of Ostrea virginica III. Stimulation of spawning in the male oyster. Biol. Bull. 1940, 78, 117–135. [Google Scholar] [CrossRef]
- Imai, T.; Hatanaka, M.; Sato, R.; Sakai, S.; Yûki, R. Artificial breeding of oysters in tanks. Tohoku J. Agric. Res. 1950, 1, 69–86. [Google Scholar]
- Huo, Y.; Yee, J.C.; Yang, H. Development of a cost-effective and portable aeration freezing system for germplasm cryopreservation for eastern oysters and other aquatic species. Aquac. Res. 2021. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium | Cryoprotectant | Equilibration | Container | Cooling Rate | Thawing | Fertilization | Post-Thaw Survival | Reference | |
|---|---|---|---|---|---|---|---|---|---|
| Sperm | Seawater pH = 7.0 | DMSO at 5% and 10% | NA | 2-mL ampoule | L °C /min from 0 to −8 °C, at 5.5 °C/min to −25 °C, then LN | 21 °C | 34 mL post-thaw sperm plus million oocytes in 250 mL | 1–5% post-thaw motility and 2% fertilization | [21] |
| 2.6× HBSS | DMSO 8% with 80 mM glycine, 55 mM NaHCO3 | 20 min at 0 °C | 0.25-mL plastic straws | 5 °C/min to −5, −20, −40, or −80 °C, then plunged into LN | 55–60 °C water bath | 0.25-mL sperm to 200–900 eggs in 300 mL seawater | 7–91% post-thaw fertility | [22] | |
| ASW or Ca-free HBSS | 0, 5, 10, 15, 20, and 25% (v/v) PG with/without 0.25 M sucrose | 20 min at 21 °C | 5-mL macrotubes | 2.5 °C/min to −30 °C, and then plunged LN | 70 °C for 15 s, 25 °C until thawing | Diluted in Ca-free HBSS at 1:1 | 57% post-thaw fertility | [23] | |
| Ca-free HBSS pH = 7.2 | Methanol, DMSO, and PG at 10% | 10, 20, 30, 40, 50 and 60 min, mixing at 1:1 | 0.5-mL of French and CBS™ straw | 1, 5, 10, 15, 20, 25, 30, and 40 °C/min from 5 to −80 °C | 30, 40, and 50 °C water bath | 2-mL post thaw sperm mix with 50-mL oocytes at 10,000/mL | 18–95% post-thaw fertilization 10% DMSO | [24] | |
| Ca-free HBSS, pH = 7.2 | 10% DMSO | 20 min after mixing at 1:1 | 0.5-mL French straw | 15 °C/min from 5 to −80 °C, then to LN | 40 °C of water bath 6 s | 3–72% fertility; self-fertilization 0–43% | 8 selfing families confirmed | [26] | |
| Swimming Larvae | ASW | 15% PG with 0.25 M Sucrose | 20 min at 21 °C | 5-mL macrotubes | 2.5 °C/min from 15 to −30 °C, hold for 5 min, then to LN | 70 °C for 15 s water bath | NA | 4 months old (850 spat) | [25] |
| HBSS | 5, 10, 15, 20, 25% PG with 0.25 M Sucrose | 20 min at 21 °C | 5-mL macrotubes | 2.5 °C/min from 15 to −30 °C, hold for 5 min, then to LN | 70 °C for 15 s water bath | D-larvae | 24% D-larvae rate | [23] |
| State | Location | Abb | Height (mm) | Length (mm) | Width (mm) | Weight (g) | Testis Weight (g) | Gonadosomatic Index (%) | Sperm Volume (mL) | Sperm Concentration (×109 cells/mL) | Sperm Production (×1010 cells) | Fresh Sperm Motility (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FL | Corrigan Reef | CR | 93.37 ± 6.57 | 51.01 ± 3.74 | 28.63 ± 4.32 | 93.86 ± 12.11 | 1.52 ± 0.00 | 0.75 ± 0.42 | 9.67 ± 4.92 | 0.09 ± 0.05 | 0.10 ± 0.11 | 11 ± 8 e |
| Lone Cabbage Reef | LR | 94.43 ± 8.43 | 51.42 ± 4.75 | 25.36 ± 1.83 | 78.58 ± 7.19 | 1.56 ± 0.41 | 1.00 ± 0.32 | 13.33 ± 6.32 | 0.60 ± 0.36 | 0.91 ± 0.90 | 23 ± 11 c,d,e | |
| Seahorse Key | SK | 87.37 ± 5.50 | 50.72 ± 2.84 | 30.52 ± 3.07 | 80.53 ± 9.63 | 1.50 ± 0.70 | 0.70 ± 0.27 | 11.75 ± 4.57 | 0.27 ± 0.13 | 0.31 ± 0.17 | 14 ± 4 d,e | |
| Oyster Bay | OB | 90.26 ± 10.56 | 56.48 ± 5.74 | 30.09 ± 5.78 | 106.30 ± 36.27 | 3.18 ± 1.57 | 2.50 ± 0.71 | 22.42 ± 9.08 | 0.73 ± 0.49 | 1.59 ± 1.07 | 70 ± 20 a | |
| Alligator Harbor | AH | 85.68 ± 11.75 | 48.38 ± 3.63 | 29.01 ± 3.69 | 79.49 ± 25.00 | 2.33 ± 1.26 | 2.00 ± 0.71 | 15.58 ± 6.25 | 1.24 ± 0.63 | 1.94 ± 1.09 | 50 ± 24 a,b,c,d | |
| Pensacola Bay | PB | 80.70 ± 7.33 | 46.57 ± 7.66 | 27.43 ± 7.89 | 72.66 ± 23.28 | 1.62 ± 0.94 | 1.17 ± 0.26 | 12.67 ± 6.05 | 0.44 ± 0.22 | 0.47 ± 0.18 | 21 ± 22 d,e | |
| AL | Alonzo Landing | AL | 88.41 ± 6.69 | 54.44 ± 4.58 | 27.84 ± 4.90 | 78.82 ± 11.09 | 1.39 ± 0.44 | / | 11.67 ± 3.40 | 1.40 ± 0.17 | 1.61 ± 0.43 | 43 ± 29 a,b,c,d,e |
| Cedar Point | CP | 85.48 ± 9.16 | 63.28 ± 2.98 | 34.61 ± 5.65 | 152.11 ± 33.08 | 2.69 ± 0.57 | 3.25 ± 1.33 | 15.08 ± 2.87 | 2.01 ± 1.12 | 3.11 ± 1.83 | 47 ± 5 a,b,c,d,e | |
| MS | Pascagoula | PS | 80.33 ± 5.00 | 53.63 ± 5.10 | 30.74 ± 5.05 | 105.95 ± 28.91 | 3.54 ± 1.71 | 3.25 ± 0.76 | 20.00 ± 7.27 | 1.39 ± 0.50 | 2.82 ± 1.52 | 60 ± 27 a,b,c |
| LA | Sister Lake | SL | 111.87 ± 13.75 | 73.40 ± 8.34 | 42.31 ± 4.80 | 280.50 ± 112.44 | 2.09 ± 1.22 | 1.00 ± 0.45 | 13.58 ± 8.69 | 0.33 ± 0.24 | 0.43 ± 0.34 | 24 ± 19 b,c,d,e |
| Lake Calcasieu | LC | 91.96 ± 14.67 | 67.64 ± 11.01 | 40.93 ± 6.58 | 211.38 ± 87.83 | 2.11 ± 0.86 | 2.25 ± 0.29 | 18.08 ± 3.67 | 0.67 ± 0.13 | 1.19 ± 0.30 | 40 ± 0 a,b,c,d,e | |
| TX | Lake Sabine | LS | 92.51 ± 11.17 | 60.67 ± 5.45 | 43.40 ± 10.45 | 221.75 ± 88.11 | 1.55 ± 0.81 | 1.08 ± 0.38 | 13.08 ± 4.34 | 0.60 ± 0.25 | 0.78 ± 0.40 | 62 ± 13 a,b |
| West Galveston | WG | 96.31 ± 16.45 | 62.71 ± 6.23 | 39.90 ± 7.25 | 198.19 ± 80.14 | 2.00 ± 0.61 | 1.83 ± 0.88 | 10.67 ± 2.66 | 0.29 ± 0.14 | 0.30 ± 0.14 | 22 ± 12 c,d,e | |
| East Galveston | EG | 83.95 ± 5.86 | 57.40 ± 5.95 | 35.13 ± 4.29 | 152.51 ± 27.26 | 3.27 ± 1.08 | 2.75 ± 0.87 | 22.25 ± 4.12 | 0.73 ± 0.18 | 1.60 ± 0.40 | 73 ± 5 a | |
| West Matagorda | WM | 89.44 ± 6.52 | 62.36 ± 7.09 | 41.57 ± 5.08 | 169.60 ± 28.76 | 1.75 ± 0.54 | 1.17 ± 0.26 | 12.00 ± 3.03 | 0.21 ± 0.15 | 0.23 ± 0.15 | 24 ± 22 b,c,d,e | |
| East Matagorda | EM | 92.30 ± 14.47 | 52.70 ± 11.46 | 33.01 ± 4.07 | 115.33 ± 17.07 | 1.19 ± 0.32 | 1.50 ± 0.89 | 15.50 ± 5.15 | 0.58 ± 0.44 | 0.94 ± 0.78 | 41 ± 34 a,b,c,d,e | |
| San Antonio | SA | 92.02 ± 11.50 | 66.97 ± 6.13 | 42.34 ± 2.99 | 213.17 ± 65.58 | 2.03 ± 1.55 | 1.17 ± 0.52 | 13.00 ± 1.87 | 0.41 ± 0.14 | 0.53 ± 0.18 | 27 ± 16 b,c,d,e | |
| Mean ± SD | 90.11 ± 7.29 | 57.80 ± 7.57 | 34.16 ± 6.31 | 141.21 ± 64.46 | 2.08 ± 0.71 | 1.77 ± 0.95 | 14.44 ± 3.39 | 0.68 ± 0.52 | 1.05 ± 0.89 | 38 ± 20 |
| Parameters | Phenotypes | Fresh Sperm Quantity and Quality | Post-Thaw Sperm Viability | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Height | Length | Width | Weight | Testis Weight | Sperm Production | Motility | Fertilization Rate | Motility | SMI | Sperm Concentration | |
| Height | 1 | ||||||||||
| Length | 0.436 | 1 | |||||||||
| Width | 0.419 | 0.687 | 1 | ||||||||
| Weight | 0.658 | 0.814 | 0.838 | 1 | |||||||
| Testis weight | 0.050 | 0.111 | 0.084 | 0.148 | 1 | ||||||
| Sperm Production | −0.256 | 0.005 | −0.158 | −0.122 | 0.463 | 1 | |||||
| Fresh sperm motility | −0.213 | −0.041 | −0.017 | −0.005 | 0.405 | 0.550 | 1 | ||||
| Fertilization rate | −0.148 | 0.081 | 0.048 | 0.000 | −0.083 | 0.127 | −0.003 | 1 | |||
| Post-thaw sperm motility | −0.177 | 0.071 | 0.058 | 0.057 | 0.368 | 0.406 | 0.446 | −0.007 | 1 | ||
| Post-thaw SMI | 0.097 | 0.079 | 0.126 | 0.150 | −0.013 | 0.138 | 0.061 | 0.165 | 0.303 | 1 | |
| Post-thaw sperm concentration | −0.092 | 0.107 | −0.040 | 0.044 | 0.186 | 0.355 | 0.183 | 0.066 | 0.295 | −0.042 | 1 |
| Height | <0.001 | ||||||||||
| Length | <0.001 | <0.001 | |||||||||
| Width | <0.001 | <0.001 | <0.001 | ||||||||
| Weight | <0.001 | <0.001 | <0.001 | <0.001 | |||||||
| Testis weight | 0.658 | 0.323 | 0.455 | 0.187 | <0.001 | ||||||
| Sperm Production | 0.021 | 0.963 | 0.158 | 0.279 | <0.001 | <0.001 | |||||
| Fresh sperm motility | 0.057 | 0.716 | 0.880 | 0.967 | <0.001 | <0.001 | <0.001 | ||||
| Fertilization rate | 0.188 | 0.474 | 0.673 | 0.997 | 0.460 | 0.260 | 0.977 | <0.001 | |||
| Post-thaw sperm motility | 0.115 | 0.528 | 0.609 | 0.613 | <0.001 | <0.001 | <0.001 | 0.952 | <0.001 | ||
| Post-thaw SMI | 0.390 | 0.484 | 0.262 | 0.180 | 0.910 | 0.219 | 0.587 | 0.141 | 0.006 | <0.001 | |
| Post-thaw sperm concentration | 0.412 | 0.343 | 0.724 | 0.694 | 0.097 | 0.001 | 0.102 | 0.560 | 0.008 | 0.710 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Huo, Y.; Yee, J.C.; Rikard, S.; Walton, W.C.; Saillant, E. Sperm Repository for a Breeding Program of the Eastern Oyster Crassostrea virginica: Sample Collection, Processing, Cryopreservation, and Data Management Plan. Animals 2021, 11, 2836. https://doi.org/10.3390/ani11102836
Yang H, Huo Y, Yee JC, Rikard S, Walton WC, Saillant E. Sperm Repository for a Breeding Program of the Eastern Oyster Crassostrea virginica: Sample Collection, Processing, Cryopreservation, and Data Management Plan. Animals. 2021; 11(10):2836. https://doi.org/10.3390/ani11102836
Chicago/Turabian StyleYang, Huiping, Yuanzi Huo, Jayme C. Yee, Scott Rikard, William C. Walton, and Eric Saillant. 2021. "Sperm Repository for a Breeding Program of the Eastern Oyster Crassostrea virginica: Sample Collection, Processing, Cryopreservation, and Data Management Plan" Animals 11, no. 10: 2836. https://doi.org/10.3390/ani11102836
APA StyleYang, H., Huo, Y., Yee, J. C., Rikard, S., Walton, W. C., & Saillant, E. (2021). Sperm Repository for a Breeding Program of the Eastern Oyster Crassostrea virginica: Sample Collection, Processing, Cryopreservation, and Data Management Plan. Animals, 11(10), 2836. https://doi.org/10.3390/ani11102836