
Relevance of Biofilm Models in Periodontal Research: From Static to Dynamic Systems

 ,
,  , ,
, ,  and
and 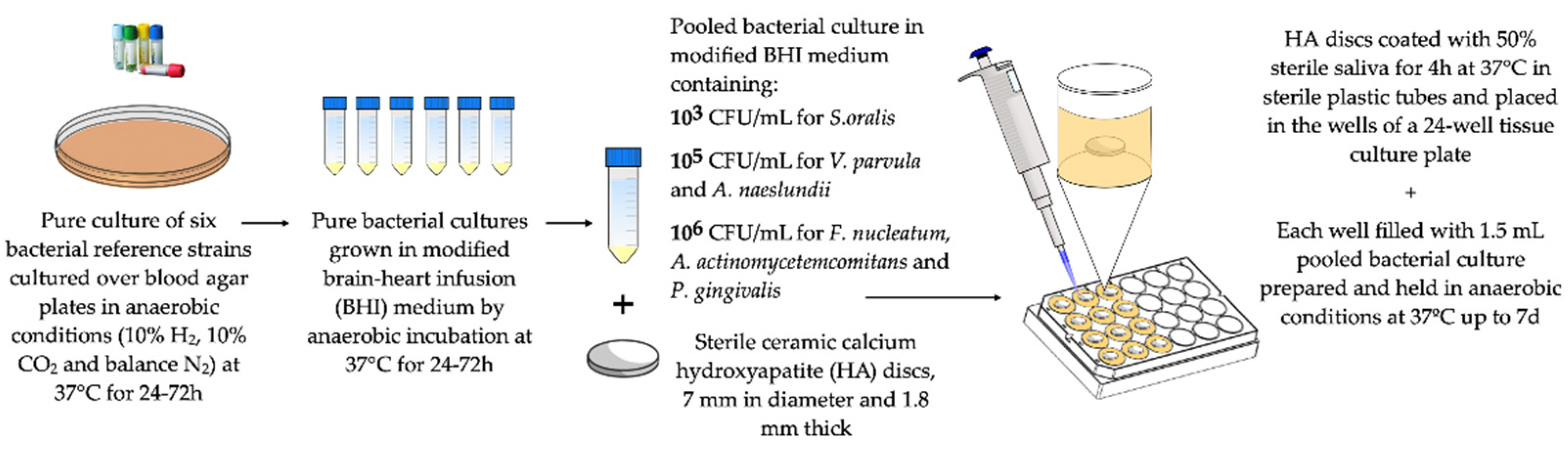{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Oral Biofilms and In Vitro Biofilm Models
Development of a Static/Dynamic Multi-Species Subgingival In Vitro Biofilm Model
3. Antimicrobial Strategies Evaluated in In Vitro Biofilm Models
4. Evaluation of New Delivery Formats of Oral Antiseptics Using In Vitro Biofilm Models
5. Transcriptomic Analysis and Protein Differential Expression in Biofilm Growth Using In Vitro Biofilm Models
6. Study of the Impact of the Material Surface and Topography of Dental Implants on Peri-Implant Biofilm Development
6.1. Static Multi-Species In Vitro Biofilm Model Over Implant-Material Discs
6.2. A Static, Multi-Species, In Vitro Biofilm Model Over Whole Dental Implants
6.3. Development of an In Vitro Multi-Species Dynamic Model to Study the Impact of Dental Implant Surface and Topography on Bacterial Biofilms
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Do, T.; Devine, D.; Marsh, P.D. Oral biofilms: Molecular analysis, challenges, and future prospects in dental diagnostics. Clin. Cosmet. Investig. Dent. 2013, 5, 11–19. [Google Scholar] [CrossRef]
- Samaranayake, L.; Matsubara, V.H. Normal Oral Flora and the Oral Ecosystem. Dent. Clin. N. Am. 2017, 61, 199–215. [Google Scholar] [CrossRef]
- Heidrich, M.; Kuhnel, M.P.; Kellner, M.; Lorbeer, R.A.; Lange, T.; Winkel, A.; Stiesch, M.; Meyer, H.; Heisterkamp, A. 3D imaging of biofilms on implants by detection of scattered light with a scanning laser optical tomograph. Biomed. Opt. Express 2011, 2, 2982–2994. [Google Scholar] [CrossRef]
- Marrie, T.J.; Nelligan, J.; Costerton, J.W. A scanning and transmission electron microscopic study of an infected endocardial pacemaker lead. Circulation 1982, 66, 1339–1341. [Google Scholar] [CrossRef] [PubMed]
- Parsek, M.R.; Singh, P.K. Bacterial biofilms: An emerging link to disease pathogenesis. Annu. Rev. Microbiol. 2003, 57, 677–701. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.G.; O’Toole, G.A. Innate and induced resistance mechanisms of bacterial biofilms. Curr. Top. Microbiol. Immunol. 2008, 322, 85–105. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Del Pozo, J.L. Biofilm-related disease. Expert Rev. Anti-Infect. Ther. 2018, 16, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Darouiche, R.O. Treatment of infections associated with surgical implants. N. Engl. J. Med. 2004, 350, 1422–1429. [Google Scholar] [CrossRef]
- Hooton, T.M.; Bradley, S.F.; Cardenas, D.D.; Colgan, R.; Geerlings, S.E.; Rice, J.C.; Saint, S.; Schaeffer, A.J.; Tambayh, P.A.; Tenke, P.; et al. Diagnosis, prevention, and treatment of catheter-associated urinary tract infection in adults: 2009 International Clinical Practice Guidelines from the Infectious Diseases Society of America. Clin. Infect. Dis. 2010, 50, 625–663. [Google Scholar] [CrossRef] [PubMed]
- Wanner, O.; Reichert, P. Mathematical modeling of mixed-culture biofilms. Biotechnol. Bioeng. 1996, 49, 172–184. [Google Scholar] [CrossRef]
- Kim, S.K.; Lee, J.H. Biofilm dispersion in Pseudomonas aeruginosa. J. Microbiol. 2016, 54, 71–85. [Google Scholar] [CrossRef]
- Klausen, M.; Gjermansen, M.; Kreft, J.U.; Tolker-Nielsen, T. Dynamics of development and dispersal in sessile microbial communities: Examples from Pseudomonas aeruginosa and Pseudomonas putida model biofilms. FEMS Microbiol. Lett. 2006, 261, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Dowd, S.E.; Smith, E.; Rhoads, D.D.; Wolcott, R.D. In vitro multispecies Lubbock chronic wound biofilm model. Wound Repair Regen. 2008, 16, 805–813. [Google Scholar] [CrossRef]
- Kojic, E.M.; Darouiche, R.O. Candida infections of medical devices. Clin. Microbiol. Rev. 2004, 17, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Horn, H.; Lackner, S. Modeling of biofilm systems: A review. Adv. Biochem. Eng. Biotechnol. 2014, 146, 53–76. [Google Scholar] [CrossRef] [PubMed]
- Williamson, K.; McCarty, P.L. Verification Studies of the Biofilm Model for Bacterial Substrate Utilization. Water Pollut. Control Fed. 1976, 48, 281–296. [Google Scholar]
- Harremoës, P. The Significance of Pore Diffusion to Filter Denitrification. Water Pollut. Control Fed. 1976, 48, 377–388. [Google Scholar]
- Rittmann, B.E.; Boltz, J.P.; Brockmann, D.; Daigger, G.T.; Morgenroth, E.; Sorensen, K.H.; Takacs, I.; van Loosdrecht, M.; Vanrolleghem, P.A. A framework for good biofilm reactor modeling practice (GBRMP). Water Sci. Technol. 2018, 77, 1149–1164. [Google Scholar] [CrossRef]
- Hauduc, H.; Rieger, L.; Oehmen, A.; van Loosdrecht, M.C.; Comeau, Y.; Heduit, A.; Vanrolleghem, P.A.; Gillot, S. Critical review of activated sludge modeling: State of process knowledge, modeling concepts, and limitations. Biotechnol. Bioeng. 2013, 110, 24–46. [Google Scholar] [CrossRef]
- Rittmann, B.E.; Manem, J.A. Development and experimental evaluation of a steady-state, multispecies biofilm model. Biotechnol. Bioeng. 1992, 39, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Mattei, M.R.; Frunzo, L.; D’Acunto, B.; Pechaud, Y.; Pirozzi, F.; Esposito, G. Continuum and discrete approach in modeling biofilm development and structure: A review. J. Math. Biol. 2018, 76, 945–1003. [Google Scholar] [CrossRef] [PubMed]
- McBain, A.J. Chapter 4: In vitro biofilm models: An overview. Adv. Appl. Microbiol. 2009, 69, 99–132. [Google Scholar] [CrossRef]
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.; Yu, W.H.; Lakshmanan, A.; Wade, W.G. The human oral microbiome. J. Bacteriol. 2010, 192, 5002–5017. [Google Scholar] [CrossRef] [PubMed]
- Selwitz, R.H.; Ismail, A.I.; Pitts, N.B. Dental caries. Lancet 2007, 369, 51–59. [Google Scholar] [CrossRef]
- Kassebaum, N.J.; Bernabe, E.; Dahiya, M.; Bhandari, B.; Murray, C.J.; Marcenes, W. Global burden of untreated caries: A systematic review and metaregression. J. Dent. Res. 2015, 94, 650–658. [Google Scholar] [CrossRef]
- Demmer, R.T.; Papapanou, P.N. Epidemiologic patterns of chronic and aggressive periodontitis. Periodontol. 2000 2010, 53, 28–44. [Google Scholar] [CrossRef]
- Holtfreter, B.; Albandar, J.M.; Dietrich, T.; Dye, B.A.; Eaton, K.A.; Eke, P.I.; Papapanou, P.N.; Kocher, T.; Joint EU/USA Periodontal Epidemiology Working Group. Standards for reporting chronic periodontitis prevalence and severity in epidemiologic studies: Proposed standards from the Joint EU/USA Periodontal Epidemiology Working Group. J. Clin. Periodontol. 2015, 42, 407–412. [Google Scholar] [CrossRef]
- Papapanou, P.N.; Sanz, M.; Buduneli, N.; Dietrich, T.; Feres, M.; Fine, D.H.; Flemmig, T.F.; Garcia, R.; Giannobile, W.V.; Graziani, F.; et al. Periodontitis: Consensus report of workgroup 2 of the 2017 World Workshop on the Classification of Periodontal and Peri-Implant Diseases and Conditions. J. Clin. Periodontol. 2018, 45 (Suppl. 20), S162–S170. [Google Scholar] [CrossRef]
- Berglundh, T.; Armitage, G.; Araujo, M.G.; Avila-Ortiz, G.; Blanco, J.; Camargo, P.M.; Chen, S.; Cochran, D.; Derks, J.; Figuero, E.; et al. Peri-implant diseases and conditions: Consensus report of workgroup 4 of the 2017 World Workshop on the Classification of Periodontal and Peri-Implant Diseases and Conditions. J. Clin. Periodontol. 2018, 45 (Suppl. 20), S286–S291. [Google Scholar] [CrossRef]
- Derks, J.; Tomasi, C. Peri-implant health and disease. A systematic review of current epidemiology. J. Clin. Periodontol. 2015, 42 (Suppl. 16), S158–S171. [Google Scholar] [CrossRef]
- Filoche, S.K.; Soma, K.J.; Sissons, C.H. Caries-related plaque microcosm biofilms developed in microplates. Oral Microbiol. Immunol. 2007, 22, 73–79. [Google Scholar] [CrossRef]
- Ccahuana-Vasquez, R.A.; Cury, J.A. S. mutans biofilm model to evaluate antimicrobial substances and enamel demineralization. Braz. Oral Res. 2010, 24, 135–141. [Google Scholar] [CrossRef] [PubMed]
- van de Sande, F.H.; Azevedo, M.S.; Lund, R.G.; Huysmans, M.C.; Cenci, M.S. An in vitro biofilm model for enamel demineralization and antimicrobial dose-response studies. Biofouling 2011, 27, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Arthur, R.A.; Waeiss, R.A.; Hara, A.T.; Lippert, F.; Eckert, G.J.; Zero, D.T. A defined-multispecies microbial model for studying enamel caries development. Caries Res. 2013, 47, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Cavalcanti, Y.W.; Bertolini, M.M.; da Silva, W.J.; Del-Bel-Cury, A.A.; Tenuta, L.M.; Cury, J.A. A three-species biofilm model for the evaluation of enamel and dentin demineralization. Biofouling 2014, 30, 579–588. [Google Scholar] [CrossRef]
- Totiam, P.; Gonzalez-Cabezas, C.; Fontana, M.R.; Zero, D.T. A new in vitro model to study the relationship of gap size and secondary caries. Caries Res. 2007, 41, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Sissons, C.H.; Cutress, T.W.; Hoffman, M.P.; Wakefield, J.S. A multi-station dental plaque microcosm (artificial mouth) for the study of plaque growth, metabolism, pH, and mineralization. J. Dent. Res. 1991, 70, 1409–1416. [Google Scholar] [CrossRef] [PubMed]
- Peters, A.C.; Wimpenny, J.W. A constant-depth laboratory model film fermentor. Biotechnol. Bioeng. 1988, 32, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, D.J.; Marsh, P.D. Effect of sugar alcohols on the composition and metabolism of a mixed culture of oral bacteria grown in a chemostat. Caries Res. 1994, 28, 251–256. [Google Scholar] [CrossRef]
- Rudney, J.D.; Chen, R.; Lenton, P.; Li, J.; Li, Y.; Jones, R.S.; Reilly, C.; Fok, A.S.; Aparicio, C. A reproducible oral microcosm biofilm model for testing dental materials. J. Appl. Microbiol. 2012, 113, 1540–1553. [Google Scholar] [CrossRef] [PubMed]
- Seemann, R.; Bizhang, M.; Kluck, I.; Loth, J.; Roulet, J.F. A novel in vitro microbial-based model for studying caries formation--development and initial testing. Caries Res. 2005, 39, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Seemann, R.; Kluck, I.; Kage, A. An in vitro microbial-based model for studying caries-preventive agents. Acta Odontol. Scand. 2006, 64, 27–30. [Google Scholar] [CrossRef]
- Lynch, R.J.; ten Cate, J.M. Effect of calcium glycerophosphate on demineralization in an in vitro biofilm model. Caries Res. 2006, 40, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Mei, M.L.; Chu, C.H.; Lo, E.C.; Samaranayake, L.P. Preventing root caries development under oral biofilm challenge in an artificial mouth. Med. Oral Patol. Oral Cir. Bucal 2013, 18, e557–e563. [Google Scholar] [CrossRef]
- Mei, M.L.; Li, Q.L.; Chu, C.H.; Lo, E.C.; Samaranayake, L.P. Antibacterial effects of silver diamine fluoride on multi-species cariogenic biofilm on caries. Ann. Clin. Microbiol. Antimicrob. 2013, 12, 4. [Google Scholar] [CrossRef]
- Kramer, N.; Schmidt, M.; Lucker, S.; Domann, E.; Frankenberger, R. Glass ionomer cement inhibits secondary caries in an in vitro biofilm model. Clin. Oral Investig. 2018, 22, 1019–1031. [Google Scholar] [CrossRef]
- Schwendicke, F.; Dorfer, C.; Kneist, S.; Meyer-Lueckel, H.; Paris, S. Cariogenic effects of probiotic Lactobacillus rhamnosus GG in a dental biofilm model. Caries Res. 2014, 48, 186–192. [Google Scholar] [CrossRef]
- Wecke, J.; Kersten, T.; Madela, K.; Moter, A.; Gobel, U.B.; Friedmann, A.; Bernimoulin, J. A novel technique for monitoring the development of bacterial biofilms in human periodontal pockets. FEMS Microbiol. Lett. 2000, 191, 95–101. [Google Scholar] [CrossRef]
- Takeuchi, H.; Yamanaka, Y.; Yamamoto, K. Morphological analysis of subgingival biofilm formation on synthetic carbonate apatite inserted into human periodontal pockets. Aust. Dent. J. 2004, 49, 72–77. [Google Scholar] [CrossRef]
- Periasamy, S.; Kolenbrander, P.E. Mutualistic biofilm communities develop with Porphyromonas gingivalis and initial, early, and late colonizers of enamel. J. Bacteriol. 2009, 191, 6804–6811. [Google Scholar] [CrossRef] [PubMed]
- Hope, C.K.; Wilson, M. Biofilm structure and cell vitality in a laboratory model of subgingival plaque. J. Microbiol. Methods 2006, 66, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Kuramitsu, H.K.; Chen, W.; Ikegami, A. Biofilm formation by the periodontopathic bacteria Treponema denticola and Porphyromonas gingivalis. J. Periodontol. 2005, 76, 2047–2051. [Google Scholar] [CrossRef]
- Walker, C.; Sedlacek, M.J. An in vitro biofilm model of subgingival plaque. Oral Microbiol. Immunol. 2007, 22, 152–161. [Google Scholar] [CrossRef]
- Tamura, A.; Ara, T.; Imamura, Y.; Fujii, T.; Wang, P.L. The effects of antibiotics on in vitro biofilm model of periodontal disease. Eur. J. Med. Res. 2008, 13, 439–445. [Google Scholar]
- Fernandez, Y.M.M.; Exterkate, R.A.M.; Buijs, M.J.; Beertsen, W.; van der Weijden, G.A.; Zaura, E.; Crielaard, W. A reproducible microcosm biofilm model of subgingival microbial communities. J. Periodontal Res. 2017, 52, 1021–1031. [Google Scholar] [CrossRef] [PubMed]
- Song, W.S.; Lee, J.K.; Park, S.H.; Um, H.S.; Lee, S.Y.; Chang, B.S. Comparison of periodontitis-associated oral biofilm formation under dynamic and static conditions. J. Periodontal Implant. Sci. 2017, 47, 219–230. [Google Scholar] [CrossRef]
- Ceri, H.; Olson, M.E.; Stremick, C.; Read, R.R.; Morck, D.; Buret, A. The Calgary Biofilm Device: New technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J. Clin. Microbiol. 1999, 37, 1771–1776. [Google Scholar] [CrossRef]
- Dibdin, G.; Wimpenny, J. Steady-state biofilm: Practical and theoretical models. Methods Enzymol. 1999, 310, 296–322. [Google Scholar] [CrossRef] [PubMed]
- Guggenheim, B.; Giertsen, E.; Schupbach, P.; Shapiro, S. Validation of an in vitro biofilm model of supragingival plaque. J. Dent. Res. 2001, 80, 363–370. [Google Scholar] [CrossRef]
- Zijnge, V.; van Leeuwen, M.B.; Degener, J.E.; Abbas, F.; Thurnheer, T.; Gmur, R.; Harmsen, H.J. Oral biofilm architecture on natural teeth. PLoS ONE 2010, 5, e9321. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.C.; Llama-Palacios, A.; Blanc, V.; Leon, R.; Herrera, D.; Sanz, M. Structure, viability and bacterial kinetics of an in vitro biofilm model using six bacteria from the subgingival microbiota. J. Periodontal Res. 2011, 46, 252–260. [Google Scholar] [CrossRef]
- Auschill, T.M.; Arweiler, N.B.; Netuschil, L.; Brecx, M.; Reich, E.; Sculean, A. Spatial distribution of vital and dead microorganisms in dental biofilms. Arch. Oral Biol. 2001, 46, 471–476. [Google Scholar] [CrossRef]
- Datey, A.; Thaha, C.S.A.; Patil, S.R.; Gopalan, J.; Chakravortty, D. Shockwave Therapy Efficiently Cures Multispecies Chronic Periodontitis in a Humanized Rat Model. Front. Bioeng. Biotechnol. 2019, 7, 382. [Google Scholar] [CrossRef]
- Dabija-Wolter, G.; Al-Zubaydi, S.S.; Mohammed, M.M.A.; Bakken, V.; Bolstad, A.I. The effect of metronidazole plus amoxicillin or metronidazole plus penicillin V on periodontal pathogens in an in vitro biofilm model. Clin. Exp. Dent. Res. 2018, 4, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Blanc, V.; Isabal, S.; Sanchez, M.C.; Llama-Palacios, A.; Herrera, D.; Sanz, M.; Leon, R. Characterization and application of a flow system for in vitro multispecies oral biofilm formation. J. Periodontal Res. 2014, 49, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Dabholkar, C.S.; Shah, M.; Kathariya, R.; Bajaj, M.; Doshi, Y. Comparative Evaluation of Antimicrobial Activity of Pomegranate-Containing Mouthwash Against Oral-Biofilm Forming Organisms: An Invitro Microbial Study. J. Clin. Diagn Res. 2016, 10, ZC65–ZC69. [Google Scholar] [CrossRef]
- Muller, H.D.; Eick, S.; Moritz, A.; Lussi, A.; Gruber, R. Cytotoxicity and Antimicrobial Activity of Oral Rinses In Vitro. Biomed. Res. Int. 2017, 2017, 4019723. [Google Scholar] [CrossRef]
- Rabin, N.; Zheng, Y.; Opoku-Temeng, C.; Du, Y.; Bonsu, E.; Sintim, H.O. Biofilm formation mechanisms and targets for developing antibiofilm agents. Future Med. Chem. 2015, 7, 493–512. [Google Scholar] [CrossRef]
- Macia, M.D.; Rojo-Molinero, E.; Oliver, A. Antimicrobial susceptibility testing in biofilm-growing bacteria. Clin. Microbiol. Infect. 2014, 20, 981–990. [Google Scholar] [CrossRef]
- Jiao, Y.; Tay, F.R.; Niu, L.N.; Chen, J.H. Advancing antimicrobial strategies for managing oral biofilm infections. Int. J. Oral Sci. 2019, 11, 28. [Google Scholar] [CrossRef] [PubMed]
- Cawthorn, D.M.; Witthuhn, R.C. Selective PCR detection of viable Enterobacter sakazakii cells utilizing propidium monoazide or ethidium bromide monoazide. J. Appl. Microbiol. 2008, 105, 1178–1185. [Google Scholar] [CrossRef]
- Nocker, A.; Cheung, C.Y.; Camper, A.K. Comparison of propidium monoazide with ethidium monoazide for differentiation of live vs. dead bacteria by selective removal of DNA from dead cells. J. Microbiol. Methods 2006, 67, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.C.; Marin, M.J.; Figuero, E.; Llama-Palacios, A.; Leon, R.; Blanc, V.; Herrera, D.; Sanz, M. Quantitative real-time PCR combined with propidium monoazide for the selective quantification of viable periodontal pathogens in an in vitro subgingival biofilm model. J. Periodontal Res. 2014, 49, 20–28. [Google Scholar] [CrossRef]
- Sanchez, M.C.; Marin, M.J.; Figuero, E.; Llama-Palacios, A.; Herrera, D.; Sanz, M. Analysis of viable vs. dead Aggregatibacter actinomycetemcomitans and Porphyromonas gingivalis using selective quantitative real-time PCR with propidium monoazide. J. Periodontal Res. 2013, 48, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.C.; Fernandez, E.; Llama-Palacios, A.; Figuero, E.; Herrera, D.; Sanz, M. Response to antiseptic agents of periodontal pathogens in in vitro biofilms on titanium and zirconium surfaces. Dent. Mater. 2017, 33, 446–453. [Google Scholar] [CrossRef]
- Serrano, J.; Escribano, M.; Roldan, S.; Martin, C.; Herrera, D. Efficacy of adjunctive anti-plaque chemical agents in managing gingivitis: A systematic review and meta-analysis. J. Clin. Periodontol. 2015, 42 (Suppl. 16), S106–S138. [Google Scholar] [CrossRef]
- Palaska, I.; Papathanasiou, E.; Theoharides, T.C. Use of polyphenols in periodontal inflammation. Eur. J. Pharmacol. 2013, 720, 77–83. [Google Scholar] [CrossRef]
- Cozens, D.; Read, R.C. Anti-adhesion methods as novel therapeutics for bacterial infections. Expert Rev. Anti Infect. Ther. 2012, 10, 1457–1468. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, S.; Moran, A.; Martinez-Blanco, H.; Ferrero, M.A.; Rodriguez-Aparicio, L.B. The Usefulness of Non-Toxic Plant Metabolites in the Control of Bacterial Proliferation. Probiotics Antimicrob. Proteins 2017, 9, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Thimothe, J.; Bonsi, I.A.; Padilla-Zakour, O.I.; Koo, H. Chemical characterization of red wine grape (Vitis vinifera and Vitis interspecific hybrids) and pomace phenolic extracts and their biological activity against Streptococcus mutans. J. Agric. Food Chem. 2007, 55, 10200–10207. [Google Scholar] [CrossRef]
- Furiga, A.; Roques, C.; Badet, C. Preventive effects of an original combination of grape seed polyphenols with amine fluoride on dental biofilm formation and oxidative damage by oral bacteria. J. Appl. Microbiol. 2014, 116, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, M.; Millhouse, E.; Culshaw, S.; Edwards, C.A.; Ramage, G.; Combet, E. Selected dietary (poly)phenols inhibit periodontal pathogen growth and biofilm formation. Food Funct. 2015, 6, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.C.; Ribeiro-Vidal, H.; Esteban-Fernandez, A.; Bartolome, B.; Figuero, E.; Moreno-Arribas, M.V.; Sanz, M.; Herrera, D. Antimicrobial activity of red wine and oenological extracts against periodontal pathogens in a validated oral biofilm model. BMC Complement. Altern. Med. 2019, 19, 145. [Google Scholar] [CrossRef] [PubMed]
- Feghali, K.; Feldman, M.; La, V.D.; Santos, J.; Grenier, D. Cranberry proanthocyanidins: Natural weapons against periodontal diseases. J. Agric. Food Chem. 2012, 60, 5728–5735. [Google Scholar] [CrossRef]
- Bodet, C.; Grenier, D.; Chandad, F.; Ofek, I.; Steinberg, D.; Weiss, E.I. Potential oral health benefits of cranberry. Crit. Rev. Food Sci. Nutr. 2008, 48, 672–680. [Google Scholar] [CrossRef]
- Philip, N.; Walsh, L.J. Cranberry Polyphenols: Natural Weapons against Dental Caries. Dent. J. 2019, 7, 20. [Google Scholar] [CrossRef]
- Sanchez, M.C.; Ribeiro-Vidal, H.; Bartolome, B.; Figuero, E.; Moreno-Arribas, M.V.; Sanz, M.; Herrera, D. New Evidences of Antibacterial Effects of Cranberry Against Periodontal Pathogens. Foods 2020, 9, 246. [Google Scholar] [CrossRef]
- Desbois, A.P. Potential applications of antimicrobial fatty acids in medicine, agriculture and other industries. Recent Pat. Antiinfect. Drug Discov. 2012, 7, 111–122. [Google Scholar] [CrossRef]
- Desbois, A.P.; Lawlor, K.C. Antibacterial activity of long-chain polyunsaturated fatty acids against Propionibacterium acnes and Staphylococcus aureus. Mar. Drugs 2013, 11, 4544–4557. [Google Scholar] [CrossRef] [PubMed]
- Mil-Homens, D.; Bernardes, N.; Fialho, A.M. The antibacterial properties of docosahexaenoic omega-3 fatty acid against the cystic fibrosis multiresistant pathogen Burkholderia cenocepacia. FEMS Microbiol. Lett. 2012, 328, 61–69. [Google Scholar] [CrossRef]
- Serhan, C.N. Resolution phase of inflammation: Novel endogenous anti-inflammatory and proresolving lipid mediators and pathways. Annu. Rev. Immunol. 2007, 25, 101–137. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro-Vidal, H.; Sanchez, M.C.; Alonso-Espanol, A.; Figuero, E.; Ciudad, M.J.; Collado, L.; Herrera, D.; Sanz, M. Antimicrobial Activity of EPA and DHA against Oral Pathogenic Bacteria Using an In Vitro Multi-Species Subgingival Biofilm Model. Nutrients 2020, 12, 2812. [Google Scholar] [CrossRef]
- Muehler, D.; Rupp, C.M.; Keceli, S.; Brochhausen, C.; Siegmund, H.; Maisch, T.; Hiller, K.A.; Buchalla, W.; Cieplik, F. Insights Into Mechanisms of Antimicrobial Photodynamic Action Toward Biofilms Using Phenalen-1-One Derivatives as Photosensitizers. Front. Microbiol. 2020, 11, 589364. [Google Scholar] [CrossRef]
- Qi, M.; Chi, M.; Sun, X.; Xie, X.; Weir, M.D.; Oates, T.W.; Zhou, Y.; Wang, L.; Bai, Y.; Xu, H.H. Novel nanomaterial-based antibacterial photodynamic therapies to combat oral bacterial biofilms and infectious diseases. Int. J. Nanomed. 2019, 14, 6937–6956. [Google Scholar] [CrossRef]
- Esteban-Fernandez, A.; Zorraquin-Pena, I.; Ferrer, M.D.; Mira, A.; Bartolome, B.; Gonzalez de Llano, D.; Moreno-Arribas, M.V. Inhibition of Oral Pathogens Adhesion to Human Gingival Fibroblasts by Wine Polyphenols Alone and in Combination with an Oral Probiotic. J. Agric. Food Chem. 2018, 66, 2071–2082. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Tiwari, M.; Donelli, G.; Tiwari, V. Strategies for combating bacterial biofilms: A focus on anti-biofilm agents and their mechanisms of action. Virulence 2018, 9, 522–554. [Google Scholar] [CrossRef] [PubMed]
- Jenssen, H.; Hamill, P.; Hancock, R.E. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [PubMed]
- Matto, J.; Malinen, E.; Suihko, M.L.; Alander, M.; Palva, A.; Saarela, M. Genetic heterogeneity and functional properties of intestinal bifidobacteria. J. Appl. Microbiol. 2004, 97, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Oren, Z.; Shai, Y. Mode of action of linear amphipathic alpha-helical antimicrobial peptides. Biopolymers 1998, 47, 451–463. [Google Scholar] [CrossRef]
- Tenorio, E.L.; Klein, B.A.; Cheung, W.S.; Hu, L.T. Identification of interspecies interactions affecting Porphyromonas gingivalis virulence phenotypes. J. Oral Microbiol. 2011, 3, 8396. [Google Scholar] [CrossRef] [PubMed]
- Abou Neel, E.A.; Bozec, L.; Perez, R.A.; Kim, H.W.; Knowles, J.C. Nanotechnology in dentistry: Prevention, diagnosis, and therapy. Int. J. Nanomed. 2015, 10, 6371–6394. [Google Scholar] [CrossRef] [PubMed]
- Jain, N.; Jain, G.K.; Javed, S.; Iqbal, Z.; Talegaonkar, S.; Ahmad, F.J.; Khar, R.K. Recent approaches for the treatment of periodontitis. Drug Discov. Today 2008, 13, 932–943. [Google Scholar] [CrossRef] [PubMed]
- Osorio, R.; Alfonso-Rodriguez, C.A.; Medina-Castillo, A.L.; Alaminos, M.; Toledano, M. Bioactive Polymeric Nanoparticles for Periodontal Therapy. PLoS ONE 2016, 11, e0166217. [Google Scholar] [CrossRef]
- Osorio, R.; Osorio, E.; Medina-Castillo, A.L.; Toledano, M. Polymer nanocarriers for dentin adhesion. J. Dent. Res. 2014, 93, 1258–1263. [Google Scholar] [CrossRef]
- Munchow, E.A.; Pankajakshan, D.; Albuquerque, M.T.; Kamocki, K.; Piva, E.; Gregory, R.L.; Bottino, M.C. Synthesis and characterization of CaO-loaded electrospun matrices for bone tissue engineering. Clin. Oral Investig. 2016, 20, 1921–1933. [Google Scholar] [CrossRef] [PubMed]
- Baalousha, M.; Nur, Y.; Romer, I.; Tejamaya, M.; Lead, J.R. Effect of monovalent and divalent cations, anions and fulvic acid on aggregation of citrate-coated silver nanoparticles. Sci. Total Environ. 2013, 454–455, 119–131. [Google Scholar] [CrossRef]
- Kim, T.S.; Burklin, T.; Schacher, B.; Ratka-Kruger, P.; Schaecken, M.T.; Renggli, H.H.; Fiehn, W.; Eickholz, P. Pharmacokinetic profile of a locally administered doxycycline gel in crevicular fluid, blood, and saliva. J. Periodontol. 2002, 73, 1285–1291. [Google Scholar] [CrossRef]
- Demirel, K.; Baer, P.N.; McNamara, T.F. Topical application of doxycycline on periodontally involved root surfaces in vitro: Comparative analysis of substantivity on cementum and dentin. J. Periodontol. 1991, 62, 312–316. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.C.; Toledano-Osorio, M.; Bueno, J.; Figuero, E.; Toledano, M.; Medina-Castillo, A.L.; Osorio, R.; Herrera, D.; Sanz, M. Antibacterial effects of polymeric PolymP-n Active nanoparticles. An in vitro biofilm study. Dent. Mater. 2019, 35, 156–168. [Google Scholar] [CrossRef] [PubMed]
- Bueno, J.; Sanchez, M.C.; Toledano-Osorio, M.; Figuero, E.; Toledano, M.; Medina-Castillo, A.L.; Osorio, R.; Herrera, D.; Sanz, M. Antimicrobial effect of nanostructured membranes for guided tissue regeneration: An in vitro study. Dent. Mater. 2020, 36, 1566–1577. [Google Scholar] [CrossRef] [PubMed]
- Llama-Palacios, A.; Sanchez, M.C.; Diaz, L.A.; Cabal, B.; Suarez, M.; Moya, J.S.; Torrecillas, R.; Figuero, E.; Sanz, M.; Herrera, D. In vitro biofilm formation on different ceramic biomaterial surfaces: Coating with two bactericidal glasses. Dent. Mater. 2019, 35, 883–892. [Google Scholar] [CrossRef]
- Lo, A.W.; Seers, C.A.; Boyce, J.D.; Dashper, S.G.; Slakeski, N.; Lissel, J.P.; Reynolds, E.C. Comparative transcriptomic analysis of Porphyromonas gingivalis biofilm and planktonic cells. BMC Microbiol. 2009, 9, 18. [Google Scholar] [CrossRef]
- Liu, D.; Xu, J.; Wang, Y.; Chen, Y.; Shen, X.; Niu, H.; Guo, T.; Ying, H. Comparative transcriptomic analysis of Clostridium acetobutylicum biofilm and planktonic cells. J. Biotechnol. 2016, 218, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Whiteley, M.; Bangera, M.G.; Bumgarner, R.E.; Parsek, M.R.; Teitzel, G.M.; Lory, S.; Greenberg, E.P. Gene expression in Pseudomonas aeruginosa biofilms. Nature 2001, 413, 860–864. [Google Scholar] [CrossRef]
- Waite, R.D.; Papakonstantinopoulou, A.; Littler, E.; Curtis, M.A. Transcriptome analysis of Pseudomonas aeruginosa growth: Comparison of gene expression in planktonic cultures and developing and mature biofilms. J. Bacteriol. 2005, 187, 6571–6576. [Google Scholar] [CrossRef] [PubMed]
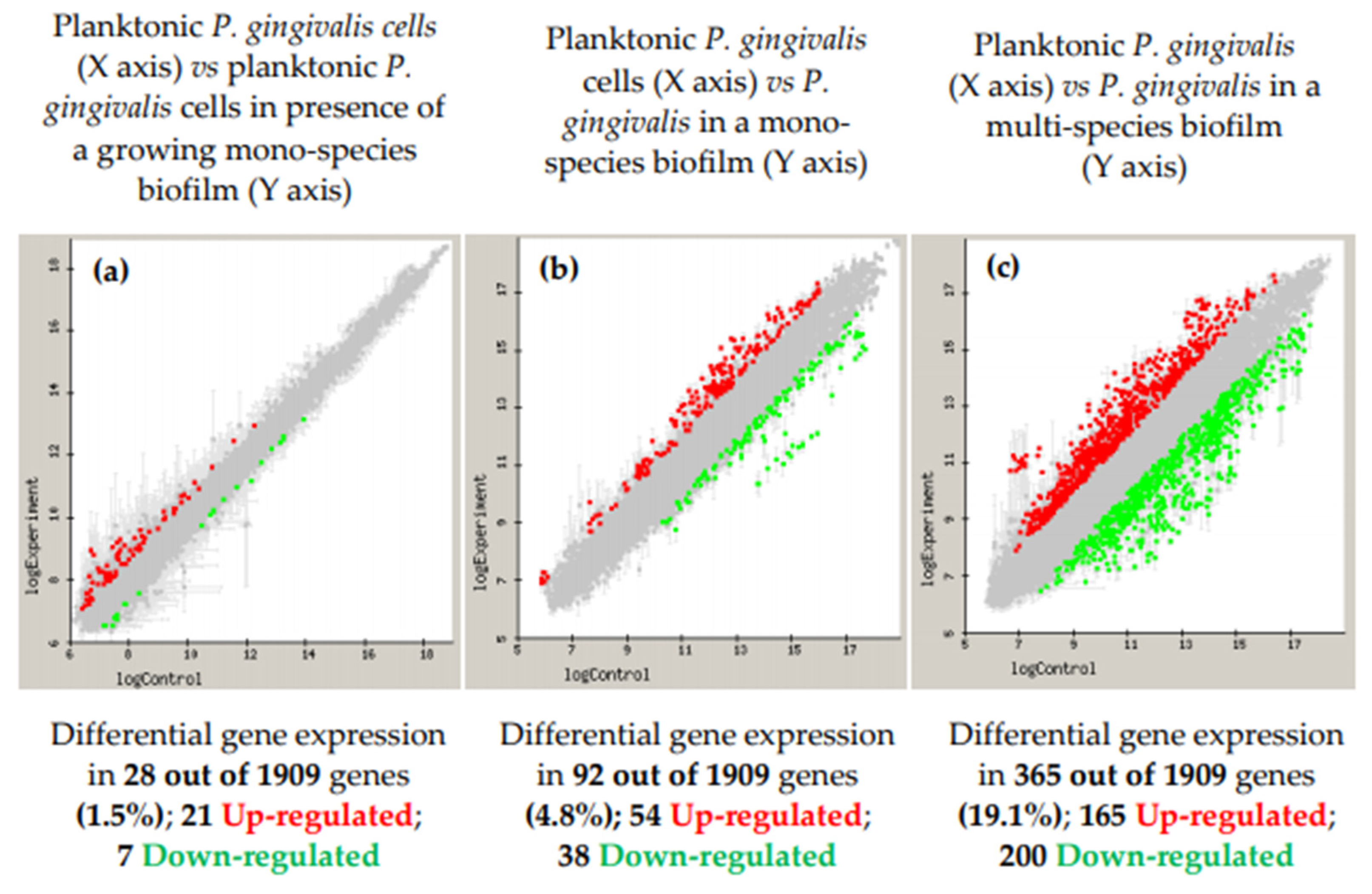
- Sanchez, M.C.; Romero-Lastra, P.; Ribeiro-Vidal, H.; Llama-Palacios, A.; Figuero, E.; Herrera, D.; Sanz, M. Comparative gene expression analysis of planktonic Porphyromonas gingivalis ATCC 33277 in the presence of a growing biofilm versus planktonic cells. BMC Microbiol. 2019, 19, 58. [Google Scholar] [CrossRef]
- Romero-Lastra, P.; Sanchez, M.C.; Ribeiro-Vidal, H.; Llama-Palacios, A.; Figuero, E.; Herrera, D.; Sanz, M. Comparative gene expression analysis of Porphyromonas gingivalis ATCC 33277 in planktonic and biofilms states. PLoS ONE 2017, 12, e0174669. [Google Scholar] [CrossRef] [PubMed]
- Romero-Lastra, P.; Sanchez, M.C.; Llama-Palacios, A.; Figuero, E.; Herrera, D.; Sanz, M. Gene expression of Porphyromonas gingivalis ATCC 33277 when growing in an in vitro multispecies biofilm. PLoS ONE 2019, 14, e0221234. [Google Scholar] [CrossRef]
- Llama-Palacios, A.; Potupa, O.; Sanchez, M.C.; Figuero, E.; Herrera, D.; Sanz, M. Aggregatibacter actinomycetemcomitans Growth in Biofilm versus Planktonic State: Differential Expression of Proteins. J. Proteome Res. 2017, 16, 3158–3167. [Google Scholar] [CrossRef]
- Llama-Palacios, A.; Potupa, O.; Sanchez, M.C.; Figuero, E.; Herrera, D.; Sanz, M. Proteomic analysis of Fusobacterium nucleatum growth in biofilm versus planktonic state. Mol. Oral Microbiol. 2020, 35, 168–180. [Google Scholar] [CrossRef]
- Jung, R.E.; Zembic, A.; Pjetursson, B.E.; Zwahlen, M.; Thoma, D.S. Systematic review of the survival rate and the incidence of biological, technical, and aesthetic complications of single crowns on implants reported in longitudinal studies with a mean follow-up of 5 years. Clin. Oral Implant. Res. 2012, 23 (Suppl. 6), 2–21. [Google Scholar] [CrossRef]
- Pjetursson, B.E.; Thoma, D.; Jung, R.; Zwahlen, M.; Zembic, A. A systematic review of the survival and complication rates of implant-supported fixed dental prostheses (FDPs) after a mean observation period of at least 5 years. Clin. Oral Implant. Res. 2012, 23 (Suppl. 6), 22–38. [Google Scholar] [CrossRef]
- Rakic, M.; Lekovic, V.; Nikolic-Jakoba, N.; Vojvodic, D.; Petkovic-Curcin, A.; Sanz, M. Bone loss biomarkers associated with peri-implantitis. A cross-sectional study. Clin. Oral Implant. Res. 2013, 24, 1110–1116. [Google Scholar] [CrossRef]
- Beikler, T.; Flemmig, T.F. Oral biofilm-associated diseases: Trends and implications for quality of life, systemic health and expenditures. Periodontol. 2000 2011, 55, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Leonhardt, A.; Renvert, S.; Dahlen, G. Microbial findings at failing implants. Clin. Oral Implant. Res. 1999, 10, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Di Giulio, M.; Traini, T.; Sinjari, B.; Nostro, A.; Caputi, S.; Cellini, L. Porphyromonas gingivalis biofilm formation in different titanium surfaces, an in vitro study. Clin. Oral Implant. Res. 2016, 27, 918–925. [Google Scholar] [CrossRef] [PubMed]
- Pita, P.P.; Rodrigues, J.A.; Ota-Tsuzuki, C.; Miato, T.F.; Zenobio, E.G.; Giro, G.; Figueiredo, L.C.; Goncalves, C.; Gehrke, S.A.; Cassoni, A.; et al. Oral Streptococci Biofilm Formation on Different Implant Surface Topographies. Biomed. Res. Int. 2015, 2015, 159625. [Google Scholar] [CrossRef]
- Aguayo, S.; Donos, N.; Spratt, D.; Bozec, L. Nanoadhesion of Staphylococcus aureus onto Titanium Implant Surfaces. J. Dent. Res. 2015, 94, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- de Avila, E.D.; Lima, B.P.; Sekiya, T.; Torii, Y.; Ogawa, T.; Shi, W.; Lux, R. Effect of UV-photofunctionalization on oral bacterial attachment and biofilm formation to titanium implant material. Biomaterials 2015, 67, 84–92. [Google Scholar] [CrossRef]
- Papavasileiou, D.; Behr, M.; Gosau, M.; Gerlach, T.; Buergers, R. Peri-implant Biofilm Formation on Luting Agents Used for Cementing Implant-Supported Fixed Restorations: A Preliminary In Vivo Study. Int. J. Prosthodont. 2015, 28, 371–373. [Google Scholar] [CrossRef]
- Ready, D.; Theodoridis, G.; Green, I.; Ciric, L.; Pratten, J.; Tay, W.; McDonald, A. In vitro evaluation of the antibiofilm properties of chlorhexidine and delmopinol on dental implant surfaces. Int. J. Antimicrob. Agents 2015, 45, 662–666. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.E.; Auschill, T.M.; Heumann, C.; Frankenberger, R.; Eick, S.; Sculean, A.; Arweiler, N.B. Influence of different instrumentation modalities on the surface characteristics and biofilm formation on dental implant neck, in vitro. Clin. Oral Implant. Res. 2017, 28, 483–490. [Google Scholar] [CrossRef]
- Sridhar, S.; Abidi, Z.; Wilson, T.G., Jr.; Valderrama, P.; Wadhwani, C.; Palmer, K.; Rodrigues, D.C. In Vitro Evaluation of the Effects of Multiple Oral Factors on Dental Implants Surfaces. J. Oral Implantol. 2016, 42, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Marotti, J.; Tortamano, P.; Cai, S.; Ribeiro, M.S.; Franco, J.E.; de Campos, T.T. Decontamination of dental implant surfaces by means of photodynamic therapy. Lasers Med. Sci. 2013, 28, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Paster, B.J.; Boches, S.K.; Galvin, J.L.; Ericson, R.E.; Lau, C.N.; Levanos, V.A.; Sahasrabudhe, A.; Dewhirst, F.E. Bacterial diversity in human subgingival plaque. J. Bacteriol. 2001, 183, 3770–3783. [Google Scholar] [CrossRef] [PubMed]
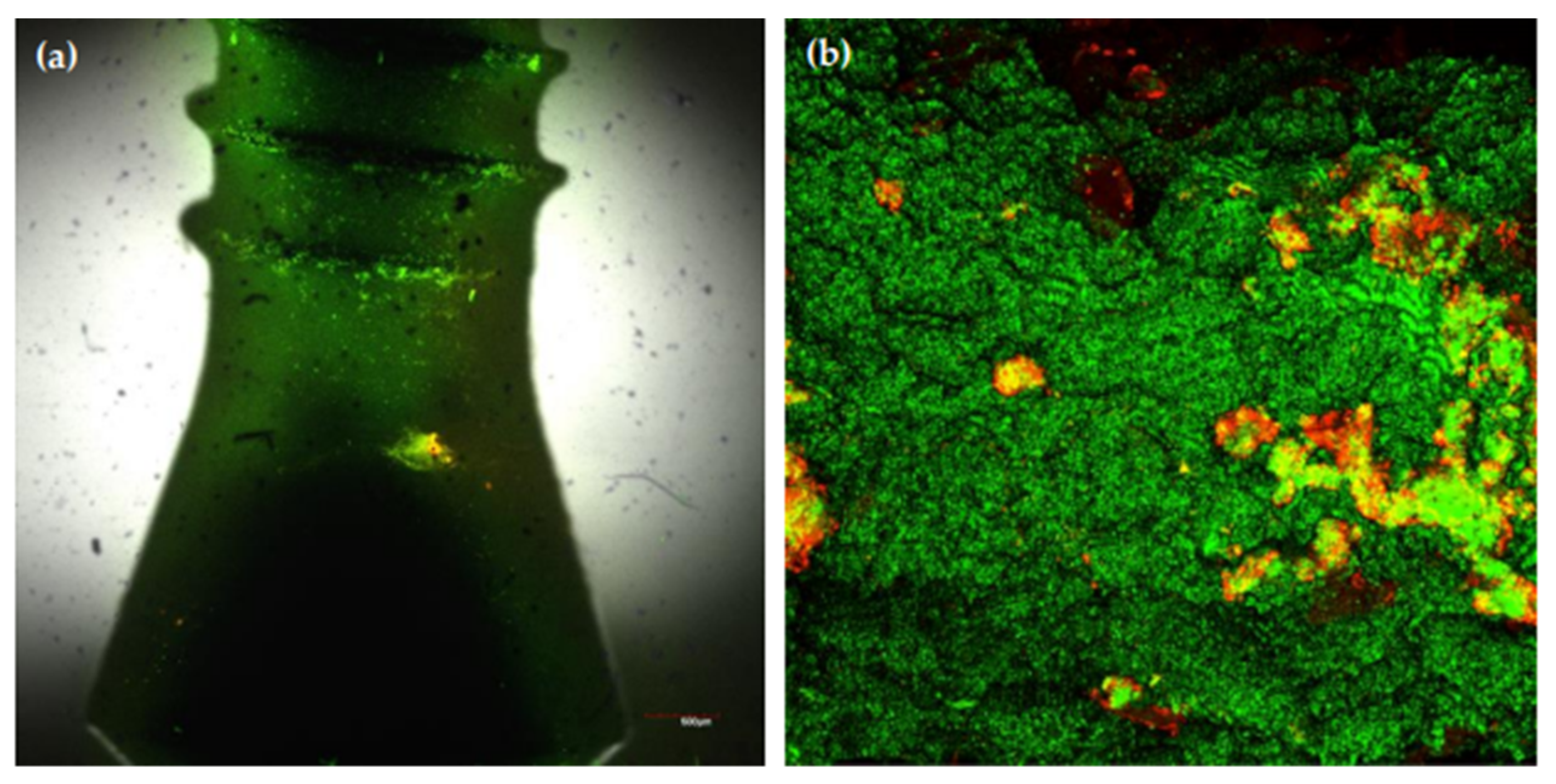
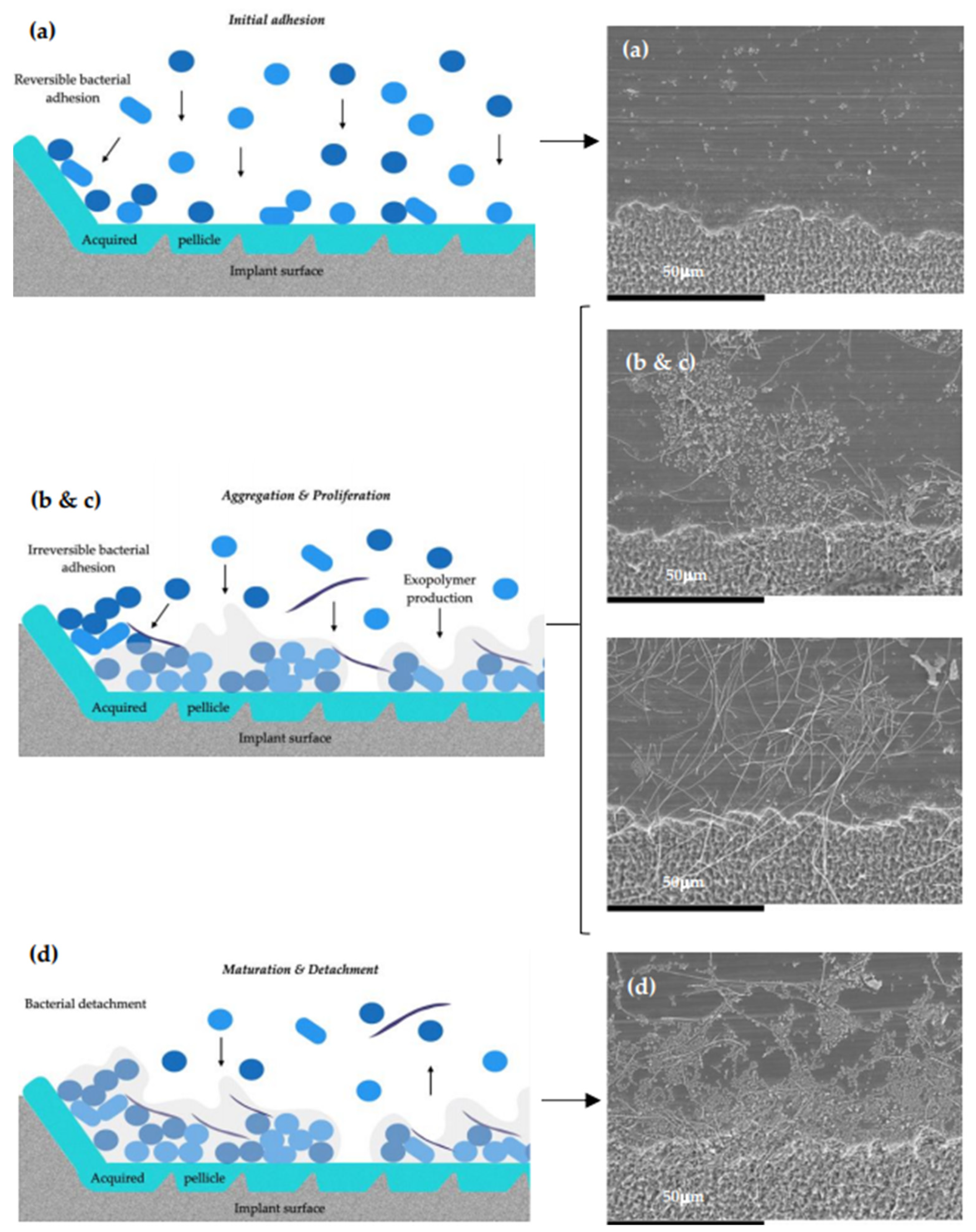
- Sanchez, M.C.; Llama-Palacios, A.; Fernandez, E.; Figuero, E.; Marin, M.J.; Leon, R.; Blanc, V.; Herrera, D.; Sanz, M. An in vitro biofilm model associated to dental implants: Structural and quantitative analysis of in vitro biofilm formation on different dental implant surfaces. Dent. Mater. 2014, 30, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Ferreira Ribeiro, C.; Cogo-Muller, K.; Franco, G.C.; Silva-Concilio, L.R.; Sampaio Campos, M.; de Mello Rode, S.; Claro Neves, A.C. Initial oral biofilm formation on titanium implants with different surface treatments: An in vivo study. Arch. Oral Biol. 2016, 69, 33–39. [Google Scholar] [CrossRef]
- Bermejo, P.; Sanchez, M.C.; Llama-Palacios, A.; Figuero, E.; Herrera, D.; Sanz, M. Topographic characterization of multispecies biofilms growing on dental implant surfaces: An in vitro model. Clin. Oral Implant. Res. 2019, 30, 229–241. [Google Scholar] [CrossRef]
- Bermejo, P.; Sanchez, M.C.; Llama-Palacios, A.; Figuero, E.; Herrera, D.; Sanz Alonso, M. Biofilm formation on dental implants with different surface micro-topography: An in vitro study. Clin. Oral Implant. Res. 2019, 30, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Drescher, K.; Dunkel, J.; Cisneros, L.H.; Ganguly, S.; Goldstein, R.E. Fluid dynamics and noise in bacterial cell-cell and cell-surface scattering. Proc. Natl. Acad. Sci. USA 2011, 108, 10940–10945. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro-Vidal, H.; Sanchez, M.D.C.; Figuero, E.; Herrera, D.; Sanz, M. Biofilm formation on dental implant surfaces—In vitro dynamic model. Clin. Oral Implant. Res. 2019, 30, 8. [Google Scholar] [CrossRef]
- Dawes, C.; Watanabe, S.; Biglow-Lecomte, P.; Dibdin, G.H. Estimation of the velocity of the salivary film at some different locations in the mouth. J. Dent. Res. 1989, 68, 1479–1482. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez, M.C.; Alonso-Español, A.; Ribeiro-Vidal, H.; Alonso, B.; Herrera, D.; Sanz, M. Relevance of Biofilm Models in Periodontal Research: From Static to Dynamic Systems. Microorganisms 2021, 9, 428. https://doi.org/10.3390/microorganisms9020428
Sánchez MC, Alonso-Español A, Ribeiro-Vidal H, Alonso B, Herrera D, Sanz M. Relevance of Biofilm Models in Periodontal Research: From Static to Dynamic Systems. Microorganisms. 2021; 9(2):428. https://doi.org/10.3390/microorganisms9020428
Chicago/Turabian StyleSánchez, María Carmen, Andrea Alonso-Español, Honorato Ribeiro-Vidal, Bettina Alonso, David Herrera, and Mariano Sanz. 2021. "Relevance of Biofilm Models in Periodontal Research: From Static to Dynamic Systems" Microorganisms 9, no. 2: 428. https://doi.org/10.3390/microorganisms9020428
APA StyleSánchez, M. C., Alonso-Español, A., Ribeiro-Vidal, H., Alonso, B., Herrera, D., & Sanz, M. (2021). Relevance of Biofilm Models in Periodontal Research: From Static to Dynamic Systems. Microorganisms, 9(2), 428. https://doi.org/10.3390/microorganisms9020428