Phylogenetic Assignment of the Fungicolous Hypoxylon invadens (Ascomycota, Xylariales) and Investigation of its Secondary Metabolites




Abstract
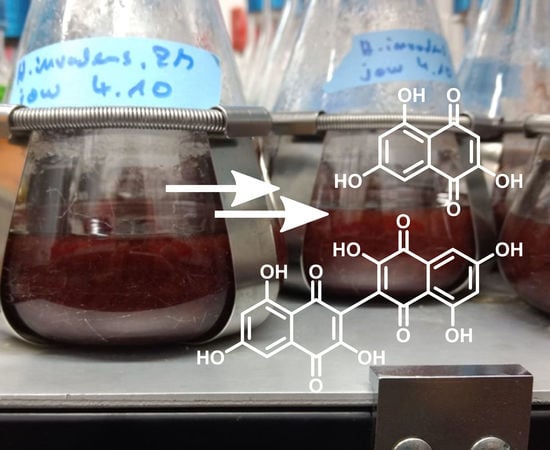
1. Introduction
2. Materials and Methods
2.1. General
2.2. Fungal Material and Molecular Phylogeny
2.3. Cultivation, Extraction, and Isolation
2.4. Antimicrobial Acticity and Cytotoxicity Assay
2.5. Spectral Data
2.5.1. Flaviolin
2.5.2. 3,3′-Biflaviolin
3. Results
3.1. Molecular Phylogeny of H. invadens
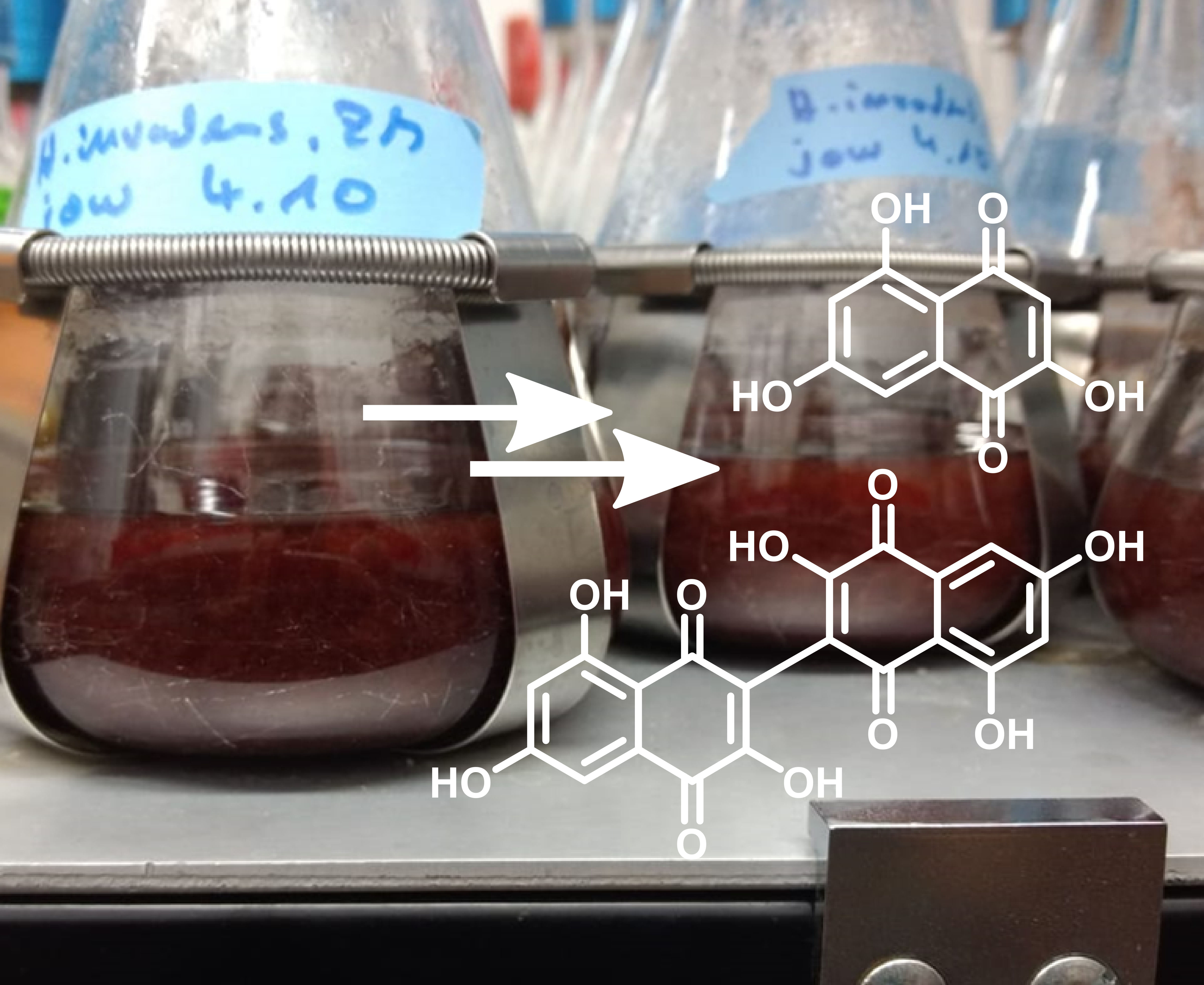
3.2. Structure Elucidation of Flaviolin (1) and 3,3′-Biflaviolin (2) and Analysis of Extracts from other Culture Media
3.3. Antimicrobial and Cytotoxic Activities of Flaviolin (1) and 3,3′-Biflaviolin (2)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hyde, K.D. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Wendt, L.; Sir, E.B.; Kuhnert, E.; Heitkämper, S.; Lambert, C.; Hladki, A.I.; Romero, A.I.; Luangsa-ard, J.J.; Srikitikulchai, P.; Peršoh, D.; et al. Resurrection and emendation of the Hypoxylaceae, recognised from a multigene phylogeny of the Xylariales. Mycol. Prog. 2018, 17, 115–154. [Google Scholar] [CrossRef]
- Lambert, C.; Wendt, L.; Hladki, A.I.; Stadler, M.; Sir, E.B. Hypomontagnella (Hypoxylaceae): a new genus segregated from Hypoxylon by a polyphasic taxonomic approach. Mycol. Prog. 2018, 18, 187–201. [Google Scholar] [CrossRef]
- Hsieh, H.M.; Ju, Y.M.; Rogers, J.D. Molecular phylogeny of Hypoxylon and closely related genera. Mycologia 2005, 97, 844–865. [Google Scholar] [CrossRef]
- Stadler, M.; Lambert, C.; Wibberg, D.; Kalinowski, J.; Cox, R.J.; Kolařík, M.; Kuhnert, E. Intragenomic polymorphisms in the ITS region of high-quality genomes of the Hypoxylaceae (Xylariales, Ascomycota). Mycol. Prog. 2020, 19, 235–245. [Google Scholar] [CrossRef]
- Helaly, S.E.; Thongbai, B.; Stadler, M. Diversity of biologically active secondary metabolites from endophytic and saprotrophic fungi of the ascomycete order Xylariales. Nat. Prod. Rep. 2018, 35, 992–1014. [Google Scholar] [CrossRef]
- Wibberg, D.; Stadler, M.; Lambert, C.; Bunk, B.; Spröer, C.; Rückert, C.; Kalinowski, J.; Cox, R.J.; Kuhnert, E. High quality genome sequences of thirteen Hypoxylaceae (Ascomycota) strengthen the phylogenetic family backbone and enable the discovery of new taxa. Fungal Divers. 2020. [Google Scholar] [CrossRef]
- Fournier, J. Two new species of Hypoxylon (Xylariaceae) from French Central Pyrénées occurring on Fagus sylvatica. Ascomycete.org 2014, 6, 53–60. [Google Scholar] [CrossRef]
- Laessoe, T.; Srikitikulchai, P.; Fournier, J.; Kopcke, B.; Stadler, M. Lepraric acid derivatives as chemotaxonomic markers in Hypoxylon aeruginosum, Chlorostroma subcubisporum and C. cyaninum sp. nov. Fungal Biol. 2010, 114, 481–489. [Google Scholar] [CrossRef]
- Dickschat, J.S.; Wang, T.; Stadler, M. Volatiles from the xylarialean fungus Hypoxylon invadens. Beilstein J. Org. Chem. 2018, 14, 734–746. [Google Scholar] [CrossRef]
- Rinkel, J.; Babczyk, A.; Wang, T.; Stadler, M.; Dickschat, J.S. Volatiles from the hypoxylaceous fungi Hypoxylon griseobrunneum and Hypoxylon macrocarpum. Beilstein J. Org. Chem. 2018, 14, 2974–2990. [Google Scholar] [CrossRef] [PubMed]
- Kuhnert, E.; Heitkamper, S.; Fournier, J.; Surup, F.; Stadler, M. Hypoxyvermelhotins A-C, new pigments from Hypoxylon lechatii sp. nov. Fungal Biol. 2014, 118, 242–252. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Otto, A.; Laub, A.; Wendt, L.; Porzel, A.; Schmidt, J.; Palfner, G.; Becerra, J.; Kruger, D.; Stadler, M.; Wessjohann, L.; et al. Chilenopeptins A and B, peptaibols from the Chilean Sepedonium aff. chalcipori KSH 883. J. Nat. Prod. 2016, 79, 929–938. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Sir, E.B.; Becker, K.; Lambert, C.; Bills, G.F.; Kuhnert, E. Observations on Texas hypoxylons, including two new Hypoxylon species and widespread environmental isolates of the H. croceum complex identified by a polyphasic approach. Mycologia 2019, 111, 832–856. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: new models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Shimodaira, H.; Hasegawa, M. Multiple comparisons of log-likelihoods with applications to phylogenetic inference. Mol. Biol. Evol. 1999, 16, 1114–1116. [Google Scholar] [CrossRef]
- Kuhnert, E.; Sir, E.B.; Lambert, C.; Hyde, K.D.; Hladki, A.I.; Romero, A.I.; Rohde, M.; Stadler, M. Phylogenetic and chemotaxonomic resolution of the genus Annulohypoxylon (Xylariaceae) including four new species. Fungal Divers. 2017, 85, 1–43. [Google Scholar] [CrossRef]
- Kuhnert, E.; Surup, F.; Halecker, S.; Stadler, M. Minutellins A-D, azaphilones from the stromata of Annulohypoxylon minutellum (Xylariaceae). Phytochemistry 2017, 137, 66–71. [Google Scholar] [CrossRef]
- Daranagama, D.A.; Camporesi, E.; Tian, Q.; Liu, X.; Chamyuang, S.; Stadler, M.; Hyde, K.D. Anthostomella is polyphyletic comprising several genera in Xylariaceae. Fungal Divers. 2015, 73, 203–238. [Google Scholar] [CrossRef]
- Triebel, D.; Peršoh, D.; Wollweber, H.; Stadler, M. Phylogenetic relationships among Daldinia, Entonaema, and Hypoxylon as inferred from ITS nrDNA analyses of Xylariales. Nova Hedw. 2005, 80, 25–43. [Google Scholar] [CrossRef]
- Stadler, M.; Laessoe, T.; Fournier, J.; Decock, C.; Schmieschek, B.; Tichy, H.V.; Persoh, D. A polyphasic taxonomy of Daldinia (Xylariaceae). Stud. Mycol. 2014, 77, 1–143. [Google Scholar] [CrossRef] [PubMed]
- Bitzer, J.; Laessoe, T.; Fournier, J.; Kummer, V.; Decock, C.; Tichy, H.V.; Piepenbring, M.; Persoh, D.; Stadler, M. Affinities of Phylacia and the daldinoid Xylariaceae, inferred from chemotypes of cultures and ribosomal DNA sequences. Mycol. Res. 2008, 112, 251–270. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Castlebury, L.A.; Miller, A.N.; Huhndorf, S.M.; Schoch, C.L.; Seifert, K.A.; Rossman, A.Y.; Rogers, J.D.; Kohlmeyer, J.; Volkmann-Kohlmeyer, B.; et al. An overview of the systematics of the Sordariomycetes based on a four-gene phylogeny. Mycologia 2017, 98, 1076–1087. [Google Scholar] [CrossRef]
- Koukol, O.; Kelnarová, I.; Černý, K.; Woodward, S. Recent observations of sooty bark disease of sycamore maple in Prague (Czech Republic) and the phylogenetic placement of Cryptostroma corticale. For. Pathol. 2015, 45, 21–27. [Google Scholar] [CrossRef]
- Stadler, M.; Eric, K.; Peršoh, D.; Fournier, J. The Xylariaceae as model example for a unified nomenclature following the “One Fungus-One Name” (1F1N) concept. Mycology 2013, 4, 5–21. [Google Scholar] [CrossRef]
- Pourmoghaddam, M.J.; Lambert, C.; Surup, F.; Khodaparast, S.A.; Krisai-Greilhuber, I.; Voglmayr, H.; Stadler, M. Discovery of a new species of the Hypoxylon rubiginosum complex from Iran and antagonistic activities of Hypoxylon spp. against the Ash Dieback pathogen, Hymenoscyphus fraxineus, in dual culture. MycoKeys 2020, 66, 105–133. [Google Scholar] [CrossRef]
- Li, G.J.; Hyde, K.D.; Zhao, R.L.; Hongsanan, S.; Abdel-Aziz, F.A.; Abdel-Wahab, M.A.; Alvarado, P.; Alves-Silva, G.; Ammirati, J.F.; Ariyawansa, H.A. Fungal diversity notes 253–366: taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 78, 1–237. [Google Scholar] [CrossRef]
- Kuhnert, E.; Fournier, J.; Peršoh, D.; Luangsa-ard, J.J.D.; Stadler, M. New Hypoxylon species from Martinique and new evidence on the molecular phylogeny of Hypoxylon based on ITS rDNA and β-tubulin data. Fungal Divers. 2013, 64, 181–203. [Google Scholar] [CrossRef]
- Bills, G.F.; Gonzalez-Menendez, V.; Martin, J.; Platas, G.; Fournier, J.; Persoh, D.; Stadler, M. Hypoxylon pulicicidum sp. nov. (Ascomycota, Xylariales), a pantropical insecticide-producing endophyte. PLoS ONE 2012, 7, e46687. [Google Scholar] [CrossRef] [PubMed]
- Stadler, M.; Flessa, F.; Rambold, G.; Peršoh, D.; Stadler, M.; Fournier, J.; Læssøe, T.; Chlebicki, A.; Lechat, C.; Peršoh, D. Chemotaxonomic and phylogenetic studies of Thamnomyces (Xylariaceae). Mycoscience 2010, 51, 189–207. [Google Scholar] [CrossRef]
- Sir, E.B. A new species of Daldinia (Xylariaceae) from the Argentine subtropical montane forest. Mycosphere 2016, 7, 1378–1388. [Google Scholar] [CrossRef]
- Becker, K.; Wongkanoun, S.; Wessel, A.-C.; Bills, G.F.; Stadler, M.; Luangsa-ard, J.J. Phylogenetic and chemotaxonomic studies confirm the affinities of Stromatoneurospora phoenix to the coprophilous Xylariaceae. J. Fungi 2020, 6. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Menendez, V.; Asensio, F.; Moreno, C.; de Pedro, N.; Monteiro, M.C.; de la Cruz, M.; Vicente, F.; Bills, G.F.; Reyes, F.; Genilloud, O.; et al. Assessing the effects of adsorptive polymeric resin additions on fungal secondary metabolite chemical diversity. Mycology 2014, 5, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Stadler, M.; Tichy, H.-V.; Katsiou, E.; Hellwig, V. Chemotaxonomy of Pochonia and other conidial fungi with Verticillium-like anamorphs. Mycol. Prog. 2003, 2, 95–122. [Google Scholar] [CrossRef]
- Becker, K.; Wessel, A.-C.; Luangsa-ard, J.J.; Stadler, M. Viridistratins A−C, antimicrobial and cytotoxic benzo[j]fluoranthenes from stromata of Annulohypoxylon viridistratum (Hypoxylaceae, Ascomycota). Biomolecules 2020, 10. [Google Scholar] [CrossRef]
- Sandargo, B.; Michehl, M.; Praditya, D.; Steinmann, E.; Stadler, M.; Surup, F. Antiviral meroterpenoid rhodatin and sesquiterpenoids rhodocoranes A-E from the Wrinkled Peach Mushroom, Rhodotus palmatus. Org. Lett. 2019, 21, 3286–3289. [Google Scholar] [CrossRef]
- Stipanovic, R.D.; Wheeler, M.H. Accumulation of 3,3′-biflaviolin, a melanin shunt product, by tricyclazole-treated Thielaviopsis basicola. Pestic. Biochem. Physiol. 1980, 13, 198–201. [Google Scholar] [CrossRef]
- Astill, B.D.; Roberts, J.C. 654. Studies in mycological chemistry. Part I. Flaviolin, 2(or 3):5:7-trihydroxy-1:4-naphthaquinone, a metabolic product of Aspergillus citricus (Wehmer) Mosseray. J. Chem. Soc. 1953. [Google Scholar] [CrossRef]
- Davies, J.E.; King, F.E.; Roberts, J.C. The structure of flaviolin. Chem. Ind. 1954, 1110–1111. [Google Scholar] [CrossRef]
- Wheeler, M.H.; Stipanovic, R.D. Melanin biosynthesis in Thielaviopsis basicola. Exp. Mycol. 1979, 3, 340–350. [Google Scholar] [CrossRef]
- Kumagai, Y.; Shinkai, Y.; Miura, T.; Cho, A.K. The chemical biology of naphthoquinones and its environmental implications. Annu. Rev. Pharmacol. Toxicol. 2012, 52, 221–247. [Google Scholar] [CrossRef] [PubMed]
- Klotz, L.O.; Hou, X.; Jacob, C. 1,4-naphthoquinones: from oxidative damage to cellular and inter-cellular signaling. Molecules 2014, 19, 14902–14918. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Species | Status | Strain | Origin | GenBank Acc. No. of DNA Loci | References | |||
|---|---|---|---|---|---|---|---|---|
| ITS | LSU | RPB2 | TUB2 | |||||
| Annulohypoxylon annulatum | ET | CBS 140775 | Texas | KY610418 | KY610418 | KY624263 | KX376353 | [2,30] |
| A. annulatum | DSM 107931 | Texas | MK287534 AJ390397 | MK287546 | MK287559 | MK287572 | [20] | |
| A. moriforme | CBS 123579 | Martinique | KX376321 | KY610425 | KY624289 | KX271261 | [2,31] | |
| A. nitens | MFLUCC 12.0832 | Thailand | KJ934991 | KJ934992 | KJ934994 | KJ934993 | [32] | |
| A. stygium | MUCL 54601 | French Guinea | KY610409 | KY610475 | KY624292 | KX271263 | [2] | |
| A. truncatum | ET | CBS 140778 | Texas | KY610419 | KY610419 | KY624277 | KX376352 | [2,30] |
| A. truncatum | DSM 107925 | Texas | MK287531 | MK287543 | MK287556 | MK287569 | [20] | |
| Daldinia concentrica | CBS 113277 | Germany | AY616683 | KY610434 | KY624243 | KC977274 | [2,12,33] | |
| D. eschscholtzii | MUCL 45435 | Benin | JX658484 | KY610437 | KY624246 | KC977266 | [2,12,34] | |
| D. petriniae | ET | MUCL 49214 | Austria | AM749937 | KY610439 | KY624248 | KC977261 | [2,12,35] |
| D. placentiformis | MUCL 47603 | Mexico | AM749921 | KY610440 | KY624249 | KC977278 | [2,12,35] | |
| Entonaema liquescens | ATCC 46302 | USA | KY610389 | KY610443 | KY624253 | KX271248 | [2] | |
| Graphostroma platystoma | HT | CBS 270.87 | France | JX658535 | DQ836906 | KY624296 | HG934108 | [2,34,36,37] |
| Hypom. barbarensis | HT | STMA 14081 | Argentina | MK131720 | MK131718 | MK135891 | MK135893 | [3] |
| Hypom. monticulosa | ET | MUCL 54604 | French Guiana | KY610404 | KY610487 | KY624305 | KX271273 | [2] |
| Hypom. submonticulosa | CBS 115280 | France | KC968923 | KY610457 | KY624226 | KC977267 | [2,12] | |
| Hypoxylon carneum | MUCL 54177 | France | KY610400 | KY610480 | KY624297 | KX271270 | [2] | |
| H. cercidicola | CBS 119009 | France | KC968908 | KY610444 | KY624254 | KU684189 | [2,12] | |
| H. croceum | DSM 107926 | Texas | MK287535 | MK287547 | MK287560 | MK287573 | [20] | |
| H. croceum | DSM 107932 | Texas | MK287532 | MK287544 | MK287557 | MK287570 | [20] | |
| H. crocopeplum | CBS 119004 | France | KC968907 | KY610445 | KY624255 | KC977268 | [2,12] | |
| H. fendleri | MUCL 54792 | French Guinea | KF234421 | KY610481 | KY624298 | KF300547 | [2,12] | |
| H. fragiforme | ET | MUCL 51264 | Germany | KC477229 | KM186295 | MK887342 | KX271282 | [2,20,32,38] |
| H. fuscum | ET | CBS 113049 | France | KY610401 | KY610482 | KY624299 | KX271271 | [2] |
| H. griseobrunneum | T | CBS 331.73 | India | KY610402 | KY610483 | KY624300 | KC977303 | [2,12] |
| H. guilanense | HT | MUCL 57726 | Iran | MT214997 | MT214992 | MT212235 | MT212239 | [39] |
| H. haematostroma | ET | MUCL 53301 | Martinique | KC968911 | KY610484 | KY624301 | KC977291 | [2,12] |
| H. howeanum | MUCL 47599 | Germany | AM749928 | KY610448 | KY624258 | KC977277 | [2,12,35] | |
| H. hypomiltum | MUCL 51845 | Guadeloupe | KY610403 | KY610449 | KY624302 | KX271249 | [2] | |
| H. invadens | HT | MUCL 51475 | France | MT809133 | MT809132 | MT813037 | MT813038 | This Study |
| H. investiens | CBS 118183 | Malaysia | KC968925 | KY610450 | KY624259 | KC977270 | [2,12] | |
| H. lateripigmentum | HT | MUCL 53304 | Martinique | KC968933 | KY610486 | KY624304 | KC977290 | [2,12] |
| H. lenormandii | CBS 119003 | Ecuador | KC968943 | KY610452 | KY624261 | KC977273 | [2,12] | |
| H. lienhwacheense | MFLUCC 14-1231 | Thailand | KU604558 | MK287550 | MK287563 | KU159522 | [20,40] | |
| H. musceum | MUCL 53765 | Guadeloupe | KC968926 | KY610488 | KY624306 | KC977280 | [2,12] | |
| H. olivaceopigmentum | T | DSM 107924 | Texas | MK287530 | MK287542 | MK287555 | MK287568 | [20] |
| H. papillatum | HT | ATCC 58729 | USA | KC968919 | KY610454 | KY624223 | KC977258 | [2,12] |
| H. perforatum | CBS 115281 | France | KY610391 | KY610455 | KY624224 | KX271250 | [2] | |
| H. petriniae | HT | CBS 114746 | France | KY610405 | KY610491 | KY624279 | KX271274 | [2,30] |
| H. pilgerianum | STMA 13455 | Martinique | KY610412 | KY610412 | KY624308 | KY624315 | [2] | |
| H. porphyreum | CBS 119022 | France | KC968921 | KY610456 | KY624225 | KC977264 | [2,41] | |
| H. pulicicidum | HT | CBS 122622 | Martinique | JX183075 | KY610492 | KY624280 | JX183072 | [2,42] |
| H. rickii | ET | MUCL 53309 | Martinique | KC968932 | KY610416 | KY624281 | KC977288 | [2,12] |
| H. rubiginosum | ET | MUCL 52887 | Germany | KC477232 | KY610469 | KY624266 | KY624311 | [2,38] |
| H. samuelsii | ET | MUCL 51843 | Guadeloupe | KC968916 | KY610466 | KY624269 | KC977286 | [2,12] |
| H. texense | T | DSM 107933 | Texas | MK287536 | MK287548 | MK287561 | MK287574 | [20] |
| H. ticinense | CBS 115273 | France | JQ009317 | KY610471 | KY624272 | AY951757 | [2,4] | |
| H. trugodes | ET | MUCL 54794 | Sri Lanka | KF234422 | KY610493 | KY624282 | KF300548 | [2,12] |
| H. vogesiacum | CBS 115273 | France | KC968920 | KY610417 | KY624283 | KX271275 | [2,12] | |
| Jackrogersella cohaerens | CBS 119126 | Germany | KY610396 | KY610497 | KY624270 | KY624314 | [2] | |
| J. minutella | CBS 119015 | Portugal | KY610381 | KY610424 | KY624235 | KX271240 | [2] | |
| J. multiformis | ET | CBS 119016 | Germany | KC477234 | KY610473 | KY624290 | KX271262 | [2,12,30] |
| Pyrenopolyporus hunteri | ET | MUCL 52673 | Ivory Coast | KY610421 | KY610472 | KY624309 | KU159530 | [2,30] |
| P. laminosus | HT | MUCL 53305 | Martinique | KC968934 | KY610485 | KY624303 | KC977292 | [2,12] |
| P.nicaraguensis | CBS 117739 | Burkina Faso | AM749922 | KY610489 | KY624307 | KC977272 | [2,12,35] | |
| Rhopalostroma angolense | CBS 126414 | Ivory Coast | KY610420 | KY610459 | KY624228 | KX271277 | [2] | |
| Thamnomyces dendroidea | HT | CBS 123578 | French Guinea | FN428831 | KY610467 | KY624232 | KY624313 | [2,43] |
| Xylaria hypoxylon | ET | CBS 122620 | Sweden | KY610407 | KY610495 | KY624231 | KX271279 | [2,44] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Becker, K.; Lambert, C.; Wieschhaus, J.; Stadler, M. Phylogenetic Assignment of the Fungicolous Hypoxylon invadens (Ascomycota, Xylariales) and Investigation of its Secondary Metabolites. Microorganisms 2020, 8, 1397. https://doi.org/10.3390/microorganisms8091397
Becker K, Lambert C, Wieschhaus J, Stadler M. Phylogenetic Assignment of the Fungicolous Hypoxylon invadens (Ascomycota, Xylariales) and Investigation of its Secondary Metabolites. Microorganisms. 2020; 8(9):1397. https://doi.org/10.3390/microorganisms8091397
Chicago/Turabian StyleBecker, Kevin, Christopher Lambert, Jörg Wieschhaus, and Marc Stadler. 2020. "Phylogenetic Assignment of the Fungicolous Hypoxylon invadens (Ascomycota, Xylariales) and Investigation of its Secondary Metabolites" Microorganisms 8, no. 9: 1397. https://doi.org/10.3390/microorganisms8091397
APA StyleBecker, K., Lambert, C., Wieschhaus, J., & Stadler, M. (2020). Phylogenetic Assignment of the Fungicolous Hypoxylon invadens (Ascomycota, Xylariales) and Investigation of its Secondary Metabolites. Microorganisms, 8(9), 1397. https://doi.org/10.3390/microorganisms8091397