The Ability of Respiratory Commensal Bacteria to Beneficially Modulate the Lung Innate Immune Response Is a Strain Dependent Characteristic

 , , , , ,
, , , , , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Animals and TLRs Agonist Administration
2.3. Respiratory Syncytial Virus and Streptococcus Pneumoniae Infections
2.4. Cytokine Concentrations in Broncho-Alveolar Lavages (BAL)
2.5. Lung Cell Suspensions and Flow Cytometry Studies
2.6. Lung Tissue Injury Studies
2.7. Statistical Analysis
3. Results
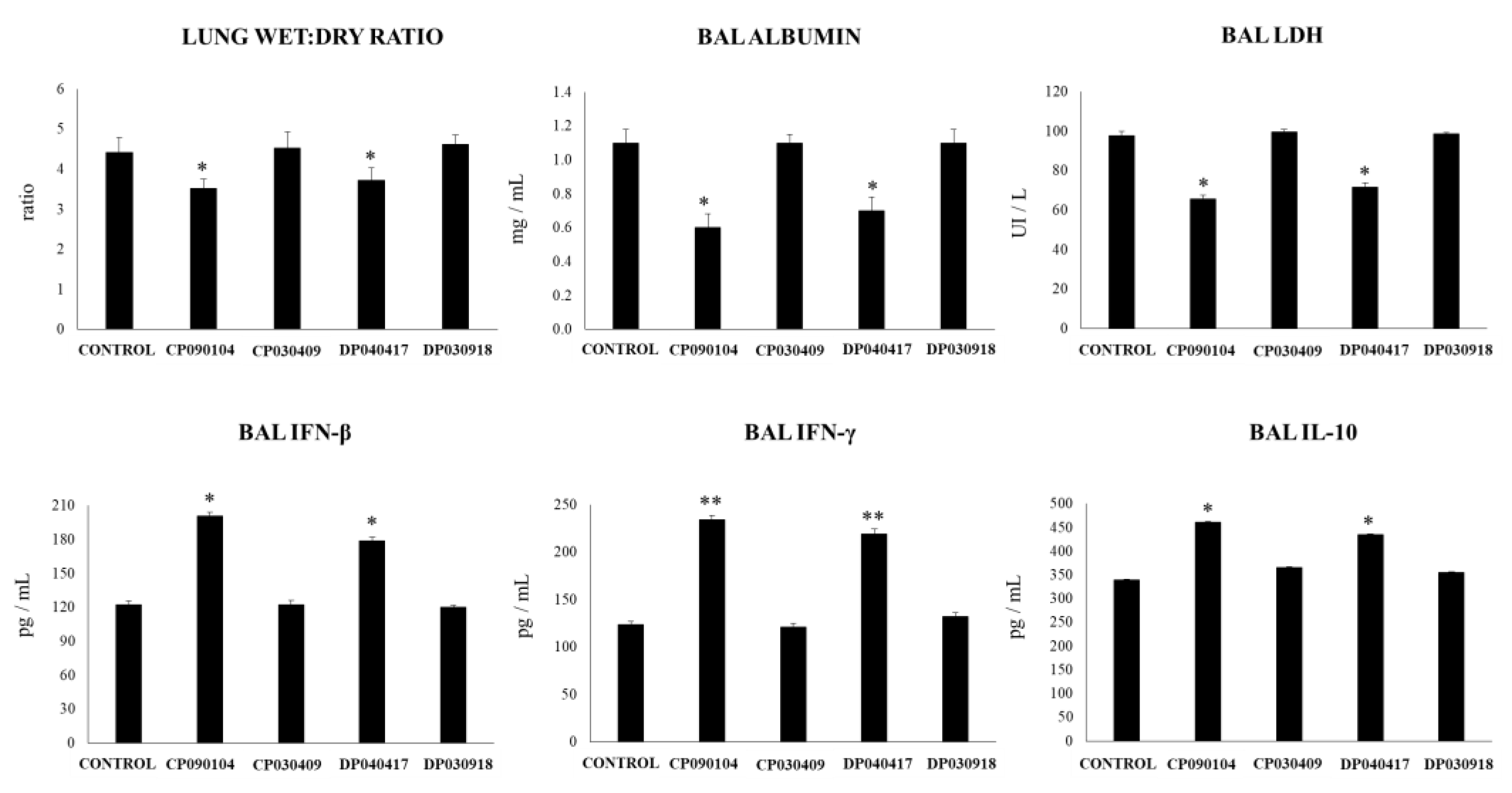
3.1. Strain-Dependent Ability of Respiratory Commensal Bacteria to Modulate Lung Innate Immunity
3.2. Dolosigranulum Pigrum 040417 Modulates Respiratory and Systemic Cytokine Profiles
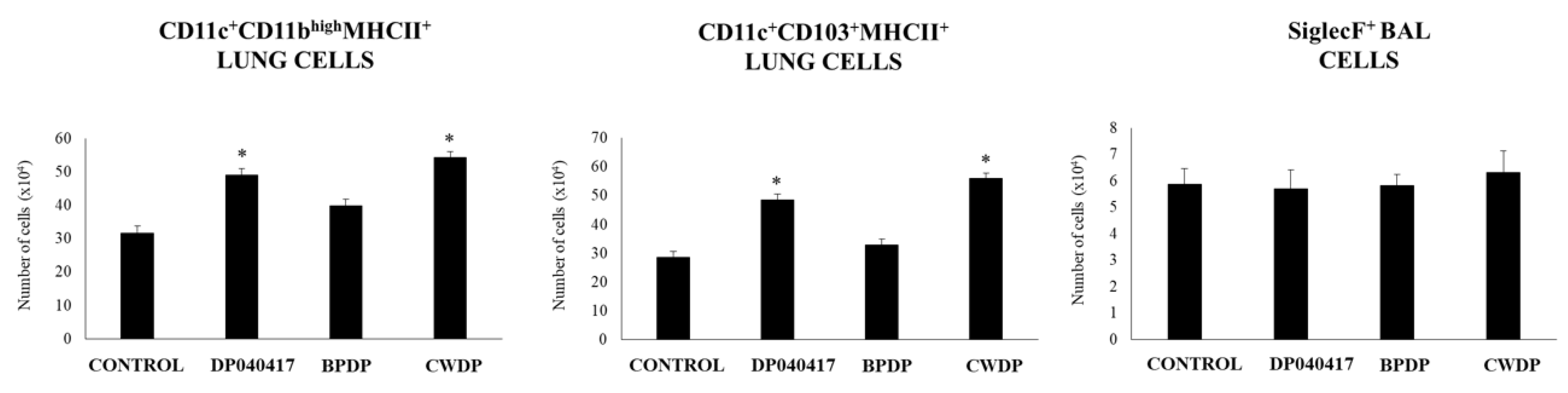
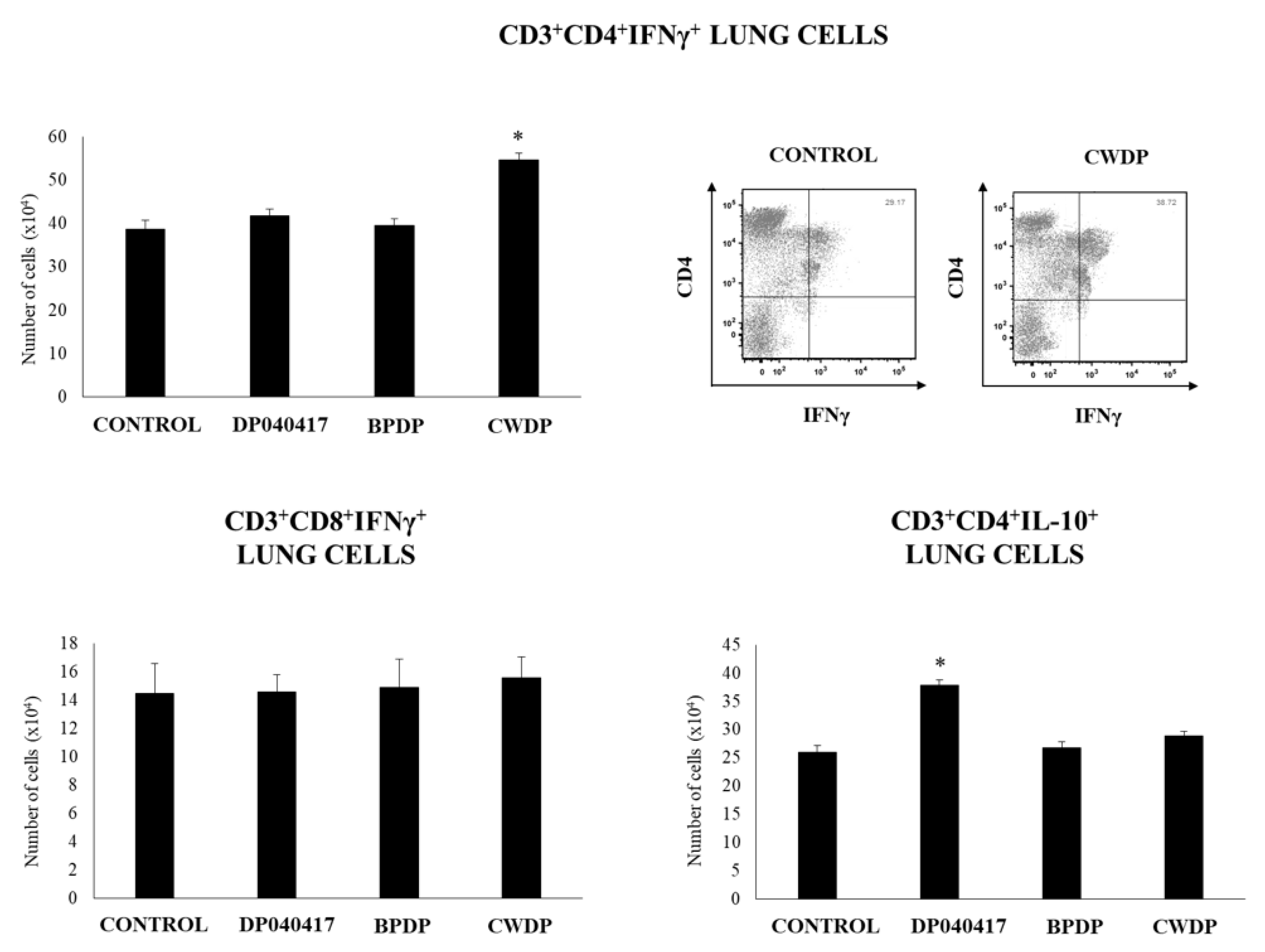
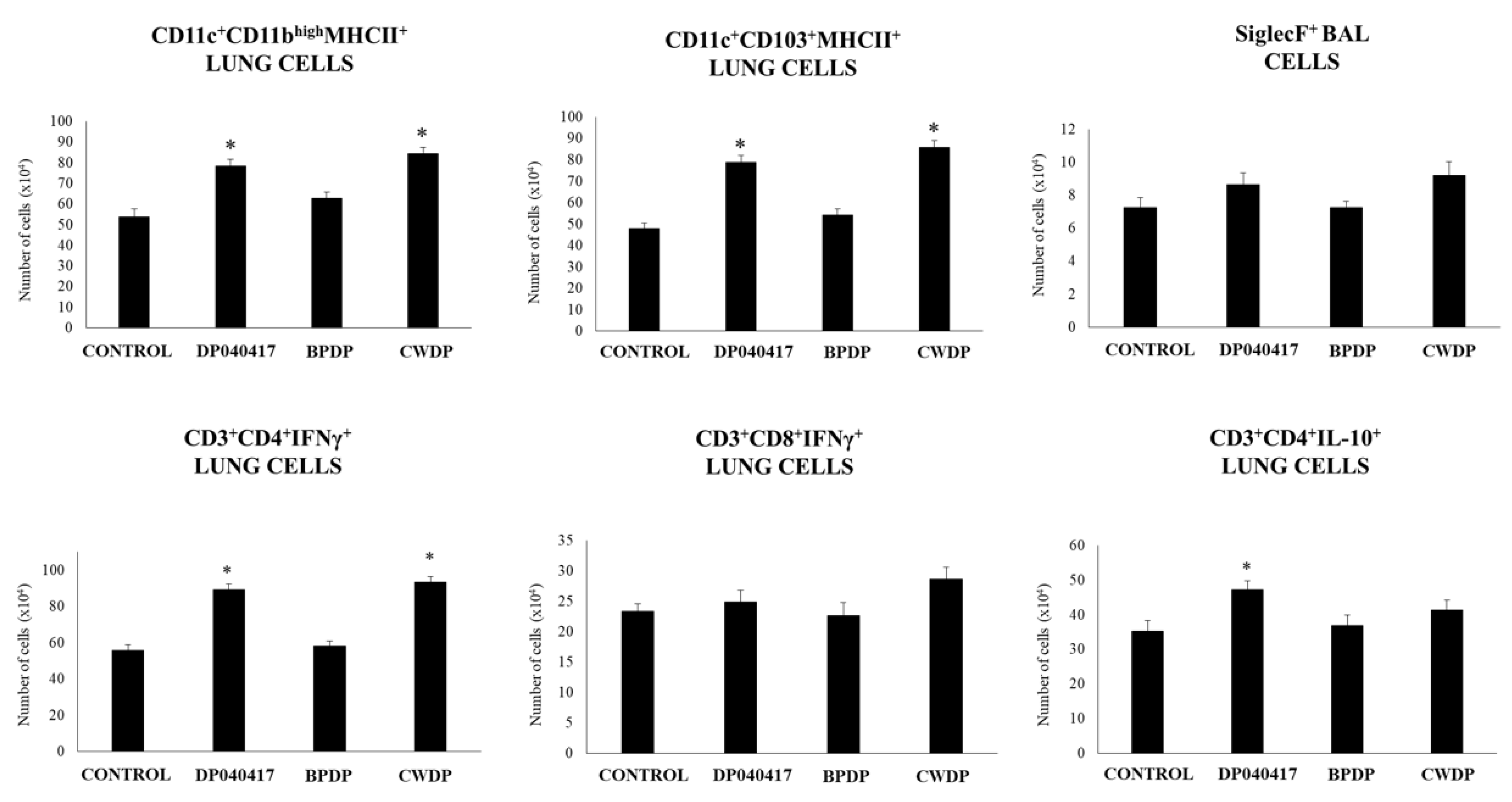
3.3. Dolosigranulum Pigrum 040417 Differentially Modulates Lung Immune Cells
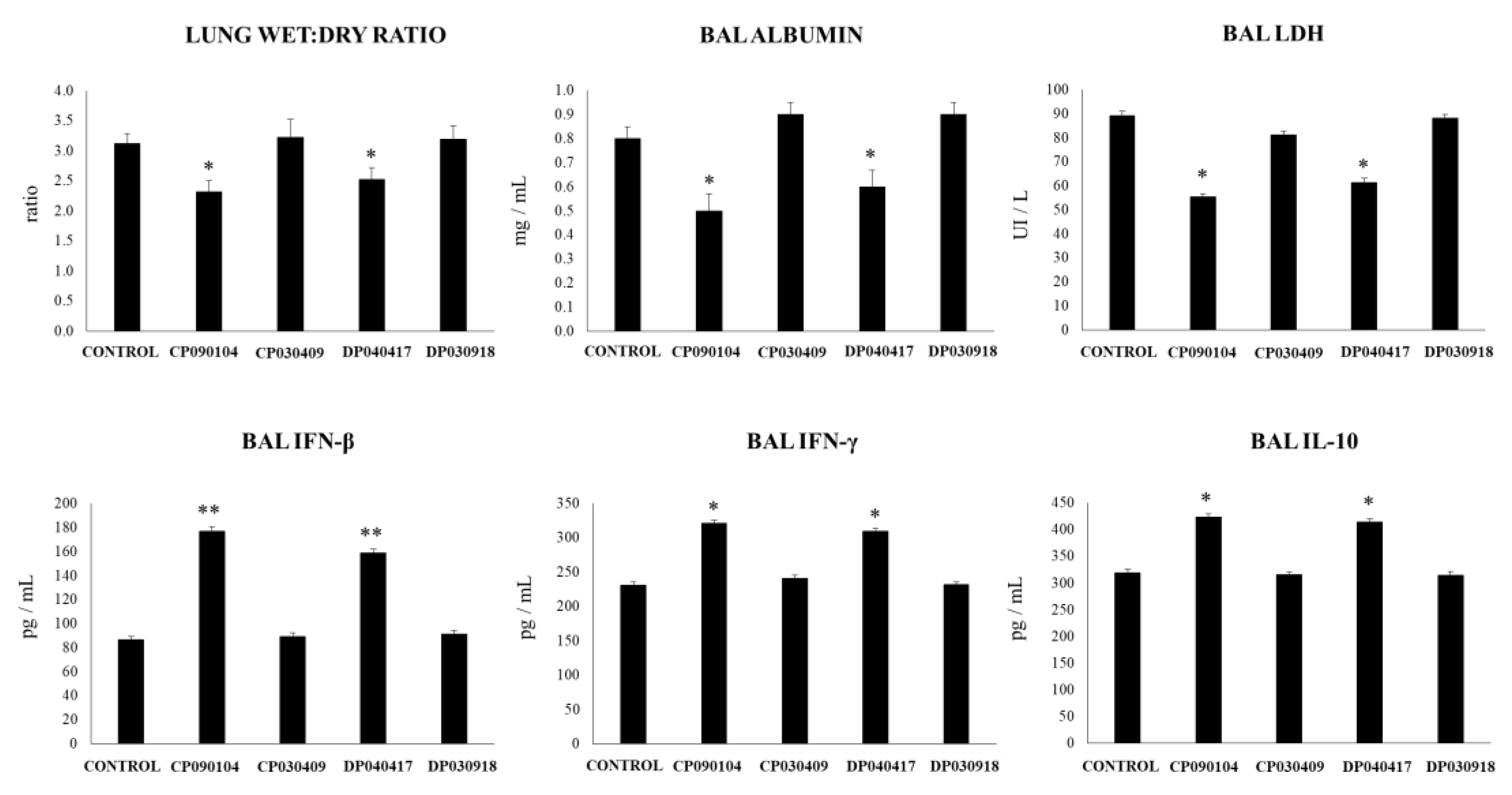
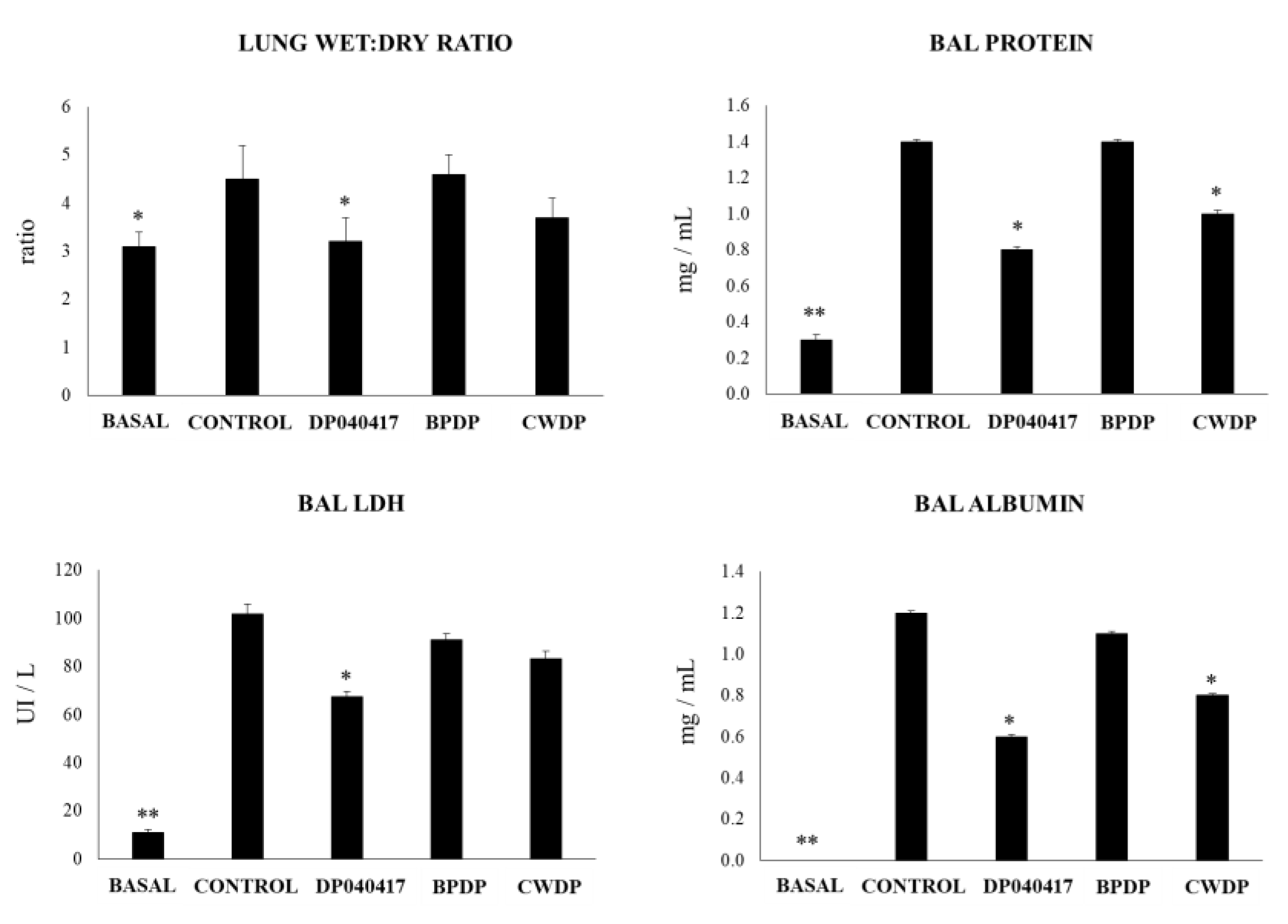
3.4. Dolosigranulum Pigrum 040417 Diminish the TLR3-Mediated Inflammatory Lung Damage
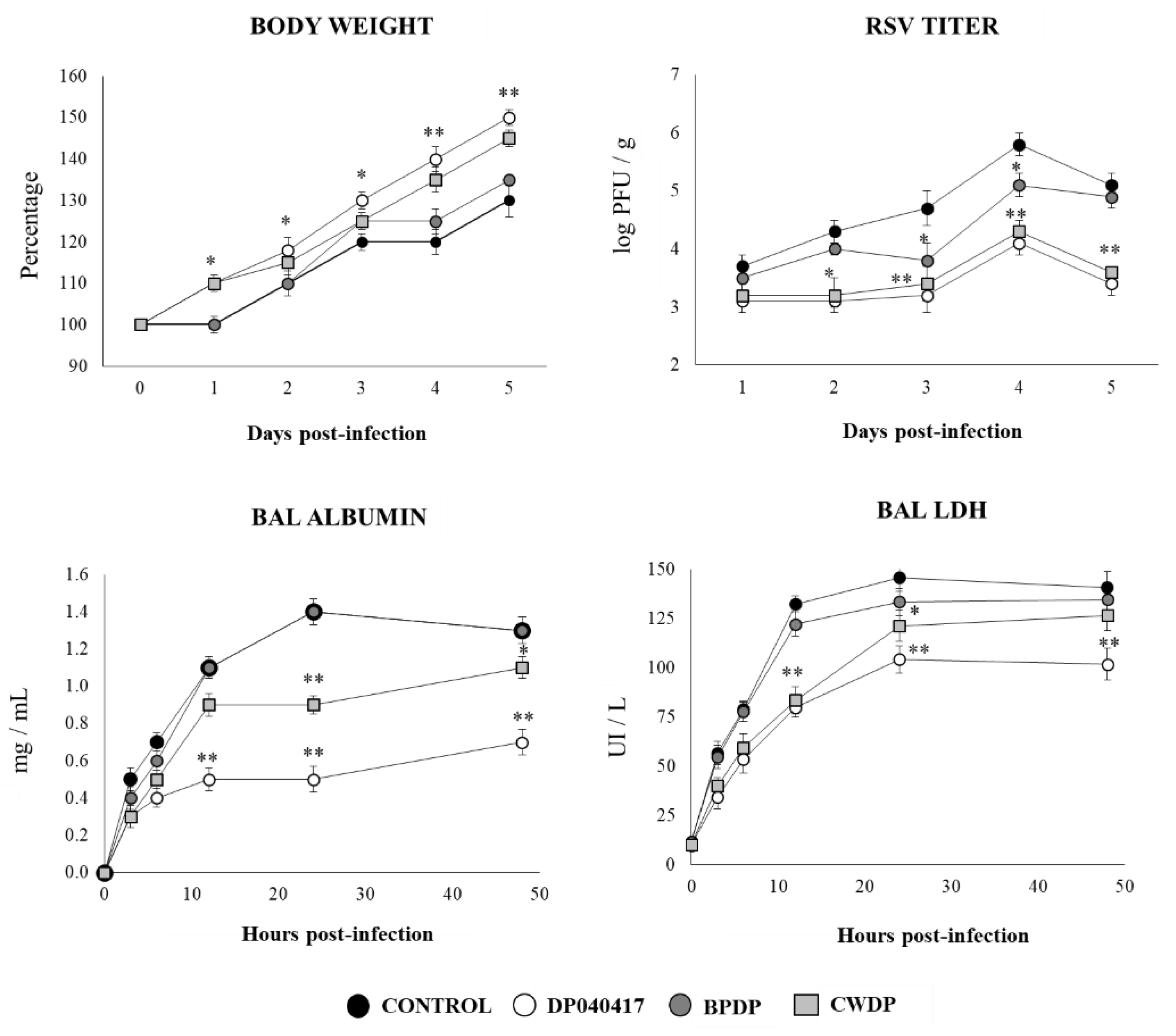
3.5. Dolosigranulum Pigrum 040417 Improves Resistance to RSV Infection
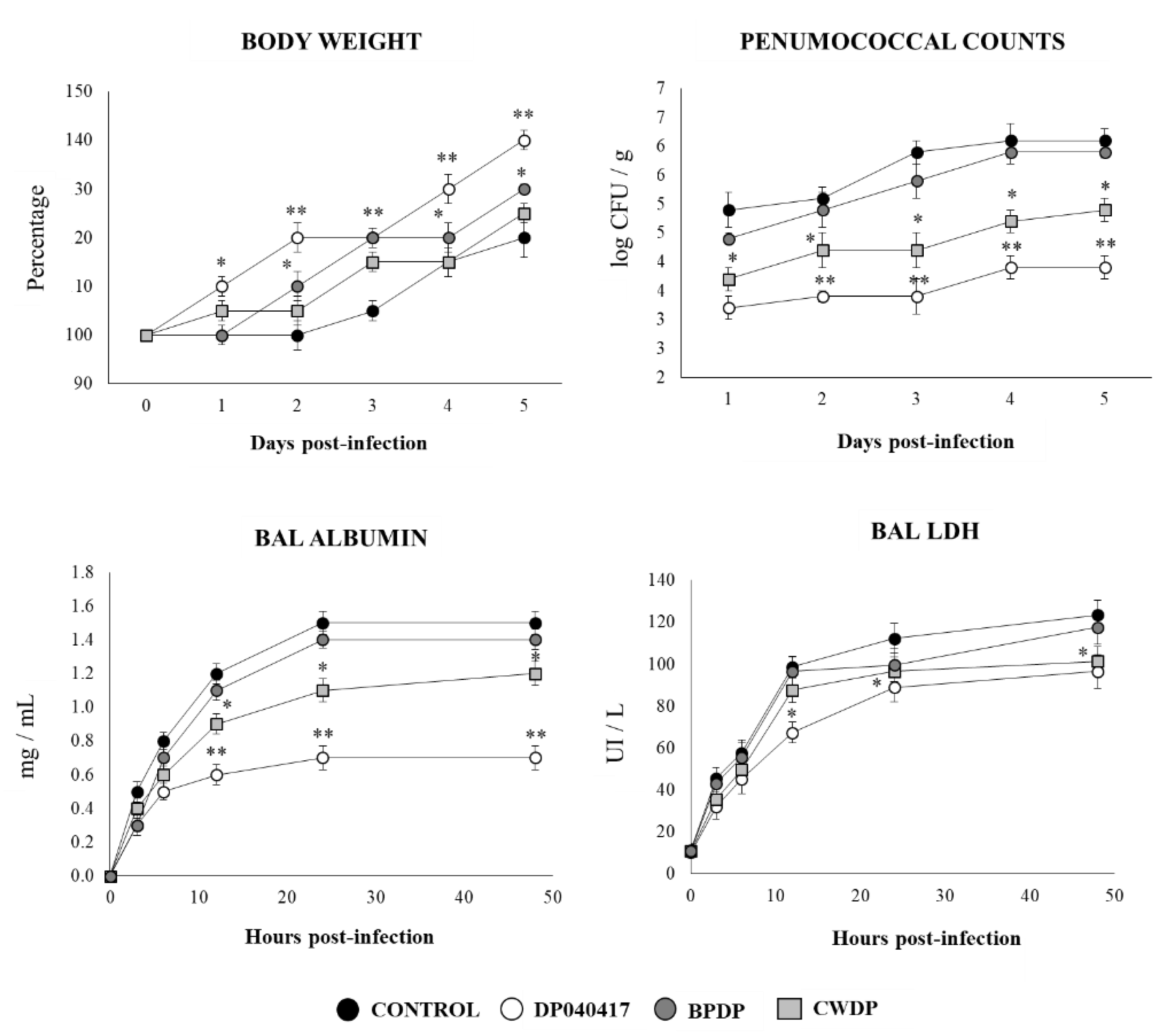
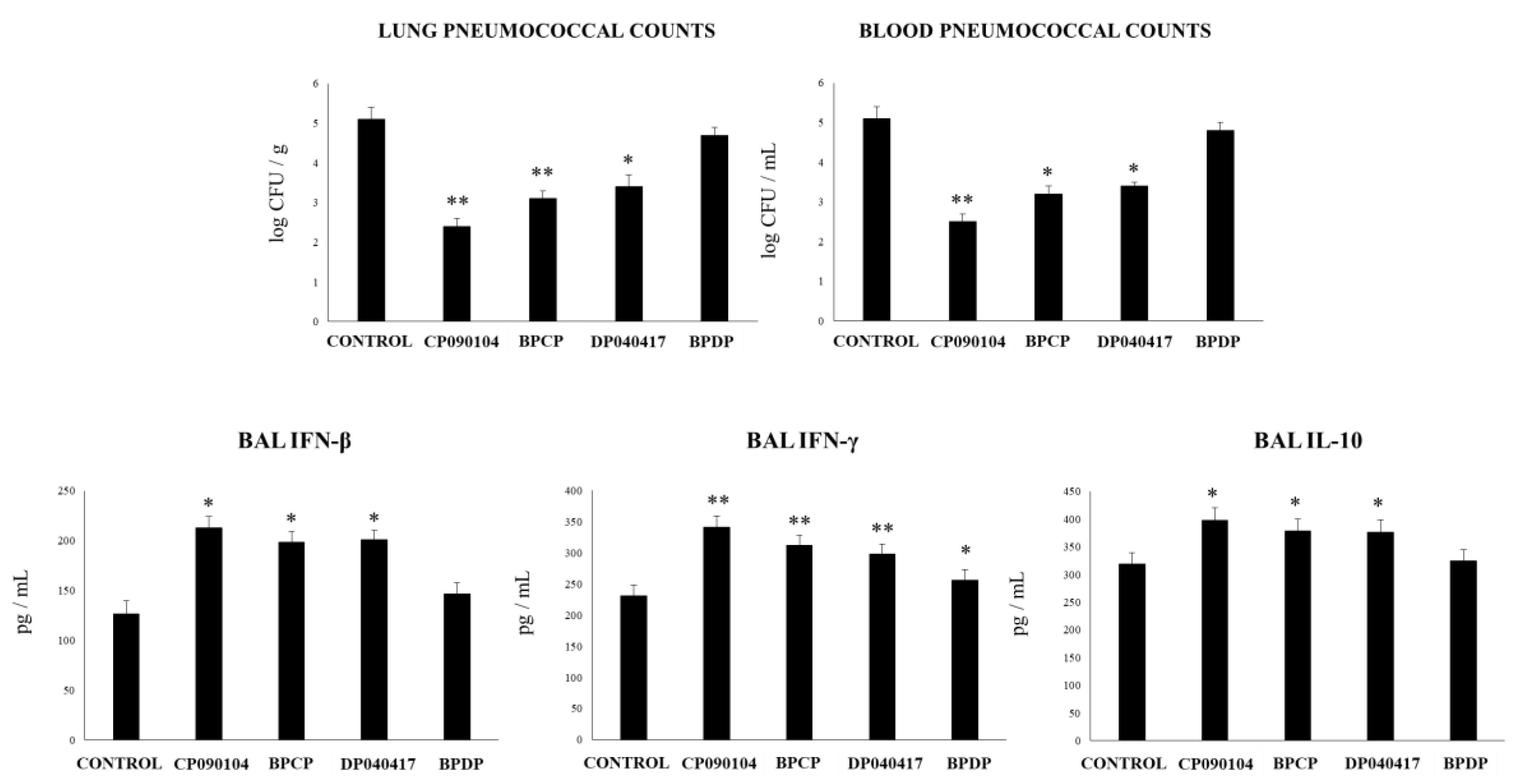
3.6. Respiratory Commensal Bacteria Improve Resistance to Pneumococcal Infection
4. Discussion
4.1. The Ability of Respiratory Commensal Bacteria to Modulate the Innate Immunity in the Respiratory Tract is a Strain-Dependent Characteristic
4.2. D. pigrum 040417, Similar to C. Pseudodiphteriticum 090104, Is Capable of Modulating the Respiratory Innate Immunity and Improve the Resistance to RSV and S. Pneumoniae Infections
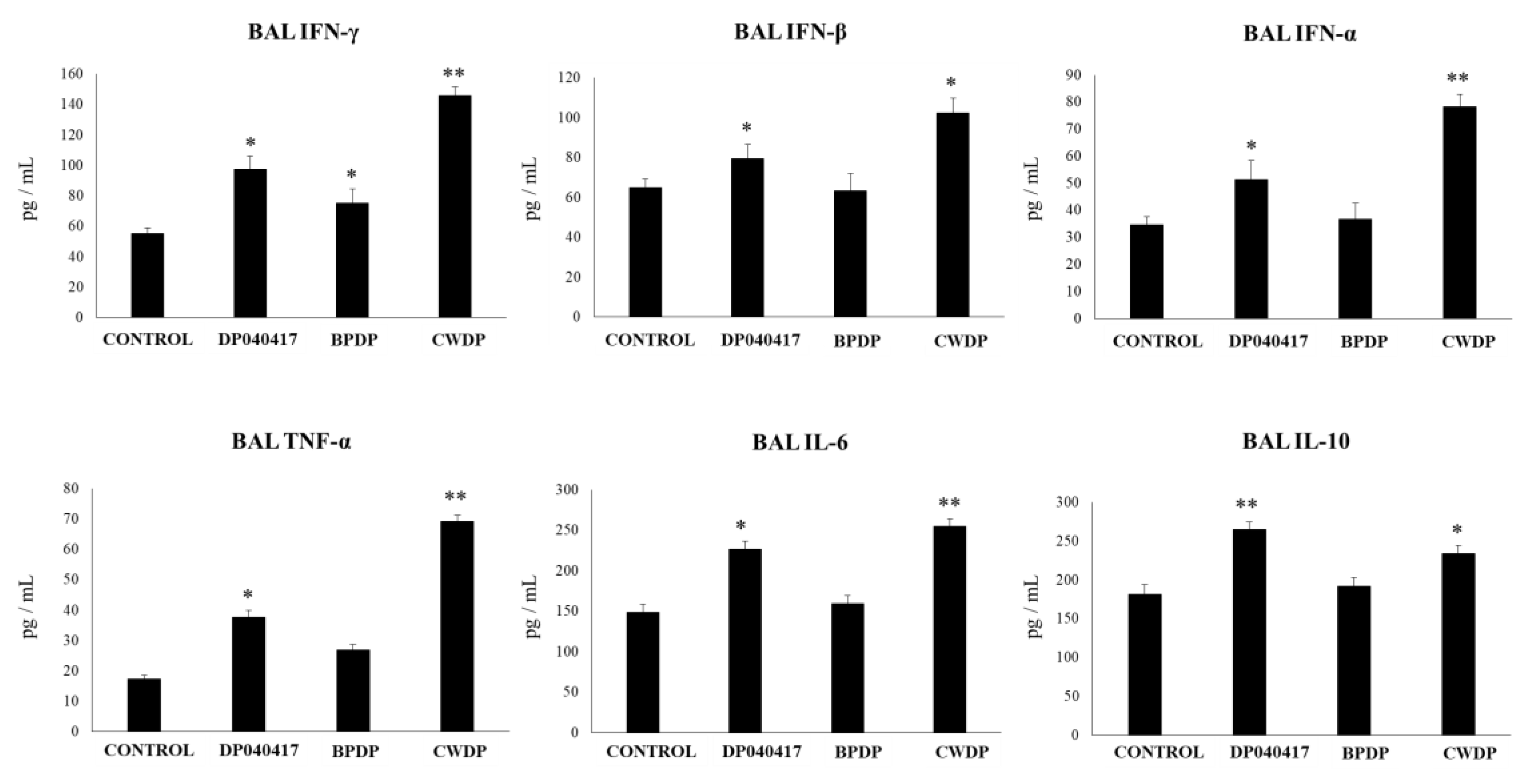
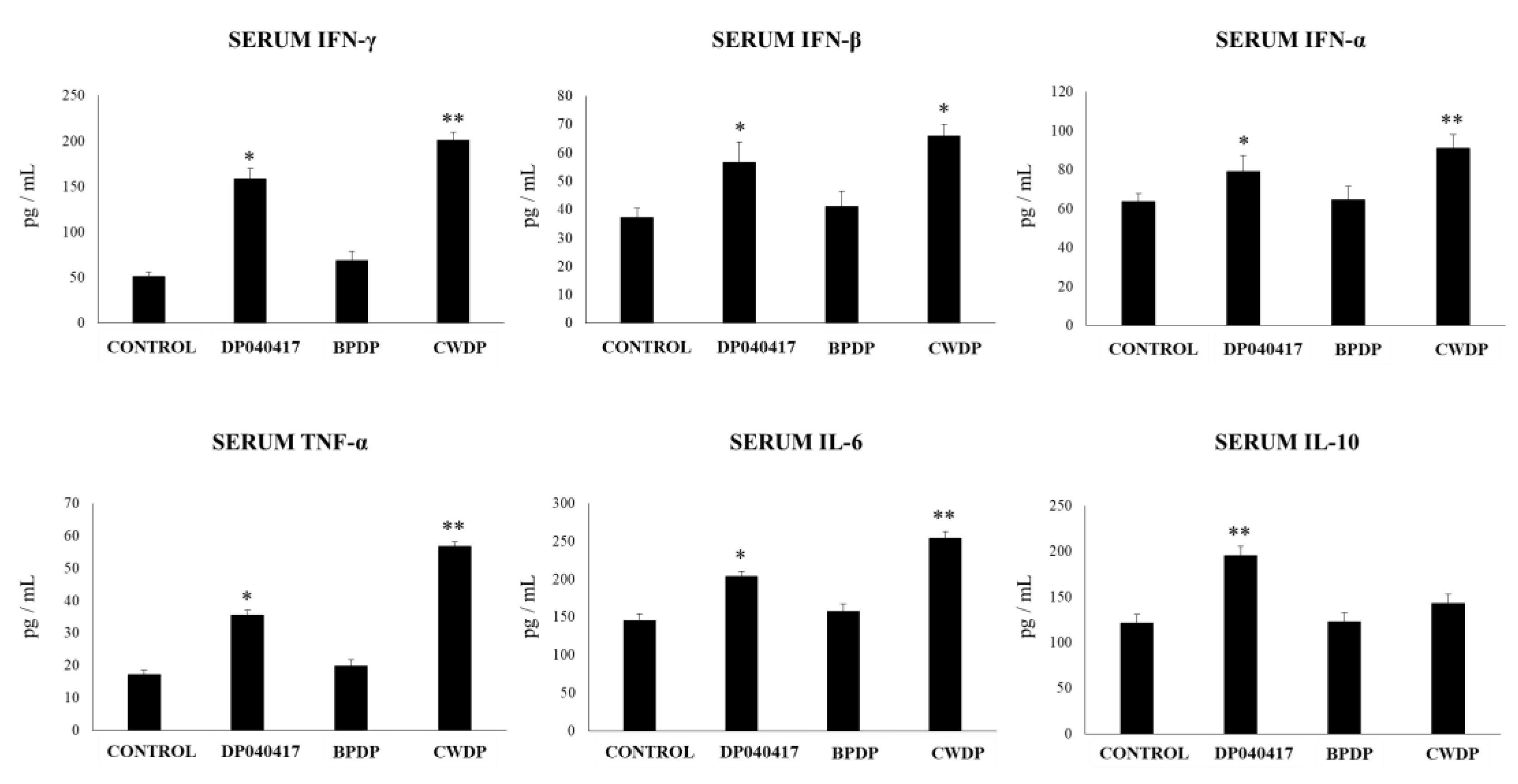
4.3. The Ability of BLP Derived from Respiratory Commensal Bacteria to Modulate the Innate Immunity in the Respiratory Tract is also a Strain Dependent Characteristic
4.4. The Cell Wall of D. pigrum 040417 is an Interesting Alternative to Beneficially Modulate the Respiratory Innate Immune Response in High-Risk Populations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Abubakar, A.; Malik, M.; Pebody, R.G.; Elkholy, A.; Khan, W.; Bellos, A.; Mala, P. World Health Organization Burden of acute respiratory disease of epidemic and pandemic potential in the WHO Eastern Mediterranean Region: A literature review. East. Mediterr. Heal. J. 2016, 22, 509–522. [Google Scholar] [CrossRef]
- Weinberger, D.; Givon-Lavi, N.; Shemer-Avni, Y.; Bar-Ziv, J.; Alonso, W.J.; Greenberg, D.; Dagan, R. Influence of Pneumococcal Vaccines and Respiratory Syncytial Virus on Alveolar Pneumonia, Israel. Emerg. Infect. Dis. 2013, 19, 1084–1091. [Google Scholar] [CrossRef]
- Cebey-López, M.; Pardo-Seco, J.; Gómez-Carballa, A.; Martinón-Torres, N.; Martinón-Sánchez, J.M.; Grande, A.J.; Calle, I.R.; Pinnock, E.; Salas, A.; Fink, C.; et al. Bacteremia in Children Hospitalized with Respiratory Syncytial Virus Infection. PLoS ONE 2016, 11, e0146599. [Google Scholar] [CrossRef]
- Hament, J.-M.; Aerts, P.C.; Fleer, A.; Van Dijk, H.; Harmsen, T.; Kimpen, J.L.L.; Wolfs, T.F. Direct Binding of Respiratory Syncytial Virus to Pneumococci: A Phenomenon That Enhances Both Pneumococcal Adherence to Human Epithelial Cells and Pneumococcal Invasiveness in a Murine Model. Pediatr. Res. 2005, 58, 1198–1203. [Google Scholar] [CrossRef]
- Smith, C.M.; Sandrini, S.; Datta, S.; Freestone, P.; Shafeeq, S.; Radhakrishnan, P.; Williams, G.; Glenn, S.M.; Kuipers, O.P.; Hirst, R.A.; et al. Respiratory syncytial virus increases the virulence of Streptococcus pneumoniae by binding to penicillin binding protein 1a. A new paradigm in respiratory infection. Am. J. Respir. Crit. Care Med. 2014, 190, 196–207. [Google Scholar] [CrossRef]
- Avadhanula, V.; Rodriguez, C.A.; DeVincenzo, J.P.; Wang, Y.; Webby, R.J.; Ulett, G.C.; Adderson, E.E. Respiratory Viruses Augment the Adhesion of Bacterial Pathogens to Respiratory Epithelium in a Viral Species- and Cell Type-Dependent Manner. J. Virol. 2006, 80, 1629–1636. [Google Scholar] [CrossRef]
- Avadhanula, V.; Wang, Y.; Portner, A.; Adderson, E. Nontypeable Haemophilus influenzae and Streptococcus pneumoniae bind respiratory syncytial virus glycoprotein. J. Med. Microbiol. 2007, 56, 1133–1137. [Google Scholar] [CrossRef]
- Martín, R.; Langella, P. Emerging Health Concepts in the Probiotics Field: Streamlining the Definitions. Front. Microbiol. 2019, 10, 1047. [Google Scholar] [CrossRef]
- Kayama, H.; Takeda, K. Polysaccharide A of Bacteroides fragilis: Actions on dendritic cells and T cells. Mol. Cell 2014, 54, 206–207. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Miquel, S.; Benevides, L.; Bridonneau, C.; Robert, V.; Hudault, S.; Chain, F.; Berteau, O.; Azevedo, V.; Chatel, J.M.; et al. Functional Characterization of Novel Faecalibacterium prausnitzii Strains Isolated from Healthy Volunteers: A Step Forward in the Use of F. prausnitzii as a Next-Generation Probiotic. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Bian, X.; Wu, W.; Yang, L.; Lv, L.; Wang, Q.; Li, Y.; Ye, J.; Fang, D.; Wu, J.; Jiang, X.; et al. Administration of Akkermansia muciniphila Ameliorates Dextran Sulfate Sodium-Induced Ulcerative Colitis in Mice. Front. Microbiol. 2019, 10, 2259. [Google Scholar] [CrossRef]
- Man, W.H.; Piters, W.D.S.; Bogaert, D. The microbiota of the respiratory tract: Gatekeeper to respiratory health. Nat. Rev. Genet. 2017, 15, 259–270. [Google Scholar] [CrossRef]
- Bellussi, L.M.; Passali, F.M.; Ralli, M.; De Vincentiis, M.; Greco, A.; Passali, D. An overview on upper respiratory tract infections and bacteriotherapy as innovative therapeutic strategy. Eur. Rev. Med Pharmacol. Sci. 2019, 23, 27–38. [Google Scholar]
- De Boeck, I.; Wittouck, S.; Wuyts, S.; Oerlemans, E.F.M.; Broek, M.F.L.V.D.; Vandenheuvel, D.; Vanderveken, O.; Lebeer, S. Comparing the Healthy Nose and Nasopharynx Microbiota Reveals Continuity As Well As Niche-Specificity. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Laufer, A.S.; Metlay, J.P.; Gent, J.F.; Fennie, K.; Kong, Y.; Pettigrew, M. Microbial Communities of the Upper Respiratory Tract and Otitis Media in Children. mBio 2011, 2, e00245-10. [Google Scholar] [CrossRef]
- Pettigrew, M.; Laufer, A.S.; Gent, J.F.; Kong, Y.; Fennie, K.; Metlay, J.P. Upper Respiratory Tract Microbial Communities, Acute Otitis Media Pathogens, and Antibiotic Use in Healthy and Sick Children. Appl. Environ. Microbiol. 2012, 78, 6262–6270. [Google Scholar] [CrossRef] [PubMed]
- Castillo, A.C.; Henares, D.; Brotons, P.; Galiana, A.; Rodríguez, J.C.; Mira, A.; Muñoz-Almagro, C. Nasopharyngeal Microbiota in Children With Invasive Pneumococcal Disease: Identification of Bacteria With Potential Disease-Promoting and Protective Effects. Front. Microbiol. 2019, 10, 11. [Google Scholar] [CrossRef]
- Wen, Z.; Xie, G.; Zhou, Q.; Qiu, C.; Li, J.; Hu, Q.; Dai, W.; Li, N.; Zheng, Y.; Wen, F. Distinct Nasopharyngeal and Oropharyngeal Microbiota of Children with Influenza A Virus Compared with Healthy Children. BioMed Res. Int. 2018, 2018, 1–9. [Google Scholar] [CrossRef]
- Kanmani, P.; Clua, P.; Vizoso-Pinto, M.G.; Rodriguez, C.; Alvarez, S.; Мельникoв, В.Г.; Takahashi, H.; Kitazawa, H.; Villena, J. Respiratory Commensal Bacteria Corynebacterium pseudodiphtheriticum Improves Resistance of Infant Mice to Respiratory Syncytial Virus and Streptococcus pneumoniae Superinfection. Front. Microbiol. 2017, 8, 8. [Google Scholar] [CrossRef]
- Kolling, Y.; Salva, S.; Villena, J.; Alvarez, S. Are the immunomodulatory properties of Lactobacillus rhamnosus CRL1505 peptidoglycan common for all Lactobacilli during respiratory infection in malnourished mice? PLoS ONE 2018, 13, e0194034. [Google Scholar] [CrossRef]
- Tonetti, F.R.; Arce, L.; Salva, S.; Alvarez, S.; Takahashi, H.; Kitazawa, H.; Vizoso-Pinto, M.G.; Villena, J. Immunomodulatory Properties of Bacterium-Like Particles Obtained From Immunobiotic Lactobacilli: Prospects for Their Use as Mucosal Adjuvants. Front. Immunol. 2020, 11, 15. [Google Scholar] [CrossRef] [PubMed]
- Shida, K.; Kiyoshima-Shibata, J.; Kaji, R.; Nagaoka, M.; Nanno, M. Peptidoglycan from lactobacilli inhibits interleukin-12 production by macrophages induced by Lactobacillus casei through Toll-like receptor 2-dependent and independent mechanisms. Immunology 2009, 128, e858–e869. [Google Scholar] [CrossRef] [PubMed]
- Tomosada, Y.; Chiba, E.; Zelaya, M.H.D.R.; Takahashi, T.; Tsukida, K.; Kitazawa, H.; Alvarez, S.; Villena, J. Nasally administered Lactobacillus rhamnosus strains differentially modulate respiratory antiviral immune responses and induce protection against respiratory syncytial virus infection. BMC Immunol. 2013, 14, 40. [Google Scholar] [CrossRef]
- Laiño, J.; Villena, J.; Suvorov, A.; Zelaya, H.; Moyano, R.O.; Salva, S.; Alvarez, S. Nasal immunization with recombinant chimeric pneumococcal protein and cell wall from immunobiotic bacteria improve resistance of infant mice to Streptococcus pneumoniae infection. PLoS ONE 2018, 13, e0206661. [Google Scholar] [CrossRef] [PubMed]
- Villena, J.; Chiba, E.; Tomosada, Y.; Salva, S.; Marranzino, G.; Kitazawa, H.; Alvarez, S. Orally administered Lactobacillus rhamnosus modulates the respiratory immune response triggered by the viral pathogen-associated molecular pattern poly(I:C). BMC Immunol. 2012, 13, 53. [Google Scholar] [CrossRef] [PubMed]
- Clua, P.; Kanmani, P.; Zelaya, H.; Tada, A.; Kober, A.K.M.H.; Salva, S.; Alvarez, S.; Kitazawa, H.; Villena, J. Peptidoglycan from Immunobiotic Lactobacillus rhamnosus Improves Resistance of Infant Mice to Respiratory Syncytial Viral Infection and Secondary Pneumococcal Pneumonia. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef]
- Zähringer, U.; Lindner, B.; Inamura, S.; Heine, H.; Alexander, C. TLR2—Promiscuous or specific? A critical re-evaluation of a receptor expressing apparent broad specificity. Immunobiology 2008, 213, 205–224. [Google Scholar] [CrossRef]
- Bogaert, D.; Keijser, B.; Huse, S.; Rossen, J.W.A.; Veenhoven, R.; Van Gils, E.; Bruin, J.; Montijn, R.; Bonten, M.; Sanders, E. Variability and Diversity of Nasopharyngeal Microbiota in Children: A Metagenomic Analysis. PLoS ONE 2011, 6, e17035. [Google Scholar] [CrossRef]
- Bosch, A.A.T.M.; Piters, W.D.S.; Van Houten, M.A.; Chu, M.L.J.N.; Biesbroek, G.; Kool, J.; Pernet, P.; De Groot, P.-K.C.M.; Eijkemans, M.J.C.; Keijser, B.J.F.; et al. Maturation of the Infant Respiratory Microbiota, Environmental Drivers, and Health Consequences. A Prospective Cohort Study. Am. J. Respir. Crit. Care Med. 2017, 196, 1582–1590. [Google Scholar] [CrossRef]
- Chonmaitree, T.; Jennings, K.; Golovko, G.; Khanipov, K.; Pimenova, M.; Patel, J.A.; McCormick, D.P.; Loeffelholz, M.J.; Fofanov, Y. Nasopharyngeal microbiota in infants and changes during viral upper respiratory tract infection and acute otitis media. PLoS ONE 2017, 12, e0180630. [Google Scholar] [CrossRef]
- Nishimura, M.; Naito, S. Tissue-Specific mRNA Expression Profiles of Human Toll-Like Receptors and Related Genes. Boil. Pharm. Bull. 2005, 28, 886–892. [Google Scholar] [CrossRef] [PubMed]
- Hoebe, K.; Georgel, P.; Rutschmann, S.; Du, X.; Mudd, S.; Crozat, K.; Sovath, S.; Shamel, L.; Hartung, T.; Zähringer, U.; et al. CD36 is a sensor of diacylglycerides. Nature 2005, 433, 523–527. [Google Scholar] [CrossRef]
- Pabst, R.; Durak, D.; Roos, A.; Lührmann, A.; Tschernig, T. TLR2/6 stimulation of the rat lung: Effects on lymphocyte subsets, natural killer cells and dendritic cells in different parts of the air-conducting compartments and at different ages. Immunology 2009, 126, 132–139. [Google Scholar] [CrossRef]
- Reppe, K.; Tschernig, T.; Lührmann, A.; Van Laak, V.; Grote, K.; Zemlin, M.V.; Gutbier, B.; Müller, H.C.; Kursar, M.; Schütte, H.; et al. Immunostimulation with Macrophage-Activating Lipopeptide-2 Increased Survival in Murine Pneumonia. Am. J. Respir. Cell Mol. Boil. 2009, 40, 474–481. [Google Scholar] [CrossRef]
- Chen, H.; Cowan, M.J.; Hasday, J.D.; Vogel, S.N.; Medvedev, A.E. Tobacco Smoking Inhibits Expression of Proinflammatory Cytokines and Activation of IL-1R-Associated Kinase, p38, and NF-κB in Alveolar Macrophages Stimulated with TLR2 and TLR4 Agonists. J. Immunol. 2007, 179, 6097–6106. [Google Scholar] [CrossRef]
- Natarajan, S.; Kim, J.; Remick, D.G. Chronic pulmonary LPS tolerance induces selective immunosuppression while maintaining the neutrophilic response. Shock 2010, 33, 162–169. [Google Scholar] [CrossRef]
- Waters, V.; Sokol, S.; Reddy, B.; Soong, G.; Chun, J.; Prince, A. The Effect of Cyclosporin A on Airway Cell Proinflammatory Signaling and Pneumonia. Am. J. Respir. Cell Mol. Boil. 2005, 33, 138–144. [Google Scholar] [CrossRef]
- Netea, M.G.; Van De Veerdonk, F.; Verschueren, I.; Van Der Meer, J.W.M.; Kullberg, B.-J. Role of TLR1 and TLR6 in the host defense against disseminated candidiasis. FEMS Immunol. Med. Microbiol. 2008, 52, 118–123. [Google Scholar] [CrossRef]
- Gomez, J.C.; Yamada, M.; Martin, J.R.; Dang, H.; Brickey, W.J.; Bergmeier, W.; Dinauer, M.C.; Doerschuk, C.M. Mechanisms of Interferon-γ Production by Neutrophils and Its Function during Streptococcus pneumoniae Pneumonia. Am. J. Respir. Cell Mol. Boil. 2015, 52, 349–364. [Google Scholar] [CrossRef]
- Marques, J.M.; Rial, A.; Munoz, N.; Pellay, F.X.; Van Maele, L.; Leger, H.; Camou, T.; Sirard, J.C.; Benecke, A.; Chabalgoity, J.A. Protection against Streptococcus pneumoniae serotype 1 acute infection shows a signature of Th17- and IFN-gamma-mediated immunity. Immunobiology 2012, 217, 420–429. [Google Scholar] [CrossRef]
- Eichinger, K.; Egaña, L.; Orend, J.G.; Resetar, E.; Anderson, K.B.; Patel, R.; Empey, K.M. Alveolar macrophages support interferon gamma-mediated viral clearance in RSV-infected neonatal mice. Respir. Res. 2015, 16, 122. [Google Scholar] [CrossRef]
- Eichinger, K.; Kosanovich, J.L.; Empey, K.M. Localization of the T-cell response to RSV infection is altered in infant mice. Pediatr. Pulmonol. 2017, 53, 145–153. [Google Scholar] [CrossRef]
- Remot, A.; Descamps, D.; Noordine, M.-L.; Boukadiri, A.; Mathieu, E.; Robert, V.; Riffault, S.; Lambrecht, B.; Langella, P.; Hammad, H.; et al. Bacteria isolated from lung modulate asthma susceptibility in mice. ISME J. 2017, 11, 1061–1074. [Google Scholar] [CrossRef]
- Cervantes-Ortiz, S.L.; Grandvaux, N.; Grandvaux, N. Respiratory Syncytial Virus and Cellular Stress Responses: Impact on Replication and Physiopathology. Viruses 2016, 8, 124. [Google Scholar] [CrossRef]
- Penaloza, H.; Nieto, P.A.; Muñoz-Durango, N.; Salazar-Echegarai, F.J.; Torres, J.; Parga, M.J.; Alvarez-Lobos, M.; Riedel, C.A.; Kalergis, A.M.; Bueno, S.M. Interleukin-10 plays a key role in the modulation of neutrophils recruitment and lung inflammation during infection by Streptococcus pneumoniae. Immunology 2015, 146, 100–112. [Google Scholar] [CrossRef]
- Peñaloza, H.; Nieto, P.; Salazar, F.; Kalergis, A.; Bueno, S. Production of IL-10 during a streptococcus pneumoniae infection prevents excessive inflammation in lungs (MPF1P.778). J. Immunol. 2014, 192, 66. [Google Scholar]
- Sun, L.; Cornell, T.T.; Levine, A.; Berlin, A.A.; Hinkovska-Galcheva, V.; Fleszar, A.J.; Lukacs, N.W.; Shanley, T.P. Dual role of interleukin-10 in the regulation of respiratory syncitial virus (RSV)-induced lung inflammation. Clin. Exp. Immunol. 2013, 172, 263–279. [Google Scholar] [CrossRef]
- Van Der Sluijs, K.F.; Van Elden, L.J.R.; Nijhuis, M.; Schuurman, R.; Pater, J.M.; Florquin, S.; Goldman, M.; Jansen, H.M.; Lutter, R.; Van Der Poll, T. IL-10 is an important mediator of the enhanced susceptibility to pneumococcal pneumonia after influenza infection. J. Immunol. 2004, 172, 7603–7609. [Google Scholar] [CrossRef]
- Bosch, A.A.; Levin, E.; Van Houten, M.A.; Hasrat, R.; Kalkman, G.; Biesbroek, G.; Piters, W.D.S.; De Groot, P.-K.C.; Pernet, P.; Keijser, B.J.; et al. Development of Upper Respiratory Tract Microbiota in Infancy is Affected by Mode of Delivery. EBioMedicine 2016, 9, 336–345. [Google Scholar] [CrossRef]
- Teo, S.M.; Mok, D.; Pham, K.; Kusel, M.; Serralha, M.; Troy, N.; Holt, B.J.; Hales, B.J.; Walker, M.L.; Hollams, E.; et al. The infant nasopharyngeal microbiome impacts severity of lower respiratory infection and risk of asthma development. Cell Host Microbe 2015, 17, 704–715. [Google Scholar] [CrossRef]
- Dyer, K.D.; Drummond, R.; Rice, T.A.; Percopo, C.M.; Brenner, T.; Barisas, D.A.G.; Karpe, K.A.; Moore, M.L.; Rosenberg, H.F. Priming of the Respiratory Tract with Immunobiotic Lactobacillus plantarum Limits Infection of Alveolar Macrophages with Recombinant Pneumonia Virus of Mice (rK2-PVM). J. Virol. 2015, 90, 979–991. [Google Scholar] [CrossRef] [PubMed]
- LeMessurier, K.; Häcker, H.; Chi, L.; Tuomanen, E.I.; Redecke, V. Type I Interferon Protects against Pneumococcal Invasive Disease by Inhibiting Bacterial Transmigration across the Lung. PLOS Pathog. 2013, 9, e1003727. [Google Scholar] [CrossRef] [PubMed]
- Lécuyer, H.; Audibert, J.; Bobigny, A.; Eckert, C.; Jannière-Nartey, C.; Buu-Hoï, A.; Mainardi, J.-L.; Podglajen, I. Dolosigranulum pigrum Causing Nosocomial Pneumonia and Septicemia. J. Clin. Microbiol. 2007, 45, 3474–3475. [Google Scholar] [CrossRef] [PubMed]
- Hoedemaekers, A.; Schulin, T.; Tonk, B.; Melchers, W.J.; Sturm, P.D.J. Ventilator-Associated Pneumonia Caused by Dolosigranulum pigrum. J. Clin. Microbiol. 2006, 44, 3461–3462. [Google Scholar] [CrossRef]
- Bittar, F.; Richet, H.; Dubus, J.-C.; Reynaud-Gaubert, M.; Stremler, N.; Sarles, J.; Raoult, D.; Rolain, J.-M. Molecular Detection of Multiple Emerging Pathogens in Sputa from Cystic Fibrosis Patients. PLoS ONE 2008, 3, e2908. [Google Scholar] [CrossRef]
- Kolling, Y.; Salva, S.; Villena, J.; Marranzino, G.; Álvarez, S. Non-viable immunobiotic Lactobacillus rhamnosus CRL1505 and its peptidoglycan improve systemic and respiratory innate immune response during recovery of immunocompromised-malnourished mice. Int. Immunopharmacol. 2015, 25, 474–484. [Google Scholar] [CrossRef]












© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortiz Moyano, R.; Raya Tonetti, F.; Tomokiyo, M.; Kanmani, P.; Vizoso-Pinto, M.G.; Kim, H.; Quilodrán-Vega, S.; Melnikov, V.; Alvarez, S.; Takahashi, H.; et al. The Ability of Respiratory Commensal Bacteria to Beneficially Modulate the Lung Innate Immune Response Is a Strain Dependent Characteristic. Microorganisms 2020, 8, 727. https://doi.org/10.3390/microorganisms8050727
Ortiz Moyano R, Raya Tonetti F, Tomokiyo M, Kanmani P, Vizoso-Pinto MG, Kim H, Quilodrán-Vega S, Melnikov V, Alvarez S, Takahashi H, et al. The Ability of Respiratory Commensal Bacteria to Beneficially Modulate the Lung Innate Immune Response Is a Strain Dependent Characteristic. Microorganisms. 2020; 8(5):727. https://doi.org/10.3390/microorganisms8050727
Chicago/Turabian StyleOrtiz Moyano, Ramiro, Fernanda Raya Tonetti, Mikado Tomokiyo, Paulraj Kanmani, María Guadalupe Vizoso-Pinto, Hojun Kim, Sandra Quilodrán-Vega, Vyacheslav Melnikov, Susana Alvarez, Hideki Takahashi, and et al. 2020. "The Ability of Respiratory Commensal Bacteria to Beneficially Modulate the Lung Innate Immune Response Is a Strain Dependent Characteristic" Microorganisms 8, no. 5: 727. https://doi.org/10.3390/microorganisms8050727
APA StyleOrtiz Moyano, R., Raya Tonetti, F., Tomokiyo, M., Kanmani, P., Vizoso-Pinto, M. G., Kim, H., Quilodrán-Vega, S., Melnikov, V., Alvarez, S., Takahashi, H., Kurata, S., Kitazawa, H., & Villena, J. (2020). The Ability of Respiratory Commensal Bacteria to Beneficially Modulate the Lung Innate Immune Response Is a Strain Dependent Characteristic. Microorganisms, 8(5), 727. https://doi.org/10.3390/microorganisms8050727