Abstract
Pectobacterium and Dickeya species, usually referred to as soft rot Enterobacteriaceae, are phytopathogenic genera of bacteria that cause soft rot and blackleg diseases and are responsible for significant yield losses in many crops across the globe. Diagnosis of soft rot disease is difficult through visual disease symptoms. Pathogen detection and identification methods based on cultural and morphological identification are time-consuming and not always reliable. A polymerase chain reaction (PCR)-based detection method with the species-specific primers is fast and reliable for detecting soft rot pathogens. We have developed a specific and sensitive detection system for some species of soft rot Pectobacteriaceae pathogens in the Pectobacterium and Dickeya genera based on the use of species-specific primers to amplify unique genomic segments. The specificities of primers were verified by PCR analysis of genomic DNA from 14 strains of Pectobacterium, 8 strains of Dickeya, and 6 strains of non-soft rot bacteria. This PCR assay provides a quick, simple, powerful, and reliable method for detection of soft rot bacteria.
1. Introduction
Bacterial soft rot is a destructive disease in plants, notably vegetables, fruits, and ornamentals, and is caused by a group of Gram-negative bacteria collectively referred to as soft rot Enterobacteriaceae. Recently, the Proteobacterial family of Enterobacteriaceae was subdivided in to six new families, in which the soft rot pathogens were put into the new family, Pectobacteriaceae [1]. Thus, we are revising the name soft rot Enterobacteriaceae into soft rot Pectobacteriaceae (SRP). The soft rot disease occurs worldwide, mostly on fresh succulent plant tissues, and can occur in the field, in transit, in storage, and retail stores. The SRP contains two genera, Pectobacterium and Dickeya, which are globally widespread and cause high economic losses in ornamental plants, fruits, and vegetable productions [1,2,3,4,5,6,7]. Almost all vegetables are susceptible to soft rot disease, which causes a greater total loss than any other bacterial disease [2,78,9].
Through a series of taxonomic revisions starting in 1998, the new family Pectobacteriaceae now comprises Brenneria, Dickeya, Lonsdalea, Pectobacterium, and Sodalis genera [8,9,10,11,12]. Recently, several new Pectobacterium species have been reported, such as P. polaris, P. peruviense, P. punjabense, and P. versatile. As a result, the genus Pectobacterium is currently divided into 18 recognized species, including P. carotovorum [13,14,15,16,17,18,19,20,21,22]. Bacterial strains in the Dickeya genus are also divided into nine species, with the recent addition of two new species—Dickeya lacustris sp. nov. and Dickeya undicola sp. nov. [11,23,24,25,26,27,28,29].
Despite the diversity in SRP, a ubiquitous and key characteristic of SRP is their ability to produce copious amounts of plant cell wall-degrading enzymes (PCWDE), a characteristic which differentiates them from other members of the new Pectobacteriaceae or previous Enterobacteriaceae families. As a result of the devastation they cause and the research attention they have received, the SRP have been included among the top ten important plant pathogenic bacteria based on scientific or economic importance [3]. Among SRP, P. atrosepticum, P. carotovorum, P. brasiliense, and D. dadantii are the major pathogens causing important diseases such as aerial stem rot, blackleg, and soft rot of the potato, and many crop plants worldwide in the field and in storage [6,30,31,32,33,34,35,36,37,38]. Soft rot disease is difficult to manage, and available management strategies are not enough to reduce the effect of the disease [39]. Once the bacteria infect plants, there is no way to control this disease effectively, so accurate early pathogen detection is important for continuous SRP pathogen monitoring and prevention.
Although soft rot pathogens have received considerable research attention compared to other bacterial plant pathogens, there is still room for improvement in effective detection and diagnosis of these pathogens [4,12,40,41,42,43]. Traditional techniques of detection and identification of SRP, techniques based on isolation, bioassays, straining, microscopical observation, pathogenicity test, and biochemical methods are cumbersome [12]. These methods have two main drawbacks. First, they are used mostly to detect in vitro culturable organisms. Secondly, based on biochemical characteristics, some bacterial pathogen isolates have patterns which do not fit as a characteristic of any known genus and species. While it is relatively easy to identify bacterial soft rot disease symptomatically, separating the pathogens between the two genera can sometimes be tricky [12]. It is possible to have mixed infections of both Pectobacterium and Dickeya causing disease on a single host [12,34,44,45]. Over the last decade, we have seen the introduction of modern DNA-based approaches for the diagnosis and detection of pathogens [12,44]. Among them, PCR–based detection systems using specific primers are more reliable for detecting them.
Various target genes, including pmrA, pelADE, pel genes, pelY, pelI, cfa6, rhsA, recA, and 16S rDNA, have been used until now to identify Pectobacterium and Dickeya species by PCR assays [12,46,47,48,49,50,51]. Detection systems based on these targets have suffered from one major drawback. Primers developed from these genes are mostly specific for either one or two isolates of Pectobacterium and Dickeya. As a result, these systems have challenges detecting all strains of soft rot or separating them into genera. Based on bioinformatics approaches, conserved signature proteins (CSPs) have been identified for the two major genera of SRP [45]. Among these CSPs, two are uniquely present in the Dickeya species, three in the Pectobacterium species, and two are present in both but unique to Pectobacteriaceae. Due to the conservation of these proteins, detection systems developed based on either the proteins or the genes specifying them will be specific for detecting all soft rot strains.
The main purpose of this study was to develop a PCR-based detection tool that can serve as a rapid identification technique to detect some species of SRP in the genera of Pectobacterium and Dickeya. Specifically, we have designed a set of PCR primers that can be used to detect strains of SRP as a group and also separate them into Pectobacterium and Dickeya. These tools should make easier the detection of soft rot pathogens and the distinction between the two genera.
2. Materials and Methods
2.1. Bacterial Strains, Media, and Growth Conditions
Bacterial strains used in this study and their relevant characteristics are shown in Table S1. Strains were obtained from our laboratory culture collection, including the collection of Arun Chatterjee. Bacteria were grown at 28 °C or 37 °C in Luria broth (LB), nutrient yeast agar (NY), or King’s B (KB) media. The components and preparation of all media have been previously described [52].
2.2. Genomic DNA Extraction
Genomic DNA was extracted from overnight broth cultures in LB, NY, or KB media using the Promega Wizard Genomic DNA Purification kit (Promega, Madison, WI, USA). Rehydrated and purified DNA was stored in TE buffer at −20 °C.
2.3. Primer Design and Optimization
Primers for this study were designed from conserved regions using the sequences of the genes for proteins listed in Table S2. These proteins have been identified previously to exist in Pectobacterium alone, Dickeya alone, or both [45,52]. Based on the above information, these CSPs were used to design primers for the detection of both Pectobacterium and Dickeya species separately and together. Primers were picked from conserved segments of CSP-coding sequences in a series of steps. First, each CSP was blasted against the NR (nonredundant) protein database at NCBI. Only CSPs whose coding sequences hit SRP, Pectobacterium, or Dickeya alone were retained and used for further development. The coding sequence of each CSP was blasted against SRP genomes, and returning sequences were aligned in a multiple sequence alignment. Primers were then hand-picked in conserved regions and analyzed for properties and compatibility with the Oligo Analyzer tool (https://www.idtdna.com/analyzer/Applications/OligoAnalyzer/). Each primer pair (Table 1) was tested by PCR using the Eppendorf Mastercycler Nexus X1.

Table 1.
List of oligonucleotide primers used for conventional PCR amplification.
2.4. End Point PCR Conditions
Test PCR using P. carotovorum Ecc71 and Dickeya dadantii 3937 genomic DNA as the template was performed with gradient annealing temperature to determine the optimum annealing temperature for each primer pair. A 25 µL (microliter) PCR reaction contained: 1 µL (100 ng) DNA, 0.25 µL (1.25 units) Taq DNA polymerase (Thermo Fisher Scientific, Waltham, MA, USA), 2.5 µL of 10 × PCR buffer, 0.25 µL of 20 mM dNTPS, 0.25 µL each of 100 pmol each of forward and reverse primer, and 21.5 µL of sterile distilled water. Gradient PCR conditions were as follows: initial denaturation of 94 °C for 5 min (min), followed by 45 cycles at 94 °C for 1 min, 45−62 °C or 45−57 °C for 1 min, 72 °C for 1 min, and a final extension step of 72 °C for 5 min. Primers were tested with different genomic DNA listed in Table 1. Among the seven tested primer pairs, we selected the best three pairs for this study (SR1F-SR1R1 primer pair for SRP, Pcc3F-Pcc3R for Pectobacterium, and Dda1F-Dda1R for Dickeya species), and their optimized PCR protocol was run in a 15 µL reaction volume. The conditions for PCR were as follows: initial denaturation of 94 °C for 5 min, followed by 45 cycles of second denaturation 94 °C for 1 min, annealing (44.9 °C for SR1F-SR1R1, 49.9 °C for Pcc3F-Pcc3R, and 56.6 °C for Dda1F-Dda1R) for 1 min, extension 72 °C for 1 min, and a one-time final extension step of 72 °C for 5 min. The products of PCR (5 µL) were separated with 2% agarose in Tris-Boric EDTA (TBE; 0.5x) buffer and stained with ethidium bromide.
2.5. DNA Quantification and Plotting of Standard Curves by qPCR
Bacterial genomic DNA (gDNA) concentrations were measured using a Synergy H1 hybrid spectrophotometer microplate reader (Biotek, Winooski VT, USA). For standard curve and sensitivity testing, 100 ng µL−1 gDNA was 10–fold serially diluted with concentrations ranging from 100 ng µL−1 to 1 pg µL−1. These diluted samples were used as a template for quantitative real-time PCR (qPCR) using SYBR green. The Pectobacterium-specific primer set (Pcc3F–Pcc3R) and Dickeya-specific primer set (Dda1F–Dda1R) were used to run qPCR to test the sensitivity of detection with serial dilutions of Ecc71 and Dd3937 gDNA. Each dilution had three technical replications in each qPCR reaction, and every experiment was repeated two times. Standard curves were used to estimate the detection limits of each primer set and to quantify target soft rot bacterial gDNA in the samples. The detection threshold was calculated automatically using StepOne software (Applied Biosystems, Foster City, CA, USA).
The 25 µL qPCR reaction contained 11.25 µL SYBR green with real master mix (Applied Biosystems, Foster City, CA, USA), 0.125 µL each of forward and reverse primers (100 µm), 12.5 µL of sterile distilled water, and 1 µL of template gDNA. The qPCR reaction was performed with the StepOnePlus Real-Time PCR systems using MicroAmp fast optical 96-well reaction plates. The reaction conditions were as follows: initial denaturation at 94 °C for 5 min, followed by 40 cycles of denaturation at 94 °C for 1 min, annealing at 49.9 °C (for Pcc3F–Pcc3R) or 56.6 °C (for Dda1F–Dda1R) for 1 min, and extension at 72 °C for 1 min. It was considered a positive for the presence of the target DNA when reactions with threshold cycle (CT) values are ≤ 35.
2.6. Phylogenetic Analysis
The sequence of global regulatory protein RsmC (Dd586-0685 global regulatory protein) was used in a blast search against the whole NR (nonredundant) GenBank protein database. The sequences of Pectobacteriaceae (including non-soft rot genera) proteins with significant similarity were obtained from the GenBank database. These sequences were aligned with the Clustal W (Clustal Omega) program using default parameters [53], and the output file was formatted using MEGA 6 software. Phylogenetic trees for the dataset were constructed using the neighbor-joining program with MEGA 6 software [54,55]. Based on 500 resamplings, the stability of the relationships was evaluated by performing bootstrap analyses of the neighbor-joining data.
3. Results
3.1. Polymerase Chain Reaction Assay
The main purpose of this study was to develop a PCR-based system that can serve as a rapid identification technique to detect soft rot Pectobacteriaceae, Pectobacterium spp., and Dickeya spp. Twenty-eight bacterial strains (Table S1) belonging to some species of Pectobacterium and Dickeya genera, Enterobacteriaceae, and non-enterobacteria were used in this study. Of the 28, 14 were Pectobacterium strains, eight were Dickeya strains, and six were non-soft rot bacteria—of which, four were Enterobacteriaceae strains, and the remaining two were non-Enterobacteriaceae strains. Due to the recent extensive revision of taxonomy and nomenclature of this group, we were not immediately able to assign these strains to species levels based on the latest taxonomy. Therefore, we grouped the strains simply at a generic level.
When the RsmC protein (Dd586-0685 homologue) was first described as a global regulatory protein, it was also reported to be present only in soft rot bacteria using Southern blot analysis with the genomic DNA of soft rot bacteria and PCR with the rsmC primers on diverse strains from Pectobacteriaceae and Enterobacteriaceae [52]. These findings suggested that rsmC is both conserved and specific to soft rot Pectobacteriaceae. Interestingly, many years and sequenced bacterial genomes later, this protein remains essentially Pectobacteriaceae-specific. Amplification with soft rot-specific primers (SR1F-SR1R1), which were designed for the rsmC sequence, yielded a product with an expected size around 299 bp from all soft rot Pectobacteriaceae strains comprising both Pectobacterium and Dickeya strains. All non-SRPs which served as negative controls did not show amplification of this specific band (Figure 1).
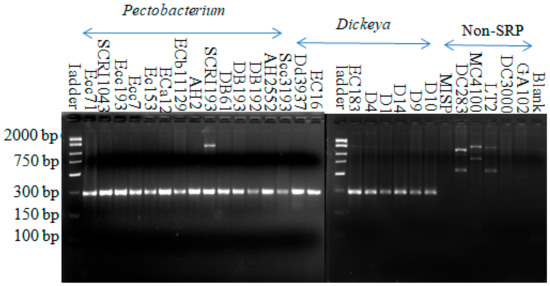
Figure 1.
Amplification from all Pectobacterium and Dickeya strains with the soft rot-specific primer (SR1F-SR1R1). The names of the strains are indicated above each lane. The product size is around the expected value of 299 bp. Fifteen-microliter reactions were carried out, followed by 45 cycles of annealing at 44.9 °C for 1 min and an extension of 72 °C for 1 min.
Using the Pectobacterium-specific primer set (Pcc3F-Pcc3R), amplification from all 14 Pectobacterium strains yielded a 177-bp DNA fragment in PCR. Using this primer set, all Dickeya species, non-soft rot Enterobacteria, and non-Enterobacterial strains did not amplify this specific fragment (Figure 2). Although faint bands were produced with Dickeya dadantii D14, Escherichia coli MC4100, and Salmonella LT2, none of these were in the same size range. This indicates that this primer set is specific for Pectobacterium species.
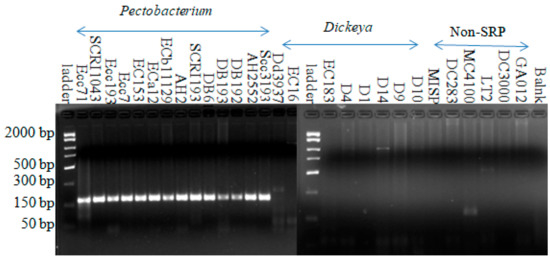
Figure 2.
Amplification with Pectobacterium-specific primer (Pcc3F-Pcc3R) from all Pectobacterium strains yielded a product with the expected size around 177 bp. Fifteen microliter reactions were carried out in 45 cycles with annealing at 49.9 °C for 1 min and an extension at 72 °C for 1 min.
All Dickeya strains yielded an expected 157-bp DNA fragment in PCR using the Dda1F–Dda1R primer set (Figure S1). Using this same primer set, seven out of fourteen Pectobacterium strains used in this study generated products in the size range of 220 bp, one of these being a very faint band on the gel. The remaining seven Pectobacterium strains, four non-soft rot Pectobacteriaceae, and two non-Enterobacteriaceae strains did not yield amplified fragments (Figure S1). To clearly show that it is possible to distinguish between Pectobacterium and Dickeya strains using this primer set, another PCR reaction was run with Dda1F-Dda1R primer using six Pectobacterium strains that produced 220-bp bright bands in the previous reaction (Figure S1). These six Pectobacterium strains were SCRI1043, Eca12, Ecc193, AH2, SCRI193, and AH2552. All other strains were same as in Figure S1. These six Pectobacterium strains yielded a product in the range size of 220-bp bands that were clearly distinguishable from the 157-bp product obtained from Dickeya strains (Figure 3). There were also slight differences in the sizes of the amplified products with this primer set, although the signal intensity appeared similar in all Dickeya strains. We speculate that there are slight differences in this locus among the Dickeya strains we tested. A summary of the PCR amplification results using the primers designed in this study is shown in Table 2.
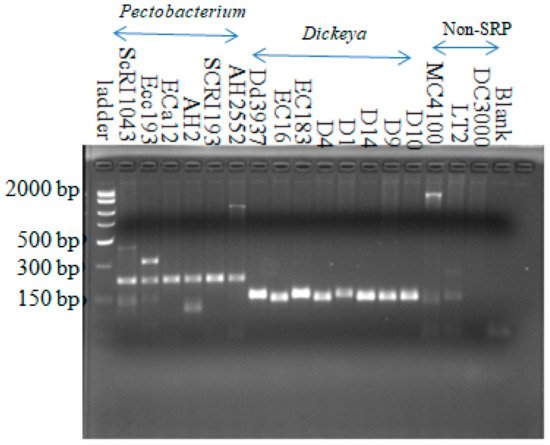
Figure 3.
Amplification from all Dickeya strains with the Dickeya-specific primer set (Dda1F-Dda1R). The product size is around the expected size of 157 bp. Six Pectobacterium strains generated about 220-bp bands. Three others, two Enterobacteriaceae (MC4100 and LT2), and Pseudomonas syringae (DC3000) strains did not show the 157-bp bands. Fifteen-microliter reactions were carried out in 45 cycles with annealing at 56.6 °C for 1 min and an extension at 72 °C for 1 min.
Table 2.
PCR amplification with primers developed in this study.
3.2. Sensitivity of the Polymerase Chain Reaction Assay
We also used these primers in qPCR to test the sensitivity of the primers to detect these bacteria. qPCR was performed using serial dilutions of Pectobacterium (Ecc71) and Dickeya (Dd3937) gDNA ranging in concentration from 100 ng to 1 pg per reaction. Pectobacterium-specific (Pcc3F-Pcc3R) and Dickeya-specific (Dda1F-Dda1R) primer sets were used for qPCR analysis of their respective genomic DNAs. The threshold cycle (CT) was 24.84 cycles for gDNA (1 pg/µL) of Ecc71 by Pectobacterium-specific primers (Figure S2A) and 27.29 cycles for gDNA (1 pg/µL) of Dd3937 by Dickeya-specific primers (Figure S2B). Both Pcc3F-Pcc3R and Dda1F-Dda1R primer sets could be used to detect up to 1 pg/µL DNA. These results indicate that both primer sets can be used to specifically detect SRP pathogens with very low inoculum levels.
3.3. Phylogenetic Analysis Based on Regulatory Protein, RsmC
Since rsmC remained essentially Pectobacteriaceae-specific and its sequence could be used to distinguish these bacteria from others, we wanted to determine if we could use it for the phylogenetic analysis of both soft rot and non-soft rot Pectobacteriaceae as well. For that, we constructed a phylogenetic tree of Pectobacteriaceae strains based on RsmC (Dd586-0685) protein sequences. The sequences were obtained from the NCBI database using a blast search against the whole NR (nonredundant) GenBank database based on the sequence of global regulatory protein RsmC (Dd586-0685). Based on the analysis, the four taxa (Pectobacterium, Brenneria, Dickeya, and Lonsdalea) were clearly divided into four clades (I -IV). Clade I groups together all the proteins from the Pectobacterium species, while the third clade comprises sequences from the Dickeya species. We also noted that there was slightly more diversity among the Dickeya species than among the Pectobacterium species (Figure 4).
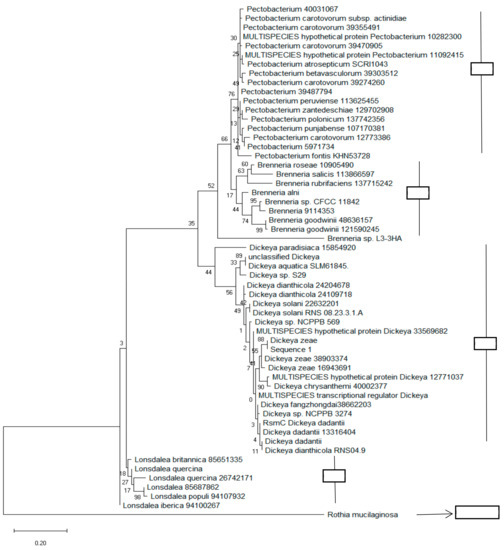
Figure 4.
Phylogenetic tree constructed based on the rsmC sequence. The branching pattern was constructed using the neighbor-joining method [54]. The numbers which are present at the nodes representing the levels of bootstrap support based on a neighbor-joining study of a set of 500 resampled data. The evolutionary distances were calculated by the maximum composite likelihood method [55]. It also calculates the base substitutions per site. The whole phylogenetic tree was generated using MEGA 6 software [39].
4. Discussion
Traditional techniques, including microbiological, serological, and biochemical methods for the detection and identification of plant pathogenic bacteria, are time-consuming and, most of the time, do not have enough sensitivity and specificity [12,56]. As a result, these methods are not specifically suited for the routine analysis of many samples [56]. At times, there are also problems of low reproducibility of detection by phenotypic characteristics, lack of phylogenetic meaning, and artificial negative results due to injured bacteria [57]. As high-throughput DNA sequencing becomes common and costs reduce, pathogen detection and diagnostic techniques are slowly progressing from these traditional methods to molecular methods [58,59]. This change is expected to continue into the future as more technologies are developed [60,61]. The overall aims of all these techniques have remained the same—accurate, reliable, and fast identification and differentiation of specific bacteria from others and of the disease symptoms they cause from those by other causes. Nucleic acids-based technologies, such as conventional PCR with its variants and qPCR, have some advantages over traditional methods. Here, in vitro culture of the pathogens is not necessary for their identification [12,56]. Additionally, PCR-based methods are reliable, cost-effective, specific, sensitive, and rapid to detect pathogens from environmental samples. Detection techniques for plant pathogenic bacteria have greatly benefited from these modern technological advancements, which is needed to identify and monitor plant diseases [56]. However, these approaches have some limitations, such as difficulty in distinguishing between viable and nonviable bacterial cells [62], possible false-positives with nontargets, and a limitation of detection by the presence of inhibitors in host extracts for PCR reactions [63]. These downsides are systematically overcome with progressive improvements in the techniques.
Three different pairs of primers were developed and used in this study, one for Pectobacterium spp. (Pcc3F-Pcc3R), one for Dickeya spp. (Dda1F-Dda1R), and the third one (SR1F-SR1R1) for soft rot bacteria comprising both Pectobacterium and Dickeya species. The Pectobacterium-specific primer set could detect all tested Pectobacterium strains, and Dickeya-specific primers could also detect all strains of Dickeya species that we tested. The global regulatory gene rsmC, on which the soft rot-specific primer was based, was first described in 1999 as a regulator of virulence and extracellular enzyme production in Pectobacterium carotovorum [52]. At the time, the authors observed through Southern blot hybridization analysis and PCR that this gene was only present in SRP. Probably because of the number of strains tested and lack of the complete genome sequence data at the time, the utility of the rsmC sequence as a molecular diagnostic tool was not immediately considered. It is therefore not surprising that the genomic analysis work of Naushad et al. [45] also identified rsmC as a soft rot-specific gene [52]. The most recent database search also returned proteins from members of Pectobacteriaceae. There were significant scores with proteins from Serratia spp. and Samsonia erythrinae, both members of the new family of Yersiniaceae. To our knowledge, this is the first primer set yet developed that can detect Pectobacterium and Dickeya species together and separately.
We showed in this study that the rsmC sequence could be used to separate strains of Pectobacterium, Dickeya, Brenneria, and Lonsdalea phylogenetically. Despite the divergence of rsmC sequences that allows it to be used for a phylogenetic analysis of the Pectobacteriaceae family, our diagnostic system could still correctly detect both soft rot genera, because the primers were specially designed for regions of the rsmC gene that are conserved in both genera. The primers we designed here are a first and an important step in the analysis of some Pectobacterium and Dickeya species. As our developed primers could detect several Pectobacterium and Dickeya alone and together, so this genome-based approach in developing a detection system can be a useful tool in research work to develop an effective detection system of other pathogen groups.
Each of the two genera, Pectobacterium and Dickeya, is comprised of twenty-seven species [7,8,13,64,65,66]. Some of these species in both genera are new and have only existed for a short time, following the period of heavy revision in taxonomy and phylogeny of soft rot bacteria. As a result, we were unable to test our primer set with strains belonging to all presently known species of both genera. The collection of strains belonging to all species of Pectobacterium and Dickeya across different phytosanitary jurisdictions presented a logistical hurdle that we could not overcome within the period of this project. The above shortfall notwithstanding, we anticipate that the genome-directed, PCR-based molecular diagnostic system we described here will be useful for monitoring the presence of soft rot bacteria under various conditions, whether in the field or in storage or in seed certification programs for blackleg/soft rot-free planting materials. We hope that this method will prove valuable in the routine detection of soft rot bacteria for environmental studies. We anticipate that the system will be further improved by others in an ongoing effort to improve pathogen detection and diagnosis. For example, it should be possible to develop another equally reliable detection system using the conservation of the RsmC protein sequence based on immunogenic properties.
5. Conclusions
In conclusion, we have developed a PCR-based detection technique which will be very helpful for detecting SRP pathogens with specificity and sensitivity. This detection method can also be valuable in developing an efficient monitoring and management system to manage soft rot disease.
Supplementary Materials
Supplementary materials can be found at https://www.mdpi.com/2076-2607/8/3/358/s1. Table S1. Bacterial Strains. Table S2. Proteins whose coding sequences were used to design primers. Figure S1. Amplification with Dickeya specific primer (Dda1F-Dda1R). All Dickeya strains yielded a product with the expected size of 157-bp. Seven Pectobacterium strains yielded products in the range size of 220-bp DNA fragment. Other seven Pectobacterium strains, two Entrobacteriaceae, two Erwiniaceae, and two non Enterobacteriales strains did not yield any fragment. Fifteen µl reactions were carried out in 45 cycles with annealing at 56.6 °C for 1 min and extension at 72 °C for 1 min. Figure S2A. Standard curves showing real-time PCR assay CT values vs. template DNA concentrations from Ecc71 with Pcc3F-Pcc3R primer set. Here Y = threshold cycles (CT) of target DNA detected, x = amount of target DNA (pg) used as a template to generate each data point in the standard curve. Twenty five µl reactions were carried out in 40 cycles with annealing at 49.9 °C for 1 min and extension at 72 °C for 1 min. Red color represents target samples and green color represents positive control. The R2 for the plot is 0.89. Figure S2B. Standard curves showing real-time PCR assays. CT values vs template DNA concentrations from Dd3937 with Dda1F-Dda1R primer set. Here Y-axis represents threshold cycles (CT) of target DNA detected and the, X-axis represents amount of target DNA (pg) used as a template to generate each data point in the standard curve. Twenty-five µl reactions were carried out in 40 cycles with annealing at 56.6 °C for 1 min and extension at 72 °C for 1 min. Blue color represents positive control and red color represents positive control. The R2 for the plot is 0.99.
Author Contributions
Conceptualization, C.K.D.; methodology, M.N.K., A.T. and C.K.D.; validation, M.N.K., and C.K.D.; resources, C.K.D.; writing—original draft preparation, M.N.K.; writing—review and editing, M.N.K., A.T. and C.K.D.; supervision, C.K.D.; project administration, C.K.D.; funding acquisition, C.K.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was publicly funded by National Institute of Food and Agriculture of USDA: TENX-1828-GF SH.
Acknowledgments
We thank A. Chatterjee for bacterial strains and Andrew Dickey for reviewing the manuscript. The first author was supported by graduate assistantships from the Department of Agricultural and Environmental Sciences.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Adeolu, M.; Alnajar, S.; Naushad, S.; Gupta, S.R. Genome-based phylogeny and taxonomy of the ‘Enterobacteriales’: Proposal for Enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam.nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5575–5599. [Google Scholar] [PubMed]
- Bhat, K.A.; Masood, S.D.; Bhat, N.A.; Bhat, M.A.; Razvi, S.M.; Mir, M.R.; Akhtar, S.; Wani, N.; Habib, M. Current Status of Post Harvest Soft Rot in Vegetables: A Review. Asian J. Plant Sci. 2010, 9, 200–208. [Google Scholar] [CrossRef]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum., M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [PubMed]
- Czajkowski, R.; Pérombelon, M.C.M.; van Veen, J.A.; van der Wolf, J.M. Control of blackleg and tuber soft rot of potato caused by Pectobacterium and Dickeya species: A review. Plant Pathol. 2011, 60, 999–1013. [Google Scholar] [CrossRef]
- Perombelon, M.C.M.; Kelman, A. Ecology of the soft rot Pectobacterium. Ann. Rev. Phytopathol. 1980, 18, 316–387. [Google Scholar] [CrossRef]
- Pérombelon, M.C.M. Potato diseases caused by soft rot Erwinias: An overview of pathogenesis. Plant Pathol. 2002, 51, 1–12. [Google Scholar] [CrossRef]
- Ma, B.; Hibbing, M.E.; Kim, H.S.; Reedy, R.M.; Yedidia, I.; Breuer, J.; Breuer, J.; Glasner, J.D.; Perna, N.T.; Kelman, A.; et al. Host range and molecular phylogenies of the soft rot enterobacterial genera Pectobacterium and Dickeya. Phytopathol 2007, 97, 1150–1163. [Google Scholar] [CrossRef]
- Charkowski, A.O.; Lind, J.; Rubio-salazar, I. Genomics of Plant-Associated Bacteria; Springer: Berlin/Heidelberg, Germany, 2014; pp. 37–59. [Google Scholar] [CrossRef]
- Tarasova, N.; Gorshkov, V.; Petrova, O.; Gogolev, Y. Potato signal molecules that activate pectate lyase synthesis in Pectobacterium atrosepticum SCRI1043. World J. Microbiol. Biotechnol. 2013, 29, 1189–1196. [Google Scholar] [CrossRef]
- Hauben, L.; Moore, E.R.; Vauterin, L.; Steenackers, M.; Mergaert, J.; Verdonck, L.; Swings, J. Phylogenetic position of phytopathogens within the Enterobacteriaceae. Syst. Appl. Microbiol. 1998, 21, 384–397. [Google Scholar] [CrossRef]
- Samson, R.; Legendre, J.B.; Christen, R.; Fischer-Le Saux, M.; Achouak, W.; Garden, L. Transfer of Pectobacterium chrysanthemi and Brenneria paradisiaca to the genus Dickeya gen. nov. as Dickeya chrysanthemi comb. nov. and Dickeya paradisiaca comb. nov. and delineation of four novel species, Dick. Int. J. Syst. Evol. Microbiol. 2005, 55, 1415–1427. [Google Scholar] [CrossRef]
- Czajkowski, R.; Pérombelon, M.C.M.; Jafra, S.; Lojkowska, E.; Potrykus, M.; van der Wolf, J.M.; Sledz, W. Detection, identification and differentiation of Pectobacterium and Dickeya species causing potato blackleg and tuber soft rot: A review. Ann. Appl. Biol. 2015, 166, 18–38. [Google Scholar] [CrossRef] [PubMed]
- Gardan, L.; Gouy, C.; Christen, R.; Samson, R. Elevation of three subspecies of Pectobacterium carotovorum to species level: Pectobacterium atrosepticum sp. nov., Pectobacterium betavasculorum sp. nov. and Pectobacterium wasabiae sp. nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Nabhan, S.; De Boer, S.H.; Maiss, E.; Wydra, K. Pectobacterium aroidearum sp. nov., a soft rot pathogen with preference for monocotyledonous plants. Int. J. Syst. Evol. Microbiol. 2013, 61, 498–508. [Google Scholar] [CrossRef] [PubMed]
- Khayi, S.; Cigna, J.; Chong, T.M.; Quetu-Laurent, A.; Chan, K.G.; Helias, V.; Faure, D. Transfer of the potato plant isolates of Pectobacterium wasabiae to Pectobacterium parmentieri sp. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5379–5383. [Google Scholar] [CrossRef] [PubMed]
- Waleron, M.; Waleron, K.; Lojkowska, E. Characterization of Pectobacterium carotovorum subsp odoriferum causing soft rot of stored vegetables. Eur. J. Plant Pathol. 2014, 139, 457–469. [Google Scholar]
- Zaczek-Moczydłowska, M.A.; Fleming, C.C.; Young, G.K.; Campbell, K.; O’Hanlon, R. Pectobacterium and Dickeya species detected in vegetables in Northern Ireland. Eur. J. Plant Pathol. 2019, 154, 635–647. [Google Scholar] [CrossRef]
- Dees, M.W.; Lysoe, E.; Rossmann, S.; Perminow, J.; Brurberg, M.B. Pectobacterium polaris sp. nov., isolated from potato (Solanum tuberosum). Int. J. Syst. Evol. Microbiol 2017, 67, 5222–5229. [Google Scholar] [CrossRef]
- Waleron, M.; Misztak, A.; Waleron, M.; Franczuk, M.; Wielgomas, B.; Waleron, K. Transfer of Pectobacterium carotovorum subsp. carotovorumstrains isolated from potatoes grown at high altitudes to Pectobacterium peruviense sp. nov. Syst. Appl. Microbiol. 2017, 41, 85–93. [Google Scholar] [CrossRef]
- Sarfraz, S.; Riaz, K.; Oulghazi, S.; Cigna, J.; Sahi, S.T.; Khan, S.H.; Faure, D. Pectobacterium punjabense sp. nov., isolated from blackleg symptoms of potato plants in Pakistan. Int. J. Syst. Evol. Microbiol. 2018, 68, 3551–3556. [Google Scholar] [CrossRef]
- Shirshikov, F.V.; Korzhenkov, A.A.; Miroshnikov, K.K.; Kabanova, A.P.; Barannik, A.P.; Ignatov, A.N. Draft genome sequences of new genomospecies “Candidatus Pectobacterium maceratum” strains, which cause soft rot in plants. Genome Announc. 2018, 6, 218–260. [Google Scholar] [CrossRef]
- Portier, P.; Pédron, J.; Taghouti, G.; Fischer-Le Saux, M.; Caullireau, E.; Bertrand, C.; Laurent, A.; Chawki, K.; Oulgazi, S.; Moumni, M.; et al. Elevation of Pectobacterium carotovorum subsp. odoriferum to species level as Pectobacterium odoriferum sp. nov., proposal of Pectobacterium brasiliense sp. nov. and Pectobacterium actinidiae sp. nov., emended description of Pectobacterium carotovorum and description of Pectobacterium versatile sp. nov., isolated from streams and symptoms on diverse plants. Int. J. Syst. Evol. Microbiol. 2019, 69, 3207–3216. [Google Scholar]
- Brady, C.L.; Cleenwerck, I.; Denman, S.; Venter, S.N.; Rodríguez-Palenzuela, P.; Coutinho, T.A.; De Vos, P. Proposal to reclassify Brenneria quercina (Hildebrand & Schroth 1967) Hauben 1999 into a novel genus, Lonsdalea gen. nov., as Lonsdalea quercina comb. nov., descriptions of Lonsdalea quercina subsp. quercina comb. nov., Lonsdalea quercina subsp. iberica subsp. nov. and Lonsdalea quercina subsp. britannica subsp. nov., emendation of the description of the genus Brenneria, reclassification of Dickeya dieffenbachiae as Dickeya dadantii subsp. dieffenbachiae comb. nov., and emendation of the description of Dickeya dadantii. Int. J. Syst. Evol. Microbiol. 2012, 62, 1592–1602. [Google Scholar] [PubMed]
- Parkinson, N.; DeVos, P.; Pirhonen, M.; Elphinstone, J. Dickeya aquatica sp. nov., isolated from waterways. Int. J. Syst. Evol. Microbiol. 2014, 64, 2264–2266. [Google Scholar] [CrossRef] [PubMed]
- van derWolf, J.M.; Nijhuis, E.H.; Kowalewska, M.J.; Saddler, G.S.; Parkinson, N.; Elphinstone, J.G.; Pritchard, L.; Toth, I.K.; Łojkowska, E.; Potrykus, M.; et al. Dickeya solani sp. nov., a pectinolytic plant pathogenic bacterium isolated from potato (Solanum tuberosum). Int. J. Syst. Evol. Microbiol. 2014, 64, 768–774. [Google Scholar] [CrossRef]
- Tian, Y.; Zhao, Y.; Yuan, X.; Yi, J.; Fan, J.; Xu, Z.; Hu, B.; De Boer, S.H.; Li, X. Dickeya fangzhongdai sp. nov., a plant-pathogenic bacterium isolated from pear trees (Pyrus pyrifolia). Int. J. Syst. Evol. Microbiol. 2016, 66, 2831–2835. [Google Scholar] [CrossRef]
- Duprey, A.; Taib, N.; Leonard, S.; Garin, T.; Flandrois, J.P.; Nasser, W.; Brochier-Armanet, C.; Reverchon, S. The phytopathogenic nature of Dickeya aquatica 174/2 and the dynamic early evolution of Dickeya pathogenicity. Environmen. Microbiol 2019, 21, 2809–2835. [Google Scholar] [CrossRef]
- Hugouvieux-Cotte-Pattat, N.; Jacot-des-Combes, C.; Briolay, J. Dickeya lacustris sp. nov., a water-living pectinolytic bacterium isolated from lakes in France. Int. J. Syst. Evol. Microbiol. 2019, 69, 721–726. [Google Scholar] [CrossRef]
- Oulghazi, S.; Pédron, J.; Cigna, J.; Lau, Y.Y.; Moumni, M.; Van Gijsegem, F.; Chan, K.G.; Faure, D. Dickeya undicola sp. nov., a novel species for pectinolytic isolates from surface waters in Europe and Asia. Int. J. Syst. Evol. Microbiol. 2019, 69, 2440–2444. [Google Scholar] [CrossRef]
- Nunes Leite, L.; de Haan, E.G.; Krijger, M.; Kastelein, P.; van der Zouwen, P.S.; van den Bovenkamp, G.W.; Tebaldi, N.D.; van der Wolf, J.M. First report of potato blackleg caused by Pectobacterium carotovorum subsp. brasiliensis in the Netherlands. New Dis. Rep. 2014, 29, 24. [Google Scholar] [CrossRef]
- De Werra, P.; Bussereau, F.; Keiser, A. First report of potato blackleg caused by Pectobacterium carotovorum subsp. brasiliense in Switzerland. Plant Disease. 2015, 99, 551. [Google Scholar] [CrossRef]
- Toth, I.K.; van der Wolf, J.M.; Saddler, G.; Lojkowska, E.; Hélias, V.; Pirhonen, M.; Tsror Lahkim, L.; Elphinstone, J.G. Dickeya species: An emerging problem for potato production in Europe. J. Plant Pathol. 2011, 60, 385–399. [Google Scholar] [CrossRef]
- Van der Wolf, J.M.; de Haan, E.G.; Kastelein, P.; Krijger, M.; de Haas, B.H.; Velvis, H.; Mendes, O.; Kooman-Gersmann, M.; van der Zouwen, P.S. Virulence of Pectobacterium carotovorum subsp. brasiliense on potato compared with that of other. Pectobacterium and Dickeya species under climatic conditions prevailing in the Netherlands. Plant Pathol. 2017, 66, 571–583. [Google Scholar] [CrossRef]
- De Boer, S.H.; Li, X.; Ward, L. Pectobacterium spp. associated with bacterial stem rot syndrome of potato in Canada. Phytopathol. 2012, 102, 937–947. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dees, M.W.; Lebecka, R.; Perminow, J.I.; Czajkowski, R.; Grupa, A.; Motyka, A.; Zoledowska, S.; Śliwka, J.; Łojkowska, E.; Brurberg, M.B. Characterization of Dickeya and Pectobacterium strains obtained from diseased potato plants in different climatic conditions of Norway and Poland. Eur. J. Plant Pathol. 148, 839–851. [CrossRef]
- Waleron, M.; Waleron, K.; Łojkowska, E. Occurrence of Pectobacterium wasabiae in potato field samples. Eur. J. Plant Pathol. 2013, 137, 149–158. [Google Scholar] [CrossRef][Green Version]
- Elphistone, J. Blackleg Survey-English and Welsh Seed Crops 2013−2015. Final Report. 2016. Available online: https://potatoes.ahdb.org.uk/sites/default/files/publication_upload/R475%20Final%20Report%20for%20publication.pdf (accessed on 10 February 2020).
- Elphistone, J. Blackleg Survey Samples. Final Report. 2016. Available online: http://potatoes.ahdb.org.uk/sites/default/files/publication_upload/2016%20monitoring%20stocks%20of%20a%20single%20variety.pdf (accessed on 10 February 2020).
- Potrykus, M.; Sledz, W.; Golanowska, M.; Slawiak, M.; Binek, A.; Motyka, A.; Zoledowska, S.; Czajkowski, R.; Lojkowska, E. Simultaneous detection of major blackleg and soft rot bacterial pathogens in potato by multiplex polymerase chain reaction. Ann. Appl. Biol. 2014, 165, 474–487. [Google Scholar] [CrossRef]
- Barras, F.; van Gijsegem, F.; Chatterjee, A.K. Extracellular Enzymes and Pathogenesis of Soft-Rot Erwinia. Annu. Rev. Phytopathol. 1994, 32, 201–234. [Google Scholar] [CrossRef]
- Hauben, L.; Steenackers, M.; Swings, J. PCR-Based detection of the causal agent of watermark disease in willows (Salix spp.). Appl. Environ. Microbiol. 1998, 64, 3966–3971. [Google Scholar] [CrossRef]
- De Boer, S.H. Characterization of pectolytic erwinias as highly sophisticated pathogens of plants. Eur. J. Plant Pathol. 2003, 109, 893–899. [Google Scholar] [CrossRef]
- Charkowski, A.; Blanco, C.; Condemine, G.; Expert, T.; Franza, T.; Hayes, C.; Hugouvieux-Cotte-Pattat, N.; López Solanilla, E.; Low, D.; Moleleki, L.; et al. The role of secretion systems and small molecules in soft-rot Enterobacteriaceae pathogenicity. Annu. Rev. Phytopathol. 2012, 50, 425–449. [Google Scholar] [CrossRef]
- Yasuhara-Bell, J.; Marrero, G.; De Silva, A.; Alvarez, A.M. Specific detection of Pectobacterium carotovorum by loop-mediated isothermal amplification. Mol. Plant Pathol. 2016, 17, 1499–1505. [Google Scholar] [CrossRef] [PubMed]
- Naushad, H.S.; Lee, B.; Gupta, R.S. Conserved signature indels and signature proteins as novel tools for understanding microbial phylogeny and systematics: Identification of molecular signatures that are specific for the phytopathogenic genera Dickeya, Pectobacterium and Brenneria. Int. J. Syst. Evol. Microbiol. 2014, 64, 366–383. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Nie, J.; Ward, L.J.; Nickerson, J.; De Boer, S.H. Development and evaluation of a loop-mediated isothermal amplification assay for rapid detection and identification of Pectobacterium atrosepticum. Can. J. Plant Pathol. 2011, 33, 447–457. [Google Scholar] [CrossRef]
- Diallo, S.; Latou, R.X.; Groboillot, A.; Smadja, B.; Copin, P.; Orange, N.; Feuilloley, M.G.J.; Chevalier, S. Simultaneous and selective detection of two major soft rot pathogens of potato: Pectobacterium atrosepticum (Erwinia carotovora subsp atrosepticum) and Dickeya spp. (Erwinia chrysanthemi). Eur. J. Plant Pathol. 2009, 125, 349–354. [Google Scholar] [CrossRef]
- Park, D.S.; Shim, J.K.; Kim, J.S.; Kim, B.Y.; Kang, M.J.; Seol, Y.J.; Hahn, J.H.; Sherstha, R.; Lim, C.K.; Go, S.J.; et al. PCR-based sensitive and specific detection of Pectobacterium atrosepticum using primers based on Rhs family gene sequences. Plant Pathol. 2006, 55, 625–629. [Google Scholar] [CrossRef]
- Frechon, D.; Exbrayat, P.; Helias, V.; Hyman, L.J.; Jouan, B.; Llop, P.; Lopez, M.M.; Payet, N.; Perombelon, M.C.M.; Toth, I.K.; et al. Evaluation of a PCR kit for the detection of Erwinia carotovora subsp. atroseptica on potato tubers. Potato Res. 1998, 41, 163–173. [Google Scholar] [CrossRef]
- Nassar, A.; Darrasse, A.; Lemattre, M.; Kotoujansky, A.; Dervin, C.; Vedel, R.; Bertheau, Y. Characterization of Erwinia chrysanthemi by pectinolytic isozyme polymorphism and restriction fragment length polymorphism analysis of PCR-amplified fragments of pel genes. Appl. Environ. Microbiol. 1996, 62, 2228–2235. [Google Scholar] [CrossRef]
- Darrasse, A.; Priou, S.; Kotoujansky, A.; Bertheau, Y. PCR and restriction fragment length polymorphism of a pel gene as a tool to identify Erwinia carotovora in relation to potato diseases. Appl. Environ. Microbiol. 1994, 60, 1437–1443. [Google Scholar] [CrossRef]
- Cui, Y.; Mukherjee, A.; Dumenyo, C.K.; Liu, Y.; Chatterjee, A.K. rsmC of the soft-rotting bacterium Erwinia carotovora subsp. carotovora negatively controls extracellular enzyme and harpin(Ecc) production and virulence by modulating levels of regulatory RNA (rsmB) and RNA-binding protein (RsmA). J. Bacteriol. 1999, 181, 6042–6052. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol Biol Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Awllace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [PubMed]
- De Boer, S.H.; Elphinstone, J.G.; Saddler, G.S. Molecular detection strategies for phytopathogenic bacteria. Biotechnol. Plant Dis. Manag. 2007, 165–194. [Google Scholar]
- Eberle, K.N.; Kiess, A.S. Phenotypic and genotypic methods for typing Campylobacter jejuni and Campylobacter coli in poultry. Poult. Sci. 2012, 91, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, A.M. Integrated approaches for detection of plant pathogenic bacteria and diagnosis of bacterial diseases. Annu. Rev. Phytopathol. 2004, 42, 339–366. [Google Scholar] [CrossRef] [PubMed]
- López, M.M.; Bertolini, E.; Olmos, A.; Caruso, P.; Gorris, M.T.; Llop, P.; Penyalver, R.; Cambra, M. Innovative tools for detection of plant pathogenic viruses and bacteria. Int. Microbiol. 2003, 6, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Aslam, S.; Tahir, A.; Aslam, M.F.; Alam, M.W.; Shedayi, A.A.; Sadia, S. Recent advances in molecular techniques for the identification of phytopathogenic fungi–a mini review. J. Plant Interact. 2017, 12, 493–504. [Google Scholar] [CrossRef]
- Scala, V.; Pucci, N.; Loreti, S. The diagnosis of plant pathogenic bacteria: A state of art. Front. Biosci. 2018, 10, 449–460. [Google Scholar] [CrossRef]
- Kang, H.W.; Kwon, S.W.; Go, S.J. PCR-based specific and sensitive detection of Pectobacterium carotovorum ssp. carotovorum by primers generated from a URP-PCR fingerprinting-derived polymorphic band. Plant Pathol. 2003, 52, 127–133. [Google Scholar] [CrossRef]
- Smit, M.L.; Giesendorf, B.J.; Vet, J.M.; Trijbels, F.J.M.; Blom, H.J. Semiautomated DNA mutation analysis using a robotic workstation and molecular beacons. Clin. Chem. 2001, 47, 739–744. [Google Scholar] [CrossRef]
- Duarte, V.; de Boer, S.H.; Ward, L.J.; de Oliveira, A.M. Characterization of atypical Erwinia carotovora strains causing blackleg of potato in Brazil. J. Appl. Microbiol. 2004, 96, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed]
- Kerremans, J.J.; Verboom, P.; Stiphen, T.; Hakkaart van Roijen, L.; Goessens, W.; Verburgh, H.A.; Vos, M.C. Rapid identification and antimicrobial susceptibility testing reduce antibiotic use accelerate pathogen-directed antibiotic use. J. Antimicrob. Chemother. 2008, 61, 428–435. [Google Scholar] [CrossRef] [PubMed]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).