Abstract
This study describes the first complete genomic sequence of an NDM-19 and QnrS11-producing multidrug-resistant (MDR) Escherichia coli isolate collected from a fecal swab from a poultry farm in 2019 in Egypt. The blaNDM-19 was identified by PCR screening and DNA sequencing. The isolate was then subjected to antimicrobial susceptibility testing, conjugation and transformation experiments, and complete genome sequencing. The chromosome of strain M2-13-1 measures 4,738,278 bp and encodes 4557 predicted genes, with an average G + C content of 50.8%. M2-13-1 is classified under ST167, serotype O101:H5, phylogroup A, and shows an MDR phenotype, having minimum inhibitory concentrations (MICs) of 64 mg/L for both meropenem and doripenem. The genes blaNDM-19 and qnrS11 are present on 49,816 bp IncX3 and 113,285 bp IncFII: IncFIB plasmids, respectively. M2-13-1 harbors genes that impart resistance to sulfonamides (sul1), trimethoprim (dfrA14), β-lactams (blaTEM-1B), aminoglycosides (aph(6)-Id, aph(3′)-Ia, aph(3″)-Ib, aac(3)-IV, and aph(4)-Ia), tetracycline (tet(A)), and chloramphenicol (floR). It was susceptible to aztreonam, colistin, fosfomycin, and tigecycline. The genetic context surrounding blaNDM-19 includes ISAba125-IS5-blaNDM-19-bleMBL-trpF-hp1-hp2-IS26. Hierarchical clustering of the core genome MLST (HierCC) indicated M2-13-1 clusters with global ST167 E. coli lineages, showing HC levels of 100 (HC100) core genome allelic differences. Plasmids of the IncX3 group and the insertion sequence (ISAba125) are critical vehicles for the dissemination of blaNDM and its related variants. To our knowledge, this is the first genomic report of a blaNDM-19/IncX3-carrying E. coli isolate of animal origin globally.
1. Introduction
Carbapenem antibiotics (e.g., imipenem, meropenem, doripenem, and ertapenem) are considered as last-resort treatments for infections caused by multidrug-resistant bacteria [1,2]. This class exhibits a broad spectrum of activity against bacteria and improved stability against β-lactamases [1,2]. In 2021, the WHO developed the Access, Watch, Reserve (AWaRe) classification of antibiotics to help reduce bacterial resistance and improve antimicrobial stewardship (https://www.who.int/publications/i/item/2021-aware-classification, accessed on 15 January 2024). It was noted that most of the carbapenems were on the WHO watch list, while imipenem/cilastatin/relebactam and meropenem/vaborbactam were on the WHO reserve list.
The spread and emergence of resistance to carbapenems is a serious public health concern. In Enterobacteriaceae, the resistance to carbapenem antibiotics is facilitated by the production of a great number of carbapenemases, which include (i) Ambler class A β-lactamases (KPC, FRI, GES, SME), (ii) Ambler class B metallo-β-lactamases (VIM, NDM, IMP, GIM, SPM), and (iii) Ambler class D oxacillinases (OXA-type) prevalent mainly in Acinetobacter baumannii and Pseudomonas aeruginosa [1,2,3]. We have previously reported, from Egypt, (i) the first NDM-5-producing E. coli clinical isolate [4], (ii) the coexistence of NDM-5 and NDM-4 in Klebsiella pneumoniae from a 6-month-old infant [5], (iii) the first NDM-1-producing-Providencia stuartii isolates in an African burn unit [3], (iv) two NDM-5 and OXA-181 co-producing E. coli clinical isolates [3], and (v) the first whole genome sequence of E. coli clinical isolate co-carrying blaNDM-1 and blaOXA-244 [6]. Additionally, we indicated the elevated incidence of carbapenemase-producing Gram-negative bacteria in Egypt, with OXA-48-like and NDM-1 being the most prevalent [7].
As of 19 January 2023, 48 variants of blaNDM are accessible in the Bacterial Antimicrobial Resistance Reference Gene Database (https://www.ncbi.nlm.nih.gov/pathogens/refgene/#NDM, accessed on 19 January 2023). The New Delhi metallo-β-lactamase (NDM) variant, NDM-19, differs from NDM-1 and NDM-7 by three (Asp130Asn, Met154Leu, and Ala233Val) and a single amino acid substitution (Ala233Val), respectively, leading to decreased susceptibility towards carbapenems and extended-spectrum cephalosporins [2,8,9,10]. NDM-19 was previously identified from human clinical E. coli and/or K. pneumoniae isolates from different countries, including China, Egypt, Lebanon, and Switzerland [2,8,9,10].
El-Kholy et al., 2021 reported a high prevalence of carbapenem-resistant Enterobacteriaceae (CRE) (13.8–100% of E. coli and 35–100% of K. pneumoniae) in Egypt [11]. In the United Arab Emirates, an analysis was conducted on a total of 381,535 non-repetitive Enterobacteriaceae from 2010 to 2021, with CRE accounting for 3.8%, representing 74 different species [12]. Meanwhile, 5% of CRE was identified in a recent surveillance study from Saudi Arabia [12]. In Lebanon, the CRE rate was 2.8% in 2022, while in Jordan, it was 1.6% in 2015 [12].
CRE is a group of Gram-negative Enterobacteriaceae that are resistant to carbapenems and can cause serious illnesses such as urinary tract infections, sepsis, and pneumonia [13]. CRE can be found in various settings, including hospitals, the environment, and animals, highlighting a zoonotic transmission [13]. Chickens are raised close to humans, such as in backyards and small-scale farms, and can act as a critical reservoir of CRE. An increased incidence of CRE has been observed in poultry farms, predominantly in E. coli, K. pneumoniae, and P. mirabilis, carrying blaNDM genes, with blaNDM-1 and blaNDM-5 being the most prevalent [14].
The genomic features of NDM-19-producing E. coli in Egypt remain scarce. Therefore, in the current study, we aimed to illustrate the first complete genomic sequence, produced by the Oxford Nanopore and Illumina MiniSeq sequencing techniques, of the NDM-19-producing E. coli strain from a poultry farm in Egypt.
2. Materials and Methods
2.1. Isolation and Identification of E. coli M2-13-1
The E. coli strain M2-13-1 was isolated from a fecal sample collected from a poultry farm in March 2019 in Sidi Ghazy City, Kafr El-Sheikh province, Egypt. The farm has healthy commercial chickens aged 15 to 30 days. The fecal sample was first incubated in Luria-Bertani (LB) (Lennox) broth at 37 °C for 18–24 h. After enrichment, the culture was plated onto MacConkey agar with 2 µg/mL meropenem. The resulting isolate showed resistance to carbapenems. The 16S ribosomal RNA gene amplification and sequencing using primers 27F and 1492R was conducted to identify the isolate [3]. PCR screening was performed for several carbapenemase encoding genes (NDM, VIM, IMP, KPC, OXA-48-like), as previously reported [3]. NDM variants were detected by PCR screening and DNA sequencing by NDM-F (5′-GGTTTGGCGATCTGGTTTTC-3′) and NDM-R (5′-CGGAATGGCTCATCACGATC-3′), NDM-WHF (5′-ATGGAATTGCCCAATATTATGCACC-3′), and NDM-WHR (5′-TCAGCGCAGCTTGTCGG-3′), amplifying PCR products of 621 and 813 base pairs (bp), respectively [3,15]. Sanger sequencing confirmed the presence of the NDM-19 variant. Consequently, the strain was submitted for whole-genome sequencing.
2.2. Minimum Inhibitory Concentration (MIC) of Various Antimicrobial Agents Against M2-13-1
MIC of various antimicrobial agents against M2-13-1 was determined according to the standards and interpretive criteria defined by the Clinical and Laboratory Standards Institute (CLSI, document M100-S24). The following antibiotics were used: aztreonam, cefotaxime, chloramphenicol, ciprofloxacin, colistin, ertapenem, fosfomycin, kanamycin, meropenem, tetracycline, and tigecycline. E. coli ATCC 25922 was used as a control.
2.3. Conjugation and Transformation Experiments
To assess the mobility of the plasmid carrying blaNDM-19, a filter-mating conjugation was achieved using M2-13-1 as the donor and the azide-resistant E. coli strain J53 as a recipient [3]. An E. coli strain carrying NDM-5 on an IncX3 plasmid from our collection was used as a control. The transconjugants were selected on LB agar plates containing 50 μg/mL ampicillin and 100 μg/mL sodium azide or 1 μg/mL imipenem and 100 μg/mL sodium azide [3]. For transformation, plasmid DNA was extracted by a standardized alkaline lysis method, as previously described [3]. Plasmids were CaCl2-transformed into E. coli DH5α. Transformants were selected on LB agar plates plus 1 μg/mL meropenem or 100 μg/mL ampicillin. Colony-direct PCR was accomplished utilizing the primers NDM-F and NDM-R to verify the transmission of the blaNDM-19-containing plasmid [15].
2.4. Complete Genome Sequencing (WGS) and Analysis
The total genomic DNA (gDNA) of the E. coli M2-13-1 strain was prepared from an overnight bacterial culture using a standard proteinase K method, as previously reported [16]. DeNovix (Wilmington, DE, USA), Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA), and a Microdrop (Thermo Scientific™, Multiskan™ Sky Microplate Spectrophotometer, Waltham, MA, USA) were utilized to assess the quality of the isolated gDNA. For Illumina MiniSeq sequencing, a Nextera DNA Flex Library Prep Kit (Illumina, San Diego, CA, USA) was employed to construct the library. For Oxford Nanopore sequencing, the Rapid Barcoding Sequencing kit (SQK-RBK004) (Oxford Nanopore Technologies, Oxford, UK) was used to construct the DNA library, which was then loaded onto a flow cell (FLO-MIN106) and sequenced for 48 h with GridION (Oxford Nanopore Technologies). Flye v2.6 (https://github.com/fenderglass/Flye) was used to perform the hybrid assembly of both Illumina short reads and Nanopore long reads. The genome assembly was further refined using Pilon-v1.23 [17]. Annotation was performed using DFAST (https://dfast.nig.ac.jp/) [18]. The complete genome of the E. coli M2-13-1 strain was examined using ResFinder-3.2, PlasmidFinder-2.0, SerotypeFinder-2.0, MLST 2.0, pMLST-2.0, and CHTyper-1.0, accessible at the Center for Genomic Epidemiology (http://www.genomicepidemiology.org/). Phylogenetic grouping was identified using the Clermont Typing scheme available at (http://clermontyping.iame-research.center/). A circular comparison between the complete sequence of pAMS-X3-NDM-19 and the highly similar plasmids identified via a NCBI nucleotide BLAST (BLASTn) (https://blast.ncbi.nlm.nih.gov/Blast.cgi) search was conducted using the BRIG tool [19]. The genetic environment of blaNDM-19 was illustrated through a BLASTn search and ISFinder (https://www-is.biotoul.fr/blast.php) to detect insertion sequence (IS) elements. Visualization and comparison of the genetic environment of blaNDM-19 in pAMS-X3-NDM-19 was accomplished with the EasyFig_win_2.1 tool (http://mjsull.github.io/Easyfig/). The virulome of the NDM-19-producing E. coli strain M2-13-1 was predicted using the virulence factor database, a JavaScript-rich interface with VFanalyzer (http://www.mgc.ac.cn/VFs/).
For a WGS-based phylogenetic analysis, fastq files of the sequenced E. coli isolate were imported into Enterobase (https://enterobase.warwick.ac.uk/, version 1.2.0) and compared to publicly available genomes of ST167 E. coli (n = 198) that were randomly selected to represent different sources and countries, using single-nucleotide polymorphisms (SNPs) and hierarchical clustering (HierCC) of core genome (cg) MLST. Different sets of hierarchical clusters (HCs) were defined in Enterobase to cluster bacterial genomes based on variations in core genomic loci across bacteria [20]. The present study isolates and selected isolates from Enterobase were mapped to the E. coli K-12 MG1655 reference genome for SNPs analysis.
2.5. Nucleotide Sequence Accession Numbers
The complete genome sequence of the E. coli M2-13-1 strain was submitted to DDBJ/ENA/GenBank under BioProject ID: PRJNA946682.
3. Results and Discussion
3.1. Minimum Inhibitory Concentration (MIC) of Various Antimicrobials Against M2-13-1
The 16S ribosomal RNA gene sequence identified the strain as belonging to E. coli. Broth microdilution exhibited that M2-13-1 was resistant to ceftriaxone, chloramphenicol, meropenem, ertapenem, kanamycin, ciprofloxacin, and tetracycline. The strain showed sensitivity to aztreonam, colistin, tigecycline, and fosfomycin (Table 1). PCR screening and DNA sequencing recognized the existence of the metallo-β-lactamase genes, blaNDM-19.

Table 1.
MICs of antimicrobials for NDM-19- and QnrS11-producing Escherichia coli strain and its transformant/transconjugants isolated in this study.
3.2. Genomic Characterization of E. coli M2-13-1
The complete genome of E. coli M2-13-1 was obtained from high-quality hybrid assemblies after the combination of Illumina short reads and Nanopore long reads. The assembly was adequate for circularizing the chromosome and the plasmids. The final polished assembly of M2-13-1 comprised 10 contigs totaling 5.13 Mb with an N50 of 4.74 Mb (L50 = 1). The mean sequencing depth was ~85×, and the overall GC content was 50.7%. A total of 5010 coding sequences, 22 rRNAs, 90 tRNAs, and 2 CRISPRs were predicted. The chromosome of the E. coli M2-13-1 strain was 4,738,278 bp in size with an average GC content of 50.8% (Table 2). The MLST2.0 server revealed that M2-13-1 belonged to ST167 [allelic profile (adk 10, fumC 11, icd 8, purA 13, gyrB 4, recA 2, and mdh 8)]. SerotypeFinder-2 and ClermonTyping servers revealed that M2-13-1 belonged to serotype O101:H5 and phylogroup A, respectively.
Table 2.
Features of the chromosome and the plasmids of E. coli strain M2-13-1 isolated from a poultry farm in Egypt.
ST167 E. coli producing different NDM enzymes were reported from different countries, including China (NDM-1 and NDM-7), Egypt (NDM-1 and NDM-5), Guatemala (NDM-1), Italy (NDM-5), and Thailand (NDM-5) [21,22,23].
Although the strain was isolated from a poultry farm fecal sample, it still lacks the virulence genes necessary for classifying it as an avian pathogenic E. coli (APEC) genetic pathotype. According to Johnson et al., 2008, an isolate is often considered APEC if it carries five or more of the following virulence genes: iroN, ompT, hlyF, iss, iutA [24]. M2-13-1 is not APEC because it only carries iss and lacks the other four genes. Based on the resistance gene profile (NDM-19 on IncX3 plasmid) and ST167, this strain is likely associated with human infection and may belong to the ExPEC pathotype, as previously reported, suggesting transmission between animals and humans and vice versa [25].
3.3. Plasmidome and Resistome of E. coli M2-13-1
The strain carried four different plasmids belonging to various Inc groups, including IncFII: IncFIB, IncI1-Iγ, IncY, and IncX3, ranging in size from 49,816 bp to 113,285 bp (Table 2). Additionally, the strain carried 13 different acquired antimicrobial resistance genes potentially conferring resistance to quinolone (qnrS11), tetracycline [tet(A)], trimethoprim (dfrA14), β-lactams (blaNDM-19 and blaTEM-1B), aminoglycosides [aac(3)-IV, aph(3″)-Ib, aph(4)-Ia, aph(6)-Id, and aph(3′)-Ia], macrolides [mdf(A)], chloramphenicol and florfenicol (floR), and sulfonamides (sul2) (Table 2). The quinolone resistance-determining region was found to be mutated (parC: S80I, parE: S458A, gyrA: S83L, and D87N) (Table 2), conferring resistance to ciprofloxacin and nalidixic acid.
3.4. Identification of a blaNDM-19/IncX3 Plasmid in an Egyptian E. coli Poultry Strain
New Delhi metallo-β-lactamase (NDM) is a widely distributed Ambler class B carbapenemase that confers high-level resistance to all β-lactams, except aztreonam, and is found in various Gram-negative bacteria from different sources, including humans, animals, and the environment [26]. NDM-19 was first detected in an E. coli strain isolated from an Egyptian patient in a Swiss hospital [8]. Subsequently, NDM-19 was identified on an IncX3 plasmid in a carbapenem-resistant (CR) ST15 K. pneumoniae strain isolated from a sputum swab of a patient with chronic obstructive pulmonary disease in China [9]. Additionally, an Egyptian clinical CR K. pneumoniae ST353 was reported to carry blaNDM-19 in 2018 [2]. Moreover, nine NDM-19-positive E. coli clinical strains were reported in Northern Lebanon, isolated between 2015 and 2019 [10].
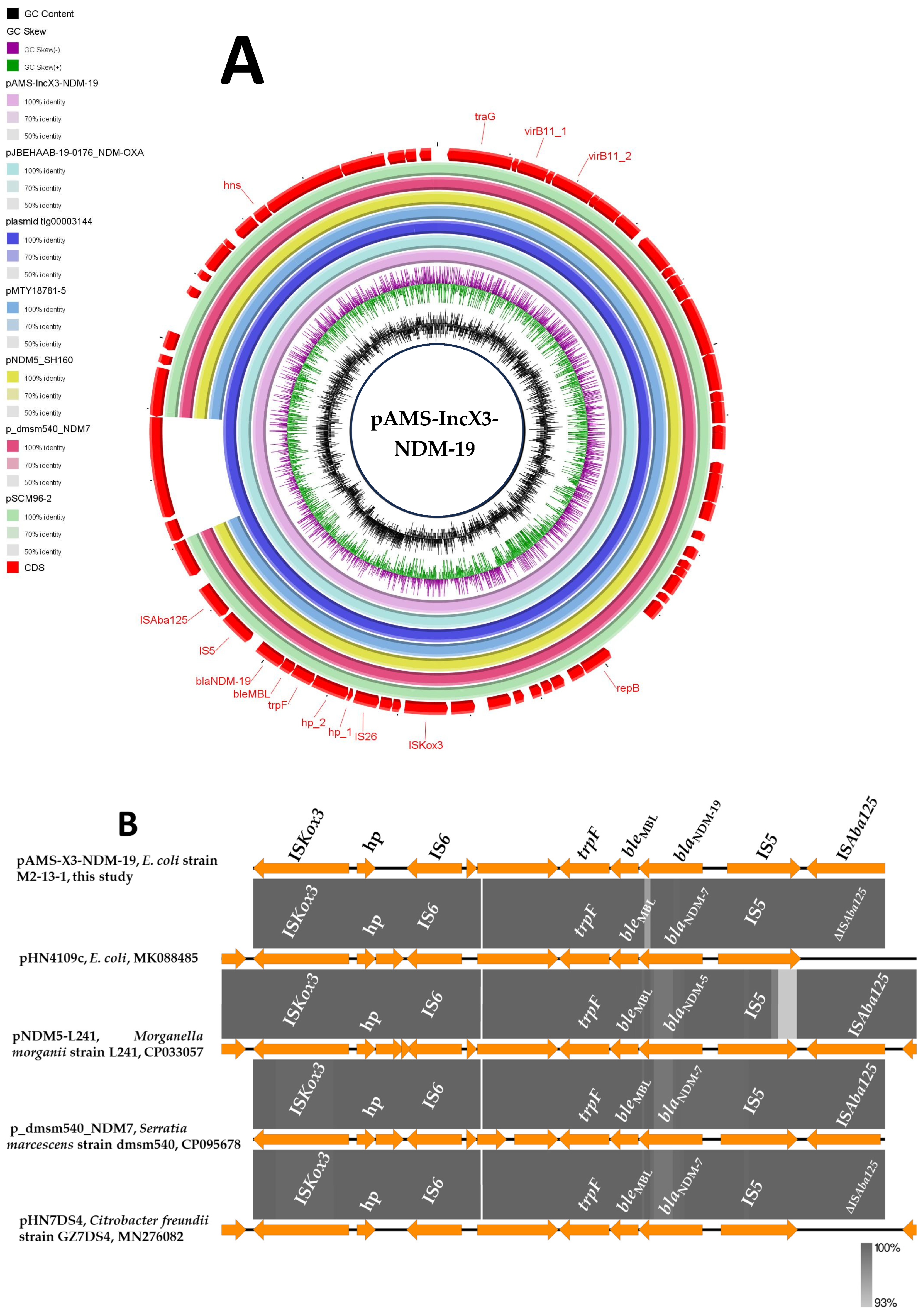
blaNDM-19 was located on the plasmid pAMS-X3-NDM-19. pAMS-X3-NDM-19 is an IncX3 plasmid of 49,816 bp in size encoding 64 CDS with an average G + C content of 47.3% (Figure 1 and Table 2). Plasmids of IncX3 replicons (i) have narrow host range [27]; (ii) have been frequently identified in Enterobacteriaceae of environmental, clinical, and animal origin [27]; (iii) are featured by carrying carbapenemase-encoding genes (e.g., blaNDM, blaKPC, and blaOXA-181) [27]; and (iv) are considered as a critical vehicle for the transmission of blaNDM and its rare new variants with the significant role of the insertion sequence (ISAba125) [27].
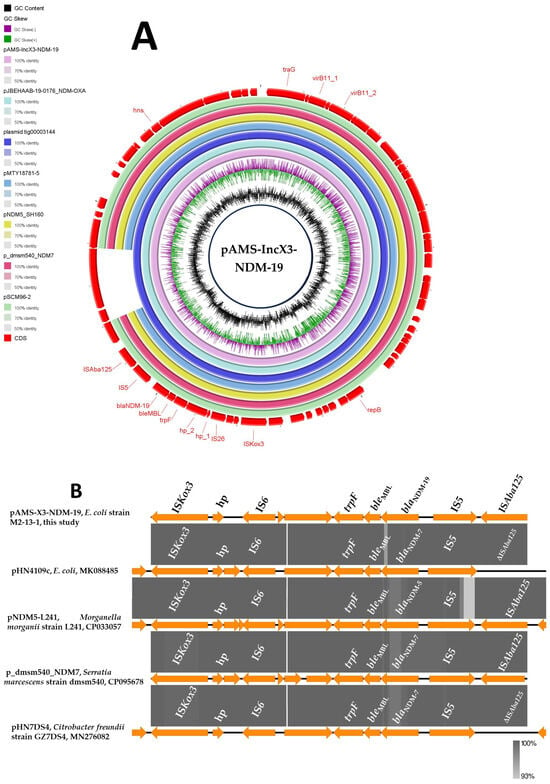
Figure 1.
Schematic representation of IncX3 plasmid carrying blaNDM-19 (A), and the genetic environment of blaNDM-19 (B) identified from the genomic sequence of E. coli strain M2-13-1 analyzed in this study. In panel A, six IncX3 plasmids, pJBEHAAB-19-0176_NDM-OXA, plasmid tig00003144, pMTY18781-5, pNDM5_SH160, p_dmsm540_NDM7, and pSCM96-2 carrying blaNDM (accession no. AP028870.1, CP021682.1, AP023210.1, CP053295.1, CP095678.1, and CP028718.1, respectively) have been detected from NCBI GenBank and were included in the figure. The whole sequence of pAMS-IncX3-NDM-19 was used as the reference. The external ring represents the annotation of pAMS-IncX3-NDM-19. The plasmids were included in the following order: pAMS-IncX3-NDM-19 (identified in this study), pJBEHAAB-19-0176_NDM-OXA, plasmid tig00003144, pMTY18781-5, pNDM5_SH160, pSCM96-2, p_dmsm540_NDM7, and pAMS-X3-NDM-19.
A BLASTn search with the entire pAMS-X3-NDM-19 sequence query identified that it has high similarity to other IncX3 plasmids carrying blaNDM. For instance, pAMS-3-NDM-19 showed more than 99.9% sequence identity to E. coli plasmids (i) plasmid tig00003144 carrying blaNDM-7 (100% coverage; GenBank: CP021682.1), (ii) blaNDM-5-carrying pMTY18781-5 isolated from human stool in Japan in 2018 (92% coverage; GenBank: AP023210.1), and (iii) blaNDM-5- and blaOXA-181-carrying pJBEHAAB-19-0176_NDM-OXA isolated from human stool in Japan in 2020 (100% coverage; GenBank: AP028870.1) (Figure 1A). In addition, pAMS-X3-NDM-19 has >99.9% identity to blaNDM-19-bearing K. pneumoniae IncX3 plasmid pSCM96-2 isolated from a sputum sample in China in 2017 (92% coverage; GenBank: CP028718.1), blaNDM-7-bearing Serratia marcescens IncX3 plasmid p_dmsm540_NDM7 isolated from a blood sample in Bangladesh in 2017 (92% coverage; GenBank: CP095678.1), and Salmonella enterica subsp. enterica serovar Typhimurium plasmid pNDM5_SH160 carrying blaNDM-5 (92% coverage; GenBank: CP053295.1) (Figure 1A).
The results of conjugation experiments showed that the pAMS-X3-NDM-19 plasmid was not successfully transferred to E. coli J53 after multiple attempts, despite the successful transfer of the control NDM-5 carrying IncX3 plasmid. Most IncX3 plasmids are capable of conjugation, with a frequency ranging from 10−9 to 10−2 [21]. Several reports described the impaired conjugation ability of IncX3 plasmids (reviewed in [21]). In some cases, this is caused by the destruction or deletion of conjugation genes. However, other IncX3 plasmids remain non-conjugative despite having the entire conjugation backbone, indicating additional regulatory mechanisms. For example, plasmid pWLK-NDM (GenBank: CP038280) was isolated from urban river debris in China and is non-conjugable due to disruption of the pilX3-4 gene, even though the conjugation genes are present [27]. Furthermore, a recent study identified two key factors controlling IncX3 plasmid conjugation: H-NS, a putative DNA-binding protein, inhibits conjugation by directly repressing the expression of the activator PrfaH, whose deletion eliminates conjugative transfer [28].
The genetic environment of blaNDM-19 was as follows: ISAba125 (IS30 family)-IS5 (IS5 family)-blaNDM-19-bleMBL (bleomycin resistance gene)-trpF (phosphoribosylanthranilate isomerase)-hp-hp-IS26 (IS6 family)-hp-ISKox3 (ISL3 family). A BLASTn search using the immediate genetic vicinity of blaNDM-19 identified the similarity to other sequences (100% query coverage and ≥99.94% sequence identity) from different organisms, including E. coli plasmid pHN4109c (GenBank: MK088485.1), plasmid pKpN01-NDM7 (GenBank: CP012990.1) in clinical K. pneumoniae strain KpN01 from Canada, plasmid p_dmsm540_NDM7 (GenBank: CP095678.1) in clinical S. marcescens strain dmsm540 from Bangladesh, plasmid pHN7DS4 (GenBank: MN276082.1) from Citrobacter freundii strain GZ7DS4 isolated from a dog in China, and plasmid pNDM5-L241 (GenBank: CP033057.1) from clinical Morganella morganii strain L241 detected in China (Figure 1B).
3.5. Detection of Other Multidrug-Resistant (MDR) Plasmids from E. coli M2-13-1
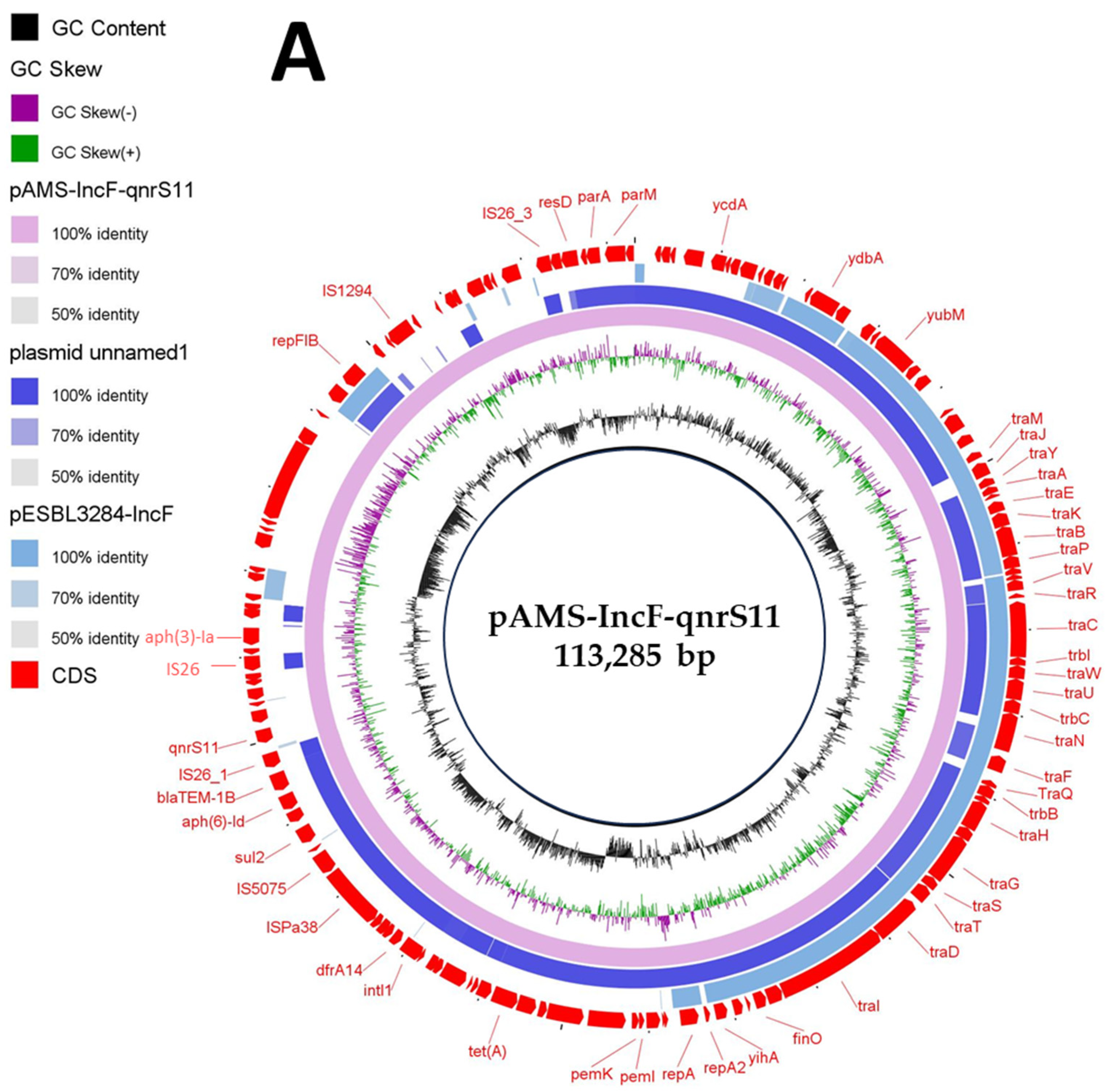
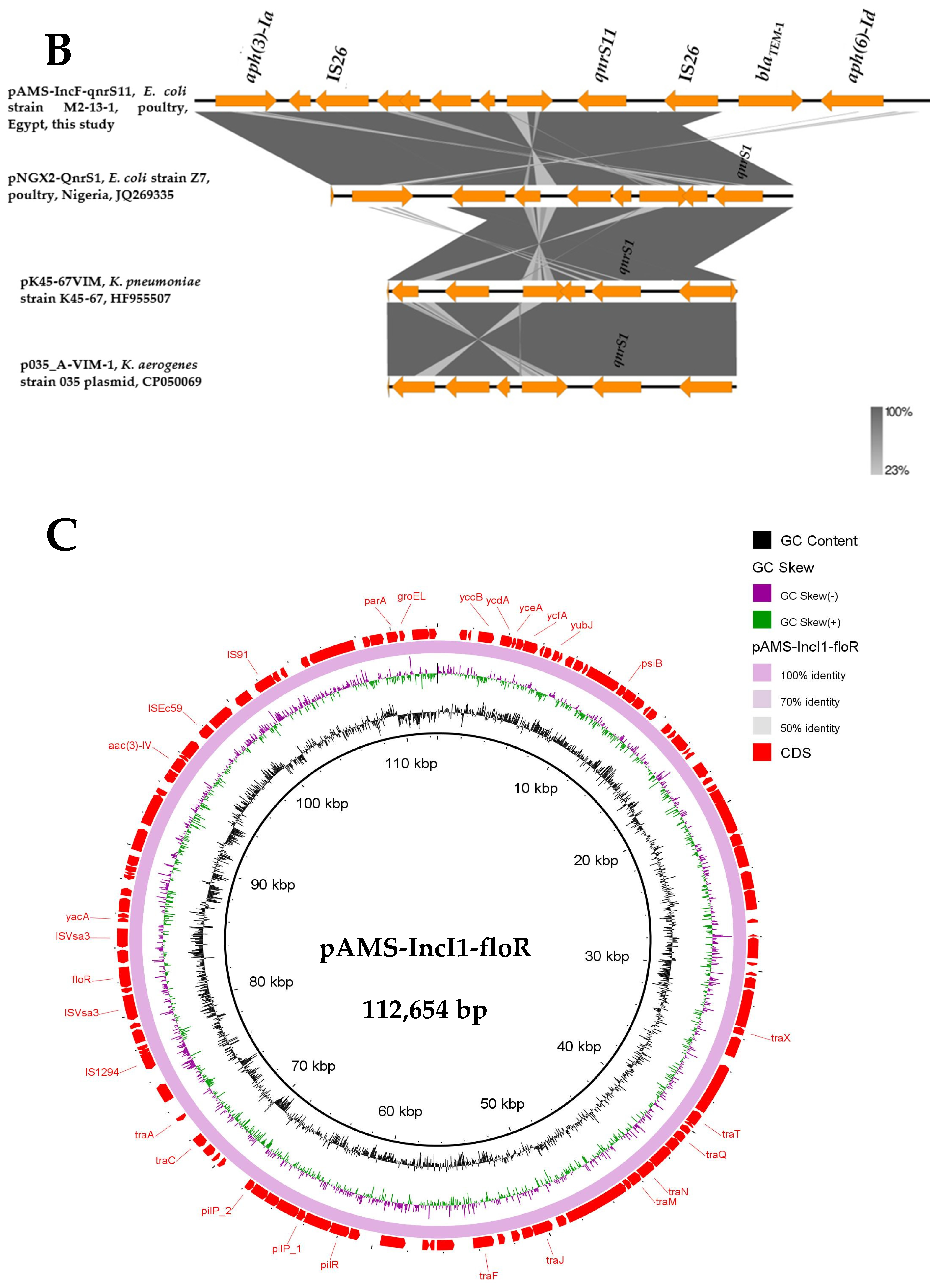
Here, MDR plasmids is defined as plasmids carrying genes conferring resistance to multiple antibiotics and can be transferred by horizontal gene transfer (HGT). E. coli M2-13-1, detected in the current study, contained another two MDR plasmids [pAMS-IncF-qnrs11 (∼113 Kb IncFII: IncFIB) (Figure 2A,B) and pAMS-IncI1-floR (∼113 Kb IncI1-Iγ) (Figure 2C)] carrying eight (aph(6)-Id, aph(3′)-Ia, aph(3″)-Ib, qnrS11, dfrA14, sul2, tet(A), blaTEM-1B) and four (aac(3)-IV, aph(4)-Ia, sul2, floR) resistant genes, respectively (Table 2, Figure 2).
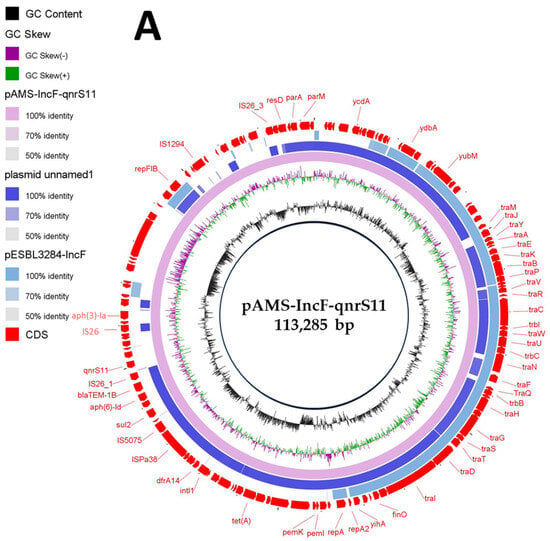
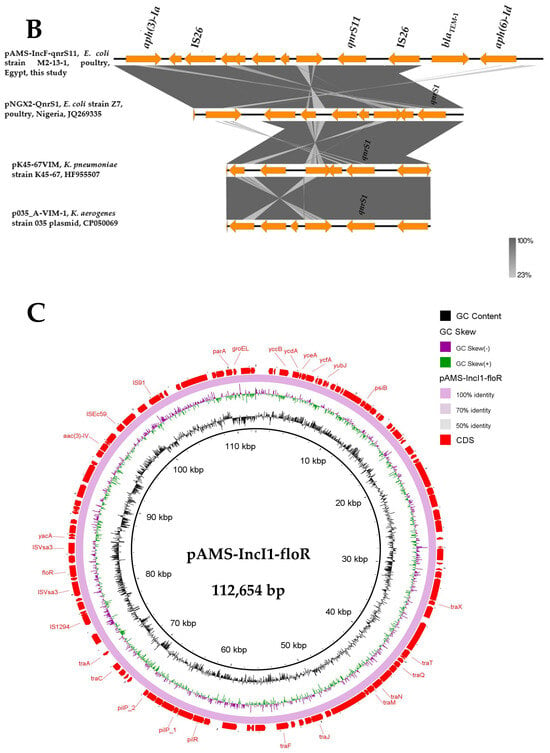
Figure 2.
Schematic representation of IncF plasmid carrying qnrS (A), the genetic environment of qnrS11 (B), and IncF plasmid carrying floR (C) identified from the genome sequences of E. coli strain M2-13-1 analyzed in this study. In panel A, two IncF plasmids, plasmid unnamed1 (GenBank: CP020510.1), and plasmid pESBL3284-IncF (GenBank: MW390540.1) have been detected from NCBI GenBank and were included in the figure. The whole sequence of pAMS-IncF-qnrS11 was used as the reference. The external ring represents the annotation of pAMS-IncF-qnrS11. The plasmids were included in the following order: pAMS-IncF-qnrS11 (identified in this study), plasmid unnamed1, and plasmid pESBL3284-IncF.
IncF plasmids are (i) one of the most commonly detected plasmid types in Enterobacteriaceae, (ii) frequently of low copy number, (iii) MDR with more than 100 kb in size, and (iv) multireplicon to enhance the replication initiation (FII, FIA, and FIB) [29]. A BLASTn search with the whole pAMS-IncF-qnrs11 sequence query identified that it has more than 99.1% identity to plasmid unnamed1 isolated from E. coli strain 165 isolated from a patient in the USA in 2015 (76% coverage; GenBank: CP020510.1), and 98.7% identity to plasmid pESBL3284-IncF E. coli strain ESBL3284 (46.8% coverage; GenBank: MW390540.1) (Figure 2A). pAMS-IncF-qnrs11 was successfully transferred by conjugation to E. coli J53. The transconjugant showed resistance to kanamycin (MIC, >256 μg/mL) and tetracycline (MIC, 64 μg/mL) but showed sensitivity to aztreonam (MIC, <2 μg/mL), ciprofloxacin (MIC, 0.5 μg/mL), chloramphenicol (MIC, 4 μg/mL), ceftriaxone (MIC, <2 μg/mL), colistin (MIC, <0.5 μg/mL), meropenem (MIC, <0.5 μg/mL), ertapenem (MIC, <0.5 μg/mL), tigecycline (MIC, <0.5 μg/mL), and fosfomycin (MIC, 4 μg/mL).
The genetic context of qnrS11 was aph(6)-Id-blaTEM-1B-IS26-qnrS11-a gene encoding TniR protein-hp-hp-IS26 (Figure 2B). qnrS11 was found between two copies of IS26 in a transposon-like organization, which might be involved in its mobilization. A BLASTn search using the genetic environment of qnrS11 identified the similarity to other sequences (≥99.89% sequence identity) from (i) plasmid pNGX2-QnrS1 identified from E. coli strain Z7 isolated from poultry in Nigeria (84% coverage; GenBank: JQ269335), (ii) plasmid pK45-67VIM identified from K. pneumoniae strain K45-67, (56% coverage; GenBank: HF955507), and (iii) plasmid p035_A-VIM-1 detected from K. aerogenes strain 035 (72% coverage; GenBank: CP050069). QnrS11 was previously reported on an IncFII plasmid in an environmental quinolone-resistant E. coli strain [30].
3.6. Analysis of the Virulome and Heavy Metal Resistance Genes
Numerous virulence-associated genes have been predicted, illustrating the pathogenicity potential associated with the consumption of contaminated chicken meat (Table 3). E. coli common pilus (ECP) (ecpABCDE), E. coli laminin-binding fimbriae (elfACD), eaeH, hemorrhagic E. coli pilus (hcpABC), and type I fimbriae (fimDF) were detected from the M2-13-1 chromosome and were mainly associated with adherence (Table 3). Type IV pili (pilQRSVW) was predicted from pAMS-IncI1-floR. It was suggested that pathogenic E. coli strains might use ECP to simulate commensal E. coli with an advantage for evasion of the immune system after the host colonization [31]. EhaB autotransporter protein promotes biofilm formation and binds to collagen I and laminin, inducing a serum IgA response in E. coli O157:H7-challenged cattle [32,33]. The invasion genes ibeBC contribute to E. coli invasion of brain microvascular endothelial cells (BMECs) or the blood–brain barrier, causing positive culture of the cerebrospinal fluid [34]. The epithelial cell adherence gene tia is an outer membrane protein implicated in the initial infection in ETEC strain H10407 and was detected in M2-13-1 analyzed in the current study [35]. Additionally, M2-13-1 is predicted to have genes encoding a type VI secretion system (T6SS), which penetrates epithelial cells, replicates within macrophages, and results in fatal illness in chicks [36].
Table 3.
The virulome of the NDM-19-producing E. coli strain M2-13-1 according to the virulence factor database (http://www.mgc.ac.cn/VFs/) a.
3.7. Clonal Relationship of ST167 E. coli Isolate
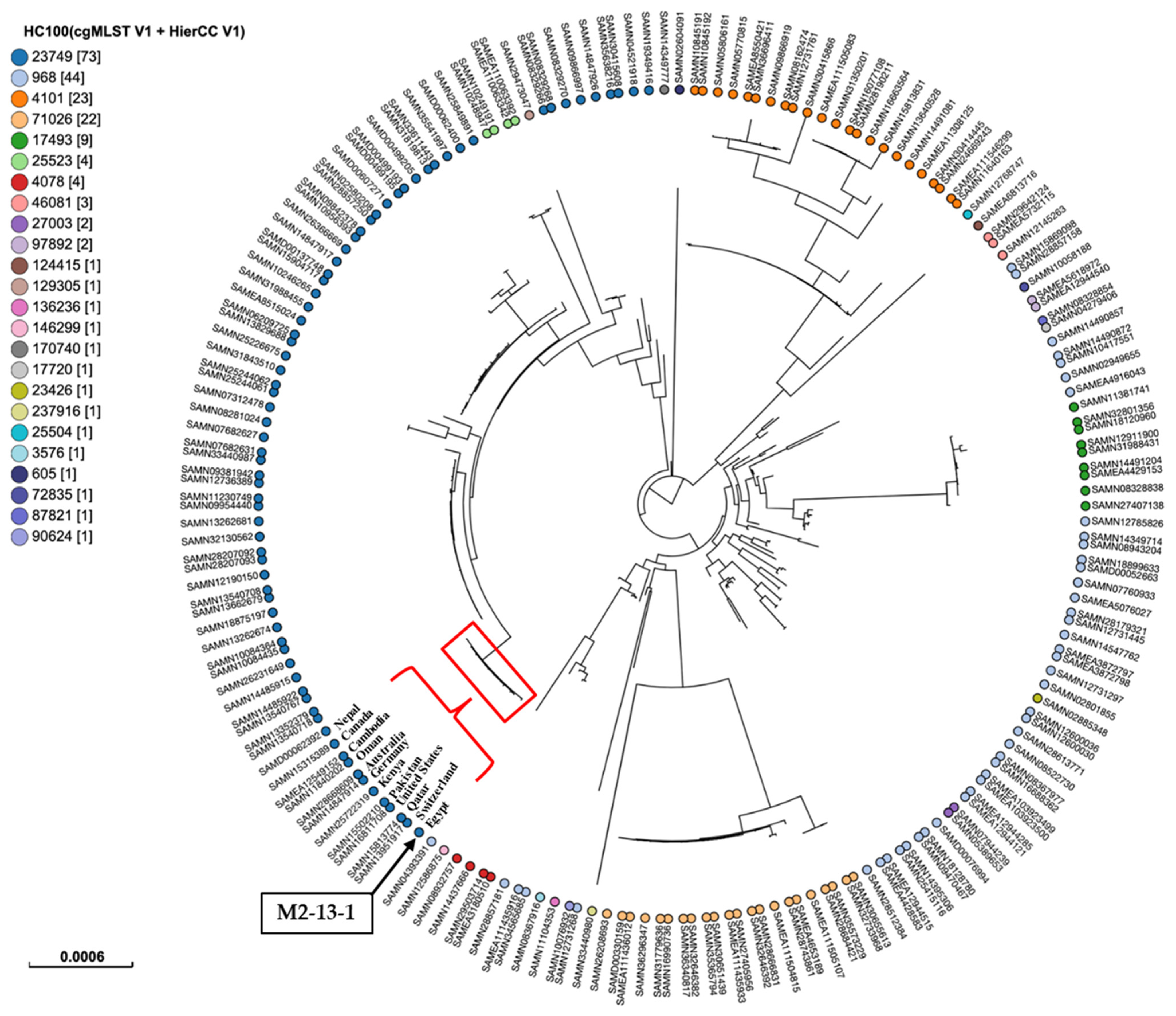
To determine the clonal relatedness between our isolate and global lineages of ST167, we performed a phylogenetic analysis based on SNPs and HierCC of cgMLST (Figure 3). The isolate was compared to 198 publicly available isolates of ST167 sourced from different hosts. Metadata of all strains were mentioned in Supplementary Table S1, associated with this article. The findings revealed the classification of isolates into 24 patterns at the HC100 level, differing by 100 core genomic loci. Of these HC100 patterns, four patterns: HC100|23749, HC100|968, HC100|4101, and HC100|71026 were recognized as the major patterns, assigned to 73, 44, 23, and 22 isolates, respectively. Our isolate shared the same HC100 pattern, HC100|23749, with other ST167 E. coli isolates from various countries, including the United States, Canada, Switzerland, Germany, the United Kingdom, Finland, Qatar, Oman, Pakistan, India, Nepal, Bangladesh, Kenya, Australia, Cambodia, China, Thailand, and Japan. The circulation of similar bacterial clones across different countries highlights the importance of implementing WGS-based phylogeny for routine epidemiological surveillance of infectious diseases, particularly those caused by clones that cross borders [37].
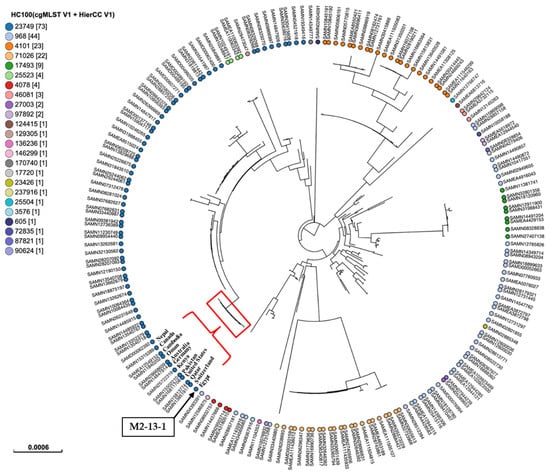
Figure 3.
SNPs and hierarchical clustering of cgMLST (HierCC) of the present E. coli isolate ST167 M2-13-1 with the publicly available E. coli isolates ST167 sourced from different hosts in EnteroBase (https://enterobase.warwick.ac.uk/). Tip labels indicate the cgMLST pattern HC100 (allelic differences no more than 100) for ST167. The numbers in brackets indicate the number of isolates assigned to each HC100 cluster. The position of M2-13-1 was marked with a black box and a black arrow. M2-13-1 was grouped with several isolates from different countries and marked with a red triangle and curly bracket.
4. Conclusions
Here, we reported, to our knowledge, the first (i) detection of NDM-19-producing E. coli strain of chicken origin globally, and (ii) complete genome sequence of a carbapenem-resistant E. coli strain producing NDM-19 from Africa and the Middle East. This study demonstrated that breeding animals may serve as a crucial reservoir of multidrug-resistant bacteria carrying plasmids encoding resistance genes (blaNDM-19) to clinically important antibiotics, including imipenem and meropenem. Additionally, the finding of these E. coli clones from Egyptian poultry is disturbing and causes food security and public health concerns, as they might be transmitted to individuals. Plasmids of the IncX3 type and the two insertion sequences (ISAba125 and IS26) might be important tools for the spread of blaNDM-19. A survey for blaNDM-19 and other carbapenemase-encoding genes in animals is essential in Egypt to define its incidence and to stop its spread. Infection prevention and control policies, antimicrobial stewardship and surveillance plans, as well as the careful use of β-lactams in humans and animals are effectively required. The major limitation of the study is its dependence on a single bacterial strain for experimental analysis, restricting the broader application of the results.
Supplementary Materials
The following Supporting Information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms13081769/s1, Supplementary Table S1: Metadata of the strains used in the phylogenetic tree.
Author Contributions
A.M.S., T.S. (Tadashi Shimamoto), conceptualization and design of the study; A.M.S., methodology and investigation; T.S. (Tadashi Shimamoto), supervision; T.S. (Tadashi Shimamoto), F.M., funding acquisition; A.M.S., F.M., T.S. (Toshi Shimamoto), T.K., H.R., analysis and interpretation of the results; A.M.S., T.S. (Tadashi Shimamoto), writing—original and final draft. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported in part by a grant to F.M. from the Japan Agency for Medical Research and Development (grant no. 23wm0225038h0001).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The complete genome sequence of the E. coli M2-13-1 strain was submitted to DDBJ/ENA/GenBank under BioProject ID: PRJNA946682.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- El-Gamal, M.I.; Brahim, I.; Hisham, N.; Aladdin, R.; Mohammed, H.; Bahaaeldin, A. Recent updates of carbapenem antibiotics. Eur. J. Med. Chem. 2017, 131, 185–195. [Google Scholar] [CrossRef]
- Gamal, D.; Egea, P.; Elías, C.; Fernández-Martínez, M.; Causse, M.; Pérez-Nadales, E.; Salem, D.; Fam, N.; Diab, M.; Aitta, A.A.; et al. High-risk clones and novel sequence type ST4497 of Klebsiella pneumoniae clinical isolates producing different alleles of NDM-type and other carbapenemases from a single tertiary-care centre in Egypt. Int. J. Antimicrob. Agents 2020, 56, 106164. [Google Scholar] [CrossRef]
- Soliman, A.M.; Zarad, H.O.; Nariya, H.; Shimamoto, T.; Shimamoto, T. Genetic analysis of carbapenemase-producing Gram-negative bacteria isolated from a university teaching hospital in Egypt. Infect. Genet. Evol. 2020, 77, 104065. [Google Scholar] [CrossRef]
- Soliman, A.M.; Khalifa, H.O.; Ahmed, A.M.; Shimamoto, T.; Shimamoto, T. Emergence of an NDM-5-producing clinical Escherichia coli isolate in Egypt. Int. J. Infect. Dis. 2016, 48, 46–48. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, H.O.; Soliman, A.M.; Ahmed, A.M.; Shimamoto, T.; Shimamoto, T. NDM-4- and NDM-5-producing Klebsiella pneumoniae coinfection in a 6-month-old infant. Antimicrob. Agents Chemother. 2016, 60, 4416–4417. [Google Scholar] [CrossRef]
- Soliman, A.M.; Ramadan, H.; Sadek, M.; Nariya, H.; Shimamoto, T.; Hiott, L.M.; Frye, J.G.; Jackson, C.R.; Shimamoto, T. Draft genome sequence of a blaNDM-1- and blaOXA-244-carrying multidrug-resistant Escherichia coli D-ST69 clinical isolate from Egypt. J. Glob. Antimicrob. Resist. 2020, 22, 832–834. [Google Scholar] [CrossRef]
- Khalifa, H.O.; Soliman, A.M.; Ahmed, A.M.; Shimamoto, T.; Hara, T.; Ikeda, M.; Kuroo, Y.; Kayama, S.; Sugai, M.; Shimamoto, T. High carbapenem resistance in clinical Gram-negative pathogens isolated in Egypt. Microb. Drug Resist. 2017, 23, 838–844. [Google Scholar] [CrossRef] [PubMed]
- Mancini, S.; Keller, P.M.; Greiner, M.; Bruderer, V.; Imkamp, F. Detection of NDM-19, a novel variant of the New Delhi metallo-β-lactamase with increased carbapenemase activity under zinc-limited conditions, in Switzerland. Diagn. Microbiol. Infect. Dis. 2019, 95, 114851. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, H.; Zhang, X.; Jiang, N.; Zhang, Z.; Zhang, J.; Zhu, B.; Wang, G.; Zhao, K.; Zhou, Y. Characterization of an NDM-19-producing Klebsiella pneumoniae strain harboring 2 resistance plasmids from China. Diagn. Microbiol. Infect. Dis. 2019, 93, 355–361. [Google Scholar] [CrossRef]
- Rima, M.; Oueslati, S.; Dabos, L.; Daaboul, D.; Mallat, H.; Bou Raad, E.; Achkar, M.; Mawlawi, O.; Bernabeu, S.; Bonnin, R.A.; et al. Prevalence and molecular mechanisms of carbapenem resistance among Gram-negative Bacilli in three hospitals of northern Lebanon. Antibiotics 2022, 11, 1295. [Google Scholar] [CrossRef] [PubMed]
- El-Kholy, A.; El-Mahallawy, H.A.; Elsharnouby, N.; Abdel Aziz, M.; Helmy, A.M.; Kotb, R. Landscape of Multidrug-Resistant Gram-Negative Infections in Egypt: Survey and Literature Review. Infect. Drug Resist. 2021, 14, 1905–1920. [Google Scholar] [CrossRef]
- Thomsen, J.; Abdulrazzaq, N.M.; The UAE AMR Surveillance Consortium; Everett, D.B.; Menezes, G.A.; Senok, A.; Ayoub Moubareck, C. Carbapenem resistant Enterobacterales in the United Arab Emirates: A retrospective analysis from 2010 to 2021. Front. Public Health 2023, 11, 1244482. [Google Scholar] [CrossRef]
- Tuhamize, B.; Bazira, J. Carbapenem-resistant Enterobacteriaceae in the livestock, humans and environmental samples around the globe: A systematic review and meta-analysis. Sci. Rep. 2024, 14, 16333. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Castillo, F.Y.; Guerrero-Barrera, A.L.; Avelar-González, F.J. An overview of carbapenem-resistant organisms from food-producing animals, seafood, aquaculture, companion animals, and wildlife. Front. Vet. Sci. 2023, 10, 1158588. [Google Scholar] [CrossRef]
- Poirel, L.; Walsh, T.R.; Cuvillier, V.; Nordmann, P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn. Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Soliman, A.M.; Maruyama, F.; Zarad, H.O.; Ota, A.; Nariya, H.; Shimamoto, T.; Shimamoto, T. Emergence of a multidrug-resistant Enterobacter hormaechei clinical isolate from Egypt co-harboring mcr-9 and blaVIM-4. Microorganisms 2020, 8, 595. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Tanizawa, Y.; Fujisawa, T.; Nakamura, Y. DFAST: A flexible prokaryotic genome annotation pipeline for faster genome publication. Bioinformatics 2018, 34, 1037–1039. [Google Scholar] [CrossRef]
- Alikhan, N.F.; Petty, N.K.; Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef]
- Zhou, Z.; Alikhan, N.F.; Mohamed, K.; Fan, Y.; Achtman, M.; Agama Study Group. The EnteroBase user’s guide, with case studies on Salmonella transmissions, Yersinia pestis phylogeny, and Escherichia core genomic diversity. Genome Res. 2020, 30, 138–152. [Google Scholar] [CrossRef]
- Peirano, G.; Chen, L.; Nobrega, D.; Finn, T.J.; Kreiswirth, B.N.; DeVinney, R.; Pitout, J.D. Genomic epidemiology of global carbapenemase-producing Escherichia coli, 2015–2017. Emerg. Infect. Dis. 2022, 28, 924–931. [Google Scholar] [CrossRef]
- Shen, P.; Yi, M.; Fu, Y.; Ruan, Z.; Du, X.; Yu, Y.; Xie, X. Detection of an Escherichia coli sequence type 167 strain with two tandem copies of blaNDM-1 in the chromosome. J. Clin. Microbiol. 2016, 55, 199–205. [Google Scholar] [CrossRef]
- Xia, S.; Wang, W.; Cheng, J.; Zhang, T.; Xia, Z.; Zhao, X.; Han, Y.; Li, Y.; Shi, X.; Qin, S. Emergence of a novel hybrid mcr-1-bearing plasmid in an NDM-7-producing ST167 Escherichia coli strain of clinical origin. Front. Microbiol. 2020, 13, 950087. [Google Scholar] [CrossRef]
- Johnson, T.J.; Wannemuehler, Y.; Doetkott, C.; Johnson, S.J.; Rosenberger, S.C.; Nolan, L.K. Identification of minimal predictors of avian pathogenic Escherichia coli virulence for use as a rapid diagnostic tool. J. Clin. Microbiol. 2008, 46, 3987–3996. [Google Scholar] [CrossRef]
- Pan, S.; Liu, S.; Tai, S.; Yu, J.; Yuan, E.; Duan, Y. Genomic analysis of an Escherichia coli sequence type 167 isolate harboring a multidrug-resistant conjugative plasmid, suggesting the potential transmission of the type strains from animals to humans. Infect. Drug Resist. 2023, 16, 5077–5084. [Google Scholar] [CrossRef] [PubMed]
- Boyd, S.E.; Livermore, D.M.; Hooper, D.C.; Hope, W.W. Metallo-β-lactamases: Structure, function, epidemiology, treatment options, and the development pipeline. Antimicrob. Agents Chemother. 2020, 64, e00397-20. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Chen, R.; Wang, Q.; Li, C.; Ge, H.; Qiao, J.; Li, Y. Global prevalence, characteristics, and future prospects of IncX3 plasmids: A review. Front. Microbiol. 2022, 13, 979558. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lu, Y.; Yu, J.; Cai, X.; Wang, C.; Lv, L.; Moran, R.A.; Zhao, X.; Hu, Z.; Deng, M.; et al. Comprehensive analysis of Enterobacteriaceae IncX plasmids reveals robust conjugation regulators PrfaH, H-NS, and conjugation-fitness tradeoff. Commun. Biol. 2025, 8, 363. [Google Scholar] [CrossRef]
- Carattoli, A. Plasmids and the spread of resistance. Int. J. Med. Microbiol. 2013, 303, 298–304. [Google Scholar] [CrossRef]
- Aworh, M.K.; Kwaga, J.K.P.; Hendriksen, R.S.; Okolocha, E.C.; Harrell, E.; Thakur, S. Quinolone-resistant Escherichia coli at the interface between humans, poultry and their shared environment—A potential public health risk. One Health Outlook 2023, 5, 2. [Google Scholar] [CrossRef]
- Rendón, M.A.; Saldaña, Z.; Erdem, A.L.; Monteiro-Neto, V.; Vázquez, A.; Kaper, J.B.; Puente, J.L.; Girón, J.A. Commensal and pathogenic Escherichia coli use a common pilus adherence factor for epithelial cell colonization. Proc. Natl. Acad. Sci. USA 2007, 104, 10637–10642. [Google Scholar] [CrossRef] [PubMed]
- Wells, T.J.; McNeilly, T.N.; Totsika, M.; Mahajan, A.; Gally, D.L.; Schembri, M.A. The Escherichia coli O157:H7 EhaB autotransporter protein binds to laminin and collagen I and induces a serum IgA response in O157:H7 challenged cattle. Environ. Microbiol. 2009, 11, 1803–1814. [Google Scholar] [CrossRef]
- Clarke, K.R.; Hor, L.; Pilapitiya, A.; Luirink, J.; Paxman, J.J.; Heras, B. Phylogenetic classification and functional review of autotransporters. Front. Immunol. 2022, 13, 921272. [Google Scholar] [CrossRef]
- Zhu, N.; Liu, W.; Prakash, A.; Zhang, C.; Kim, K.S. Targeting E. coli invasion of the blood-brain barrier for investigating the pathogenesis and therapeutic development of E. coli meningitis. Cell. Microbiol. 2020, 22, e13231. [Google Scholar] [CrossRef]
- Zhang, Y.; Tan, P.; Zhao, Y.; Ma, X. Enterotoxigenic Escherichia coli: Intestinal pathogenesis mechanisms and colonization resistance by gut microbiota. Gut Microbes 2022, 14, 2055943. [Google Scholar] [CrossRef]
- de Pace, F.; Nakazato, G.; Pacheco, A.; de Paiva, J.B.; Sperandio, V.; da Silveira, W.D. The type VI secretion system plays a role in type 1 fimbria expression and pathogenesis of an avian pathogenic Escherichia coli strain. Infect. Immun. 2010, 78, 4990–4998. [Google Scholar] [CrossRef] [PubMed]
- Quainoo, S.; Coolen, J.P.M.; van Hijum, S.; Huynen, M.A.; Melchers, W.J.G.; van Schaik, W.; Wertheim, H.F. Whole-genome sequencing of bacterial pathogens: The future of nosocomial outbreak analysis. Clin. Microbiol. Rev. 2017, 30, 1015–1063. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).