
Alternatives Integrating Omics Approaches for the Advancement of Human Skin Models: A Focus on Metagenomics, Metatranscriptomics, and Metaproteomics

 ,
,  , , and
, , and 
Abstract
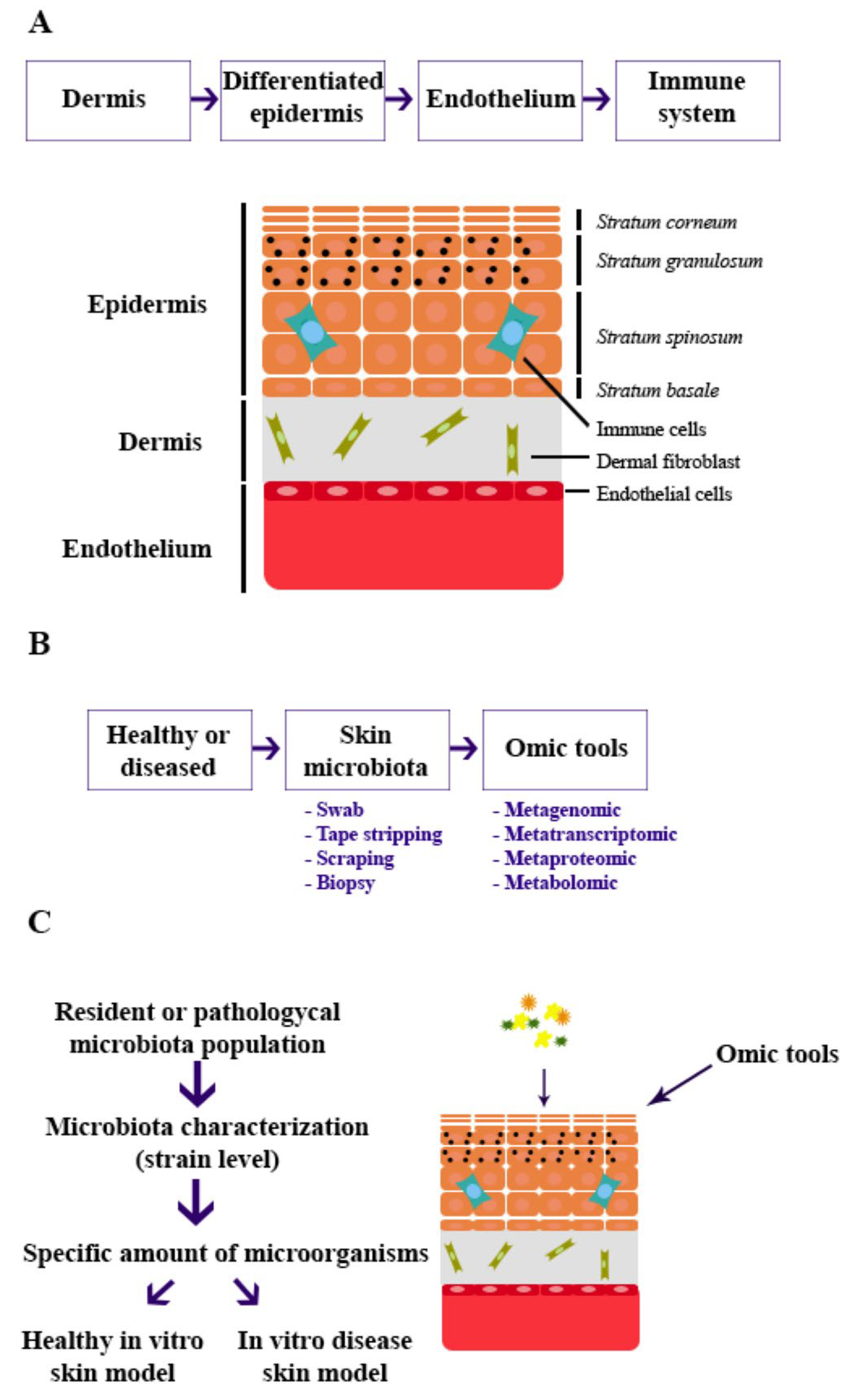
1. Introduction
2. Metagenomics
3. Metatranscriptomics
4. Metaproteomics
5. Metabolomics
6. Building In Vitro Skin Models
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Smythe, P.; Wilkinson, H.N. The Skin Microbiome: Current Landscape and Future Opportunities. Int. J. Mol. Sci. 2023, 24, 3950. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Chiang, H.-I.; Jiang, S.B.; Nagarajan, H.; Zengler, K.; Gallo, R.L. The Microbiome Extends to Subepidermal Compartments of Normal Skin. Nat. Commun. 2013, 4, 1431. [Google Scholar] [CrossRef]
- Grice, E.A.; Segre, J.A. The Skin Microbiome. Nat. Rev. Microbiol. 2011, 9, 244–253. [Google Scholar] [CrossRef]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The Human Skin Microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery Mode Shapes the Acquisition and Structure of the Initial Microbiota across Multiple Body Habitats in Newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef]
- Chu, D.M.; Ma, J.; Prince, A.L.; Antony, K.M.; Seferovic, M.D.; Aagaard, K.M. Maturation of the Infant Microbiome Community Structure and Function across Multiple Body Sites and in Relation to Mode of Delivery. Nat. Med. 2017, 23, 314–326. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Conlan, S.; Polley, E.C.; Segre, J.A.; Kong, H.H. Shifts in Human Skin and Nares Microbiota of Healthy Children and Adults. Genome Med. 2012, 4, 77. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Byrd, A.L.; Park, M.; NISC Comparative Sequencing Program; Kong, H.H.; Segre, J.A. Temporal Stability of the Human Skin Microbiome. Cell 2016, 165, 854–866. [Google Scholar] [CrossRef]
- Grice, E.A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; NISC Comparative Sequencing Program; Bouffard, G.G.; Blakesley, R.W.; Murray, P.R.; et al. Topographical and Temporal Diversity of the Human Skin Microbiome. Science 2009, 324, 1190–1192. [Google Scholar] [CrossRef] [PubMed]
- Celoria, V.; Rosset, F.; Pala, V.; Dapavo, P.; Ribero, S.; Quaglino, P.; Mastorino, L. The Skin Microbiome and Its Role in Psoriasis: A Review. Psoriasis 2023, 13, 71–78. [Google Scholar] [CrossRef]
- Martins, A.M.; Ascenso, A.; Ribeiro, H.M.; Marto, J. The Brain-Skin Connection and the Pathogenesis of Psoriasis: A Review with a Focus on the Serotonergic System. Cells 2020, 9, 796. [Google Scholar] [CrossRef] [PubMed]
- Adalsteinsson, J.A.; Kaushik, S.; Muzumdar, S.; Guttman-Yassky, E.; Ungar, J. An Update on the Microbiology, Immunology and Genetics of Seborrheic Dermatitis. Exp. Dermatol. 2020, 29, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Sowell, J.; Pena, S.M.; Elewski, B.E. Seborrheic Dermatitis in Older Adults: Pathogenesis and Treatment Options. Drugs Aging 2022, 39, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, P.; Farina, S.; Cristofolini, M.; Girolomoni, G.; Tett, A.; Segata, N. Experimental Metagenomics and Ribosomal Profiling of the Human Skin Microbiome. Exp. Dermatol. 2017, 26, 211–219. [Google Scholar] [CrossRef]
- Sandhu, S.S.; Pourang, A.; Sivamani, R.K. A Review of next Generation Sequencing Technologies Used in the Evaluation of the Skin Microbiome: What a Time to Be Alive. Dermatol. Online J. 2019, 25. [Google Scholar] [CrossRef]
- Roux, P.-F.; Oddos, T.; Stamatas, G. Deciphering the Role of Skin Surface Microbiome in Skin Health: An Integrative Multiomics Approach Reveals Three Distinct Metabolite‒Microbe Clusters. JID 2022, 142, 469–479.e5. [Google Scholar] [CrossRef]
- Rusiñol, L.; Puig, L. Multi-Omics Approach to Improved Diagnosis and Treatment of Atopic Dermatitis and Psoriasis. Int. J. Mol. Sci. 2024, 25, 1042. [Google Scholar] [CrossRef]
- Guo, Y.; Luo, L.; Zhu, J.; Li, C. Multi-Omics Research Strategies for Psoriasis and Atopic Dermatitis. Int. J. Mol. Sci. 2023, 24, 8018. [Google Scholar] [CrossRef]
- Handelsman, J. Metagenomics: Application of Genomics to Uncultured Microorganisms. Microbiol. Mol. Biol. Rev. 2004, 68, 669–685. [Google Scholar] [CrossRef]
- Meisel, J.S.; Hannigan, G.D.; Tyldsley, A.S.; SanMiguel, A.J.; Hodkinson, B.P.; Zheng, Q.; Grice, E.A. Skin Microbiome Surveys Are Strongly Influenced by Experimental Design. JID 2016, 136, 947–956. [Google Scholar] [CrossRef]
- Kong, H.H.; Segre, J.A. The Molecular Revolution in Cutaneous Biology: Investigating the Skin Microbiome. JID 2017, 137, e119–e122. [Google Scholar] [CrossRef]
- De Filippis, F.; Laiola, M.; Blaiotta, G.; Ercolini, D. Different Amplicon Targets for Sequencing-Based Studies of Fungal Diversity. Appl. Environ. Microbiol. 2017, 83, e00905-17. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.-H.; Kennedy, E.A.; Kong, H.H. Research Techniques Made Simple: Bacterial 16S Ribosomal RNA Gene Sequencing in Cutaneous Research. JID 2016, 136, e23–e27. [Google Scholar] [CrossRef]
- Jovel, J.; Patterson, J.; Wang, W.; Hotte, N.; O’Keefe, S.; Mitchel, T.; Perry, T.; Kao, D.; Mason, A.L.; Madsen, K.L.; et al. Characterization of the Gut Microbiome Using 16S or Shotgun Metagenomics. Front. Microbiol. 2016, 7, 459. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, R.; Rani, A.; Metwally, A.; McGee, H.S.; Perkins, D.L. Analysis of the Microbiome: Advantages of Whole Genome Shotgun versus 16S Amplicon Sequencing. Biochem. Biophys. Res. Commun. 2016, 469, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Bashiardes, S.; Zilberman-Schapira, G.; Elinav, E. Use of Metatranscriptomics in Microbiome Research. Bioinform. Biol. Insights 2016, 10, BBI.S34610. [Google Scholar] [CrossRef]
- Mathieu, A.; Vogel, T.M.; Simonet, P. The Future of Skin Metagenomics. Res. Microbiol. 2014, 165, 69–76. [Google Scholar] [CrossRef]
- Gao, Z.; Tseng, C.; Strober, B.E.; Pei, Z.; Blaser, M.J. Substantial Alterations of the Cutaneous Bacterial Biota in Psoriatic Lesions. PLoS ONE 2008, 3, e2719. [Google Scholar] [CrossRef]
- Van Den Bossche, T.; Verschaffelt, P.; Moortele, T.V.; Dawyndt, P.; Martens, L.; Mesuere, B. Biodiversity Analysis of Metaproteomics Samples with Unipept: A Comprehensive Tutorial. In Protein Bioinformatics; Humana: New York, NY, USA, 2023. [Google Scholar]
- Kelly, D.; Yang, L.; Pei, Z. A Review of the Oesophageal Microbiome in Health and Disease. In Methods in Microbiology; Academic Press Inc.: Cambridge, MA, USA, 2017; pp. 19–35. [Google Scholar]
- Franzosa, E.A.; Hsu, T.; Sirota-Madi, A.; Shafquat, A.; Abu-Ali, G.; Morgan, X.C.; Huttenhower, C. Sequencing and beyond: Integrating Molecular “omics” for Microbial Community Profiling. Nat. Rev. Microbiol. 2015, 13, 360–372. [Google Scholar] [CrossRef]
- Benítez-Páez, A.; Belda-Ferre, P.; Simón-Soro, A.; Mira, A. Microbiota Diversity and Gene Expression Dynamics in Human Oral Biofilms. BMC Genom. 2014, 15, 311. [Google Scholar] [CrossRef]
- Wilmes, P.; Heintz-Buschart, A.; Bond, P.L. A Decade of Metaproteomics: Where We Stand and What the Future Holds. Proteomics 2015, 15, 3409–3417. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, W.; Ning, Z.; Mayne, J.; Mack, D.; Stintzi, A.; Tian, R.; Figeys, D. Deep Metaproteomics Approach for the Study of Human Microbiomes. Anal. Chem. 2017, 89, 9407–9415. [Google Scholar] [CrossRef]
- Heyer, R.; Schallert, K.; Zoun, R.; Becher, B.; Saake, G.; Benndorf, D. Challenges and Perspectives of Metaproteomic Data Analysis. J. Biotechnol. 2017, 261, 24–36. [Google Scholar] [CrossRef]
- Van Den Bossche, T.; Arntzen, M.Ø.; Becher, D.; Benndorf, D.; Eijsink, V.G.H.; Henry, C.; Jagtap, P.D.; Jehmlich, N.; Juste, C.; Kunath, B.J.; et al. The Metaproteomics Initiative: A Coordinated Approach for Propelling the Functional Characterization of Microbiomes. Microbiome 2021, 9, 243. [Google Scholar] [CrossRef]
- Armengaud, J. Metaproteomics to Understand How Microbiota Function: The Crystal Ball Predicts a Promising Future. Environ. Microbiol. 2023, 25, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Heintz-Buschart, A.; Wilmes, P. Human Gut Microbiome: Function Matters. Trends Microbiol. 2018, 26, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Grenga, L.; Pible, O.; Miotello, G.; Culotta, K.; Ruat, S.; Roncato, M.; Gas, F.; Bellanger, L.; Claret, P.; Dunyach-Remy, C.; et al. Taxonomical and Functional Changes in COVID-19 Faecal Microbiome Could Be Related to SARS-CoV-2 Faecal Load. Environ. Microbiol. 2022, 24, 4299–4316. [Google Scholar] [CrossRef]
- Cheng, L.; Qi, C.; Yang, H.; Lu, M.; Cai, Y.; Fu, T.; Ren, J.; Jin, Q.; Zhang, X. GutMGene: A Comprehensive Database for Target Genes of Gut Microbes and Microbial Metabolites. Nucleic Acids Res. 2022, 50, D795–D800. [Google Scholar] [CrossRef]
- Xiong, W.; Abraham, P.E.; Li, Z.; Pan, C.; Hettich, R.L. Microbial Metaproteomics for Characterizing the Range of Metabolic Functions and Activities of Human Gut Microbiota. Proteomics 2015, 15, 3424–3438. [Google Scholar] [CrossRef]
- Xiao, M.; Yang, J.; Feng, Y.; Zhu, Y.; Chai, X.; Wang, Y. Metaproteomic Strategies and Applications for Gut Microbial Research. Appl. Microbiol. Biotechnol. 2017, 101, 3077–3088. [Google Scholar] [CrossRef] [PubMed]
- Aguiar-Pulido, V.; Huang, W.; Suarez-Ulloa, V.; Cickovski, T.; Mathee, K.; Narasimhan, G. Metagenomics, Metatranscriptomics, and Metabolomics Approaches for Microbiome Analysis. Evol. Bioinform. 2016, 12s1, EBO.S36436. [Google Scholar] [CrossRef]
- Kuehne, A.; Hildebrand, J.; Soehle, J.; Wenck, H.; Terstegen, L.; Gallinat, S.; Knott, A.; Winnefeld, M.; Zamboni, N. An Integrative Metabolomics and Transcriptomics Study to Identify Metabolic Alterations in Aged Skin of Humans in Vivo. BMC Genom. 2017, 18, 169. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Wen, B.; Hou, G.; Lei, L.; Mei, Z.; Jia, X.; Chen, X.; Zhu, W.; Li, J.; Kuang, Y.; et al. Lipidomics Profiling Reveals the Role of Glycerophospholipid Metabolism in Psoriasis. Gigascience 2017, 6, 1–11. [Google Scholar] [CrossRef]
- Xiong, Q.; Zhong, D.; Li, Q.; Yu, Y.; Zhang, S.; Liang, J.; Zhang, X. LC–MS Metabolomics Reveal Skin Metabolic Signature of Psoriasis Vulgaris. Exp. Dermatol. 2023, 32, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Kalan, L.R.; Meisel, J.S.; Loesche, M.A.; Horwinski, J.; Soaita, I.; Chen, X.; Uberoi, A.; Gardner, S.E.; Grice, E.A. Strain- and Species-Level Variation in the Microbiome of Diabetic Wounds Is Associated with Clinical Outcomes and Therapeutic Efficacy. Cell Host Microbe 2019, 25, 641–655.e5. [Google Scholar] [CrossRef] [PubMed]
- Teertam, S.K.; Setaluri, V.; Ayuso, J.M. Advances in Microengineered Platforms for Skin Research. JID Innov. 2025, 5, 100315. [Google Scholar] [CrossRef]
- Barros, N.R.; Kang, R.; Kim, J.; Ermis, M.; Kim, H.-J.; Dokmeci, M.R.; Lee, J. A Human Skin-on-a-Chip Platform for Microneedling-Driven Skin Cancer Treatment. Mater. Today Bio 2025, 30, 101399. [Google Scholar] [CrossRef]
- Ko, B.; Son, J.; In Won, J.; Kang, B.M.; Choi, C.W.; Kim, R.; Sung, J.H. Gut Microbe–Skin Axis on a Chip for Reproducing the Inflammatory Crosstalk. Lab Chip 2025, 25, 2609–2619. [Google Scholar] [CrossRef]
- Holland, D.B.; Bojar, R.A.; Jeremy, A.H.T.; Ingham, E.; Holland, K.T. Microbial Colonization of an in Vitro Model of a Tissue Engineered Human Skin Equivalent--a Novel Approach. FEMS Microbiol. Lett. 2008, 279, 110–115. [Google Scholar] [CrossRef]
- Laclaverie, M.; Rouaud-Tinguely, P.; Grimaldi, C.; Jugé, R.; Marchand, L.; Aymard, E.; Closs, B. Development and Characterization of a 3D in Vitro Model Mimicking Acneic Skin. Exp. Dermatol. 2021, 30, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Kanwar, I.L.; Haider, T.; Kumari, A.; Dubey, S.; Jain, P.; Soni, V. Models for Acne: A Comprehensive Study. Drug Discov. Ther. 2018, 12, 329–340. [Google Scholar] [CrossRef]
- Fitz-Gibbon, S.; Tomida, S.; Chiu, B.-H.; Nguyen, L.; Du, C.; Liu, M.; Elashoff, D.; Erfe, M.C.; Loncaric, A.; Kim, J.; et al. Propionibacterium Acnes Strain Populations in the Human Skin Microbiome Associated with Acne. J. Investig. Dermatol. 2013, 133, 2152–2160. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, C.C.; Eady, A.; Philpott, M.; Goldsmith, L.A.; Orfanos, C.; Cunliffe, W.C.; Rosenfield, R. What Is the Pathogenesis of Acne? Exp. Dermatol. 2005, 14, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Byrd, A.L.; Deming, C.; Cassidy, S.K.B.; Harrison, O.J.; Ng, W.-I.; Conlan, S.; NISC Comparative Sequencing Program; Belkaid, Y.; Segre, J.A.; Kong, H.H. Staphylococcus Aureus and Staphylococcus Epidermidis Strain Diversity Underlying Pediatric Atopic Dermatitis. Sci. Transl. Med. 2017, 9, eaal4651. [Google Scholar] [CrossRef]
- Ahn, K.; Kim, B.E.; Kim, J.; Leung, D.Y. Recent Advances in Atopic Dermatitis. Curr. Opin. Immunol. 2020, 66, 14–21. [Google Scholar] [CrossRef]
- Wu, X.; Zhao, Y.; Gu, Y.; Li, K.; Wang, X.; Zhang, J. Interferon-Lambda 1 Inhibits Staphylococcus Aureus Colonization in Human Primary Keratinocytes. Front. Pharmacol. 2021, 12, 652302. [Google Scholar] [CrossRef]
- Jung, S.-Y.; You, H.J.; Kim, M.-J.; Ko, G.; Lee, S.; Kang, K.-S. Wnt-Activating Human Skin Organoid Model of Atopic Dermatitis Induced by Staphylococcus Aureus and Its Protective Effects by Cutibacterium Acnes. iScience 2022, 25, 105150. [Google Scholar] [CrossRef]
- Schwartz, J.R.; Messenger, A.G.; Tosti, A.; Todd, G.; Hordinsky, M.; Hay, R.J.; Wang, X.; Zachariae, C.; Kerr, K.M.; Henry, J.P.; et al. A Comprehensive Pathophysiology of Dandruff and Seborrheic Dermatitis—Towards a More Precise Definition of Scalp Health. Acta Derm. Venereol. 2013, 93, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, R.A.; Janusz, C.A.; Janniger, C.K. Seborrheic Dermatitis: An Overview. Am. Fam. Physician 2006, 74, 125–130. [Google Scholar]
- Dawson, T.L. Malassezia Globosa and Restricta: Breakthrough Understanding of the Etiology and Treatment of Dandruff and Seborrheic Dermatitis through Whole-Genome Analysis. J. Investig. Dermatol. Symp. Proc. 2007, 12, 15–19. [Google Scholar] [CrossRef]
- Donnarumma, G.; Perfetto, B.; Paoletti, I.; Oliviero, G.; Clavaud, C.; Del Bufalo, A.; Guéniche, A.; Jourdain, R.; Tufano, M.A.; Breton, L. Analysis of the Response of Human Keratinocytes to Malassezia Globosa and Restricta Strains. Arch. Dermatol. Res. 2014, 306, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Meloni, M.; Balzaretti, S.; Collard, N.; Desaint, S.; Laperdrix, C. Reproducing the Scalp Microbiota Community: Co-Colonization of a 3D Reconstructed Human Epidermis with C. Acnes and M. Restricta. Int. J. Cosmet. Sci. 2021, 43, 235–245. [Google Scholar] [CrossRef]
- Cui, M.; Wiraja, C.; Zheng, M.; Singh, G.; Yong, K.; Xu, C. Recent Progress in Skin-on-a-Chip Platforms. Adv. Ther. 2022, 5, 2100138. [Google Scholar] [CrossRef]
- Fernandez-Carro, E.; Remacha, A.R.; Orera, I.; Lattanzio, G.; Garcia-Barrios, A.; del Barrio, J.; Alcaine, C.; Ciriza, J. Human Dermal Decellularized ECM Hydrogels as Scaffolds for 3D In Vitro Skin Aging Models. Int. J. Mol. Sci. 2024, 25, 4020. [Google Scholar] [CrossRef]
- Risueño, I.; Valencia, L.; Jorcano, J.L.; Velasco, D. Skin-on-a-Chip Models: General Overview and Future Perspectives. APL Bioeng. 2021, 5, 030901. [Google Scholar] [CrossRef]
- Pontiggia, L.; Klar, A.S.; Michalak-Micka, K.; Moehrlen, U.; Biedermann, T. Isolation, Characterization, and Utilization of Human Skin Basal and Suprabasal Epidermal Stem Cells. In Skin Stem Cells; Humana: New York, NY, USA, 2024; pp. 1–15. [Google Scholar] [CrossRef]
- Kwak, B.S.; Jin, S.; Kim, S.J.; Kim, E.J.; Chung, J.H.; Sung, J.H. Microfluidic Skin Chip with Vasculature for Recapitulating the Immune Response of the Skin Tissue. Biotechnol. Bioeng. 2020, 117, 1853–1863. [Google Scholar] [CrossRef]
- Ogai, K.; Nagase, S.; Mukai, K.; Iuchi, T.; Mori, Y.; Matsue, M.; Sugitani, K.; Sugama, J.; Okamoto, S. A Comparison of Techniques for Collecting Skin Microbiome Samples: Swabbing Versus Tape-Stripping. Front. Microbiol. 2018, 9, 2362. [Google Scholar] [CrossRef] [PubMed]
- Gotschlich, E.C.; Colbert, R.A.; Gill, T. Methods in Microbiome Research: Past, Present, and Future. Best. Pract. Res. Clin. Rheumatol. 2019, 33, 101498. [Google Scholar] [CrossRef]
- Ederveen, T.H.A.; Smits, J.P.H.; Boekhorst, J.; Schalkwijk, J.; van den Bogaard, E.H.; Zeeuwen, P.L.J.M. Skin Microbiota in Health and Disease: From Sequencing to Biology. J. Dermatol. 2020, 47, 1110–1118. [Google Scholar] [CrossRef] [PubMed]
- Dariush Gholami, Z.E.A.R.N.S.A. Advances in Bacterial Identification and Characterization: Methods and Applications. Microbiol. Metab. Biotechnol. 2019, 2, 119–136. [Google Scholar]
- van den Bogaard, E.; Ilic, D.; Dubrac, S.; Tomic-Canic, M.; Bouwstra, J.; Celli, A.; Mauro, T. Perspective and Consensus Opinion: Good Practices for Using Organotypic Skin and Epidermal Equivalents in Experimental Dermatology Research. JID 2021, 141, 203–205. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Collecting Area | Advantages | Disadvantages | |
|---|---|---|---|
| Swab | Surface and epidermal layers | Easy No invasive Commercially available Standardized | Often contain small bacterial yields |
| Tape stripping | Deep epidermal layers | Easy Commercially available More cultivable bacteria collected | Different adhesives Less efficient in oily, wet, or undulating skin |
| Scraping | Deep epidermal layers | Easy | Invasive Mechanical scraping damages the skin. DNA contamination (host DNA) |
| Biopsy | Epidermis and dermis | Most representative skin-microbiota detection | Invasive DNA contamination (host DNA) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Carro, E.; Letsiou, S.; Tsironi, S.; Chaniotis, D.; Ciriza, J.; Beloukas, A. Alternatives Integrating Omics Approaches for the Advancement of Human Skin Models: A Focus on Metagenomics, Metatranscriptomics, and Metaproteomics. Microorganisms 2025, 13, 1771. https://doi.org/10.3390/microorganisms13081771
Fernández-Carro E, Letsiou S, Tsironi S, Chaniotis D, Ciriza J, Beloukas A. Alternatives Integrating Omics Approaches for the Advancement of Human Skin Models: A Focus on Metagenomics, Metatranscriptomics, and Metaproteomics. Microorganisms. 2025; 13(8):1771. https://doi.org/10.3390/microorganisms13081771
Chicago/Turabian StyleFernández-Carro, Estibaliz, Sophia Letsiou, Stella Tsironi, Dimitrios Chaniotis, Jesús Ciriza, and Apostolos Beloukas. 2025. "Alternatives Integrating Omics Approaches for the Advancement of Human Skin Models: A Focus on Metagenomics, Metatranscriptomics, and Metaproteomics" Microorganisms 13, no. 8: 1771. https://doi.org/10.3390/microorganisms13081771
APA StyleFernández-Carro, E., Letsiou, S., Tsironi, S., Chaniotis, D., Ciriza, J., & Beloukas, A. (2025). Alternatives Integrating Omics Approaches for the Advancement of Human Skin Models: A Focus on Metagenomics, Metatranscriptomics, and Metaproteomics. Microorganisms, 13(8), 1771. https://doi.org/10.3390/microorganisms13081771