
Unveiling Species Diversity Within Early-Diverging Fungi from China VI: Four Absidia sp. nov. (Mucorales) in Guizhou and Hainan


Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Morphology
2.2. DNA Extraction and Amplification
3. Results
3.1. Phylogenetic Analyses
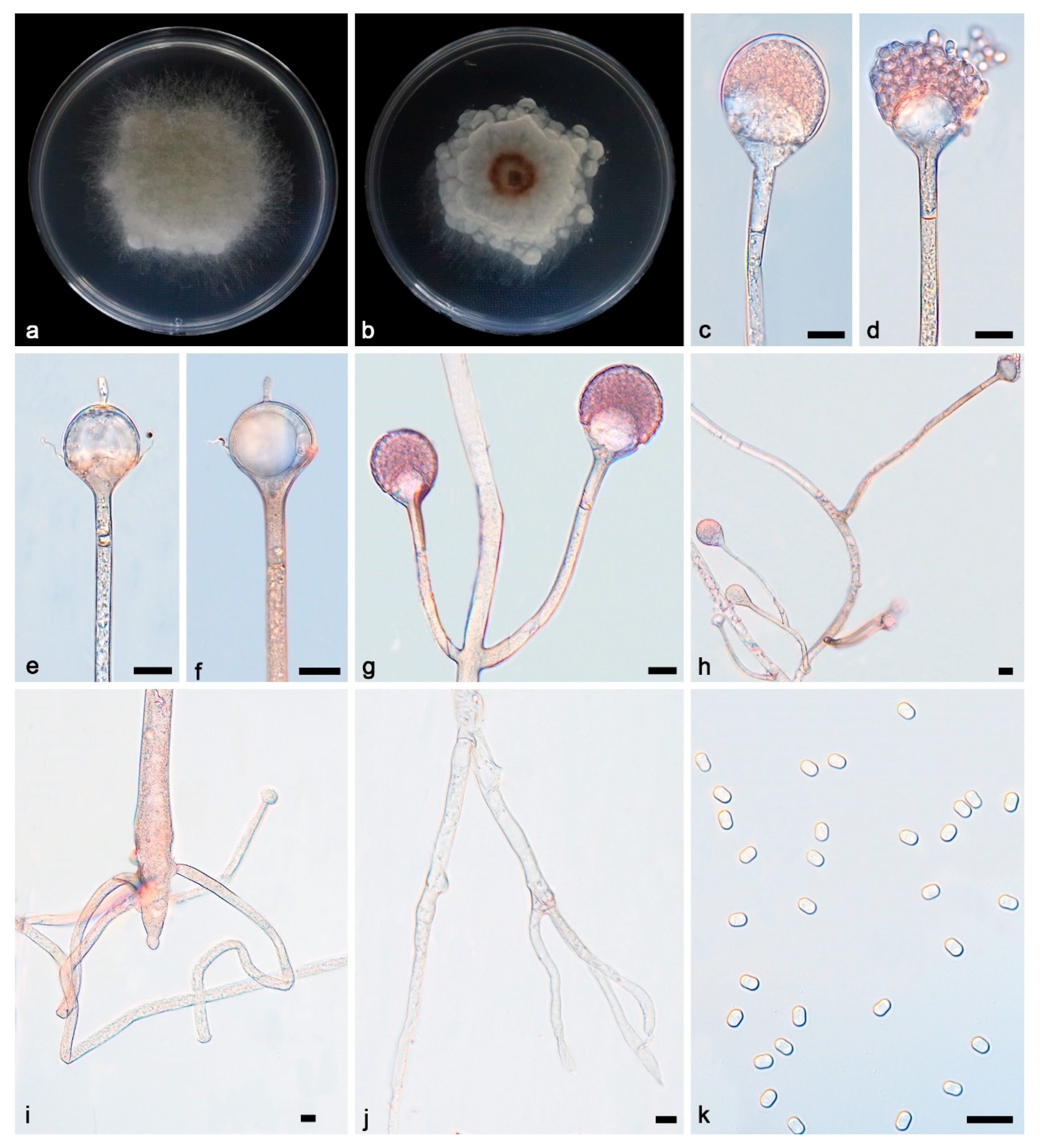
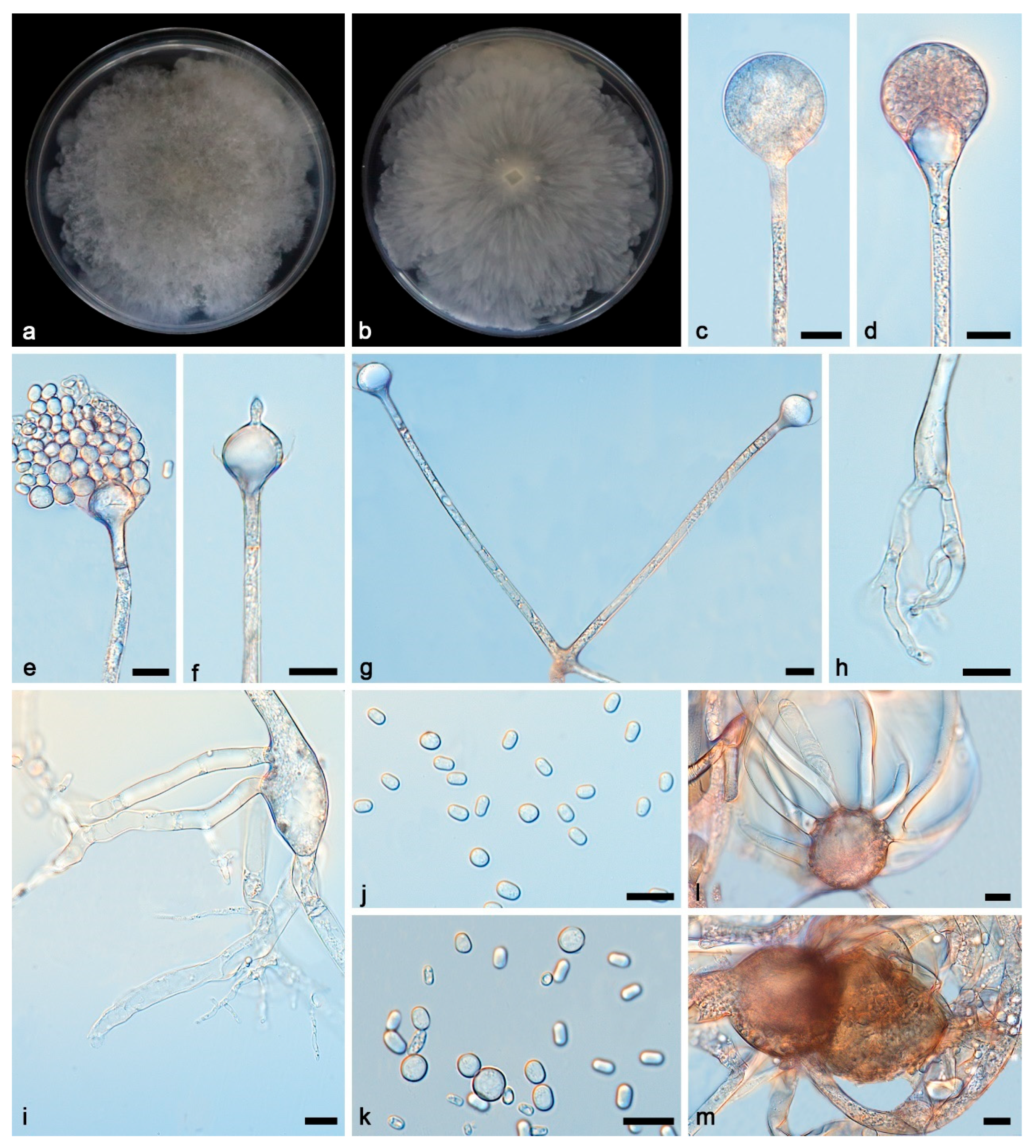
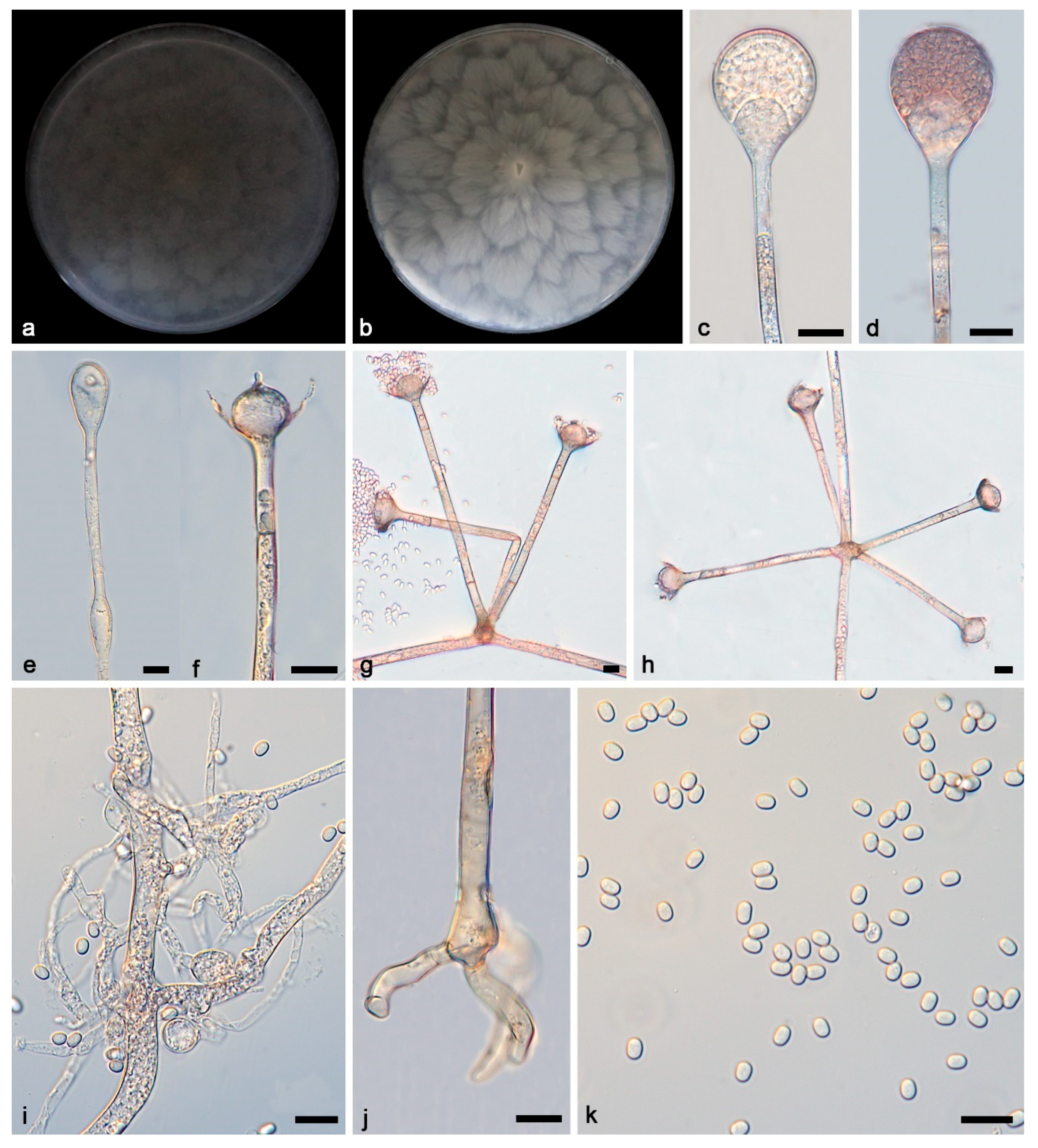
3.2. Taxonomy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- van Tieghem, P. Troisieme memoire sur les Mucorinees. Ann. Sci. Nat. Bot. Ser. 1876, 4, 312–398. [Google Scholar]
- Tedersoo, L.; Sánchez-Ramírez, S.; Kõljalg, U.; Bahram, M.; Döring, M.; Schigel, D.; May, T.; Ryberg, M.; Abarenkov, K. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Divers. 2018, 90, 135–159. [Google Scholar] [CrossRef]
- Von Arx, J.A. On Mucoraceae s. str. and other families of the Mucorales. Sydowia 1982, 35, 10–26. [Google Scholar]
- Benny, G.; Humber, R.; Morton, J. Zygomycota: Zygomycetes. In The Mycota. VII. Systematics and Evolution: Part A; McLaughlin, D.J., McLaughlin, E.G., Lemke, P.A., Eds.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 113–146. [Google Scholar]
- Kirk, P.; Cannon, P.; Minter, D.; Stalpers, J. Ainsworth and Bisby’s Dictionary of the Fungi (10th edition). Ref. Rev. 2009, 23, 42. [Google Scholar]
- Hoffmann, K. Identification of the genus Absidia (Mucorales, Zygomycetes): A comprehensive taxonomic revision. In Molecular Identification of Fungi; Gherbawy, Y., Voigt, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 439–460. [Google Scholar]
- Hoffmann, K.; Discher, S.; Voigt, K. Revision of the genus Absidia (Mucorales, Zygomycetes) based on physiological, phylogenetic, and morphological characters; thermotolerant Absidia spp. form a coherent group, Mycocladiaceae fam. nov. Mycol. Res. 2007, 111, 1169–1183. [Google Scholar] [CrossRef]
- Hoffmann, K.; Voigt, K. Absidia parricida plays a dominant role in biotrophic fusion parasitism among mucoralean fungi (Zygomycetes): Lentamyces, a new genus for A. parricida and A. zychae. Plant Biol. 2009, 11, 537–554. [Google Scholar] [CrossRef]
- Zhao, H.; Nie, Y.; Tongkai, Z.; Wang, Y.-J.; Wang, M.; Dai, Y.-C.; Liu, X.-Y. Species diversity and ecological habitat of Absidia (Cunninghamellaceae, Mucorales) with emphasis on five new species from forest and grassland soil in China. J. Fungi 2022, 8, 471. [Google Scholar] [CrossRef]
- Zhao, H.; Nie, Y.; Zong, T.; Dai, Y.; Liu, X. Three new species of Absidia (Mucoromycota) from China based on phylogeny, morphology and physiology. Diversity 2022, 14, 132. [Google Scholar] [CrossRef]
- Braga, A.R.; Nunes, M.C.; Raymundo, A. The experimental development of emulsions enriched and stabilized by recovering matter from spirulina biomass: Valorization of residue into a sustainable protein source. Molecules 2023, 28, 6179. [Google Scholar] [CrossRef]
- Liu, X.; Qiao, L.; Xie, D.; Zhang, Y.; Zou, J.; Chen, X.; Dai, J. Microbial transformation of ginsenoside-Rg1 by Absidia coerulea and the reversal activity of the metabolites towards multi-drug resistant tumor cells. Fitoterapia 2011, 82, 1313–1317. [Google Scholar] [CrossRef]
- Tao, M.-F.; Ding, Z.-Y.; Wang, Y.-X.; Zhang, Z.-X.; Zhao, H.; Meng, Z.; Liu, X.-Y. Unveiling species diversity within early-diverging fungi from China II: Three new species of Absidia (Cunninghamellaceae, Mucoromycota) from Hainan Province. MycoKeys 2024, 110, 255–272. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-X.; Zhao, H.; Jiang, Y.; Liu, X.-Y.; Tao, M.-F.; Liu, X.-Y. Unveiling species diversity within early-diverging fungi from China III: Six new species and a new record of Gongronella (Cunninghamellaceae, Mucoromycota). MycoKeys 2024, 110, 287–317. [Google Scholar] [CrossRef]
- Zhao, H.; Nie, Y.; Huang, B.; Liu, X.-Y. Unveiling species diversity within early-diverging fungi from China I: Three new species of Backusella (Backusellaceae, Mucoromycota). MycoKeys 2024, 109, 285–304. [Google Scholar] [CrossRef]
- Ding, Z.-Y.; Ji, X.-Y.; Tao, M.-F.; Jiang, Y.; Liu, W.-X.; Wang, Y.-X.; Meng, Z.; Liu, X.-Y. Unveiling species diversity within early-diverging fungi from China IV: Four new species of Absidia (Cunninghamellaceae, Mucoromycota). MycoKeys 2025, 117, 267. [Google Scholar]
- Ji, X.-Y.; Ding, Z.-Y.; Nie, Y.; Zhao, H.; Wang, S.; Huang, B.; Liu, X.-Y. Unveiling species diversity within early-diverging fungi from China V: Five new species of Absidia (Cunninghamellaceae, Mucoromycota). MycoKeys 2025, 117, 267–288. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wei, Y.; Zou, Y.; Liu, P.; Li, Z.; Gontcharov, A.; Stephenson, S.; Wang, Q.; Zhang, S.; Li, Y. Dictyostelid cellular slime molds from the russian far east. Protist 2020, 171, 125756. [Google Scholar] [CrossRef]
- Zou, Y.; Hou, J.; Guo, S.; Li, C.; Li, Z.; Stephenson, S.L.; Pavlov, I.N.; Liu, P.; Li, Y. Diversity of dictyostelid cellular slime molds, including two species new to science, in forest soils of Changbai Mountain, China. Microbiol. Spectr. 2022, 10, e0240222. [Google Scholar] [CrossRef]
- Corry, J.E.L.; Curtis, G.D.W.; Baird, R.M. Rose Bengal Chloramphenicol (RBC) agar. Prog. Ind. Microbiol. 1995, 34, 431–433. [Google Scholar]
- Wang, Y.; Zhao, H.; Ding, Z.; Ji, X.; Zhang, Z.; Wang, S.; Zhang, X.; Liu, X. Three new species of Gongronella (Cunninghamellaceae, Mucorales) from soil in Hainan, China based on morphology and molecular phylogeny. J. Fungi 2023, 9, 1182. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, R.; Liu, S.; Mu, T.; Zhang, X.; Xia, J. Morphological and phylogenetic analyses reveal two new species of Sporocadaceae from Hainan, China. MycoKeys 2022, 88, 171–192. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Voigt, K.; Wostemeyer, J. Reliable amplification of actin genes facilitates deep-level phylogeny. Microbiol. Res. 2000, 155, 179–195. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, J.; Innis, M.; Gelfand, D.; Sninsky, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics; PCR Protocols; Academic Press, Inc.: Cambridge, MA, USA, 1990; Volume 31, pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Lutzoni, F.; Ward, T.; Benny, G. Evolutionary relationships among mucoralean fungi (Zygomycota): Evidence for family polyphyly on a large scale. Mycologia 2001, 93, 286–296. [Google Scholar] [CrossRef]
- Nylander, J. MrModeltest V2. Program distributed by the author. Bioinformatics 2004, 24, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Mark, P.; Ayres, D.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.; Huelsenbeck, J. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Zhao, H.; Nie, Y.; Zong, T.-K.; Wang, K.; Lv, M.-L.; Cui, Y.-J.; Tohtirjap, A.; Chen, J.-J.; Zhao, C.-L.; Wu, F.; et al. Species diversity, updated classification and divergence times of the phylum Mucoromycota. Fungal Divers. 2023, 123, 49–157. [Google Scholar] [CrossRef]
- Zhang, Z.; Shang, Y.; Liu, Q.; Li, D.; Yin, C.; Liu, X.; Tao, M.; Jiang, Y.; Wang, Y.; Zhang, M.; et al. Deciphering the evolutionary and taxonomic complexity of Diaporthales (Sordariomycetes, Ascomycota) through integrated phylogenomic and divergence time estimation. Fungal Divers. 2025. [Google Scholar] [CrossRef]
- Hesseltine, C.W.; Ellis, J.J. The genus Absidia: Gongronella and cylindrical-spored species of Absidia. Mycologia 1964, 4, 568–601. [Google Scholar] [CrossRef]
- Zhang, T.-Y.; Yu, Y.; Zhu, H.; Yang, S.-Z.; Yang, T.-M.; Zhang, M.-y.; Zhang, Y.-X. Absidia panacisoli sp. nov., isolated from rhizosphere of Panax notoginseng. Int. J. Syst. Evol. Microbiol. 2018, 68, 2468–2472. [Google Scholar] [CrossRef] [PubMed]
- Hurdeal, V.G.; Gentekaki, E.; Lee, H.B.; Jeewon, R.; Hyde, K.D.; Tibpromma, S.; Mortimer, P.E.; Xu, J. Mucoralean fungi in Thailand: Novel species of Absidia from tropical forest soil. Cryptogam. Mycol. 2021, 42, 39–61. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Santiago, A.; Hallsworth, J.E.; Cordeiro, T.; Voigt, K.; Kirk, P.; Crous, P.W.; Júnior, M.A.M.; Elsztein, C.; Lee, H. New Mucorales from opposite ends of the world. Stud. Mycol. 2024, 109, 273–321. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Loci | PCR Primers | Sequence (5′–3′) | PCR Cycles | Reference |
|---|---|---|---|---|
| Act | ACT-512F | ATG TGC AAG GCC GGT TTC GC | 95 °C 3 min; (95 °C 1 min, 55 °C 1 min, 72 °C 1 min) × 30 cycles; 72 °C 10 min | [25] |
| ACT-783R | TAC GAG TCC TTC TGG CCC AT | |||
| ITS | ITS5 | GGA AGT AAA AGT CGT AAC AAG G | 95 °C 5 min; (95 °C 30 s, 55 °C 30 s, 72 °C 1 min) × 35 cycles; 72 °C 10 min | [26] |
| ITS4 | TCC TCC GCT TAT TGA TAT GC | |||
| LSU | LR0R | GTA CCC GCT GAA CTT AAG C | 95 °C 5 min; (95 °C 50 s, 47 °C 30 s, 72 °C 1.5 min) × 35 cycles; 72 °C 10 min | [27] |
| LR7 | TAC TAC CAC CAA GAT CT | |||
| SSU | NS1 | GTA GTC ATA TGC TTG TCT C C | 95 °C 5 min; (94 °C 60 s, 54 °C 50 s, 72 °C 60 s) × 37 cycles; 72 °C 10 min | [26] |
| NS4 | CTT CCG TCA ATT CCT TTA AG | |||
| TEF1α | EF1-728F | CAT CGA GAA GTT CGA GAA GG | 95 °C 5 min; (95 °C 30 s, 55 °C 60 s, 72 °C 1 min) × 30 cycles; 72 °C 10 min | [28] |
| EF2 | GGA RGT ACC AGT SAT CAT GTT |
| Species | Strains | GenBank Accession Numbers | ||||
|---|---|---|---|---|---|---|
| ITS | LSU | TEF1α | Act | SSU | ||
| Absidia abundans | CGMCC 3.16255 * | NR_182590 | ON074683 | n.a. | n.a. | n.a. |
| A. abundans | XY09274 | ON074696 | ON074682 | n.a. | n.a. | n.a. |
| A. aguabelensis | URM 8213 * | NR_189383 | NG_241934 | n.a. | n.a. | n.a. |
| A. alpina | CGMCC 3.16104 | OL678133 | n.a. | n.a. | n.a. | n.a. |
| A. ampullacea | CGMCC 3.16054 | MZ354138 | MZ350132 | n.a. | n.a. | n.a. |
| A. anomala | CBS 125.68 * | MH859085 | MH870799 | n.a. | n.a. | n.a. |
| A. anomala | FSU5798 | EF030523 | n.a. | n.a. | EF030535 | n.a. |
| A. biappendiculata | CBS 187.64 | MZ354153 | MZ350147 | MZ357420 | MZ357438 | |
| A. bonitoensis | URM 7889 * | MN977786 | MN977805 | n.a. | n.a. | n.a. |
| A. brunnea | CGMCC 3.16055 * | MZ354139 | MZ350133 | MZ357403 | MZ357421 | n.a. |
| A. caatinguensis | URM 7156 * | NR_154704 | NG_058582 | n.a. | n.a. | n.a. |
| A. caerulea | CBS101.36 * | MH855718 | MH867230 | n.a. | n.a. | n.a. |
| A. californica | CBS 314.78 | JN205816 | MH872902 | n.a. | n.a. | n.a. |
| A. chinensis | CGMCC 3.16057 | MZ354141 | MZ350135 | n.a. | MZ357422 | n.a. |
| A. chinensis | CGMCC 3.16056 * | MZ354140 | MZ350134 | n.a. | n.a. | n.a. |
| A. cinerea | CGMCC 3.16062 | MZ354146 | MZ350140 | MZ357407 | MZ357427 | n.a. |
| A. cornuta | URM 6100 * | NR_172976 | MN625255 | n.a. | n.a. | n.a. |
| A. crystalloides | CGMCC3.27496 * | PP377803 | PP373736 | PP790574 | PP790582 | PP779723 |
| A. crystalloides | SAUCC693201 | PP377804 | PP373737 | PP790573 | PP790581 | PP779722 |
| A. cuneospora | CBS 101.59 * | MH857828 | MH869361 | n.a. | n.a. | n.a. |
| A. cuneospora | FSU5890 | EF030524 | n.a. | n.a. | EF030533 | n.a. |
| A. cylindrospora | CBS 100.08 | JN205822 | JN206588 | n.a. | n.a. | n.a. |
| A. digitula | CGMCC 3.16058 * | MZ354142 | MZ350136 | MZ357404 | MZ357423 | n.a. |
| A. edaphica | MFLUCC 20-0088 | NR_172305 | NG_075367 | n.a. | MT410739 | NG_074951 |
| A. frigida | CGMCC 3.16201 * | NR_182565 | OM030223 | n.a. | n.a. | n.a. |
| A. fusca | CBS 102.35 * | NR_103625 | NG_058552 | n.a. | n.a. | n.a. |
| A. gemella | CGMCC 3.16202 * | OM108488 | OM030224 | n.a. | n.a. | n.a. |
| A. glauca | CBS 101.08 * | MH854573 | MH866105 | n.a. | n.a. | n.a. |
| A. glauca | FSU660 | AY944879 | EU736302 | EU736248 | EU736225 | EU736275 |
| A. globospora | CGMCC 3.16031 * | NR_189829 | MW671544 | MZ357412 | MZ357431 | n.a. |
| A. globospora | CGMCC 3.16036 | MW671539 | MW671546 | MZ357414 | MZ357433 | n.a. |
| A. healeyae | UoMAU1 | n.a. | MT436027 | n.a. | n.a. | n.a. |
| A. heterospora | SHTH021 | JN942683 | JN982936 | n.a. | n.a. | JQ004928 |
| A. irregularis | CGMCC 3.27812 * | PQ306325 | PQ289020 | PV126019 | PQ807209 | PQ799254 |
| A. irregularis | XG05674-7 | PQ306326 | PQ289021 | PV126020 | PQ807210 | PQ799255 |
| A. jiangxiensis | CGMCC 3.16105 * | OL678134 | PP780377 | n.a. | n.a. | n.a. |
| A. jindoensis | CNUFC-PTI1-1 | MF926622 | MF926616 | MF926513 | MF926510 | MF926626 |
| A. koreana | EML-IFS45-1 * | KR030062 | KR030056 | KR030060 | KR030058 | KT321298 |
| A. koreana | XY00816 | OL620083 | ON123771 | n.a. | n.a. | n.a. |
| A. lobata | CGMCC 3.16256 | ON074690 | ON074679 | n.a. | n.a. | n.a. |
| A. longissima | CGMCC 3.16203 * | NR_182566 | OM030225 | n.a. | n.a. | n.a. |
| A. macrospora | FSU4746 | AY944882 | EU736303 | EU736249 | AY944760 | EU736276 |
| A. medulla | CGMCC 3.16034 | NR_189832 | MW671549 | MZ357417 | MZ357436 | n.a. |
| A. montepascoalis | URM 8218 | NR_172995 | n.a. | n.a. | n.a. | n.a. |
| A. multiformis | CGMCC 3.27807 * | PQ306319 | PQ803168 | PV126021 | PQ807203 | PQ799260 |
| A. multiformis | XG04016-3 | PQ306320 | PQ803169 | PV126022 | PQ807204 | PQ799261 |
| A. multispora | URM 8210 * | MN953780 | MN953782 | n.a. | n.a. | n.a. |
| A. nigra | CBS 127.68 * | NR_173068 | MZ350146 | MZ357419 | MZ357437 | n.a. |
| A. nigra | CGMCC 3.16060 | MZ354144 | MZ350138 | MZ357406 | MZ357425 | n.a. |
| A. oblongispora | CGMCC 3.16061 | MZ354145 | MZ350139 | n.a. | MZ357426 | n.a. |
| A. ovalispora | CGMCC 3.16019 | NR_176748 | MW264131 | n.a. | n.a. | n.a. |
| A. ovoidospora | CGMCC 3.27811 clone1 * | PQ306327 | PQ803164 | PV126015 | PQ807207 | PQ799256 |
| A. ovoidospora | CGMCC 3.27811 clone2 * | PV069753 | PQ803165 | PV126017 | PV126023 | PQ799257 |
| A. ovoidospora | XG05673-3 clone1 | PQ306328 | PQ803166 | PV126016 | PQ807208 | PQ799258 |
| A. ovoidospora | XG05673-3 clone2 | PV069754 | PQ803167 | PV126018 | PV126024 | PQ799259 |
| A. pacifica | CGMCC3.27497 * | PP377802 | PP373735 | PP839793 | PP790579 | PP779720 |
| A. pacifica | SAUCC413601 | PP377801 | PP373734 | PP839794 | PP790580 | PP779721 |
| A. panacisoli | SYPF 7183 * | MF522181 | MF522180 | MF624251 | n.a. | MF522179 |
| A. pararepens | CCF 6352 | MT193669 | MT192308 | n.a. | n.a. | n.a. |
| A. pateriformis | CGMCC3.27495 * | PP377805 | PP373738 | PP790575 | PP790583 | PP779724 |
| A. pateriformis | SAUCC634702 | PP377806 | PP373739 | PP790576 | PP790584 | PP779725 |
| A. pernambucoensis | URM<BRA>7219 | MN635568 | MN635569 | n.a. | n.a. | n.a. |
| A. pseudocylindrospora | EML-FSDY6-2 | KU923817 | KU923814 | n.a. | KU923815 | KU923819 |
| A. pseudocylindrospora | CBS 100.62 * | NR_145276 | MH869688 | n.a. | n.a. | n.a. |
| A. psychrophilia | FSU4745 | AY944874 | EU736306 | EU736252 | AY944762 | EU736279 |
| A. radiata | CGMCC 3.16257 | ON074698 | ON074684 | n.a. | n.a. | n.a. |
| A. radiata | XY09330-1 | ON074699 | ON074685 | n.a. | n.a. | n.a. |
| A. repens | CBS 115583 * | NR_103624 | NG_058551 | n.a. | n.a. | n.a. |
| A. saloaensis | URM 8209 * | MN953781 | MN953783 | n.a. | n.a. | n.a. |
| A. sichuanensis | CGMCC 3.16258 * | NR_182589 | ON074688 | n.a. | n.a. | n.a. |
| A. soli | MFLU-20-0414 * | MT396373 | MT393988 | n.a. | n.a. | MT394049 |
| A. soli | MFLU 20-0413 | MT396371 | MT393985 | n.a. | n.a. | MT394046 |
| A. spinosa | FSU551 | AY944887 | EU736307 | EU736253 | EU736227 | EU736280 |
| A. stercoraria | EML-DG8-1 * | KU168828 | KT921998 | KT922002 | KT922000 | NG_065640 |
| A. sympodialis | CGMCC 3.16063 * | MZ354147 | MZ350141 | n.a. | n.a. | n.a. |
| A. sympodialis | CGMCC 3.16064 | MZ354148 | MZ350142 | MZ357408 | n.a. | n.a. |
| A. terrestris | FMR 14989 * | LT795003 | LT795005 | n.a. | n.a. | n.a. |
| A. turgida | CGMCC 3.16032 * | NR_189830 | NG_241931 | MZ357415 | MZ357434 | n.a. |
| A. varians | CGMCC 3.16065 * | MZ354149 | MZ350143 | MZ357409 | MZ357428 | n.a. |
| A. verticilliformis | CGMCC 3.27810 clone1 * | PQ306315 | PQ803170 | PV126011 | PQ807205 | PQ799262 |
| A. verticilliformis | CGMCC 3.27810 clone2 * | PV069755 | PQ803171 | PV126013 | PV126025 | PQ799263 |
| A. verticilliformis | XG04088-4 clone1 | PQ306316 | PQ803172 | PV126012 | PQ807206 | PQ799264 |
| A. verticilliformis | XG04088-4 clone2 | PV069756 | PQ803173 | PV126014 | PV126026 | PQ799265 |
| A. virescens | CGMCC 3.16066 * | MZ354150 | MZ350144 | MZ357410 | MZ357429 | n.a. |
| A. virescens | CGMCC 3.16067 | MZ354151 | MZ350145 | MZ357411 | MZ357430 | n.a. |
| A. xinjiangensis | CGMCC 3.16107 * | OL678136 | n.a. | n.a. | n.a. | n.a. |
| A. yunnanensis | XY09528 | ON074701 | ON074686 | n.a. | n.a. | n.a. |
| A. yunnanensis | CGMCC 3.16259 * | NR_182591 | NG_149054 | n.a. | n.a. | n.a. |
| A. zonata | CGMCC 3.16033 * | NR_189831 | MW671548 | MZ357416 | MZ357435 | n.a. |
| A. zygospora | RSPG 214 | KC478527 | n.a. | n.a. | n.a. | n.a. |
| A. zygospora | ANG28 | DQ914420 | n.a. | n.a. | n.a. | n.a. |
| Cunninghamella blakesleeana | CBS 782.68 | JN205869 | MH870950 | n.a. | n.a. | n.a. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-X.; Ding, Z.-Y.; Ji, X.-Y.; Meng, Z.; Liu, X.-Y. Unveiling Species Diversity Within Early-Diverging Fungi from China VI: Four Absidia sp. nov. (Mucorales) in Guizhou and Hainan. Microorganisms 2025, 13, 1315. https://doi.org/10.3390/microorganisms13061315
Wang Y-X, Ding Z-Y, Ji X-Y, Meng Z, Liu X-Y. Unveiling Species Diversity Within Early-Diverging Fungi from China VI: Four Absidia sp. nov. (Mucorales) in Guizhou and Hainan. Microorganisms. 2025; 13(6):1315. https://doi.org/10.3390/microorganisms13061315
Chicago/Turabian StyleWang, Yi-Xin, Zi-Ying Ding, Xin-Yu Ji, Zhe Meng, and Xiao-Yong Liu. 2025. "Unveiling Species Diversity Within Early-Diverging Fungi from China VI: Four Absidia sp. nov. (Mucorales) in Guizhou and Hainan" Microorganisms 13, no. 6: 1315. https://doi.org/10.3390/microorganisms13061315
APA StyleWang, Y.-X., Ding, Z.-Y., Ji, X.-Y., Meng, Z., & Liu, X.-Y. (2025). Unveiling Species Diversity Within Early-Diverging Fungi from China VI: Four Absidia sp. nov. (Mucorales) in Guizhou and Hainan. Microorganisms, 13(6), 1315. https://doi.org/10.3390/microorganisms13061315