
Effects of Hops Treatment on Nitrogen Retention, Volatile Fatty Acid Accumulations, and Select Microbial Populations of Composting Poultry Litter Intended for Use as a Ruminant Feedstuff

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Poultry Litter, Ground Forage and Hop Extracts
2.2. Design of Compost Simulations
2.3. Analytical
2.4. Statistics
3. Results
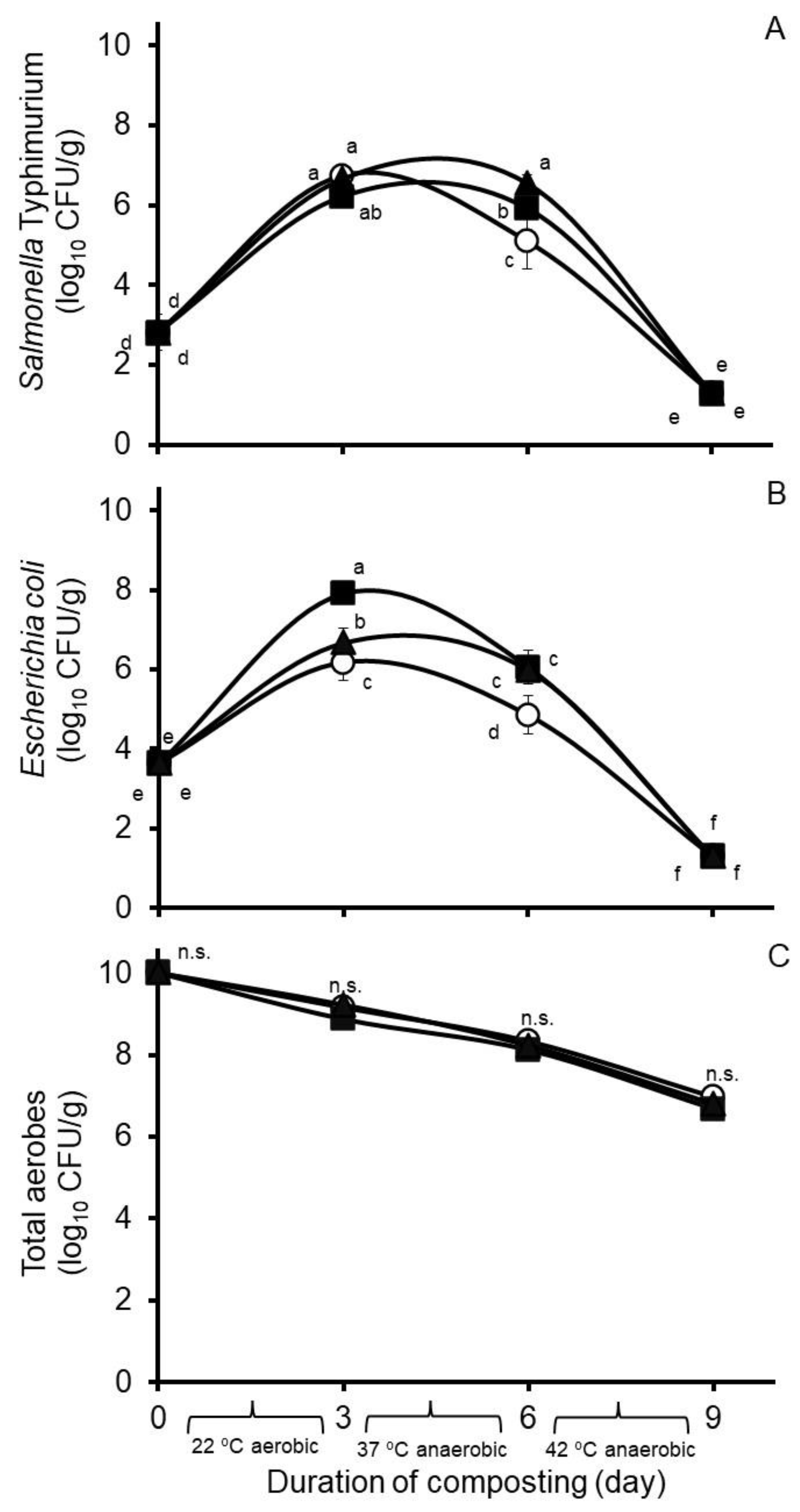
3.1. Experiment 1
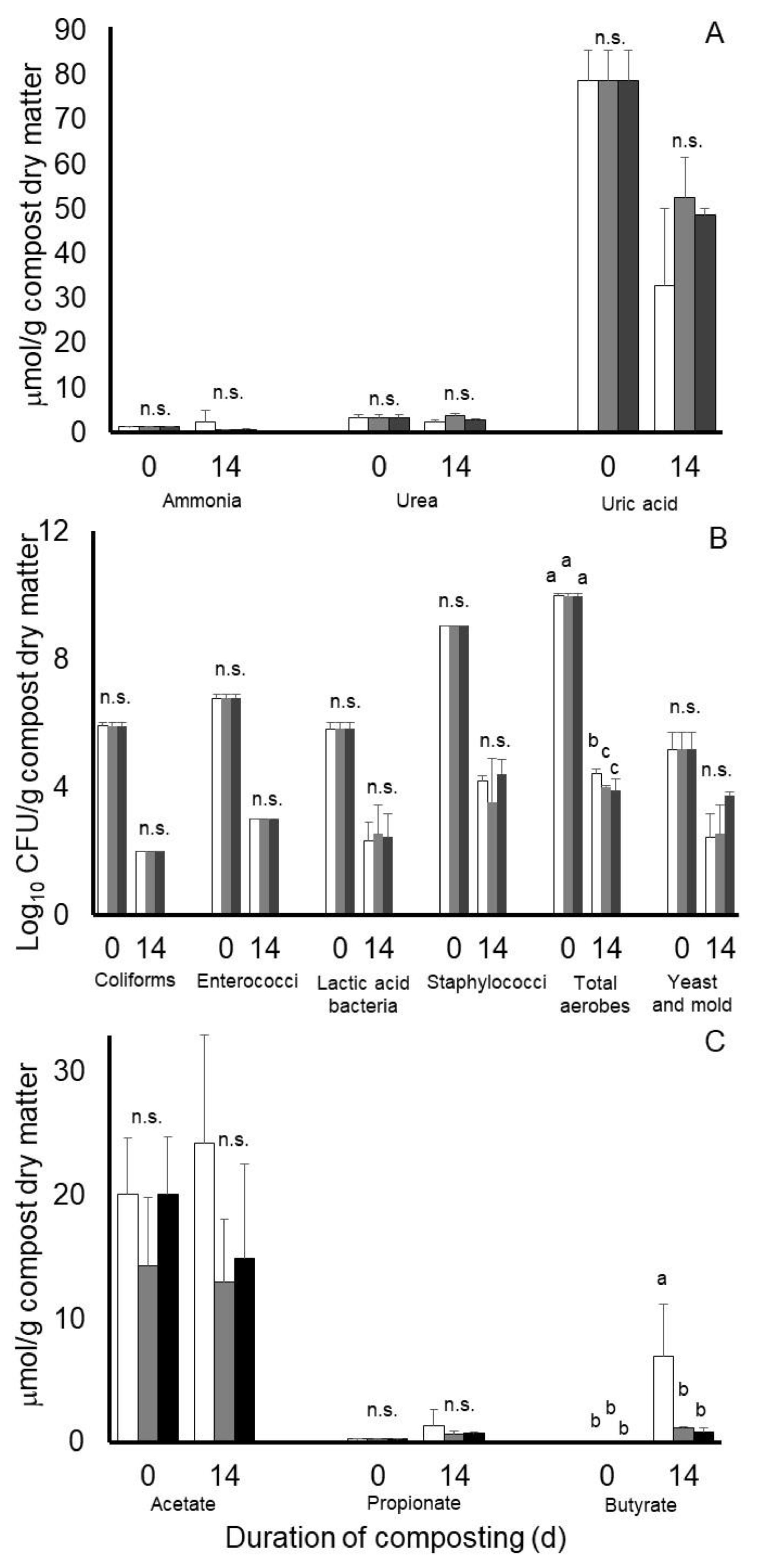
3.2. Experiment 2
3.3. Experiment 3
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bolan, N.S.; Szogi, A.A.; Chuasavathi, T.; Seshadri, B.; Rothrock, M.J.; Panneerselvam, P. Uses and management of poultry litter. World Poult. Sci. J. 2010, 66, 673–698. [Google Scholar] [CrossRef]
- Edwards, D.R.; Daniel, T.C. Environmental impacts of on-farm poultry waste disposal—A review. Bioresour. Technol. 1992, 41, 9–33. [Google Scholar] [CrossRef]
- Bhattacharya, A.N.; Fontenot, J.P. Utilization of different levels of poultry nitrogen by sheep. J. Anim. Sci. 1965, 24, 1174–1178. [Google Scholar] [CrossRef]
- Bhattacharya, A.N.; Fontenot, J.P. Protein and energy value of peanut hull and wood shaving poultry litters. J. Anim. Sci. 1966, 25, 367–371. [Google Scholar] [CrossRef]
- Chen, Z.; Jiang, X. Microbiological safety of chicken litter or chicken litter-based organic fertilizers: A review. Agriculture 2014, 4, 1–29. [Google Scholar] [CrossRef]
- Jeffrey, J.; Kirk, J.; Atwill, E.; Cullor, J.S. Research notes: Prevalence of selected microbial pathogens in processed poultry waste used as dairy cattle feed. Poult. Sci. 1998, 77, 808–811. [Google Scholar] [CrossRef]
- Bakshi, M.P.S.; Fontenot, J.P. Processing and nutritive evaluation of broiler litter as a feed. Anim. Feed Sci. Technol. 1988, 74, 337–345. [Google Scholar] [CrossRef]
- Kirchmann, H.; Witter, E. Ammonia volatilization during aerobic and anaerobic manure decomposition. Plant Soil 1989, 115, 35–41. [Google Scholar] [CrossRef]
- Tiquia, S.T.; Tamb, N.F.Y. Fate of nitrogen during composting of chicken litter. Environ. Pollut. 2020, 110, 535–541. [Google Scholar] [CrossRef]
- Chaudhry, S.M.; Naseer, Z.; Chaudhry, D.M. Fermentation characteristics and nutritive value of broiler litter ensiled with corn forage. Food Chem. 1993, 48, 51–55. [Google Scholar] [CrossRef]
- Chaudhry, S.M.; Naseer, Z. Safety of ensiling poultry litter with sugar cane tops. Pak. J. Agric. Sci. 2008, 45, 322–326. [Google Scholar]
- Steiner, C.; Das, K.C.; Melear, N.; Lakly, D. Reducing nitrogen loss during poultry litter composting using biochar. J. Environ. Qual. 2010, 39, 1236–1242. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Bai, M.; Han, B.; Impraim, R.; Butterly, C.; Hu, H.; He, J.; Chen, D. Enhanced nitrogen retention by lignite during poultry litter composting. J. Clean. Prod. 2020, 277, 122422. [Google Scholar] [CrossRef]
- Arzola-Alavarez, C.; Castillo-Castillo, Y.; Anderson, R.C.; Hume, M.E.; Ruiz-Barrera, O.; Min, B.R.; Arzola-Rubio, A.; Beier, R.C.; Salinas-Chavira, J. Influence of pine bark tannin on bacterial pathogens growth and nitrogen compounds on changes in composted poultry litter. Braz. J. Poult. Sci. 2020, 22, 1–5. [Google Scholar] [CrossRef]
- Anderson, R.C.; Vodovnik, M.; Min, B.R.; Pinchak, W.E.; Krueger, N.A.; Harvey, R.B.; Nisbet, D.J. Bactericidal effect of hydrolysable and condensed tannin extracts on Campylobacter jejuni In Vitro. Folia Microbiol. 2012, 4, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Owen, P.L.; Johns, T. Xanthine oxidase inhibitory activity of northeastern North American plant remedies used for gout. J. Ethnopharmacol. 1999, 64, 149–160. [Google Scholar] [CrossRef]
- Vogels, G.D.; van der Drift, C. Degradation of purines and pyrimidines by microorganisms. Bacteriol. Rev. 1976, 40, 403–468. [Google Scholar] [CrossRef]
- Ruiz-Barrera, O.; Ontiveros-Magadan, M.; Anderson, R.C.; Byrd, J.A.; Hume, M.E.; Latham, E.A.; Nisbet, D.J.; Arzola-Alvarez, C.; Salinas-Chavira, J.; Castillo-Castillo, Y. Nitro-treatment of composted poultry litter; effects on Salmonella, E. coli and nitrogen metabolism. Bioresour. Technol. 2020, 310, 123459. [Google Scholar] [CrossRef]
- Arzola-Alvarez, C.; Anderson, R.C.; Hume, M.E.; Ledezma, E.; Ruiz-Barrera, O.; Castillo-Castillo, Y.; Arzola-Rubio, A.; Ontiveros-Magadan, M.; Min, B.R.; Wottlin, L.R.; et al. Effect of select tannin sources on pathogen control and microbial nitrogen metabolism in composted poultry litter intended for use as a ruminant crude protein feedstuff. Front. Vet. Sci. 2022, 9, 930980. [Google Scholar] [CrossRef] [PubMed]
- Kwak, W.S.; Huh, J.W.; McCaskey, T.A. Effect of processing time on enteric bacteria survival and on temperature and chemical composition of broiler poultry litter processed by two methods. Bioresour. Technol. 2005, 96, 1529–1536. [Google Scholar] [CrossRef] [PubMed]
- Čermák, P.; Palečková, V.; Houška, M.; Strohalm, J.; Novotná, P.; Mikyška, A.; Jurková, M.; Sikorová, M. Inhibitory effects of fresh hops on Helicobacter pylori strains. Czech J. Food Sci. 2015, 33, 302–307. [Google Scholar] [CrossRef]
- Bocquet, L.B.; Sahpaz, S.; Riviève, C. An overview of the antimicrobial properties of hops. In Natural Antimicrobial Agents; Mérillon, J.-M., Rivière, C., Eds.; Springer International Publishing: New York, NY, USA, 2018; pp. 31–54. [Google Scholar] [CrossRef]
- Flythe, M.D. The antimicrobial effects of hops (Humulus lupulus L.) on ruminal hyper ammonia-producing bacteria. Lett. Appl. Microbiol. 2009, 48, 712–717. [Google Scholar] [CrossRef] [PubMed]
- Siragusa, G.R.; Haas, G.J.; Matthews, P.D.; Smith, R.J.; Buhr, R.J.; Dale, N.M.; Wise, M.G. Antimicrobial activity of lupulone against Clostridium perfringens in the chicken intestinal tract jejunum and caecum. J. Antimicrob. Chemother. 2008, 61, 853–858. [Google Scholar] [CrossRef]
- Tillman, G.E.; Haas, G.J.; Wise, M.G.; Oakley, B.; Smith, A.M.; Siragusa, G.R. Chicken intestine microbiota following the administration of lupolone, a hop-based antimicrobial. FEMS Microbiol. Ecol. 2011, 77, 395–403. [Google Scholar] [CrossRef]
- Galperin, M.Y.; Brover, V.; Tolstoy, I.; Yutin, N. Phylogenomic analysis of the family Peptostreptococcaceae (Clostridium cluster XI) and proposal for reclassification of Clostridium litorale (Fendrich et al. 1991) and Eubacterium acidaminophilum (Zindel et al. 1989) as Peptoclostridium litorale gen. nov. comb. nov. and Peptoclostridium acidaminophilum comb. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5506–5513. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, T.R.; Cotta, M.A. Isolation and identification of hyper-ammonia producing bacteria from swine manure storage pits. Curr. Microbiol. 2004, 48, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Hartinger, T.; Gresner, N.; Südekum, K.H. Does intra-ruminal nitrogen recycling waste valuable resources? A review of major players and their manipulation. J. Anim. Sci. Biotechnol. 2018, 9, 33. [Google Scholar] [CrossRef]
- Biglar, M.; Sufi, M.; Bagherzadeh, K.; Amanlou, M.; Mojab, F. Screening of 20 commonly used Iranian traditional medicinal plants against urease. Iran. J. Pharm. Res. 2014, 13 (Suppl. S1), 195–198. [Google Scholar] [CrossRef] [PubMed]
- Keskin, Ş.; Şirin, Y.; Çakir, H.E.; Keskin, M. An investigation of Humulus lupulus L.: Phenolic composition, antioxidant capacity and inhibition properties of clinically important enzymes. S. Afr. J. Bot. 2019, 120, 170–174. [Google Scholar] [CrossRef]
- Flythe, M.D.; Harlow, B.E.; Aiken, G.E.; Gellin, G.L.; Kagan, I.A.; Pappas, J. Inhibition of growth and ammonia production of ruminal hyper ammonia-producing bacteria by Chinook or Galena hops after long-term storage. Fermentation 2017, 3, 68. [Google Scholar] [CrossRef]
- Sakamoto, K.; Konings, W.N. Beer spoilage bacteria and hop resistance. Inter. J. Food Microbiol. 2003, 89, 105–124. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.J.; Russell, J.B. More monensin-sensitive, ammonia-producing bacteria from the rumen. Appl. Environ. Microbiol. 1989, 55, 1052–1057. [Google Scholar] [CrossRef] [PubMed]
- Flythe, M.D.; Aiken, G.E. Effects of hops (Humulus lupulus L.) extract on volatile fatty acid production by rumen bacteria. J. Appl. Microbiol. 2010, 109, 1169–1176. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.C.; Jung, Y.S.; Oliver, C.E.; Horrocks, S.M.; Genovese, K.J.; Harvey, R.B.; Callaway, T.R.; Edrington, T.S.; Nisbet, D.J. Effects of nitrate or nitro-supplementation, with or without added chlorate, on Salmonella enterica serovar Typhimurium and Escherichia coli in swine feces. J. Food Prot. 2007, 70, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Chaney, A.L.; Marbach, E.P. Modified reagents for determination of urea and ammonia. Clin. Chem. 1962, 8, 130–132. [Google Scholar] [CrossRef]
- Cagle, C.M.; Batista, L.F.D.; Anderson, R.C.; Fonseca, M.A.; Cravey, M.D.; Julien, C.; Tedeschi, L.O. Evaluation of different inclusion levels of dry live yeast impacts on various rumen parameters and in situ digestibilities of dry matter and neutral detergent fiber in growing and finishing beef cattle. J. Anim. Sci. 2019, 97, 4987–4998. [Google Scholar] [CrossRef] [PubMed]
- Fahle, A.; Bereswill, S.; Heimesaat, M.M. Antibacterial effects of biologically active ingredients in hop provide promising options to fight infections by pathogens including multi-drug resistant bacteria. Eur. J. Microbiol. Immunol. 2022, 12, 22–30. [Google Scholar] [CrossRef]
- Tabler, T.; Liang, Y.; Wells, J.; Moon, J. Publication 3601, 2021. Broiler Litter Production and Nutrient Content Characteristics. Mississippi State University Extension. 2021. Available online: https://extension.msstate.edu/sites/default/files/publications/publications/P3601_web.pdf (accessed on 25 May 2022).
- Kim, S.C.; Yi, S.J.; Choi, I.H. Reducing volatile fatty acids in poultry litter with chemicals additives: A laboratory experiment. J. Poult. Sci. 2011, 48, 139–143. [Google Scholar] [CrossRef]
- Onrust, L.; Ducatelle, R.; Van Driessche, K.; De Maesschalck, C.; Vermeulen, K.; Haesebrouck, F.; Eeckhaut, V.; Van Immersee, l.F. Steering endogenous butyrate production in the intestinal tract of broilers as a tool to improve gut health. Front. Vet. Sci. 2015, 2, 75. [Google Scholar] [CrossRef]
- Wang, L.; Lilburn, M.; Yu, Z. Intestinal microbiota of broiler chickens as affected by litter management regimens. Front. Microbiol. 2016, 7, 593. [Google Scholar] [CrossRef]
- Blatchford, P.A.; Parkar, S.G.; Hopkins, W.; Ingram, J.R.; Sutton, K.H. Dose-dependent alterations to in vitro human microbiota composition and butyrate inhibition by a supercritical carbon dioxide hops extract. Biomolecules 2019, 9, 390. [Google Scholar] [CrossRef] [PubMed]
- Lavrenčič, A.; Levart, A.; Košir, I.J.; Čerenak, A. Influence of two hop (Humulus lupulus L.) varieties on in vitro dry matter and crude protein degradability and digestibility in ruminants. J. Sci. Food. Agric. 2014, 94, 1248–1252. [Google Scholar] [CrossRef]
- Kung, L.; Hession, A.O. Preventing in vitro lactate accumulation in ruminal fermentations by inoculation with Megasphaera elsdenii. J. Anim. Sci. 1995, 73, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Narvaez, N.; Wang, Y.; Xu, Z.; Alexander, T.; Garden, S.; McAllister, T. Effects of hop varieties on ruminal fermentation and bacterial community in an artificial rumen (rusitec). J. Sci. Food Agric. 2013, 93, 45–52. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| F Statistic (p-Value) | |||
|---|---|---|---|
| Treatment | Day | Treatment by Day | |
| Nitrogen measurements | |||
| Ammonia | 2.50 (0.0987) | 261.6 (<0.0001) | 5.22 (0.0015) |
| Urea | 6.52 (0.0044) | 0.80 (0.5039) | 1.81 (0.1394) |
| Uric acid | 0.03 (0.9710) | 3.60 (0.0280) | 0.29 (0.9341) |
| Microbiological measurements | |||
| Salmonella | 2.13 (0.1367) | 467.7 (<0.0001) | 4.11 (0.0056) |
| Escherichia coli | 8.40 (0.0013) | 840.8 (<0.0001) | 11.11 (<0.0001) |
| Total aerobes | 3.52 (0.0422) | 402.3 (<0.0001) | 0.63 (0.7018) |
| Degrees of Freedom | 2 | 3 | 6 |
| Experiment 2, F Statistic (p-Value) | Experiment 3, F Statistic (p-Value) | |||||
|---|---|---|---|---|---|---|
| Treatment | Day | Treatment by Day | Treatment | Day | Treatment by Day | |
| Nitrogen measurements | ||||||
| Ammonia | 1.19 (0.3324) | 0.00 (9570) | 1.23 (0.3260) | 0.90 (0.4351) | 230.67 (<0.0001) | 0.55 (0.5952) |
| Urea | 1.84 (0.1945) | 1.83 (0.2007) | 2.15 (0.1595) | 0.27 (0.7695) | 10.46 (0.0102) | 0.15 (0.8617) |
| Uric acid | 1.67 (0.2230) | 60.98 (<0.0001) | 1.89 (0.1941) | 0.07 (0.9354) | 809.25 (<0.0001) | 0.04 (0.9634) |
| Microbiological measurements | ||||||
| Coliforms | 0.00 (1.000) | 13,483.23 (<0.0001) | 0.00 (1.000) | 0.83 (0.4628) | 130.64 (<0.0001) | 0.50 (0.6217) |
| Enterococci | 0.00 (1.000) | 11,649.84 (<0.0001) | 0.00 (1.000) | 0.38 (0.6952) | 531.49 (<0.0001) | 0.21 (0.8107) |
| Lactic acid bacteria | 0.06 (0.9452) | 168.58 (<0.0001) | 0.05 (0.9525) | 0.97 (0.4021) | 456.55 (<0.0001) | 0.60 (0.5694) |
| Staphylococci | 0.93 (0.4183) | 327.88 (<0.0001) | 0.92 (0.4259) | 0.12 (0.8912) | 1924.19 (<0.0001) | 0.06 (0.9380) |
| Total aerobes | 2.86 (0.0906) | 55,32.72 (<0.0001) | 4.15 (0.0425) | 0.50 (0.6178) | 1080.86 (<0.0001) | 0.29 (0.7534) |
| Yeast and mold | 1.77 (0.2058) | 59.74 (<0.0001) | 2.04 (0.1733) | 0.09 (0.9158) | 54.49 (<0.0001) | 0.05 (0.9525) |
| Volatile fatty acid accumulations | ||||||
| Acetate | 2.87 (0.0900) | 0.08 (0.7808) | 0.85 (0.4521) | 2.02 (0.1795) | 3.46 (0.0960) | 1.46 (0.2833) |
| Propionate | 0.70 (0.5128) | 6.25 (0.0279) | 0.67 (0.5311) | 2.26 (0.1501) | 2.32 (0.1620) | 1.70 (0.2361) |
| Butyrate | 3.08 (0.0801) | 23.25 (0.0005) | 9.45 (0.0041) | 3.80 (0.0556) | 20.16 (0.0015) | 3.84 (0.0621) |
| Isobutyrate | 0.99 (0.3957) | 2.76 (0.1228) | 0.99 (0.4000) | 0.95 (0.4156) | 0.75 (0.4102) | 0.59 (0.5759) |
| Isovalerate | 1.13 (0.3502) | 2.40 (0.1476) | 1.16 (0.3470) | 3.38 (0.0716) | 49.17 (0.0001) | 3.13 (0.0930) |
| Valerate | 0.24 (0.7917) | 0.00 (0.9638) | 0.21 (0.8128) | 0.91 (0.4306) | 19.65 (0.0016) | 0.56 (0.5908) |
| Degrees of freedom | 2 | 1 | 2 | 2 | 1 | 2 |
| Nitrogen Characteristics (µmol/g Compost Dry Matter) | Microbiological Characteristics (log10 Colony Forming Units/g Compost Dry Matter) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Ammonia | Urea | Uric Acid | Coliforms | Enterococci | Lactic Acid Bacteria | Staphylococci | Total Aerobes | Yeast and Mold | |
| Pre-compost | 1.32 n.s. | 3.40 n.s. | 78.89 a | 5.92 a | 6.79 a | 5.83 a | 9.05 a | 9.99 a | 5.18 a |
| Ending (day 14) | 1.29 | 3.05 | 44.80 b | 2.00 b | 3.00 b | 2.43 a | 4.06 a | 4.11 a | 2.90 b |
| SEM 2 | 0.371 | 0.183 | 3.087 | 0.024 | 0.025 | 0.185 | 0.195 | 0.056 | 0.208 |
| Nitrogen Characteristics (µmol/g Compost Dry Matter) | Microbiological Characteristics (Log10 Colony Forming Units/g Compost Dry Matter) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Ammonia | Urea | Uric Acid | Coliforms | Enterococci | Lactic Acid Bacteria | Staphylococci | Total Aerobes | Yeast and Mold | |
| Pre-compost | 2.29 a | 0.74 b | 72.50 a | 5.25 a | 5.69 a | 5.18 a | 9.00 a | 9.77 a | 5.21 a |
| Ending (day 14) | 1.21 b | 1.40 a | 29.80 b | 2.19 b | 3.05 b | 2.11 b | 3.99 b | 4.31 b | 2.69 b |
| SEM 2 | 0.055 | 0.158 | 1.163 | 0.208 | 0.089 | 0.111 | 0.089 | 0.129 | 0.265 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castillo-Castillo, Y.; Arzola-Alvarez, C.; Fonseca, M.; Salinas-Chavira, J.; Ontiveros-Magadan, M.; Hume, M.E.; Anderson, R.C.; Flythe, M.D.; Byrd, J.A.; Ruiz-Barrera, O. Effects of Hops Treatment on Nitrogen Retention, Volatile Fatty Acid Accumulations, and Select Microbial Populations of Composting Poultry Litter Intended for Use as a Ruminant Feedstuff. Microorganisms 2023, 11, 839. https://doi.org/10.3390/microorganisms11040839
Castillo-Castillo Y, Arzola-Alvarez C, Fonseca M, Salinas-Chavira J, Ontiveros-Magadan M, Hume ME, Anderson RC, Flythe MD, Byrd JA, Ruiz-Barrera O. Effects of Hops Treatment on Nitrogen Retention, Volatile Fatty Acid Accumulations, and Select Microbial Populations of Composting Poultry Litter Intended for Use as a Ruminant Feedstuff. Microorganisms. 2023; 11(4):839. https://doi.org/10.3390/microorganisms11040839
Chicago/Turabian StyleCastillo-Castillo, Yamicela, Claudio Arzola-Alvarez, Mozart Fonseca, Jaime Salinas-Chavira, Marina Ontiveros-Magadan, Michael E. Hume, Robin C. Anderson, Michael D. Flythe, James Allen Byrd, and Oscar Ruiz-Barrera. 2023. "Effects of Hops Treatment on Nitrogen Retention, Volatile Fatty Acid Accumulations, and Select Microbial Populations of Composting Poultry Litter Intended for Use as a Ruminant Feedstuff" Microorganisms 11, no. 4: 839. https://doi.org/10.3390/microorganisms11040839
APA StyleCastillo-Castillo, Y., Arzola-Alvarez, C., Fonseca, M., Salinas-Chavira, J., Ontiveros-Magadan, M., Hume, M. E., Anderson, R. C., Flythe, M. D., Byrd, J. A., & Ruiz-Barrera, O. (2023). Effects of Hops Treatment on Nitrogen Retention, Volatile Fatty Acid Accumulations, and Select Microbial Populations of Composting Poultry Litter Intended for Use as a Ruminant Feedstuff. Microorganisms, 11(4), 839. https://doi.org/10.3390/microorganisms11040839