

Role of the Microbiota in Skin Neoplasms: New Therapeutic Horizons




{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
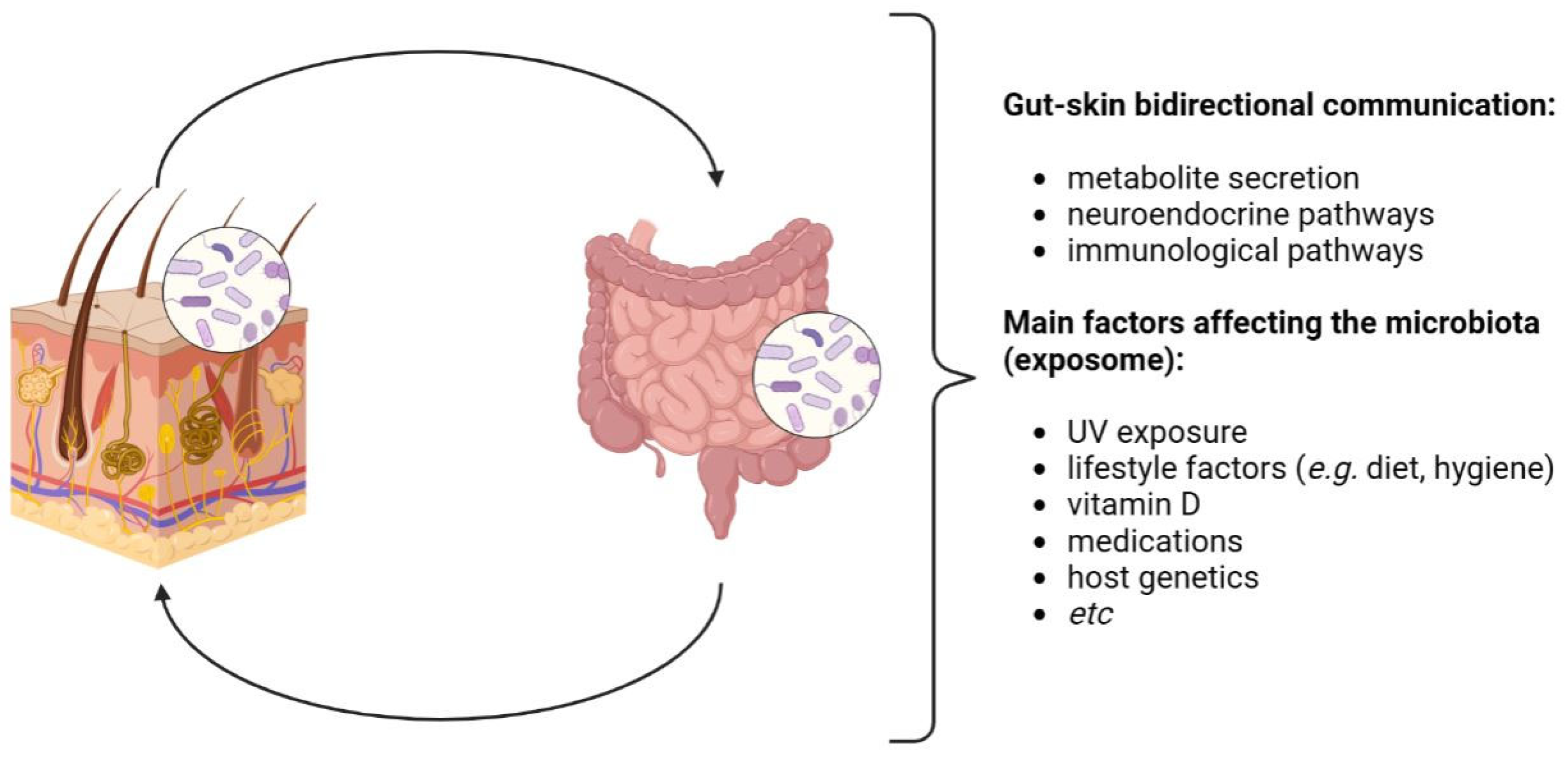
2. The Human Microbiota and the Gut–Skin Axis
3. Experimental Evidence of the Role of Microbiota in Skin Carcinogenic Processes
4. Microbiota and Melanoma
5. Microbiota and Non-Melanoma Skin Cancer
6. Microbiota and Other Cutaneous Tumors
6.1. Cutaneous T-Cell Lymphomas
6.2. Merkel Cell Carcinoma
6.3. Kaposi’s Sarcoma
7. Potential Therapeutic Implications
7.1. Microbiota Modulation
7.2. Immunomodulation by Microbiota
7.3. Bacterial Anticancer Metabolites
7.4. TLR-Targeted Therapy
7.5. Antibiotics
8. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mahmud, M.R.; Akter, S.; Tamanna, S.K.; Mazumder, L.; Esti, I.Z.; Banerjee, S.; Akter, S.; Hasan, M.R.; Acharjee, M.; Hossain, M.S.; et al. Impact of gut microbiome on skin health: Gut-skin axis observed through the lenses of therapeutics and skin diseases. Gut Microbes 2022, 14, 2096995. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Su, J.; Liu, C.; Mai, Z.; Yin, S.; Yang, C.; Fu, L. Human microbiomes in cancer development and therapy. Med. Comm. 2020, 26, e221. [Google Scholar] [CrossRef] [PubMed]
- Bibbò, S.; Ianiro, G.; Giambò, F.; Settanni, C.R.; Cammarota, G.; Gasbarrini, A. Role of gut microbiome on immunotherapy efficacy in melanoma. Hum. Vaccin. Immunother. 2022, 18, 1926759. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.; Lin, G.; Ferenczi, K. The skin microbiome and the gut-skin axis. Clin. Dermatol. 2021, 39, 829–839. [Google Scholar] [CrossRef] [PubMed]
- De Pessemier, B.; Grine, L.; Debaere, M.; Maes, A.; Paetzold, B.; Callewaert, C. Gut-Skin Axis: Current Knowledge of the Interrelationship between Microbial Dysbiosis and Skin Conditions. Microorganisms 2021, 9, 353. [Google Scholar] [CrossRef]
- Garcia-Gutierrez, E.; Mayer, M.J.; Cotter, P.D.; Narbad, A. Gut microbiota as a source of novel antimicrobials. Gut Microbes 2019, 10, 1–21. [Google Scholar] [CrossRef]
- Schauber, J.; Gallo, R.L. Antimicrobial peptides and the skin immune defense system. J. Allergy Clin. 2008, 122, 261–266. [Google Scholar] [CrossRef]
- Veniaminova, N.A.; Jia, Y.; Hartigan, A.M.; Huyge, T.J.; Tsai, S.Y.; Grachtchouk, M.; Nakagawa, S.; Dlugosz, A.A.; Atwood, S.X.; Wong, S.Y. Distinct mechanisms for sebaceous gland self-renewal and regeneration provide durability in response to injury. Cell Rep. 2023, 42, 113121. [Google Scholar] [CrossRef]
- Paone, P.; Cani, P.D. Mucus barrier, mucins and gut microbiota: The expected slimy partners? Gut 2020, 69, 2232–2243. [Google Scholar] [CrossRef]
- Li, Y.; Jin, Y.; Zhang, J.; Pan, H.; Wu, L.; Liu, D.; Liu, J.; Hu, J.; Shen, J. Recovery of human gut microbiota genomes with third-generation sequencing. Cell Death Dis. 2021, 12, 569. [Google Scholar] [CrossRef]
- Ederveen, T.H.A.; Smits, J.P.H.; Boekhorst, J.; Schalkwijk, J.; van den Bogaard, E.H.; Zeeuwen, P.L.J.M. Skin microbiota in health and disease: From sequencing to biology. J. Dermatol. 2020, 47, 1110–1118. [Google Scholar] [CrossRef] [PubMed]
- Grice, E.A.; Segre, J.A. The skin microbiome. Nat. Rev. Microbiol. 2011, 9, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Sugimura, N.; Burgermeister, E.; Ebert, M.P.; Zuo, T.; Lan, P. The gut virome: A new microbiome component in health and disease. EBioMedicine 2022, 81, 104113. [Google Scholar] [CrossRef]
- Salem, I.; Ramser, A.; Isham, N.; Ghannoum, M.A. The Gut Microbiome as a Major Regulator of the Gut-Skin Axis. Front. Microbiol. 2018, 9, 1459. [Google Scholar] [CrossRef] [PubMed]
- Pérez, J.C. Fungi of the human gut microbiota: Roles and significance. Int. J. Med. Microbiol. 2021, 311, 151490. [Google Scholar] [CrossRef]
- Skowron, K.; Bauza-Kaszewska, J.; Kraszewska, Z.; Wiktorczyk-Kapischke, N.; Grudlewska-Buda, K.; Kwiecińska-Piróg, J.; Wałecka-Zacharska, E.; Radtke, L.; Gospodarek-Komkowska, E. Human Skin Microbiome: Impact of Intrinsic and Extrinsic Factors on Skin Microbiota. Microorganisms 2021, 9, 543. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, C.A.; Monteleone, G.; McLaughlin, J.T.; Paus, R. The gut-skin axis in health and disease: A paradigm with therapeutic implications. Bioessays 2016, 38, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.S.; Wang, J.; Yannie, P.J.; Ghosh, S. Intestinal Barrier Dysfunction, LPS Translocation, and Disease Development. J. Endocr. Soc. 2020, 4, bvz039. [Google Scholar] [CrossRef] [PubMed]
- Denechaud, P.D.; Rabhi, N. Editorial: Intra and inter organ crosstalk and cellular communication. Front. Endocrinol. 2023, 14, 1209436. [Google Scholar] [CrossRef]
- Gueniche, A.; Philippe, D.; Bastien, P.; Reuteler, G.; Blum, S.; Castiel-Higounenc, I.; Breton, L.; Benyacoub, J. Randomised double-blind placebo-controlled study of the effect of Lactobacillus paracasei NCC 2461 on skin reactivity. Benef. Microbes 2014, 5, 137–145. [Google Scholar] [CrossRef]
- Alexander, H.; Brown, S.; Danby, S.; Flohr, C. Research Techniques Made Simple: Transepidermal Water Loss Measurement as a Research Tool. J. Investig. Dermatol. 2018, 138, 2295–2300. [Google Scholar] [CrossRef]
- Xiao, X.; Hu, X.; Yao, J.; Cao, W.; Zou, Z.; Wang, L.; Qin, H.; Zhong, D.; Li, Y.; Xue, P.; et al. The role of short-chain fatty acids in inflammatory skin diseases. Front. Microbiol. 2023, 13, 1083432. [Google Scholar] [CrossRef]
- Souak, D.; Barreau, M.; Courtois, A.; André, V.; Duclairoir Poc, C.; Feuilloley, M.G.J.; Gault, M. Challenging Cosmetic Innovation: The Skin Microbiota and Probiotics Protect the Skin from UV-Induced Damage. Microorganisms 2021, 9, 936. [Google Scholar] [CrossRef]
- Patra, V.; Wagner, K.; Arulampalam, V.; Wolf, P. Skin Microbiome Modulates the Effect of Ultraviolet Radiation on Cellular Response and Immune Function. iScience 2019, 15, 211–222. [Google Scholar] [CrossRef]
- Ghaly, S.; Kaakoush, N.O.; Hart, P.H. Effects of UVR exposure on the gut microbiota of mice and humans. Photochem. Photobiol. Sci. 2020, 19, 20–28. [Google Scholar] [CrossRef]
- Squarzanti, D.F.; Sorrentino, R.; Azzimonti, B. High-risk HPVs, microbiota, and epithelial carcinogenesis: State of the art and research contribution of in vitro 3D models. J. Cancer Metastasis Treat. 2019, 5, 73. [Google Scholar] [CrossRef]
- Squarzanti, D.F.; Zavattaro, E.; Pizzimenti, S.; Amoruso, A.; Savoia, P.; Azzimonti, B. Non-Melanoma Skin Cancer: News from microbiota research. Crit. Rev. Microbiol. 2020, 46, 433–449. [Google Scholar] [CrossRef] [PubMed]
- Squarzanti, D.F.; Zanetta, P.; Azzimonti, B. Non-melanoma skin cancer and the cutaneous microbiota network. Biol. Med. 2020, 12, 1b. [Google Scholar]
- Azzimonti, B.; Ballacchino, C.; Zanetta, P.; Cucci, M.A.; Monge, C.; Grattarola, M.; Dianzani, C.; Barrera, G.; Pizzimenti, S. Microbiota, Oxidative Stress, and Skin Cancer: An Unexpected Triangle. Antioxidants 2023, 12, 546. [Google Scholar] [CrossRef]
- Ke, Y.; Wang, X.J. TGFβ Signaling in Photoaging and UV-Induced Skin Cancer. J. Investig. Dermatol. 2021, 141, 1104–1110. [Google Scholar] [CrossRef]
- Bauché, D.; Marie, J.C. Transforming growth factor β: A master regulator of the gut microbiota and immune cell interactions. Clin. Transl. Immunol. 2017, 6, e136. [Google Scholar] [CrossRef]
- Hazrat, B.; Yuanyuan, X.; Muhammad Nadeem, K.; Chen, J.; Wang, Y.; Zeng, Y.; Lin, X. Stabilization of Acne Vulgaris-Associated Microbial Dysbiosis with 2% Supramolecular Salicylic Acid. Pharmaceuticals 2023, 16, 87. [Google Scholar]
- Severn, M.M.; Horswill, A.R. Staphylococcus epidermidis and its dual lifestyle in skin health and infection. Nat. Rev. Microbiol. 2023, 21, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Sandhu, E.; Yang, X.; Yang, J.; Ren, Y.; Gao, X. Bidirectional Functional Effects of Staphylococcus on Carcinogenesis. Microorganisms 2022, 10, 2353. [Google Scholar] [CrossRef]
- Kullander, J.; Forslund, O.; Dillner, J. Staphylococcus aureus and squamous cell carcinoma of the skin. Cancer Epidemiol. Biomark. Prev. 2009, 18, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Fujii, K. Pathogenesis of cutaneous T cell lymphoma: Involvement of Staphylococcus aureus. J. Dermatol. 2022, 49, 202–209. [Google Scholar] [CrossRef]
- Takei, I.; Sawamura, S.; Myangat, T.M.; Kajihara, I.; Kanemaru, H.; Kashiwada-Nakamura, K.; Makino, K.; Aoi, J.; Masuguchi, S.; Fukushima, S. Clinical significance of skin colonization of Pseudomonas aeruginosa in cutaneous squamous cell carcinoma. J. Dermatol. 2021, 48, e581–e582. [Google Scholar] [CrossRef]
- Woo, Y.R.; Cho, S.H.; Lee, J.D.; Kim, H.S. The Human Microbiota and Skin Cancer. Int. J. Mol. Sci. 2022, 23, 1813. [Google Scholar] [CrossRef]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like Receptors and the Control of Immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef]
- Vijay, K. Toll-like receptors in immunity and inflammatory diseases: Past, present, and future. Int. Immunopharmacol. 2018, 59, 391–412. [Google Scholar] [CrossRef]
- Zhao, Q.; Wang, Q.; Wang, T.; Xu, J.; Li, T.; Liu, Q.; Yao, Q.; Wang, P. Pattern Recognition Receptors (PRRs) in Macrophages Possess Prognosis and Immunotherapy Potential for Melanoma. Front. Immunol. 2021, 12, 765615. [Google Scholar] [CrossRef]
- Zhang, Y.; Liang, X.; Bao, X.; Xiao, W.; Chen, G. Toll-like receptor 4 (TLR4) inhibitors: Current research and prospective. Eur. J. Med. Chem. 2022, 235, 114291. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Liu, X.; Zhang, S.; Dang, S.; Zhang, Y.; Zhang, G. Expression of TRIM44 and its correlation with TLR4 in laryngeal squamous cell carcinoma Cell. Mol. Biol. 2022, 68, 56–61. [Google Scholar]
- Zu, Y.; Ping, W.; Deng, T.; Zhang, N.; Fu, X.; Sun, W. Lipopolysaccharide-induced toll-like receptor 4 signaling in esophageal squamous cell carcinoma promotes tumor proliferation and regulates inflammatory cytokines expression. Dis. Esophagus 2017, 30, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Davakis, S.; Kapelouzou, A.; Liakakos, T.; Mpoura, M.; Stergiou, D.; Sakellariou, S.; Charalabopoulos, A. The Role of Toll-like Receptors in Esophageal Cancer. Anticancer Res. 2022, 42, 2813–2818. [Google Scholar] [CrossRef]
- Shomali, N.; Hatamnezhad, L.S.; Tarzi, S.; Tamjidifar, R.; Xu, H.; Shotorbani, S.S. Heat Shock Proteins Regulating Toll-like Receptors and the Immune System could be a Novel Therapeutic Target for Melanoma. Curr. Mol. Med. 2021, 21, 15–24. [Google Scholar] [CrossRef]
- Theivanthiran, B.; Yarla, N.; Haykal, T.; Nguyen, Y.V.; Cao, L.; Ferreira, M.; Holtzhausen, A.; Al-Rohil, R.; Salama, A.K.S.; Beasley, G.M.; et al. Tumor-intrinsic NLRP3-HSP70-TLR4 axis drives premetastatic niche development and hyperprogression during anti-PD-1 immunotherapy. Sci. Transl. Med. 2022, 14, eabq7019. [Google Scholar] [CrossRef] [PubMed]
- Jun, Y.K.; Kwon, S.H.; Yoon, H.T.; Park, H.; Soh, H.; Lee, H.J.; Im, J.P.; Kim, J.S.; Kim, J.W.; Koh, S.J. Toll-like receptor 4 regulates intestinal fibrosis via cytokine expression and epithelial-mesenchymal transition. Sci. Rep. 2020, 10, 19867. [Google Scholar] [CrossRef]
- Chen, M.C.; Chang, W.W.; Kuan, Y.D.; Lin, S.T.; Hsu, H.C.; Lee, C.H. Resveratrol inhibits LPS-induced epithelial-mesenchymal transition in mouse melanoma model. Innate Immun. 2012, 18, 685–693. [Google Scholar] [CrossRef]
- Eiro, N.; Ovies, C.; Fernandez-Garcia, B.; Álvarez-Cuesta, C.; González, L.; González, L.; Vizoso, F. Expression of TLR3, 4, 7 and 9 in cutaneous malignant melanoma: Relationship with clinicopathological characteristics and prognosis. Arch. Dermatol. Res. 2013, 305, 59–67. [Google Scholar] [CrossRef] [PubMed]
- McAllister, F.; Kolls, J.K. Th17 cytokines in non-melanoma skin cancer. Eur. J. Immunol. 2015, 45, 692–694. [Google Scholar] [CrossRef] [PubMed]
- Kovács, T.; Mikó, E.; Ujlaki, G.; Sári, Z.; Bai, P. The Microbiome as a Component of the Tumor Microenvironment. Adv. Exp. Med. Biol. 2020, 1225, 137–153. [Google Scholar]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef]
- Yang, R.; Qian, L. Research on Gut Microbiota-Derived Secondary Bile Acids in Cancer Progression. Integr. Cancer Ther. 2022, 21, 15347354221114100. [Google Scholar] [CrossRef] [PubMed]
- Huo, X.; Juergens, S.; Zhang, X.; Rezaei, D.; Yu, C.; Strauch, E.D.; Wang, J.Y.; Cheng, E.; Meyer, F.; Wang, D.H.; et al. Deoxycholic acid causes DNA damage while inducing apoptotic resistance through NF-kappaB activation in benign Barrett’s epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G278–G286. [Google Scholar] [CrossRef]
- Wang, S.; Dong, W.; Liu, L. Interplay between bile acids and the gut microbiota promotes intestinal carcinogenesis. Mol. Carcinog. 2019, 58, 1155–1167. [Google Scholar] [CrossRef]
- Yu, J.; Luo, Y.; Zhu, Z.; Zhou, Y.; Sun, L.; Gao, J.; Sun, J.; Wang, G.; Yao, X.; Li, W. A tryptophan metabolite of the skin microbiota attenuates inflammation in patients with atopic dermatitis through the aryl hydrocarbon receptor. J. Allergy Clin. Immunol. 2019, 143, 2108–2119. [Google Scholar] [CrossRef]
- Sanford, J.A.; Zhang, L.J.; Williams, M.R.; Gangoiti, J.A.; Huang, C.M.; Gallo, L.R. Inhibition of HDAC8 and HDAC9 by microbial short-chain fatty acids breaks immune tolerance of the epidermis to TLR ligands. Sci. Immunol. 2016, 1, eaah4609. [Google Scholar] [CrossRef]
- González-Sánchez, P.; DeNicola, G.M. The microbiome(s) and cancer: Know thy neighbor(s). J. Pathol. 2021, 254, 332–343. [Google Scholar] [CrossRef]
- Fu, C.; Yang, Z.; Yu, J.; Wei, M. The interaction between gut microbiome and anti-tumor drug therapy. Am. J. Cancer Res. 2021, 11, 5812–5832. [Google Scholar] [PubMed]
- Mekadim, C.; Kupcova Skalnikova, H.; Cizkova, J.; Cizkova, V.; Palanova, A.; Horak, V.; Mrazek, J. Dysbiosis of skin microbiome and gut microbiome in melanoma progression. BMC Microbiol. 2022, 22, 63. [Google Scholar] [CrossRef] [PubMed]
- Mrázek, J.; Mekadim, C.; Kučerová, P.; Švejstil, R.; Salmonová, H.; Vlasáková, J.; Tarasová, R.; Čížková, J.; Červinková, M. Melanoma-related changes in skin microbiome. Folia Microbiol. 2019, 64, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Vitali, F.; Colucci, R.; Di Paola, M.; Pindo, M.; De Filippo, C.; Moretti, S.; Cavalieri, D. Early melanoma invasivity correlates with gut fungal and bacterial profiles. Br. J. Dermatol. 2022, 186, 106–116. [Google Scholar] [CrossRef]
- Makaranka, S.; Scutt, F.; Frixou, M.; Wensley, K.E.; Sharma, R.; Greenhowe, J. The gut microbiome and melanoma: A review. Exp. Dermatol. 2022, 31, 1292–1301. [Google Scholar] [CrossRef]
- Chen, L.; Zhou, X.; Wang, Y.; Wang, D.; Ke, Y.; Zeng, X. Propionate and Butyrate Produced by Gut Microbiota after Probiotic Supplementation Attenuate Lung Metastasis of Melanoma Cells in Mice. Mol. Nutr. Food Res. 2021, 65, e2100096. [Google Scholar] [CrossRef]
- Shin, J.; Song, I.S.; Pak, J.H.; Jang, S.W. Upregulation of annexin A1 expression by butyrate in human melanoma cells induces invasion by inhibiting E-cadherin expression. Tumour Biol. 2016, 37, 14577–14584. [Google Scholar] [CrossRef] [PubMed]
- Pinato, J.D.; Gramenitskaya, D.; Altmann, D.M.; Boyton, R.J.; Mullish, B.H.; Marchesi, J.R.; Bower, M. Antibiotic therapy and outcome from immune-checkpoint inhibitors. J. Immunother. Cancer 2019, 7, 287. [Google Scholar] [CrossRef]
- Swami, U.; Chennamadhavuni, A.; Borcherding, N.; Bossler, A.D.; Mott, S.L.; Garje, R.; Zakharia, Y.; Milhem, M. Multivariable Analysis of 169 Cases of Advanced Cutaneous Melanoma to Evaluate Antibiotic Exposure as Predictor of Survival to Anti-PD-1 Based Immunotherapies. Antibiotics 2020, 9, 740. [Google Scholar] [CrossRef]
- Spencer, C.N.; Spencer, C.N.; McQuade, J.D.; Gopalakrishnan, V.; McCulloch, J.M.; Vetizou, M.; Cogdill, A.P.; Wadud Khan, M.A.; Zhang, X.; White, M.G.; et al. Dietary fiber and probiotics influence the gut microbiome and melanoma immunotherapy response. Science 2021, 374, 1632–1640. [Google Scholar] [CrossRef]
- Davar, D.; Dzutsev, A.K.; McCulloch, J.A.; Rodrigues, R.R.; Chauvin, J.M.; Morrison, R.M.; Deblasio, R.N.; Menna, C.; Ding, Q.; Pagliano, O.; et al. Fecal microbiota transplant overcomes resistance to anti-PD-1 therapy in melanoma patients. Science 2021, 371, 595–602. [Google Scholar] [CrossRef]
- Barbosa, E.C.; Bucar, E.E.C.; Jubé, G.R.; Silveira, L.B.; Silva, N.C.D.; Faria, P.C.C.; Ramos, P.L.C.; Moraes, V.R.Y.; Barros, J.O.B. Fecal microbiota transplantation and its repercussions in patients with melanoma refractory to anti-PD-1 therapy: Scope review. Rev. Col. Bras. Cir. 2023, 50, e20233490. [Google Scholar] [CrossRef]
- Baruch, E.N.; Youngster, I.; Ben-Betzalel, G.; Ortenberg, R.; Lahat, A.; Katz, L.; Adler, K.; Dick-Necula, D.; Raskin, S.; Bloch, N.; et al. Fecal microbiota transplant promotes response in immunotherapy-refractory melanoma patients. Science 2021, 371, 602–609. [Google Scholar] [CrossRef]
- Routy, B.; Lenehan, J.G.; Miller, W.H.; Jamal, R.; Messaoudene, M.; Daisley, B.A.; Hes, C.; Al, K.F.; Martinez-Gili, L.; Punčochář, M.; et al. Fecal microbiota transplantation plus anti-PD-1 immunotherapy in advanced melanoma: A phase I trial. Nat. Med. 2023, 29, 2121–2132. [Google Scholar] [CrossRef] [PubMed]
- Chaput, N.; Lepage, P.; Coutzac, C.; Soularue, E.; Le Roux, K.; Monot, C.; Boselli, L.; Routier, E.; Cassard, L.; Collins, M.; et al. Baseline gut microbiota predicts clinical response and colitis in metastatic melanoma patients treated with ipilimumab. Ann. Oncol. 2017, 28, 1368–1379. [Google Scholar] [CrossRef]
- Andrews, M.C.; Duong, C.P.M.; Gopalakrishnan, V.; Iebba, V.; Chen, W.S.; Derosa, L.; Khan, M.A.W.; Cogdill, A.P.; White, M.G.; Wong, M.C.; et al. Gut microbiota signatures are associated with toxicity to combined CTLA-4 and PD-1 blockade. Nat. Med. 2021, 27, 1432–1441. [Google Scholar] [CrossRef]
- Wang, Y.; Wiesnoski, D.H.; Helmink, B.A.; Gopalakrishnan, V.; Choi, K.; DuPont, H.L.; Jiang, Z.D.; Abu-Sbeih, H.; Sanchez, C.A.; Chang, C.C.; et al. Fecal microbiota transplantation for refractory immune checkpoint inhibitor-associated colitis. Nat. Med. 2018, 24, 1804–1808. [Google Scholar] [CrossRef]
- Samarasinghe, V.; Madan, V. Nonmelanoma skin cancer. J. Cutan. Aesthet. Surg. 2012, 5, 3–10. [Google Scholar] [CrossRef]
- Willenbrink, T.J.; Ruiz, E.S.; Cornejo, C.M.; Schmults, C.D.; Arron, S.T.; Jambusaria-Pahlajani, A. Field cancerization: Definition, epidemiology, risk factors, and outcomes. J. Am. Acad. Dermatol. 2020, 83, 709–717. [Google Scholar] [CrossRef]
- Fernandez Figueras, M.T. From actinic keratosis to squamous cell carcinoma: Pathophysiology revisited. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 5–7. [Google Scholar] [CrossRef]
- Mittal, A.; Colegio, O. Skin cancers in Organ Transplant Recipients. Am. J. Transplant. 2017, 17, 2509–2530. [Google Scholar] [CrossRef] [PubMed]
- Wood, D.L.A.; Lachner, N.; Tan, J.M.; Tang, S.; Angel, N.; Laino, A.; Linedale, R.; Lê Cao, K.A.; Morrison, M.; Frazer, I.H.; et al. A natural history of actinic keratosis and cutaneous squamous cell carcinoma microbiomes. mBio 2018, 9, e01432-18. [Google Scholar] [CrossRef]
- Majewski, S.; Jablonska, S. Epidermodysplasia verruciformis human papillomavirusese contribute to malignant and benign epidermal proliferations? Arch. Dermatol. 2002, 138, 649–654. [Google Scholar] [CrossRef]
- Patel, T.; Morrison, L.K.; Rady, P.; Tyring, S. Epidermodysplasia verruciformis and susceptibility to HPV. Dis. Markers 2010, 29, 199–206. [Google Scholar] [CrossRef]
- Rogers, H.D.; Macgregor, J.L.; Nord, K.M.; Tyring, S.; Rady, P.; Engler, D.E.; Grossman, M.E. Acquired epidermodysplasia verruciformis. J. Am. Acad. Dermatol. 2009, 60, 315–320. [Google Scholar] [CrossRef]
- Meyer, T.; Arndt, R.; Nindl, I.; Ulrich, C.; Christophers, E.; Stockfleth, E. Association of human papillomavirus infections with cutaneous tumors in immunosuppressed patients. Transpl. Int. 2003, 16, 146–153. [Google Scholar] [CrossRef]
- Borgogna, C.; Lanfredini, S.; Peretti, A.; De Andrea, M.; Zavattaro, E.; Colombo, E.; Quaglia, M.; Boldorini, R.; Miglio, U.; Doorbar, J.; et al. Impreved detection reveals active ß-papillomavirus infection in skin lesions from kidney transplant recipients. Mod. Pathol. 2014, 27, 1101–1115. [Google Scholar] [CrossRef]
- Borgogna, C.; Olivero, C.; Lanfredini, S.; Calati, F.; De Andrea, M.; Zavattaro, E.; Savoia, P.; Trisolini, E.; Boldorini, R.; Patel, G.K.; et al. ß-HPV infection correlates with early stages of carcinogenesis in skin tumors and patient-derifed xenografts from a kidney transplant recipient cohort. Front. Microbiol. 2018, 9, 117. [Google Scholar] [CrossRef]
- Viarisio, D.; Müller-Decker, K.; Accardi, R.; Robitaille, A.; Dürst, M.; Beer, K.; Jansen, L.; Flechtenmacher, C.; Bozza, M.; Harbottle, R.; et al. Beta HPV38 oncoproteins act with a hit-and-run mechanism in ultraviolet radiation-induced skin carcino-genesis in mice. PLoS Pathog. 2018, 14, e1006783. [Google Scholar] [CrossRef]
- Strickley, J.D.; Messerschmidt, J.L.; Awad, M.E.; Tancheng, L.; Hasegawa, T.; Thinh Ha, D.; Nabeta, H.W.; Bevins, P.A.; Ngo, K.H.; Asgari, M.M.; et al. Immunity to commensal papillomaviruses protects against skin cancer. Nature 2019, 575, 519–522. [Google Scholar] [CrossRef]
- Neagu, N.; Dianzani, C.; Venuti, A.; Bonin, S.; Voidãzan, S.; Zalaudek, I.; Conforti, C. The role of HPV in keratinocyte skin cancer development: A systematic review. J. Eur. Acad. Dermatol. Venereol. 2023, 37, 40–46. [Google Scholar] [CrossRef] [PubMed]
- de Koning, M.N.C.; Struijk, L.; Bouwes Bavinck, J.N.; Kleter, B.; Ter Schegget, J.; Quint, W.G.V.; Feltkamp, M.C.W. Betapapillomaviruses frequently persist in the skin of healthy inidividuals. J. Gen. Virol. 2007, 88, 1489–1495. [Google Scholar] [CrossRef]
- Ha, M.V.; McCormick, T.S.; Salem, I.; Al-Shakhshir, H.; Ghannoum, M.A.; Carroll, B.T. Skin and gut microbial associations with squamous cell carcinoma in solid organ transplant recipients. Arch. Dermatol. Res. 2023, 315, 2709–2713. [Google Scholar] [CrossRef] [PubMed]
- Voigt, A.T.; Emiola, A.; Johnson, J.S.; Fleming, E.; Nguyen, H.; Zhou, W.; Tsai, K.Y.; Fink, C.; Oh, J. Skin microbiome variation with cancer progression in human cutaneous squamous cell carcinoma. J. Investig. Dermatol. 2022, 142, 2773–2782. [Google Scholar] [CrossRef] [PubMed]
- Madhusudhan, N.; Pausan, M.R.; Halwachs, B.; Durdević, M.; Windisch, M.; Kehrmann, J.; Patra, V.; Wolf, P.; Boukamp, P.; Moissl-Eichinger, C.; et al. Molecular profiling of keratinocyte skin tumors links Staphylococcus aureus overabundance and increased ß-defensin-2 expression to growth promotion of squamous cell carcinoma. Cancers 2020, 12, 541. [Google Scholar] [CrossRef]
- Nagakawa, S.; Matsumoto, M.; Katayama, Y.; Oguma, R.; Wakabayashi, S.; Nygaard, T.; Saijo, S.; Inohara, N.; Otto, M.; Matsue, H.; et al. Staphylococcus aureus virulent PSMα peptides induce keratinocyte alarmin release to orchestrate IL-17-dependet skin inflammation. Cell Host Microbe 2017, 22, 667–677. [Google Scholar] [CrossRef]
- Krueger, A.; Zaugg, J.; Chisholm, S.; Linedale, R.; Lachner, N.; Teoh, S.M.; Tuong, Z.K.; Lukowski, S.W.; Morrison, M.; Soyer, H.P.; et al. Secreted toxins from Staphylococcus aureus strains isolated from keratinocyte skin cancers mediate pro-tumorigenic inflammatory responses in the skin. Front. Microbiol. 2021, 12, 789042. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Chen, T.H.; Butcher, A.M.; Trzoss, L.L.; Nam, S.J.; Shirakawa, K.T.; Zhou, W.; Oh, J.; Otto, M.; Fenical, W.; et al. A commensal strain of Staphylococcus epidermidis protects against skin neoplasia. Sci. Adv. 2019, 4, eaao4502. [Google Scholar] [CrossRef]
- Calzavara-Pinton, P.; Rossi, M.T.; Sala, R.; Venturini, M. Photodynamic antifungal chemotherapy. Photochem. Photobiol. 2012, 88, 512–522. [Google Scholar] [CrossRef]
- Gilaberte, Y.; Aspiroz, C.; Alejandre, C.; Rezusta, A. Malassezia growth on peritumoral skin after routine methyl-5-aminoluvulynate Photodynamic Therapy for actinic keratosis and Nonmelanoma Skin cancer. Actas Dermosifiliogr. 2015, 106, 70–77. [Google Scholar] [CrossRef]
- Ramsey, M.M.; Freire, M.O.; Gabrilska, R.A.; Rumbaugh, K.P.; Lemon, K.P. Staphylococcus aureus shifts toward commensalism in response to Corynebacterium species. Front. Microbiol. 2016, 7, 1230. [Google Scholar] [CrossRef]
- Bromfield, J.I.; Hugenholtz, P.; Frazer, I.H.; Khosrotehrani, K.; Chandra, J. Targeting Staphylococcus aureus dominated skin dysbiosis in actinic keratosis to prevent the onset of cutaneous squamous cell carcinoma: Outlook for future therapies? Front. Oncol. 2023, 13, 091379. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, A.D.; Campo, V.E.; Cela, E.M.; Morelli, A.E.; Shufesky, W.J.; Tckacheva, O.A.; Leoni, J.; Paz, M.L.; Larregina, A.T.; González Maglio, D.H. Oral administration of lipoteichoic acid from Lactobacillus rhamnosus GG overcomes UVB-induced immunosuppression and impairs skin tumor growth in mice. Eur. J. Immunol. 2012, 49, 2095–2102. [Google Scholar] [CrossRef] [PubMed]
- Weill, F.S.; Cela, E.M.; Paz, M.L.; Ferrari, A.; Leoni, J.; Maglio, D.H.G. Lipoteichoic acid from Lactobacillus rhamnosus GG as an oral photoprotective agent against UV-induced carcinogenesis. Br. J. Nutr. 2013, 109, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Patra, V.; Gallais Sérézal, I.; Wolf, P. Potential of skin microbiome, pro- and pre-biotics to affect local cutaneous responses to UV exposure. Nutrients 2020, 12, 1795. [Google Scholar] [CrossRef]
- Kehrmann, J.; Koch, F.; Zumdick, S.; Höwner, A.; Best, L.; Masshöfer, L.; Scharfenberg, S.; Zeschnigk, M.; Becker, J.C.; Schadendorf, D.; et al. Reduced Staphylococcus abudance characterizes the lesional microbiome of actinic keratosis patients after field-directed therapies. Microbiol. Spectr. 2023, 11, e0440122. [Google Scholar] [CrossRef]
- Wu, X.; Hwang, S.T. A Microbiota-Dependent, STAT3-Driven Mouse Model of Cutaneous T-Cell Lymphoma. J. Investig. Dermatol. 2018, 138, 1022–1026. [Google Scholar] [CrossRef]
- Salava, A.; Deptula, P.; Lyyski, A.; Laine, P.; Paulin, L.; Väkevä, L.; Ranki, A.; Auvinen, P.; Lauerma, A. Skin Microbiome in Cutaneous T-Cell Lymphoma by 16S and Whole-Genome Shotgun Sequencing. J. Investig. Dermatol. 2020, 140, 2304–2308.e7. [Google Scholar] [CrossRef]
- Harkins, C.P.; MacGibeny, M.A.; Thompson, K.; Bubic, B.; Huang, X.; Brown, I.; Park, J.; Jo, J.H.; Segre, J.A.; Kong, H.H.; et al. Cutaneous T-Cell Lymphoma Skin Microbiome Is Characterized by Shifts in Certain Commensal Bacteria but not Viruses when Compared with Healthy Controls. J. Investig. Dermatol. 2021, 141, 1604–1608. [Google Scholar] [CrossRef]
- Zhang, Y.; Seminario-Vidal, L.; Cohen, L.; Hussaini, M.; Yao, J.; Rutenberg, D.; Kim, Y.; Giualiano, A.; Robinson, L.A.; Sokol, L. Alterations in the Skin Microbiota are Associated with Symptom Severity in Mycosis Fungoides. Front. Cell. Infect. Microbiol. 2022, 12, 850509. [Google Scholar] [CrossRef]
- Jackow, C.M.; Cather, J.C.; Hearne, V.; Asano, A.T.; Musser, J.M.; Duvic, M. Association of Erythrodermic Cutaneous T-Cell Lymphoma, Superantigen-Positive Staphylococcus aureus, and Oligoclonal T-Cell Receptor V Beta Gene Expansion. Blood 1997, 89, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Talpur, R.; Bassett, R.; Duvic, M. Prevalence and Treatment of Staphylococcus aureus Colonization in Patients with Mycosis. Fungoides and Sézary Syndrome. Br. J. Dermatol. 2008, 159, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Hooper, M.J.; Enriquez, G.L.; Veon, F.L.; LeWitt, T.M.; Sweeney, D.; Green, S.J.; Seed, P.C.; Choi, J.; Guitart, J.; Burns, M.B.; et al. Narrowband ultraviolet B response in cutaneous T-cell lymphoma is characterized by increased bacterial diversity and reduced Staphylococcus aureus and Staphylococcus lugdunensis. Front. Immunol. 2022, 13, 1022093. [Google Scholar] [CrossRef]
- Dehner, C.A.; Ruff, W.E.; Greiling, T.; Pereira, M.S.; Redanz, S.; McNiff, J.; Girardi, M.; Kriegel, M.A. Malignant T Cell Activation by a Bacillus Species Isolated from Cutaneous T-Cell Lymphoma Lesions. JID Innov. 2022, 2, 100084. [Google Scholar] [CrossRef]
- Hooper, M.J.; LeWitt, T.M.; Pang, Y.; Veon, F.L.; Chlipala, G.E.; Feferman, L.; Green, S.J.; Sweeney, D.; Bagnowski, K.T.; Burns, M.B.; et al. Gut dysbiosis in cutaneous T-cell lymphoma is characterized by shifts in relative abundances of specific bacterial taxa and decreased diversity in more advanced disease. J. Eur. Acad. Dermatol. Venereol. 2022, 36, 1552–1563. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.G.; Dreno, B.; da Costa, A.C.; Li, L.; Orlandi, P.; Deng, X.; Kapusinszky, B.; Siqueira, J.; Knol, A.C.; Halary, F.; et al. A New Protoparvovirus in Human Fecal Samples and Cutaneous T Cell Lymphomas (Mycosis Fungoides). Virology 2016, 496, 299–305. [Google Scholar] [CrossRef]
- Väisänen, E.; Fu, Y.; Hedman, K.; Söderlund-Venermo, M. Human Protoparvoviruses. Viruses 2017, 9, 354. [Google Scholar] [CrossRef]
- Vaïsänen, E.; Fu, Y.; Koskenmies, S.; Fyhrquist, N.; Wang, Y.; Keinonen, A.; Mäkisalo, H.; Väkevä, L.; Pitkänen, S.; Ranki, A. Cutavirus DNA in Malignant and Nonmalignant Skin of Cutaneous T-Cell Lymphoma and Organ Transplant Patients but Not of Healthy Adults. Clin. Infect. Dis. 2019, 68, 1904–1910. [Google Scholar] [CrossRef]
- Bergallo, M.; Daprà, V.; Fava, P.; Ponti, R.; Calvi, C.; Fierro, M.T.; Quaglino, P.; Galliano, I.; Montanari, P. Lack of Detection of Cutavirus DNA Using PCR Real Time in Cutaneous T-Cell Lymphomas. G. Ital. Dermatol. Venereol. Organo Uff. Soc. Ital. Dermatol. Sifilogr. 2020, 155, 772–774. [Google Scholar] [CrossRef]
- Leroux-Kozal, V.; Lévêque, N.; Brodard, V.; Lesage, C.; Dudez, O.; Makeieff, M.; Kanagaratnam, L.; Diebold, M.B. Merkel cell carcinoma: Histopathologic and prognostic features according to the immunohistochemical expression of Merkel cell polyomavirus large T antigen correlated with viral load. Hum. Pathol. 2015, 46, 443–453. [Google Scholar] [CrossRef]
- Hernandez, L.E.; Mohsin, N.; Yaghi, M.; Frech, F.S.; Dreyfuss, I.; Nouri, K. Merkel cell carcinoma: An updated review of pathogenesis, diagnosis and treatment options. Dermatol. Ther. 2022, 35, e15292. [Google Scholar] [CrossRef]
- Hashida, Y.; Nakajima, K.; Nakajima, H.; Shiga, T.; Tanaka, M.; Murakami, M.; Matsuzaki, S.; Naganuma, S.; Kuroda, N.; Seki, Y.; et al. High load of Merkel cell polyomavirus DNA detected in the normal skin of Japanese patients with Merkel cell carcinoma. J. Clin. Virol. 2016, 82, 101–107. [Google Scholar] [CrossRef]
- Dimitraki, M.G.; Sourvinos, G. Merkel cell polyomavirus (MCPyV) and cancers: Emergency bell or false alarm? Cancers 2022, 14, 5548. [Google Scholar] [CrossRef] [PubMed]
- Kassem, A.; Technau, K.; Kurz, A.K.; Pantulu, D.; Löning, M.; Kayser, G.; Stickeler, E.; Weyers, W.; Diaz, C.; Werner, M. Merkel cell polyomavirus sequences are frequently detected in nonmelanoma skin cancer of immunosuppressed patients. Int. J. Cancer 2009, 125, 356–361. [Google Scholar] [CrossRef]
- Csoboz, B.; Rasheed, K.; Sveinbiørnsson, B.; Moens, U. Merkel cell polyomavirus and non-Merkel cell carcinomas: Guilty or circumstantial evidence? APMIS 2020, 128, 104–120. [Google Scholar] [CrossRef] [PubMed]
- Venceslau, M.; da Costa, G.R.M.; Guimarães, M.A.A.M.; Varella, R.B.; Luz, F.B. Quantification of human polyomaviruses MCPyV and HPyV6 in malignant and non-malignant skin lesions. An. Bras. Dermatol. 2023, 98, 198–201. [Google Scholar] [CrossRef]
- Viscidi, R.P.; Rollinson, D.E.; Sondak, V.K.; Silver, B.; Messina, J.L.; Giuliano, A.R.; Fulp, W.; Ajidahun, A.; Rivanera, D. Age-specific seroprevalence of Merkel cell polyomavirus, BK virus, and JC virus. Clin. Vaccine Immunol. 2011, 18, 1737–1743. [Google Scholar] [CrossRef] [PubMed]
- Bhutani, M.; Polizzotto, M.N.; Uldrick, T.S.; Yarchoan, R. Kaposi sarcoma-associated herpesvirus-associated malignancies: Epidemiology, pathogenesis, and advances in treatment. Semin. Oncol. 2015, 42, 223–246. [Google Scholar] [CrossRef]
- Mataftsi, M.; Skoura, L.; Sakellari, D. HIV infection and periodontal diseases: An overview of the post-HAART era. Oral. Dis. 2011, 17, 13–25. [Google Scholar] [CrossRef]
- Slots, J. Human viruses in periodontitis. Periodontol. 2010, 2000, 89–110. [Google Scholar] [CrossRef]
- Dai, L.; Lu, Y.C.; Chen, J.; Plaisance-Bonstaff, K.; Mu, S.; Forrest, J.C.; Whitby, D.; Post, S.R.; Qin, Z. Oral Shedding of an Oncogenic Virus Alters the Oral Microbiome in HIV+ Patients. Front. Microbiol. 2022, 13, 882520. [Google Scholar] [CrossRef]
- Markazi, A.; Meng, W.; Bracci, P.M.; McGrath, M.S.; Gao, S.J. The Role of Bacteria in KSHV Infection and KSHV-Induced Cancers. Cancers 2021, 13, 4269. [Google Scholar] [CrossRef]
- Gorres, K.L.; Daigle, D.; Mohanram, S.; Miller, G. Activation and repression of Epstein-Barr Virus and Kaposi’s sarcoma-associated herpesvirus lytic cycles by short- and medium-chain fatty acids. J. Virol. 2014, 88, 8028–8044. [Google Scholar] [CrossRef] [PubMed]
- Gruffaz, M.; Zhang, T.; Marshall, V.; Gonçalves, P.; Ramaswami, R.; Labo, N.; Whitby, D.; Uldrick, T.S.; Yarchoan, R.; Huang, Y.; et al. Signatures of oral microbiome in HIV-infected individuals with oral Kaposi’s sarcoma and cell-associated KSHV DNA. PLoS Pathog. 2020, 16, e1008114. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.K.; Kumari, I.; Singh, B.; Sharma, K.K.; Tiwari, S.K. Probiotics, prebiotics and synbiotics: Safe options for next-generation therapeutics. Appl. Microbiol. Biotechnol. 2022, 106, 505–521. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Li, L.; Zhang, H.; Zhao, J.; Lu, W.; Chen, W. Gut Microbiota, Probiotics, and Their Interactions in Prevention and Treatment of Atopic Dermatitis: A Review. Front. Immunol. 2021, 12, 720393. [Google Scholar] [CrossRef]
- Goodarzi, A.; Mozafarpoor, S.; Bodaghabadi, M.; Mohamadi, M. The potential of probiotics for treating acne vulgaris: A review of literature on acne and microbiota. Dermatol. Ther. 2020, 33, e13279. [Google Scholar] [CrossRef]
- Rigon, R.B.; de Freitas, A.C.P.; Bicas, J.L.; Cogo-Müller, K.; Kurebayashi, A.K.; Magalhães, R.F.; Leonardi, G.R. Skin microbiota as a therapeutic target for psoriasis treatment: Trends and perspectives. J. Cosmet. Dermatol. 2021, 20, 1066–1072. [Google Scholar] [CrossRef]
- Yu, Y.; Dunaway, S.; Champer, J.; Kim, J.; Alikhan, A. Changing our microbiome: Probiotics in dermatology. Br. J. Dermatol. 2020, 182, 39–46. [Google Scholar] [CrossRef]
- Valdez, J.; Peral, M.; Rachid, M.; Santana, M.; Perdigon, G. Interference of Lactobacillus plantarum with Pseudomonas aeruginosa in vitro and in infected burns: The potential use of probiotics in wound treatment. Clin. Microbiol. Infect. 2005, 11, 472–479. [Google Scholar] [CrossRef]
- Riaz Rajoka, M.S.; Shi, J.; Zhu, J.; Shao, D.; Huang, Q.; Yang, H.; Jin, M. Capacity of lactic acid cells in immune enhancement and cancer prevention. Appl. Microbiol. Biotechnol. 2017, 101, 35–45. [Google Scholar] [CrossRef]
- Garbacz, K. Anticancer activity of lactic acid bacteriae. Semin. Cancer Biol. 2022, 86, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.C.; Lee, I.J.; Huang, C.; Chang, T.M. Lactic Acid Bacteria and Lactic Acid for Skin Health and Melanogenesis Inhibition. Curr. Pharm. Biotechnol. 2020, 21, 566–577. [Google Scholar] [CrossRef]
- Hosseini, S.; Imenshahidi, M.; Hosseinzadeh, H.; Karimi, G. Effects of plant extracts and bioactive compounds on attenuation of bleomycin-induced pulmonary fibrosis. Biomed. Pharmacother. 2018, 107, 1454–1465. [Google Scholar] [CrossRef] [PubMed]
- Blohm-Mangone, K.; Burkett, N.B.; Tahsin, S.; Myrdal, P.B.; Aodah, A.; Ho, B.; Janda, J.; McComas, M.; Saboda, K.; Roe, D.J. Pharmacological TLR4 antagonism using topical resatorvid blocks solar UV-induced skin tumorigenesis in SKH-1 mice. Cancer Prev. Res. 2018, 11, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.; Miller, N.J.; Lu, H.; Longino, N.V.; Ibrani, D.; Shinohara, M.M.; Byrd, D.R.; Parvathaneni, U.; Kulikauskas, R.; Ter Meulen, J. Intratumoral G100, a TLR4 agonist, induces antitumor immune responses and tumor regression in patients with merkel cell carcinoma. Clin. Cancer Res. 2019, 25, 1185–1195. [Google Scholar] [CrossRef] [PubMed]
- Burns, E.M.; Yusuf, N. Toll-like receptors and skin cancer. Front. Immunol. 2014, 5, 135. [Google Scholar] [CrossRef]
- Le, M.; Ghazawi, F.M.; Netchiporouk, E.; Litvinov, I.V. The Novel Role of Antibiotic Treatment in the Management of Cutaneous T-Cell Lymphoma (CTCL) Patients. J. Cutan. Med. Surg. 2020, 24, 410–411. [Google Scholar] [CrossRef]
- Lindahl, L.M.; Willerslev-Olsen, A.; Gjerdrum, L.M.R.; Nielsen, P.R.; Blümel, E.; Rittig, A.H.; Celis, P.; Herpers, B.; Becker, J.C.; Stausbøl-Grøn, B.; et al. Antibiotics Inhibit Tumor and Disease Activity in Cutaneous T-Cell Lymphoma. Blood 2019, 134, 1072–1083. [Google Scholar] [CrossRef]
- Lindahl, L.M.; Iversen, L.; Ødum, N.; Kilian, M. Staphylococcus aureus and Antibiotics in Cutaneous T-Cell Lymphoma. Dermatology 2021, 238, 3. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savoia, P.; Azzimonti, B.; Rolla, R.; Zavattaro, E. Role of the Microbiota in Skin Neoplasms: New Therapeutic Horizons. Microorganisms 2023, 11, 2386. https://doi.org/10.3390/microorganisms11102386
Savoia P, Azzimonti B, Rolla R, Zavattaro E. Role of the Microbiota in Skin Neoplasms: New Therapeutic Horizons. Microorganisms. 2023; 11(10):2386. https://doi.org/10.3390/microorganisms11102386
Chicago/Turabian StyleSavoia, Paola, Barbara Azzimonti, Roberta Rolla, and Elisa Zavattaro. 2023. "Role of the Microbiota in Skin Neoplasms: New Therapeutic Horizons" Microorganisms 11, no. 10: 2386. https://doi.org/10.3390/microorganisms11102386
APA StyleSavoia, P., Azzimonti, B., Rolla, R., & Zavattaro, E. (2023). Role of the Microbiota in Skin Neoplasms: New Therapeutic Horizons. Microorganisms, 11(10), 2386. https://doi.org/10.3390/microorganisms11102386