
The Low-FODMAP Diet, IBS, and BCFAs: Exploring the Positive, Negative, and Less Desirable Aspects—A Literature Review

 ,
,  ,
,  ,
,  ,
,  ,
, 
 and
and 
Abstract
:1. Introduction
The Role of BCFAs in Intestinal Inflammation
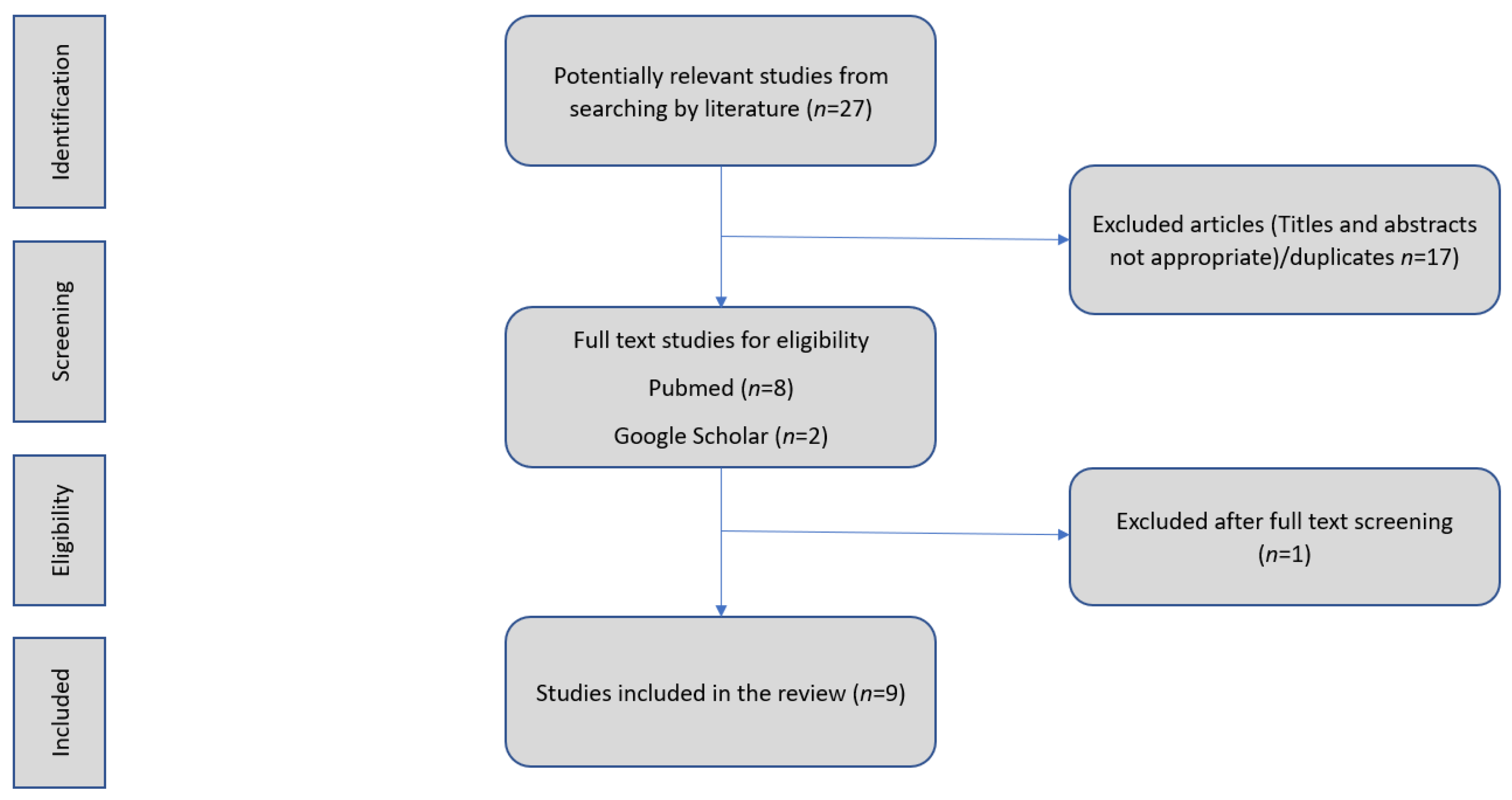
2. Literature Search
2.1. BCFAs & IBS
Human Studies, Table 1

{kind=link}
{kind=link}
| References | Type of Study | Subjects | Samples | Outcomes |
|---|---|---|---|---|
| Le Gall et al., 2011 [47] | Cohort | UC patients (n = 13). IBS patients (n = 10). Healthy controls (n = 22). | Feces | ↑ 2-methylbutyrate, isobutyrate, isovalerate in control vs. IBS group. |
| Farup et al., 2016 [48] | Case–control | IBS patients (All subtypes, n = 25). Healthy controls (n = 25). | Feces | No statistically significant difference in isovaleric or isobutyric between IBS and control. |
| Zhang et al., 2019 [49] | Case–control | IBS-D patients (n = 30). Healthy controls (n = 15). | Feces | ↑ Isobutyrate IBS-D (no statistically significant different). ↑ Isovalerate in IBS-D correlated with severity of abdominal pain. |
2.2. BCFAs, Inflammation, and Low-FODMAP Diet
2.2.1. Preclinical Trials (Table 2)
| References | Subjects | Intervention | Samples | Outcomes |
|---|---|---|---|---|
| Tuck et al., 2019 [51] | Mice (n = 35) | 3 groups Positive control + low/high FODMAP (n = 12). Negative control + low/high FODMAP (n = 12). Post-inflammatory + low/high FODMAP (n = 11). | Feces | In positive control and post-inflammatory groups that followed LFD: ↑ BCFAs; ↑ Isovalerate and isobutyric. |
| Tuck et al., 2020 [52] | Mice (n = 40) | 4 groups Group A (n = 10) LabDiet 5066—Sacrificed at baseline. Group B (n = 10) LabDiet 5066. Group C (n = 10) ResearchDiets AIN93G (lower FODMAP content). Group D (n = 10) LabDiet 5001 (higher FODMAP content). | Feces | Group C: ↑ BCFAs; ↑ Isovalerate and isobutyric. |
2.2.2. Clinical Trials (Table 3)
| References | Subjects | Intervention | Samples | Outcomes |
|---|---|---|---|---|
| Halmos et al., 2014 [53] | IBS patients (all subtypes, n = 27), healthy controls (n = 6) | Habitual diet | Feces | ↑ Isobutyrate and isovalerate in healthy controls. |
| 2 groups LFD (3.05 g FODMAP). Australian diet (23.7 g). | Feces | No statistically significant difference in BCFAs among the groups. | ||
| Wilson et al., 2020 [54] | IBS patients (n = 69) | 3 groups Sham diet with placebo supplement (control) (n = 23). LFD supplemented with placebo (n = 22). LFD supplemented with 1.4 g/d B-GOS (n = 24). | Feces | No statistically significant difference in isobutyrate and isovalerate in LFD with or without supplements of B-GOS. |
| Zhang et al., 2021 [55] | IBS-D patients (n = 100) | 2 groups LFD (n = 51). TDA (n = 49). | Feces | In LFD group: ↑ Isobutyrate and isovalerate. |
| Nordin et al., 2023 [56] | IBS patients (n = 103) | 3 groups 1. Placebo–gluten–FODMAPs (n = 35). 2. FODMAPs–placebo–gluten (n = 33). 3. Gluten–FODMAPs–placebo (n = 35). | Feces and plasma | No statistically significant difference in isobutyrate among groups (feces). ↓ Isovalerate after gluten vs. placebo. ↓ Isobutyrate after FODMAPs compared to the placebo (plasma). |
2.3. BCFAs as Potentially Harmful Metabolites
3. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BCAAs | Branched-chain amino acids |
| BCFA | Branched-chain fatty acid |
| B-GOS | β-Galactooligosaccharide |
| CTT | Colonic transit time |
| DSS | Dextran sulfate sodium |
| EC | Enterochromaffin cells |
| FFA2, FFA3 | Free fatty acid receptors 2,3 |
| FMT | Fecal microbiota transplantation |
| GI | Gastrointestinal tract |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| HDAC | Histone deacetylase |
| IBS | Irritable bowel syndrome |
| IBS-C | IBS with constipation |
| IBS-D | IBS with diarrhea |
| IBS-M | IBS mixed |
| IFN | Interferon |
| ILs | Interleukins |
| LFD | Low-FODMAP diet |
| MPO | Myeloperoxidase |
| NEC | Necrotizing enterocolitis |
| NMR | Nuclear magnetic resonance |
| NO | Nitric oxide |
| PKA | Protein kinase A |
| SCFA | Short-chain fatty acids |
| TEER | Transepithelial electrical resistance |
| TNF-a | Tumor Necrosis Factor -a |
| UC | Ulcerative colitis |
| ZO-1 | Zonulin-1 |
References
- Van Malderen, K.; De Winter, B.Y.; De Man, J.G.; De Schepper, H.U.; Lamote, K. Volatomics in inflammatory bowel disease and irritable bowel syndrome. EBioMedicine 2020, 54, 102725. [Google Scholar] [CrossRef] [PubMed]
- Ezzine, C.; Loison, L.; Montbrion, N.; Bôle-Feysot, C.; Déchelotte, P.; Coëffier, M.; Ribet, D. Fatty acids produced by the gut microbiota dampen host inflammatory responses by modulating intestinal SUMOylation. Gut Microbes 2022, 14, 2108280. [Google Scholar] [CrossRef] [PubMed]
- Ringel-Kulka, T.; Choi, C.H.; Temas, D.; Kim, A.; Maier, D.M.; Scott, K.; Galanko, J.A.; Ringel, Y. Altered Colonic Bacterial Fermentation as a Potential Pathophysiological Factor in Irritable Bowel Syndrome. Am. J. Gastroenterol. 2015, 110, 1339–1346. [Google Scholar] [CrossRef] [PubMed]
- Ran-Ressler, R.R.; Devapatla, S.; Lawrence, P.; Brenna, J.T. Branched chain fatty acids are constituents of the normal healthy newborn gastrointestinal tract. Pediatr. Res. 2008, 64, 605–609. [Google Scholar] [CrossRef]
- Ran-Ressler, R.R.; Glahn, R.P.; Bae, S.; Brenna, J.T. Branched-chain fatty acids in the neonatal gut and estimated dietary intake in infancy and adulthood. Nestle Nutr. Inst. Workshop Ser. 2013, 77, 133–143. [Google Scholar] [CrossRef]
- Gozdzik, P.; Magkos, F.; Sledzinski, T.; Mika, A. Monomethyl branched-chain fatty acids: Health effects and biological mechanisms. Prog. Lipid Res. 2023, 90, 101226. [Google Scholar] [CrossRef]
- Ramos Meyers, G.; Samouda, H.; Bohn, T. Short Chain Fatty Acid Metabolism in Relation to Gut Microbiota and Genetic Variability. Nutrients 2022, 14, 5361. [Google Scholar] [CrossRef]
- Fan, L.; Xia, Y.; Wang, Y.; Han, D.; Liu, Y.; Li, J.; Fu, J.; Wang, L.; Gan, Z.; Liu, B.; et al. Gut microbiota bridges dietary nutrients and host immunity. Sci. China Life Sci. 2023, 1–49. [Google Scholar] [CrossRef]
- Musch, M.W.; Bookstein, C.; Xie, Y.; Sellin, J.H.; Chang, E.B. SCFA increase intestinal Na absorption by induction of NHE3 in rat colon and human intestinal C2/bbe cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 280, G687–G693. [Google Scholar] [CrossRef]
- Aguirre, M.; Eck, A.; Koenen, M.E.; Savelkoul, P.H.M.; Budding, A.E.; Venema, K. Diet drives quick changes in the metabolic activity and composition of human gut microbiota in a validated in vitro gut model. Res. Microbiol. 2016, 167, 114–125. [Google Scholar] [CrossRef]
- Procházková, N.; Falony, G.; Dragsted, L.O.; Licht, T.R.; Raes, J.; Roager, H.M. Advancing human gut microbiota research by considering gut transit time. Gut 2023, 72, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Goyal, A.; Wang, T.; Dubinkina, V.; Maslov, S. Ecology-guided prediction of cross-feeding interactions in the human gut microbiome. Nat. Commun. 2021, 12, 1335. [Google Scholar] [CrossRef] [PubMed]
- Kasti, A.; Petsis, K.; Lambrinou, S.; Katsas, K.; Nikolaki, M.; Papanikolaou, I.S.; Hatziagelaki, E.; Triantafyllou, K. A Combination of Mediterranean and Low-FODMAP Diets for Managing IBS Symptoms? Ask Your Gut! Microorganisms 2022, 10, 751. [Google Scholar] [CrossRef] [PubMed]
- Bellini, M.; Tonarelli, S.; Nagy, A.G.; Pancetti, A.; Costa, F.; Ricchiuti, A.; de Bortoli, N.; Mosca, M.; Marchi, S.; Rossi, A. Low FODMAP Diet: Evidence, Doubts, and Hopes. Nutrients 2020, 12, 148. [Google Scholar] [CrossRef]
- Spiller, R. Impact of Diet on Symptoms of the Irritable Bowel Syndrome. Nutrients 2021, 13, 575. [Google Scholar] [CrossRef] [PubMed]
- Gibson, P.R. History of the low FODMAP diet. J. Gastroenterol. Hepatol. 2017, 32 (Suppl. 1), 5–7. [Google Scholar] [CrossRef] [PubMed]
- Gershon, M.D.; Tack, J. The serotonin signaling system: From basic understanding to drug development for functional GI disorders. Gastroenterology 2007, 132, 397–414. [Google Scholar] [CrossRef]
- Camilleri, M.; Boeckxstaens, G. Dietary and pharmacological treatment of abdominal pain in IBS. Gut 2017, 66, 966–974. [Google Scholar] [CrossRef]
- Hustoft, T.N.; Hausken, T.; Ystad, S.O.; Valeur, J.; Brokstad, K.; Hatlebakk, J.G.; Lied, G.A. Effects of varying dietary content of fermentable short-chain carbohydrates on symptoms, fecal microenvironment, and cytokine profiles in patients with irritable bowel syndrome. Neurogastroenterol. Motil. 2017, 29, e12969. [Google Scholar] [CrossRef]
- Staudacher, H.M.; Irving, P.M.; Lomer, M.C.E.; Whelan, K. Mechanisms and efficacy of dietary FODMAP restriction in IBS. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 256–266. [Google Scholar] [CrossRef]
- Staudacher, H.M.; Lomer, M.C.E.; Anderson, J.L.; Barrett, J.S.; Muir, J.G.; Irving, P.M.; Whelan, K. Fermentable carbohydrate restriction reduces luminal bifidobacteria and gastrointestinal symptoms in patients with irritable bowel syndrome. J. Nutr. 2012, 142, 1510–1518. [Google Scholar] [CrossRef]
- Staudacher, H.M.; Whelan, K. Altered gastrointestinal microbiota in irritable bowel syndrome and its modification by diet: Probiotics, prebiotics and the low FODMAP diet. Proc. Nutr. Soc. 2016, 75, 306–318. [Google Scholar] [CrossRef] [PubMed]
- Kolodziejczyk, A.A.; Zheng, D.; Elinav, E. Diet-microbiota interactions and personalized nutrition. Nat. Rev. Microbiol. 2019, 17, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Fredericks, E.; Theunissen, R.; Roux, S. Short chain fatty acids and monocarboxylate transporters in irritable bowel syndrome. Turk. J. Gastroenterol. 2020, 31, 840–847. [Google Scholar] [CrossRef]
- Gilbert, M.S.; Ijssennagger, N.; Kies, A.K.; van Mil, S.W.C. Protein fermentation in the gut; implications for intestinal dysfunction in humans, pigs, and poultry. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G159–G170. [Google Scholar] [CrossRef]
- Yao, C.K.; Muir, J.G.; Gibson, P.R. Review article: Insights into colonic protein fermentation, its modulation and potential health implications. Aliment. Pharmacol. Ther. 2016, 43, 181–196. [Google Scholar] [CrossRef] [PubMed]
- Davila, A.-M.; Blachier, F.; Gotteland, M.; Andriamihaja, M.; Benetti, P.-H.; Sanz, Y.; Tomé, D. Intestinal luminal nitrogen metabolism: Role of the gut microbiota and consequences for the host. Pharmacol. Res. 2013, 68, 95–107. [Google Scholar] [CrossRef]
- Dong, L.; Li, Y.; Chen, Q.; Liu, Y.; Qiao, Z.; Sang, S.; Zhang, J.; Zhan, S.; Wu, Z.; Liu, L. Research advances of advanced glycation end products in milk and dairy products: Formation, determination, control strategy and immunometabolism via gut microbiota. Food Chem. 2023, 417, 135861. [Google Scholar] [CrossRef]
- Rios-Covian, D.; González, S.; Nogacka, A.M.; Arboleya, S.; Salazar, N.; Gueimonde, M.; de Los Reyes-Gavilán, C.G. An Overview on Fecal Branched Short-Chain Fatty Acids Along Human Life and as Related with Body Mass Index: Associated Dietary and Anthropometric Factors. Front. Microbiol. 2020, 11, 973. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Ghyselinck, J.; Marzorati, M.; Koch, A.-M.; Lambert, W.; Michiels, J.; Chalvon-Demersay, T. The Effect of Amino Acids on Production of SCFA and bCFA by Members of the Porcine Colonic Microbiota. Microorganisms 2022, 10, 762. [Google Scholar] [CrossRef]
- Gasaly, N.; Hermoso, M.A.; Gotteland, M. Butyrate and the Fine-Tuning of Colonic Homeostasis: Implication for Inflammatory Bowel Diseases. Int. J. Mol. Sci. 2021, 22, 3061. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H. Immune regulation by microbiome metabolites. Immunology 2018, 154, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.; Chen, Y.; Ma, Z.; Zhang, X.; Shi, D.; Khan, J.A.; Liu, H. Gut microbiota-derived short chain fatty acids are potential mediators in gut inflammation. Anim. Nutr. (Zhongguo Xu Mu Shou Yi Xue Hui) 2022, 8, 350–360. [Google Scholar] [CrossRef]
- Kaneda, T. Iso- and anteiso-fatty acids in bacteria: Biosynthesis, function, and taxonomic significance. Microbiol. Rev. 1991, 55, 288–302. [Google Scholar] [CrossRef]
- Yan, Y.; Wang, Z.; Greenwald, J.; Kothapalli, K.S.D.; Park, H.G.; Liu, R.; Mendralla, E.; Lawrence, P.; Wang, X.; Brenna, J.T. BCFA suppresses LPS induced IL-8 mRNA expression in human intestinal epithelial cells. Prostaglandins. Leukot. Essent. Fat. Acids 2017, 116, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, Z.; Park, H.G.; Xu, C.; Lawrence, P.; Su, X.; Wijendran, V.; Walker, W.A.; Kothapalli, K.S.D.; Brenna, J.T. Human fetal intestinal epithelial cells metabolize and incorporate branched chain fatty acids in a structure specific manner. Prostaglandins Leukot. Essent. Fat. Acids 2017, 116, 32–39. [Google Scholar] [CrossRef]
- Yan, Y.; Wang, Z.; Wang, D.; Lawrence, P.; Wang, X.; Kothapalli, K.S.D.; Greenwald, J.; Liu, R.; Park, H.G.; Brenna, J.T. BCFA-enriched vernix-monoacylglycerol reduces LPS-induced inflammatory markers in human enterocytes in vitro. Pediatr. Res. 2018, 83, 874–879. [Google Scholar] [CrossRef]
- Bosnar, L.M.; Shindler, A.E.; Wood, J.; Patch, C.; Franks, A.E. The Potential to Lower the Incidence of Necrotising Enterocolitis with Branched Chain Fatty Acids. J. Clin. Gastroenterol. Treat. 2022, 8, 85. [Google Scholar] [CrossRef]
- Yi, C.; Sun, W.; Ding, L.; Yan, M.; Sun, C.; Qiu, C.; Wang, D.; Wu, L. Short-Chain Fatty Acids Weaken Ox-LDL-Induced Cell Inflammatory Injury by Inhibiting the NLRP3/Caspase-1 Pathway and Affecting Cellular Metabolism in THP-1 Cells. Molecules 2022, 27, 8801. [Google Scholar] [CrossRef]
- Taormina, V.M.; Unger, A.L.; Schiksnis, M.R.; Torres-Gonzalez, M.; Kraft, J. Branched-Chain Fatty Acids-An Underexplored Class of Dairy-Derived Fatty Acids. Nutrients 2020, 12, 2875. [Google Scholar] [CrossRef]
- Ran-Ressler, R.R.; Khailova, L.; Arganbright, K.M.; Adkins-Rieck, C.K.; Jouni, Z.E.; Koren, O.; Ley, R.E.; Brenna, J.T.; Dvorak, B. Branched chain fatty acids reduce the incidence of necrotizing enterocolitis and alter gastrointestinal microbial ecology in a neonatal rat model. PLoS ONE 2011, 6, e29032. [Google Scholar] [CrossRef] [PubMed]
- Russell, W.R.; Gratz, S.W.; Duncan, S.H.; Holtrop, G.; Ince, J.; Scobbie, L.; Duncan, G.; Johnstone, A.M.; Lobley, G.E.; Wallace, R.J.; et al. High-protein, reduced-carbohydrate weight-loss diets promote metabolite profiles likely to be detrimental to colonic health. Am. J. Clin. Nutr. 2011, 93, 1062–1072. [Google Scholar] [CrossRef]
- Boudry, G.; Jamin, A.; Chatelais, L.; Gras-Le Guen, C.; Michel, C.; Le Huërou-Luron, I. Dietary protein excess during neonatal life alters colonic microbiota and mucosal response to inflammatory mediators later in life in female pigs. J. Nutr. 2013, 143, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Carco, C.; Young, W.; Gearry, R.B.; Talley, N.J.; McNabb, W.C.; Roy, N.C. Increasing Evidence That Irritable Bowel Syndrome and Functional Gastrointestinal Disorders Have a Microbial Pathogenesis. Front. Cell. Infect. Microbiol. 2020, 10, 468. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, W.; Wang, L.; Yu, C.; Zhang, G.; Zhu, H.; Wang, C.; Zhao, S.; Hu, C.-A.A.; Liu, Y. Lentinan modulates intestinal microbiota and enhances barrier integrity in a piglet model challenged with lipopolysaccharide. Food Funct. 2019, 10, 479–489. [Google Scholar] [CrossRef]
- He, X.; Sun, W.; Ge, T.; Mu, C.; Zhu, W. An increase in corn resistant starch decreases protein fermentation and modulates gut microbiota during in vitro cultivation of pig large intestinal inocula. Anim. Nutr. (Zhongguo Xu Mu Shou Yi Xue Hui) 2017, 3, 219–224. [Google Scholar] [CrossRef]
- Le Gall, G.; Noor, S.O.; Ridgway, K.; Scovell, L.; Jamieson, C.; Johnson, I.T.; Colquhoun, I.J.; Kemsley, E.K.; Narbad, A. Metabolomics of fecal extracts detects altered metabolic activity of gut microbiota in ulcerative colitis and irritable bowel syndrome. J. Proteome Res. 2011, 10, 4208–4218. [Google Scholar] [CrossRef]
- Farup, P.G.; Rudi, K.; Hestad, K. Faecal short-chain fatty acids—A diagnostic biomarker for irritable bowel syndrome? BMC Gastroenterol. 2016, 16, 51. [Google Scholar] [CrossRef]
- Zhang, W.-X.; Zhang, Y.; Qin, G.; Li, K.-M.; Wei, W.; Li, S.-Y.; Yao, S.-K. Altered profiles of fecal metabolites correlate with visceral hypersensitivity and may contribute to symptom severity of diarrhea-predominant irritable bowel syndrome. World J. Gastroenterol. 2019, 25, 6416–6429. [Google Scholar] [CrossRef]
- El-Salhy, M.; Valeur, J.; Hausken, T.; Gunnar Hatlebakk, J. Changes in fecal short-chain fatty acids following fecal microbiota transplantation in patients with irritable bowel syndrome. Neurogastroenterol. Motil. 2021, 33, e13983. [Google Scholar] [CrossRef]
- Tuck, C.J.; Caminero, A.; Jiménez Vargas, N.N.; Soltys, C.L.; Jaramillo Polanco, J.O.; Lopez Lopez, C.D.; Constante, M.; Lourenssen, S.R.; Verdu, E.F.; Muir, J.G.; et al. The impact of dietary fermentable carbohydrates on a postinflammatory model of irritable bowel syndrome. Neurogastroenterol. Motil. 2019, 31, e13675. [Google Scholar] [CrossRef] [PubMed]
- Tuck, C.J.; De Palma, G.; Takami, K.; Brant, B.; Caminero, A.; Reed, D.E.; Muir, J.G.; Gibson, P.R.; Winterborn, A.; Verdu, E.F.; et al. Nutritional profile of rodent diets impacts experimental reproducibility in microbiome preclinical research. Sci. Rep. 2020, 10, 17784. [Google Scholar] [CrossRef] [PubMed]
- Halmos, E.P.; Christophersen, C.T.; Bird, A.R.; Shepherd, S.J.; Gibson, P.R.; Muir, J.G. Diets that differ in their FODMAP content alter the colonic luminal microenvironment. Gut 2015, 64, 93–100. [Google Scholar] [CrossRef]
- Wilson, B.; Rossi, M.; Kanno, T.; Parkes, G.C.; Anderson, S.; Mason, A.J.; Irving, P.M.; Lomer, M.C.; Whelan, K. β-Galactooligosaccharide in Conjunction with Low FODMAP Diet Improves Irritable Bowel Syndrome Symptoms but Reduces Fecal Bifidobacteria. Am. J. Gastroenterol. 2020, 115, 906–915. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Feng, L.; Wang, X.; Fox, M.; Luo, L.; Du, L.; Chen, B.; Chen, X.; He, H.; Zhu, S.; et al. Low fermentable oligosaccharides, disaccharides, monosaccharides, and polyols diet compared with traditional dietary advice for diarrhea-predominant irritable bowel syndrome: A parallel-group, randomized controlled trial with analysis of clinical and micr. Am. J. Clin. Nutr. 2021, 113, 1531–1545. [Google Scholar] [CrossRef]
- Nordin, E.; Hellström, P.M.; Dicksved, J.; Pelve, E.; Landberg, R.; Brunius, C. Effects of FODMAPs and Gluten on Gut Microbiota and Their Association with the Metabolome in Irritable Bowel Syndrome: A Double-Blind, Randomized, Cross-Over Intervention Study. Nutrients 2023, 15, 3045. [Google Scholar] [CrossRef]
- Trefflich, I.; Dietrich, S.; Braune, A.; Abraham, K.; Weikert, C. Short- and Branched-Chain Fatty Acids as Fecal Markers for Microbiota Activity in Vegans and Omnivores. Nutrients 2021, 13, 1808. [Google Scholar] [CrossRef] [PubMed]
- Windey, K.; De Preter, V.; Verbeke, K. Relevance of protein fermentation to gut health. Mol. Nutr. Food Res. 2012, 56, 184–196. [Google Scholar] [CrossRef]
- Wang, X.; Gibson, G.R.; Costabile, A.; Sailer, M.; Theis, S.; Rastall, R.A. Prebiotic Supplementation of In Vitro Fecal Fermentations Inhibits Proteolysis by Gut Bacteria, and Host Diet Shapes Gut Bacterial Metabolism and Response to Intervention. Appl. Environ. Microbiol. 2019, 85, e02749-18. [Google Scholar] [CrossRef]
- Salazar, N.; Arboleya, S.; Fernández-Navarro, T.; de Los Reyes-Gavilán, C.G.; Gonzalez, S.; Gueimonde, M. Age-Associated Changes in Gut Microbiota and Dietary Components Related with the Immune System in Adulthood and Old Age: A Cross-Sectional Study. Nutrients 2019, 11, 1765. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikolaki, M.D.; Kasti, A.N.; Katsas, K.; Petsis, K.; Lambrinou, S.; Patsalidou, V.; Stamatopoulou, S.; Karlatira, K.; Kapolos, J.; Papadimitriou, K.; et al. The Low-FODMAP Diet, IBS, and BCFAs: Exploring the Positive, Negative, and Less Desirable Aspects—A Literature Review. Microorganisms 2023, 11, 2387. https://doi.org/10.3390/microorganisms11102387
Nikolaki MD, Kasti AN, Katsas K, Petsis K, Lambrinou S, Patsalidou V, Stamatopoulou S, Karlatira K, Kapolos J, Papadimitriou K, et al. The Low-FODMAP Diet, IBS, and BCFAs: Exploring the Positive, Negative, and Less Desirable Aspects—A Literature Review. Microorganisms. 2023; 11(10):2387. https://doi.org/10.3390/microorganisms11102387
Chicago/Turabian StyleNikolaki, Maroulla D., Arezina N. Kasti, Konstantinos Katsas, Konstantinos Petsis, Sophia Lambrinou, Vasiliki Patsalidou, Sophia Stamatopoulou, Katerina Karlatira, John Kapolos, Konstantinos Papadimitriou, and et al. 2023. "The Low-FODMAP Diet, IBS, and BCFAs: Exploring the Positive, Negative, and Less Desirable Aspects—A Literature Review" Microorganisms 11, no. 10: 2387. https://doi.org/10.3390/microorganisms11102387
APA StyleNikolaki, M. D., Kasti, A. N., Katsas, K., Petsis, K., Lambrinou, S., Patsalidou, V., Stamatopoulou, S., Karlatira, K., Kapolos, J., Papadimitriou, K., & Triantafyllou, K. (2023). The Low-FODMAP Diet, IBS, and BCFAs: Exploring the Positive, Negative, and Less Desirable Aspects—A Literature Review. Microorganisms, 11(10), 2387. https://doi.org/10.3390/microorganisms11102387