One Health Approach to Leishmaniases: Understanding the Disease Dynamics through Diagnostic Tools


 ,
, 
Abstract
1. Introduction
2. Clinical Manifestations
2.1. Cutaneous/Mucocutaneous Leishmaniases
2.2. Visceral Leishmaniasis
2.3. Post-Kala-Azar Dermal Leishmaniasis
2.4. Asymptomatic Infection
3. Risk Factors for Leishmaniases
3.1. Socioeconomic Factors and Malnutrition
3.2. Migration
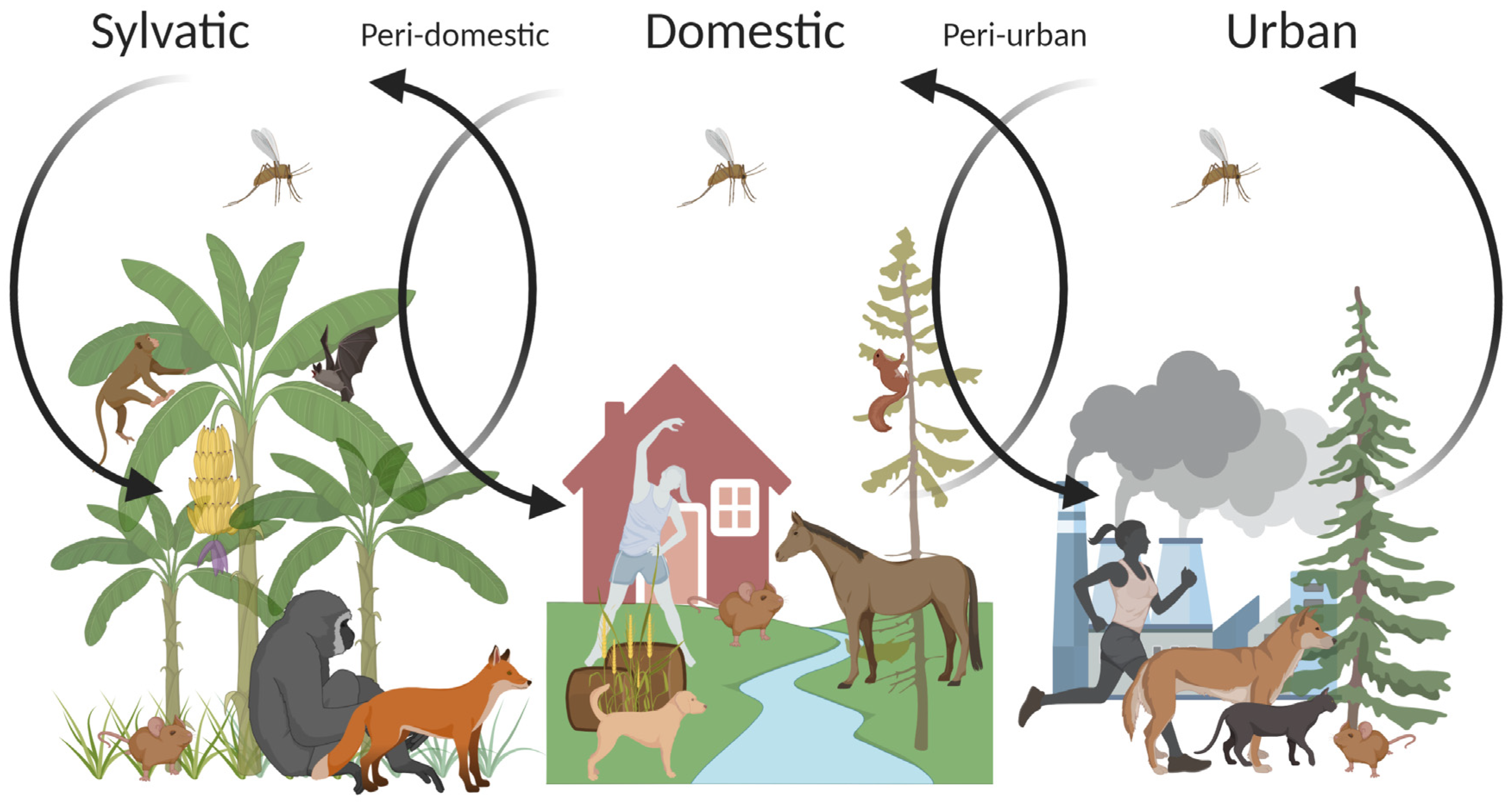
3.3. Environmental Changes
4. Diagnostic Tools for Leishmaniases
4.1. Parasitological Diagnoses
4.2. Molecular Diagnosis of Parasites
4.3. Immunological Diagnoses
5. Use of Diagnostic Tools in Epidemiological Studies
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- WHO. Control of the Leishmaniases: Report of a Meeting of the WHO Expert Committee on the Control of Leishmaniases, 949th ed.; WHO Press: Geneva, Switzerland, 2010. [Google Scholar]
- Alvar, J.; Velez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; den Boer, M. Leishmaniasis worldwide and global estimates of its incidence. PLoS ONE 2012, 7, e35671. [Google Scholar] [CrossRef]
- Palatnik-de-Sousa, C.B.; Santos, W.R.; França-Silva, J.C.; da Costa, R.T.; Barbosa Reis, A.; Palatnik, M.; Mayrink, W.; Genaro, O. Impact of canine control on the epidemiology of canine and human visceral leishmaniasis in Brazil. Am. Soc. Trop. Med. Hyg. 2001, 65, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Hubálek, Z. Emerging Human Infectious Diseases: Anthroponoses, Zoonoses, and Sapronoses. Emerg. Infect. Dis. 2003, 9, 403–404. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, O.A.; Serrano, M.G.; Camargo, E.P.; Teixeira, M.M.G.; Shaw, J.J. An appraisal of the taxonomy and nomenclature of trypanosomatids presently classified as Leishmania and Endotrypanum. Parasitology 2018, 145, 430–442. [Google Scholar] [CrossRef] [PubMed]
- Khanal, B.; Picado, A.; Bhattarai, N.R.; Van Der Auwera, G.; Das, M.L.; Ostyn, B.; Davies, C.R.; Boelaert, M.; Dujardin, J.C.; Rijal, S. Spatial analysis of Leishmania donovani exposure in humans and domestic animals in a recent kala azar focus in Nepal. Parasitology 2010, 137, 1597–1603. [Google Scholar] [CrossRef]
- Bhattarai, N.R.; Van der Auwera, G.; Rijal, S.; Picado, A.; Speybroeck, N.; Khanal, B.; De Doncker, S.; Das, M.L.; Ostyn, B.; Davies, C.; et al. Domestic animals and epidemiology of visceral leishmaniasis, Nepal. Emerg. Infect. Dis. 2010, 16, 231–237. [Google Scholar] [CrossRef]
- Talmi-Frank, D.; Kedem-Vaanunu, N.; King, R.; Bar-Gal, G.K.; Edery, N.; Jaffe, C.L.; Baneth, G. Leishmania tropica infection in golden jackals and red foxes, Israel. Emerg. Infect. Dis. 2010, 16, 1973–1975. [Google Scholar] [CrossRef]
- Lainson, R. Ecological interactions in the transmission of the leishmaniases. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1988, 321, 389–404. [Google Scholar]
- Colmenares, M.; Kar, S.; Goldsmith-Pestana, K.; McMahon-Pratt, D. Mechanisms of pathogenesis: Differences amongst Leishmania species. Trans. R. Soc. Trop. Med. Hyg. 2002, 96 (Suppl. 1), S3–S7. [Google Scholar] [CrossRef]
- Rangel, E.F.; Shaw, J.J. (Eds.) Brazilian Sand Flies: Biology, Taxonomy, Medical Importance and Control, 1st ed.; Springer International Publishing: Cham, Switzerland, 2018; Volume 1, p. 494. [Google Scholar]
- Andrade, M.S.; Courtenay, O.; Brito, M.E.; Carvalho, F.G.; Carvalho, A.W.; Soares, F.; Carvalho, S.M.; Costa, P.L.; Zampieri, R.; Floeter-Winter, L.M.; et al. Infectiousness of Sylvatic and Synanthropic Small Rodents Implicates a Multi-host Reservoir of Leishmania (Viannia) braziliensis. PLoS. Negl. Trop. Dis. 2015, 9, e0004137. [Google Scholar] [CrossRef]
- Badaró, R.; Jones, T.C.; Lorenço, R.; Cerf, B.J.; Sampaio, D.; Carvalho, E.M.; Rocha, H.; Teixeira, R.; Johnson, W.D., Jr. A prospective study of visceral leishmaniasis in an endemic area of Brazil. J. Infect. Dis. 1986, 154, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Sang, R.; Arum, S.; Chepkorir, E.; Mosomtai, G.; Tigoi, C.; Sigei, F.; Lwande, O.W.; Landmann, T.; Affognon, H.; Ahlm, C.; et al. Distribution and abundance of key vectors of Rift Valley fever and other arboviruses in two ecologically distinct counties in Kenya. PLoS. Negl. Trop. Dis. 2017, 11, e0005341. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.H.; Latham, S.M.; Woolhouse, M.E. Risk factors for human disease emergence. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2001, 356, 983–989. [Google Scholar] [CrossRef] [PubMed]
- CDC. One Health Basics. Available online: https://www.cdc.gov/onehealth/basics/index.html (accessed on 18 April 2020).
- Kahn, L.H. The need for one health degree programs. Infect. Ecol. Epidemiol. 2011, 1. [Google Scholar] [CrossRef] [PubMed]
- Gebreyes, W.A.; Dupouy-Camet, J.; Newport, M.J.; Oliveira, C.J.; Schlesinger, L.S.; Saif, Y.M.; Kariuki, S.; Saif, L.J.; Saville, W.; Wittum, T.; et al. The global one health paradigm: Challenges and opportunities for tackling infectious diseases at the human, animal, and environment interface in low-resource settings. PLoS Negl. Trop. Dis. 2014, 8, e3257. [Google Scholar] [CrossRef]
- Institute of Medicine Committee on Emerging Microbial Threats to H. In Emerging Infections: Microbial Threats to Health in the United States; Lederberg, J., Shope, R.E., Oaks, S.C., Jr., Eds.; National Academies Press (US): Washington, DC, USA, 1992. [Google Scholar]
- Institute of Medicine (US) Committee on Emerging Microbial Threats to Health in the 21st Century; National Academies Press (US): Washington, DC, USA, 2003; p. 367.
- Kumar, N.P.; Srinivasan, R.; Anish, T.S.; Nandakumar, G.; Jambulingam, P. Cutaneous leishmaniasis caused by Leishmania donovani in the tribal population of the Agasthyamala Biosphere Reserve forest, Western Ghats, Kerala, India. J. Med. Microbiol. 2015, 64 Pt 2, 157–163. [Google Scholar] [CrossRef]
- Sharma, N.L.; Mahajan, V.K.; Kanga, A.; Sood, A.; Katoch, V.M.; Mauricio, I.; Singh, C.D.; Parwan, U.C.; Sharma, V.K.; Sharma, R.C. Localized cutaneous leishmaniasis due to Leishmania donovani and Leishmania tropica: Preliminary findings of the study of 161 new cases from a new endemic focus in himachal pradesh, India. Am. J. Trop. Med. Hyg. 2005, 72, 819–824. [Google Scholar] [CrossRef]
- Khatri, M.L.; Di Muccio, T.; Fiorentino, E.; Gramiccia, M. Ongoing outbreak of cutaneous leishmaniasis in northwestern Yemen: Clinicoepidemiologic, geographic, and taxonomic study. Int. J. Dermatol. 2016, 55, 1210–1218. [Google Scholar] [CrossRef]
- Del Giudice, P.; Marty, P.; Lacour, J.P.; Perrin, C.; Pratlong, F.; Haas, H.; Dellamonica, P.; Le Fichoux, Y. Cutaneous leishmaniasis due to Leishmania infantum. Case reports and literature review. Arch. Dermatol. 1998, 134, 193–198. [Google Scholar] [CrossRef]
- Crowe, A.; Slavin, J.; Stark, D.; Aboltins, C. A case of imported Leishmania infantum cutaneous leishmaniasis; an unusual presentation occurring 19 years after travel. BMC Infect. Dis. 2014, 14, 597. [Google Scholar] [CrossRef]
- Convit, J.; Ulrich, M.; Pérez, M.; Hung, J.; Castillo, J.; Rojas, H.; Viquez, A.; Araya, L.N.; Lima, H.D. Atypical cutaneous leishmaniasis in Central America: Possible interaction between infectious and environmental elements. Trans. R. Soc. Trop. Med. Hyg. 2005, 99, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Gitari, J.W.; Nzou, S.M.; Wamunyokoli, F.; Kinyeru, E.; Fujii, Y.; Kaneko, S.; Mwau, M. Leishmaniasis recidivans by Leishmania tropica in Central Rift Valley Region in Kenya. Int. J. Infect. Dis. 2018, 74, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Afonso, L.C.; Scott, P. Immune responses associated with susceptibility of C57BL/10 mice to Leishmania amazonensis. Infect. Immun. 1993, 61, 2952–2959. [Google Scholar] [CrossRef] [PubMed]
- Aoki, J.I.; Laranjeira-Silva, M.F.; Muxel, S.M.; Floeter-Winter, L.M. The impact of arginase activity on virulence factors of Leishmania amazonensis. Curr. Opin. Microbiol. 2019, 52, 110–115. [Google Scholar] [CrossRef]
- Scorza, B.M.; Carvalho, E.M.; Wilson, M.E. Cutaneous Manifestations of Human and Murine Leishmaniasis. Int. J. Mol. Sci. 2017, 18, 1296. [Google Scholar] [CrossRef]
- Scott, P.; Novais, F.O. Cutaneous leishmaniasis: Immune responses in protection and pathogenesis. Nat. Rev. Immunol. 2016, 16, 581–592. [Google Scholar] [CrossRef]
- Tomiotto-Pellissier, F.; Bortoleti, B.; Assolini, J.P.; Gonçalves, M.D.; Carloto, A.C.M.; Miranda-Sapla, M.M.; Conchon-Costa, I.; Bordignon, J.; Pavanelli, W.R. Macrophage Polarization in Leishmaniasis: Broadening Horizons. Front. Immunol. 2018, 9, 2529. [Google Scholar] [CrossRef]
- Yanik, M.; Gurel, M.S.; Simsek, Z.; Kati, M. The psychological impact of cutaneous leishmaniasis. Clin. Exp. Dermatol. 2004, 29, 464–467. [Google Scholar] [CrossRef]
- Reithinger, R.; Dujardin, J.C.; Louzir, H.; Pirmez, C.; Alexander, B.; Brooker, S. Cutaneous leishmaniasis. Lancet Infect. Dis. 2007, 7, 581–596. [Google Scholar] [CrossRef]
- Carvalho, E.M.; Barral, A.; Costa, J.M.; Bittencourt, A.; Marsden, P. Clinical and immunopathological aspects of disseminated cutaneous leishmaniasis. Acta Trop. 1994, 56, 315–325. [Google Scholar] [CrossRef]
- Develoux, M.; Diallo, S.; Dieng, Y.; Mane, I.; Huerre, M.; Pratlong, F.; Dedet, J.P.; Ndiaye, B. Diffuse cutaneous leishmaniasis due to Leishmania major in Senegal. Trans. R. Soc. Trop. Med. Hyg. 1996, 90, 396–397. [Google Scholar] [CrossRef]
- Alcover, M.M.; Rocamora, V.; Guillén, M.C.; Berenguer, D.; Cuadrado, M.; Riera, C.; Fisa, R. Case Report: Diffuse Cutaneous Leishmaniasis by Leishmania infantum in a Patient Undergoing Immunosuppressive Therapy: Risk Status in an Endemic Mediterranean Area. Am. J. Trop. Med. Hyg. 2018, 98, 1313–1316. [Google Scholar] [CrossRef] [PubMed]
- Sangueza, O.P.; Sangueza, J.M.; Stiller, M.J.; Sangueza, P. Mucocutaneous leishmaniasis: A clinicopathologic classification. J. Am. Acad. Dermatol. 1993, 28, 927–932. [Google Scholar] [CrossRef]
- Dassoni, F.; Daba, F.; Naafs, B.; Morrone, A. Leishmaniasis recidivans in Ethiopia: Cutaneous and mucocutaneous features. J. Infect. Dev. Ctries. 2017, 11, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Davies, C.R.; Reithinger, R.; Campbell-Lendrum, D.; Feliciangeli, D.; Borges, R.; Rodriguez, N. The epidemiology and control of leishmaniasis in Andean countries. Cadernos de Saude Publica 2000, 16, 925–950. [Google Scholar] [CrossRef]
- Marsden, P.D. Mucosal leishmaniasis (“espundia” Escomel, 1911). Trans. R. Soc. Trop. Med. Hyg. 1986, 80, 859–876. [Google Scholar] [CrossRef]
- Osorio, L.E.; Castillo, C.M.; Ochoa, M.T. Mucosal leishmaniasis due to Leishmania (Viannia) panamensis in Colombia: Clinical characteristics. Am. J. Trop. Med. Hyg. 1998, 59, 49–52. [Google Scholar] [CrossRef]
- David, C.; Dimier-David, L.; Vargas, F.; Torrez, M.; Dedet, J.P. Fifteen years of cutaneous and mucocutaneous leishmaniasis in Bolivia: A retrospective study. Trans. R. Soc. Trop. Med. Hyg. 1993, 87, 7–9. [Google Scholar] [CrossRef]
- WHO. Leishmaniasis: Epidemiological Situation. Available online: https://www.who.int/leishmaniasis/burden/en/ (accessed on 1 May 2020).
- Machado-Coelho, G.L.; Caiaffa, W.T.; Genaro, O.; Magalhães, P.A.; Mayrink, W. Risk factors for mucosal manifestation of American cutaneous leishmaniasis. Trans. R. Soc. Trop. Med. Hyg. 2005, 99, 55–61. [Google Scholar] [CrossRef]
- Shaw, J.J. Further thoughts on the use of the name Leishmania (Leishmania) infantum chagasi for the aetiological agent of American visceral leishmaniasis. Memorias do Instituto Oswaldo Cruz 2006, 101, 577–579. [Google Scholar] [CrossRef]
- Dantas-Torres, F. Leishmania infantum versus Leishmania chagasi: Do not forget the law of priority. Memorias do Instituto Oswaldo Cruz 2006, 101, 117–118, discussion 118. [Google Scholar] [CrossRef] [PubMed]
- Killick-Kendrick, R. Some epidemiological consequences of the evolutionary fit between Leishmaniae and their phlebotomine vectors. Bull. Soc. Pathol. Exot. Filiales. 1985, 78 Pt 2, 747–755. [Google Scholar]
- Rioux, J.A.; Lanotte, G.; Serres, E.; Pratlong, F.; Bastien, P.; Perieres, J. Taxonomy of Leishmania. Use of isoenzymes. Suggestions for a new classification. Ann. Parasitol. Hum. Comp. 1990, 65, 111–125. [Google Scholar] [CrossRef]
- Lainson, R.; Rangel, E.F. Lutzomyia longipalpis and the eco-epidemiology of American visceral leishmaniasis, with particular reference to Brazil: A review. Memorias do Instituto Oswaldo Cruz 2005, 100, 811–827. [Google Scholar] [CrossRef] [PubMed]
- Lukes, J.; Mauricio, I.L.; Schonian, G.; Dujardin, J.C.; Soteriadou, K.; Dedet, J.P.; Kuhls, K.; Tintaya, K.W.; Jirku, M.; Chocholova, E.; et al. Evolutionary and geographical history of the Leishmania donovani complex with a revision of current taxonomy. Proc. Natl. Acad. Sci. USA 2007, 104, 9375–9380. [Google Scholar] [CrossRef] [PubMed]
- Alborzi, A.; Rasouli, M.; Shamsizadeh, A. Leishmania tropica-isolated patient with visceral leishmaniasis in southern Iran. Am. J. Trop. Med. Hyg. 2006, 74, 306–307. [Google Scholar] [CrossRef]
- Alborzi, A.; Pouladfar, G.R.; Fakhar, M.; Motazedian, M.H.; Hatam, G.R.; Kadivar, M.R. Isolation of Leishmania tropica from a patient with visceral leishmaniasis and disseminated cutaneous leishmaniasis, southern Iran. Am. J. Trop. Med. Hyg. 2008, 79, 435–437. [Google Scholar] [CrossRef][Green Version]
- Sarkari, B.; Bavarsad Ahmadpour, N.; Moshfe, A.; Hajjaran, H. Molecular Evaluation of a Case of Visceral Leishmaniasis Due to Leishmania tropica in Southwestern Iran. Iran. J. Parasitol. 2016, 11, 126–130. [Google Scholar]
- Magill, A.J.; Grögl, M.; Gasser, R.A., Jr.; Sun, W.; Oster, C.N. Visceral infection caused by Leishmania tropica in veterans of Operation Desert Storm. N. Engl. J. Med. 1993, 328, 1383–1387. [Google Scholar] [CrossRef]
- Singh, O.P.; Hasker, E.; Sacks, D.; Boelaert, M.; Sundar, S. Asymptomatic Leishmania infection: A new challenge for Leishmania control. Clin. Infect. Dis. 2014, 58, 1424–1429. [Google Scholar] [CrossRef]
- Stauch, A.; Sarkar, R.R.; Picado, A.; Ostyn, B.; Sundar, S.; Rijal, S.; Boelaert, M.; Dujardin, J.C.; Duerr, H.P. Visceral leishmaniasis in the Indian subcontinent: Modelling epidemiology and control. PLoS. Negl. Trop. Dis. 2011, 5, e1405. [Google Scholar] [CrossRef] [PubMed]
- Alvar, J.; Aparicio, P.; Aseffa, A.; Den Boer, M.; Cañavate, C.; Dedet, J.P.; Gradoni, L.; Ter Horst, R.; López-Vélez, R.; Moreno, J. The relationship between leishmaniasis and AIDS: The second 10 years. Clin. Microbiol. Rev. 2008, 21, 334–359. [Google Scholar] [CrossRef] [PubMed]
- Barral, A.; Pedral-Sampaio, D.; Grimaldi, D., Jr.; Momen, H.; McMahon-Pratt, D.; de Jesus, A.R.; Almeida, R.; Badaro, R.; Barral-Netto, M.; Carvalho, E.M.; et al. Leishmaniasis in Bahia, Brazil: Evidence that Leishmania amazonensis Produces a Wide Spectrum of Clinical Disease. Am. J. Trop. Med. Hyg. 1991, 44, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Aliaga, L.; Ceballos, J.; Sampedro, A.; Cobo, F.; López-Nevot, M.; Merino-Espinosa, G.; Morillas-Márquez, F.; Martín-Sánchez, J. Asymptomatic Leishmania infection in blood donors from the Southern of Spain. Infection 2019, 47, 739–747. [Google Scholar] [CrossRef]
- França, A.O.; Pompilio, M.A.; Pontes, E.; de Oliveira, M.P.; Pereira, L.O.R.; Lima, R.B.; Goto, H.; Sanchez, M.C.A.; Fujimori, M.; Lima-Júnior, M.; et al. Leishmania infection in blood donors: A new challenge in leishmaniasis transmission? PLoS ONE 2018, 13, e0198199. [Google Scholar] [CrossRef]
- Pineda, J.A.; Macías, J.; Morillas, F.; Fernandez-Ochoa, J.; Cara, J.; de La Rosa, R.; Mira, J.A.; Martín-Sánchez, J.; González, M.; Delgado, J.; et al. Evidence of increased risk for leishmania infantum infection among HIV-seronegative intravenous drug users from southern Spain. Eur. J. Clin. Microbiol. Infect. Dis. 2001, 20, 354–357. [Google Scholar] [CrossRef]
- Vida, B.; Toepp, A.; Schaut, R.G.; Esch, K.J.; Juelsgaard, R.; Shimak, R.M.; Petersen, C.A. Immunologic progression of canine leishmaniosis following vertical transmission in United States dogs. Vet. Immunol. Immunopathol. 2016, 169, 34–38. [Google Scholar] [CrossRef]
- Eltoum, I.A.; Zijlstra, E.E.; Ali, M.S.; Ghalib, H.W.; Satti, M.M.; Eltoum, B.; el-Hassan, A.M. Congenital kala-azar and leishmaniasis in the placenta. Am. J. Trop. Med. Hyg. 1992, 46, 57–62. [Google Scholar] [CrossRef]
- Berger, B.A.; Bartlett, A.H.; Saravia, N.G.; Galindo Sevilla, N. Pathophysiology of Leishmania Infection during Pregnancy. Trends Parasitol. 2017, 33, 935–946. [Google Scholar] [CrossRef]
- Barrett, M.P.; Croft, S.L. Management of trypanosomiasis and leishmaniasis. Br. Med. Bull. 2012, 104, 175–196. [Google Scholar] [CrossRef]
- Zijlstra, E.E.; Musa, A.M.; Khalil, E.A.; el-Hassan, I.M.; el-Hassan, A.M. Post-kala-azar dermal leishmaniasis. Lancet Infect. Dis. 2003, 3, 87–98. [Google Scholar] [CrossRef]
- Stark, D.; Pett, S.; Marriott, D.; Harkness, J. Post-kala-azar dermal leishmaniasis due to Leishmania infantum in a human immunodeficiency virus type 1-infected patient. J. Clin. Microbiol. 2006, 44, 1178–1180. [Google Scholar] [CrossRef] [PubMed]
- Rathi, S.K.; Pandhi, R.K.; Khanna, N.; Chopra, P. Mucosal and peri-orificial involvement in post-kala-azar dermal leishmaniasis. Indian J. Dermatol. Venereol. Leprol. 2004, 70, 280–282. [Google Scholar] [PubMed]
- Zijlstra, E.E.; Alves, F.; Rijal, S.; Arana, B.; Alvar, J. Post-kala-azar dermal leishmaniasis in the Indian subcontinent: A threat to the South-East Asia Region Kala-azar Elimination Programme. PLoS Negl. Trop. Dis. 2017, 11, e0005877. [Google Scholar] [CrossRef]
- Burza, S.; Croft, S.L.; Boelaert, M. Leishmaniasis. Lancet 2018, 392, 951–970. [Google Scholar] [CrossRef]
- Ganguly, S.; Saha, P.; Chatterjee, M.; Roy, S.; Ghosh, T.K.; Guha, S.K.; Kundu, P.K.; Bera, D.K.; Basu, N.; Maji, A.K. PKDL--A Silent Parasite Pool for Transmission of Leishmaniasis in Kala-azar Endemic Areas of Malda District, West Bengal, India. PLoS Negl. Trop. Dis. 2015, 9, e0004138. [Google Scholar] [CrossRef]
- Molina, R.; Ghosh, D.; Carrillo, E.; Monnerat, S.; Bern, C.; Mondal, D.; Alvar, J. Infectivity of Post-Kala-azar Dermal Leishmaniasis Patients to Sand Flies: Revisiting a Proof of Concept in the Context of the Kala-azar Elimination Program in the Indian Subcontinent. Clin. Infect. Dis. 2017, 65, 150–153. [Google Scholar] [CrossRef]
- Andrade-Narvaez, F.J.; Loría-Cervera, E.N.; Sosa-Bibiano, E.I.; Van Wynsberghe, N.R. Asymptomatic infection with American cutaneous leishmaniasis: Epidemiological and immunological studies. Memorias do Instituto Oswaldo Cruz 2016, 111, 599–604. [Google Scholar] [CrossRef]
- Da Costa-Val, A.P.; Cavalcanti, R.R.; de Figueiredo Gontijo, N.; Michalick, M.S.; Alexander, B.; Williams, P.; Melo, M.N. Canine visceral leishmaniasis: Relationships between clinical status, humoral immune response, haematology and Lutzomyia (Lutzomyia) longipalpis infectivity. Vet. J. 2007, 174, 636–643. [Google Scholar] [CrossRef]
- Topno, R.K.; Das, V.N.; Ranjan, A.; Pandey, K.; Singh, D.; Kumar, N.; Siddiqui, N.A.; Singh, V.P.; Kesari, S.; Bimal, S.; et al. Asymptomatic infection with visceral leishmaniasis in a disease-endemic area in bihar, India. Am. J. Trop. Med. Hyg. 2010, 83, 502–506. [Google Scholar] [CrossRef]
- Schaefer, K.U.; Kurtzhals, J.A.; Gachihi, G.S.; Muller, A.S.; Kager, P.A. A prospective sero-epidemiological study of visceral leishmaniasis in Baringo District, Rift Valley Province, Kenya. Trans. R. Soc. Trop. Med. Hyg. 1995, 89, 471–475. [Google Scholar] [CrossRef]
- Saha, P.; Ganguly, S.; Chatterjee, M.; Das, S.B.; Kundu, P.K.; Guha, S.K.; Ghosh, T.K.; Bera, D.K.; Basu, N.; Maji, A.K. Asymptomatic leishmaniasis in kala-azar endemic areas of Malda district, West Bengal, India. PLoS Negl. Trop. Dis. 2017, 11, e0005391. [Google Scholar] [CrossRef] [PubMed]
- Ostyn, B.; Gidwani, K.; Khanal, B.; Picado, A.; Chappuis, F.; Singh, S.P.; Rijal, S.; Sundar, S.; Boelaert, M. Incidence of symptomatic and asymptomatic Leishmania donovani infections in high-endemic foci in India and Nepal: A prospective study. PLoS Negl. Trop. Dis. 2011, 5, e1284. [Google Scholar] [CrossRef]
- Evans, T.G.; Teixeira, M.J.; McAuliffe, I.T.; Vasconcelos, I.; Vasconcelos, A.W.; Sousa Ade, A.; Lima, J.W.; Pearson, R.D. Epidemiology of visceral leishmaniasis in northeast Brazil. J. Infect. Dis. 1992, 166, 1124–1132. [Google Scholar] [CrossRef] [PubMed]
- Maia, Z.; Viana, V.; Muniz, E.; Gonçalves, L.O.; Mendes, C.M.; Mehta, S.R.; Badaro, R. Risk Factors Associated with Human Visceral Leishmaniasis in an Urban Area of Bahia, Brazil. Vector Borne Zoonotic Dis. 2016, 16, 368–376. [Google Scholar] [CrossRef]
- Davies, C.R.; Mazloumi Gavgani, A.S. Age, acquired immunity and the risk of visceral leishmaniasis: A prospective study in Iran. Parasitology 1999, 119 Pt 3, 247–257. [Google Scholar] [CrossRef]
- Fukutani, K.F.; Figueiredo, V.; Celes, F.S.; Cristal, J.R.; Barral, A.; Barral-Netto, M.; de Oliveira, C.I. Serological survey of Leishmania infection in blood donors in Salvador, Northeastern Brazil. BMC Infect. Dis. 2014, 14, 422. [Google Scholar] [CrossRef]
- Arrow, K.J.; Panosian, C.B.; Gelband, H. (Eds.) Saving Lives, Buying Time: Economics of Malaria Drugs in an Age of Resistance; National Academies Press (US): Washington, DC, USA, 2004; Volume 2. [Google Scholar]
- Alvar, J.; Yactayo, S.; Bern, C. Leishmaniasis and poverty. Trends Parasitol. 2006, 22, 552–557. [Google Scholar] [CrossRef]
- Rijal, S.; Koirala, S.; Van der Stuyft, P.; Boelaert, M. The economic burden of visceral leishmaniasis for households in Nepal. Trans. R. Soc. Trop. Med. Hyg. 2006, 100, 838–841. [Google Scholar] [CrossRef]
- Reithinger, R.; Aadil, K.; Kolaczinski, J.; Mohsen, M.; Hami, S. Social impact of leishmaniasis, Afghanistan. Emerg. Infect. Dis. 2005, 11, 634–636. [Google Scholar] [CrossRef]
- Anstead, G.M.; Chandrasekar, B.; Zhao, W.; Yang, J.; Perez, L.E.; Melby, P.C. Malnutrition alters the innate immune response and increases early visceralization following Leishmania donovani infection. Infect. Immun. 2001, 69, 4709–4718. [Google Scholar] [CrossRef]
- Saha, S.; Ramachandran, R.; Hutin, Y.J.; Gupte, M.D. Visceral leishmaniasis is preventable in a highly endemic village in West Bengal, India. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 737–742. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.C.; Campbell-Lendrum, D.H.; Lozovei, A.L.; Teodoro, U.; Davies, C.R. Phlebotomine sandfly responses to carbon dioxide and human odour in the field. Med. Vet. Entomol. 2001, 15, 132–139. [Google Scholar] [CrossRef]
- Tavares, D.D.S.; Salgado, V.R.; Miranda, J.C.; Mesquita, P.R.R.; Rodrigues, F.M.; Barral-Netto, M.; de Andrade, J.B.; Barral, A. Attraction of phlebotomine sandflies to volatiles from skin odors of individuals residing in an endemic area of tegumentary leishmaniasis. PLoS ONE 2018, 13, e0203989. [Google Scholar] [CrossRef] [PubMed]
- Bern, C.; Hightower, A.W.; Chowdhury, R.; Ali, M.; Amann, J.; Wagatsuma, Y.; Haque, R.; Kurkjian, K.; Vaz, L.E.; Begum, M.; et al. Risk factors for kala-azar in Bangladesh. Emerg. Infect. Dis. 2005, 11, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Bantie, K.; Tessema, F.; Tafere, Y. Factors Associated with Visceral Leishmaniasis Infection in North Gondar Zone, Amhara Region, North West Ethiopia, Case Control Study. Sci. J. Public Health 2014, 2, 560. [Google Scholar]
- Islam, A.; Rahman, M.H.; Islam, S.; Debnath, P.; Alam, M.; Hassan, M. Sero-prevalence of visceral leishmaniasis (VL) among dogs in VL endemic areas of Mymensingh district, Bangladesh. J. Adv. Vet. Anim. Res. 2018, 4, 241–248. [Google Scholar] [CrossRef]
- Al-Salem, W.; Herricks, J.R.; Hotez, P.J. A review of visceral leishmaniasis during the conflict in South Sudan and the consequences for East African countries. Parasit. Vectors 2016, 9, 460. [Google Scholar] [CrossRef]
- Alawieh, A.; Musharrafieh, U.; Jaber, A.; Berry, A.; Ghosn, N.; Bizri, A.R. Revisiting leishmaniasis in the time of war: The Syrian conflict and the Lebanese outbreak. Int. J. Infect. Dis. 2014, 29, 115–119. [Google Scholar] [CrossRef]
- Saúde, M.D. Manual de Vigilância e Controle da Leishmaniose Visceral, 1st ed.; Editora do Ministério da Saúde: Brasília-DF, Brasil, 2006; p. 120.
- Cardim, M.F.; Rodas, L.A.; Dibo, M.R.; Guirado, M.M.; Oliveira, A.M.; Chiaravalloti-Neto, F. Introduction and expansion of human American visceral leishmaniasis in the state of Sao Paulo, Brazil, 1999–2011. Rev. Saude Publica 2013, 47, 691–700. [Google Scholar] [CrossRef]
- Barata, R.B. Cem anos de endemias e epidemias. Ciênc Saúde Coletiva 2000, 5, 333–345. [Google Scholar] [CrossRef]
- Barrett, T.V.; Senra, M.S. Leishmaniasis in Manaus, Brazil. Parasitol. Today 1989, 5, 255–257. [Google Scholar] [CrossRef]
- Defries, R.; Rudel, T.; Uriarte, M.; Hansen, M. Deforestation driven by urban population growth and agricultural trade in the twenty-first century. Nat. Geosci. 2010, 3, 178–181. [Google Scholar] [CrossRef]
- Desjeux, P. Leishmaniasis: Current situation and new perspectives. Comp. Immunol. Microbiol. Infect. Dis. 2004, 27, 305–318. [Google Scholar] [CrossRef] [PubMed]
- Desjeux, P. The increase in risk factors for leishmaniasis worldwide. Trans. R. Soc. Trop. Med. Hyg. 2001, 95, 239–243. [Google Scholar] [CrossRef]
- Penna, G.; Pinto, L.F.; Soranz, D.; Glatt, R. High incidence of diseases endemic to the Amazon region of Brazil, 2001-2006. Emerg. Infect. Dis. 2009, 15, 626–632. [Google Scholar] [CrossRef]
- Warburg, A.; Montoya-Lerma, J.; Jaramillo, C.; Cruz-Ruiz, A.L.; Ostrovska, K. Leishmaniasis vector potential of Lutzomyia spp. in Colombian coffee plantations. Med. Vet. Entomol. 1991, 5, 9–16. [Google Scholar] [CrossRef]
- Alexander, B.; Oliveria, E.B.; Haigh, E.; Almeida, L.L. Transmission of Leishmania in coffee plantations of Minas Gerais, Brazil. Memórias do Instituto Oswaldo Cruz 2002, 97, 627–630. [Google Scholar] [CrossRef]
- Alexander, B.; Agudelo, L.A.; Navarro, J.F.; Ruiz, J.F.; Molina, J.; Aguilera, G.; Klein, A.; Quiñones, M.L. Relationship between coffee cultivation practices in Colombia and exposure to infection with Leishmania. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 1263–1268. [Google Scholar] [CrossRef]
- Barhoumi, W.; Chelbi, I.; Zhioua, E. Effects of the development of irrigation systems in the arid areas on the establishment of Phlebotomus (Larroussius) perfiliewi Parrot, 1939. Bull. Soc. Pathol. Exot. 2012, 105, 403–405. [Google Scholar] [CrossRef]
- Zhioua, E.; Kaabi, B.; Chelbi, I. Entomological investigations following the spread of visceral leishmaniasis in Tunisia. J. Vector. Ecol. 2007, 32, 371–374. [Google Scholar] [CrossRef]
- Vink, M.M.T.; Nahzat, S.M.; Rahimi, H.; Buhler, C.; Ahmadi, B.A.; Nader, M.; Zazai, F.R.; Yousufzai, A.S.; van Loenen, M.; Schallig, H.; et al. Evaluation of point-of-care tests for cutaneous leishmaniasis diagnosis in Kabul, Afghanistan. EBioMedicine 2018, 37, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, D.D. (Ed.) On the Presence of Peculiar Parasitic Organisms in the Tissue of a Specimen of Delhi; Superintendent of Government: Calcutta, India, 1885; Volume 1, pp. 21–31.
- Ross, R. Further notes on leishman’s bodies. Br. Med. J. 1903, 2, 1401. [Google Scholar] [CrossRef] [PubMed]
- Ross, R. Note on the bodies recently described by leishman and donovan. Br. Med. J. 1903, 2, 1261–1262. [Google Scholar] [CrossRef] [PubMed]
- Row, R. A simple haemoglobinized saline culture medium: For the growth of leishmania and allied protozoa. Br. Med. J. 1912, 1, 1119–1120. [Google Scholar] [CrossRef][Green Version]
- Forkner, C.E.; Zia, L.S. Further studies on kala-azar: Leishmania in nasal and oral secretions of patients and the bearing of this finding on the transmission of the disease. J. Exp. Med. 1935, 61, 183–203. [Google Scholar] [CrossRef]
- Allahverdiyev, A.M.; Uzun, S.; Bagirova, M.; Durdu, M.; Memisoglu, H.R. A sensitive new microculture method for diagnosis of cutaneous leishmaniasis. Am. J. Trop. Med. Hyg. 2004, 70, 294–297. [Google Scholar] [CrossRef]
- Van Eys, G.J.; Schoone, G.J.; Ligthart, G.S.; Laarman, J.J.; Terpstra, W.J. Detection of Leishmania parasites by DNA in situ hybridization with non-radioactive probes. Parasitol. Res. 1987, 73, 199–202. [Google Scholar] [CrossRef]
- Rodgers, M.R.; Popper, S.J.; Wirth, D.F. Amplification of kinetoplast DNA as a tool in the detection and diagnosis of Leishmania. Exp. Parasitol. 1990, 71, 267–275. [Google Scholar] [CrossRef]
- Arnot, D.E.; Barker, D.C. Biochemical identification of cutaneous leishmanias by analysis of kinetoplast DNA. II. Sequence homologies in Leishmania kDNA. Mol. Biochem. Parasitol. 1981, 3, 47–56. [Google Scholar] [CrossRef]
- Tibayrenc, M.; Neubauer, K.; Barnabé, C.; Guerrini, F.; Skarecky, D.; Ayala, F.J. Genetic characterization of six parasitic protozoa: Parity between random-primer DNA typing and multilocus enzyme electrophoresis. Proc. Natl. Acad. Sci. USA 1993, 90, 1335–1339. [Google Scholar] [CrossRef] [PubMed]
- Qubain, H.I.; Saliba, E.K.; Oskam, L. Visceral leishmaniasis from Bal’a, Palestine, caused by Leishmania donovani s.1. identified through polymerase chain reaction and restriction fragment length polymorphism analysis. Acta Trop. 1997, 68, 121–128. [Google Scholar] [CrossRef]
- Noyes, H.A.; Reyburn, H.; Bailey, J.W.; Smith, D. A nested-PCR-based schizodeme method for identifying Leishmania kinetoplast minicircle classes directly from clinical samples and its application to the study of the epidemiology of Leishmania tropica in Pakistan. J. Clin. Microbiol. 1998, 36, 2877–2881. [Google Scholar] [CrossRef] [PubMed]
- Harris, E.; Kropp, G.; Belli, A.; Rodriguez, B.; Agabian, N. Single-step multiplex PCR assay for characterization of New World Leishmania complexes. J. Clin. Microbiol. 1998, 36, 1989–1995. [Google Scholar] [CrossRef]
- Belli, A.; Rodriguez, B.; Aviles, H.; Harris, E. Simplified polymerase chain reaction detection of new world Leishmania in clinical specimens of cutaneous leishmaniasis. Am. J. Trop. Med. Hyg. 1998, 58, 102–109. [Google Scholar] [CrossRef]
- Robert-Gangneux, F.; Baixench, M.T.; Piarroux, R.; Pratlong, F.; Tourte-Schaefer, C. Use of molecular tools for the diagnosis and typing of a Leishmania major strain isolated from an HIV-infected patient in Burkina Faso. Trans. R. Soc. Trop. Med. Hyg. 1999, 93, 396–397. [Google Scholar] [CrossRef]
- Bretagne, S.; Durand, R.; Olivi, M.; Garin, J.F.; Sulahian, A.; Rivollet, D.; Vidaud, M.; Deniau, M. Real-time PCR as a new tool for quantifying Leishmania infantum in liver in infected mice. Clin. Diagn. Lab Immunol. 2001, 8, 828–831. [Google Scholar] [CrossRef]
- Takagi, H.; Itoh, M.; Islam, M.Z.; Razzaque, A.; Ekram, A.R.; Hashighuchi, Y.; Noiri, E.; Kimura, E. Sensitive, specific, and rapid detection of Leishmania donovani DNA by loop-mediated isothermal amplification. Am. J. Trop. Med. Hyg. 2009, 81, 578–582. [Google Scholar] [CrossRef]
- Nasereddin, A.; Jaffe, C.L. Rapid diagnosis of Old World Leishmaniasis by high-resolution melting analysis of the 7SL RNA gene. J. Clin. Microbiol. 2010, 48, 2240–2242. [Google Scholar] [CrossRef]
- Mouri, O.; Morizot, G.; Van der Auwera, G.; Ravel, C.; Passet, M.; Chartrel, N.; Joly, I.; Thellier, M.; Jauréguiberry, S.; Caumes, E.; et al. Easy identification of leishmania species by mass spectrometry. PLoS Negl. Trop. Dis. 2014, 8, e2841. [Google Scholar] [CrossRef]
- Montenegro, J. A cutis-reação na Leishmaniose. Anais da Faculdade de Medicina de São Paulo 1926, 1, 9. [Google Scholar]
- Ghosh, H.; Ghosh, N.N. Complement-fixation reaction in sera of rabbits actively immunized with living culture of Leishmania donovani. Ann. Biochem. Exp. Med. 1947, 7, 1. [Google Scholar] [PubMed]
- Ghosh, H.; Ghosh, N.N. Agglutination reaction in sera of rabbits immunized with different strains of Leishmania donovani. Ann. Biochem. Exp. Med. 1947, 7, 3–6. [Google Scholar] [PubMed]
- Duxbury, R.E.; Sadun, E.H. Fluorescent antibody test for the serodiagnosis of visceral leishmaniasis. Am. J. Trop. Med. Hyg. 1964, 13, 525–529. [Google Scholar] [CrossRef]
- Shaw, J.J.; Voller, A. The detection of circulating antibody to kala-azar by means of immunofluorescent techniques. Trans. R. Soc. Trop. Med. Hyg. 1964, 58, 349–352. [Google Scholar] [CrossRef]
- Baldelli, B.; Orfei, A.B.; Fioretti, D.P.; Polidori, G.A.; Ambrosi, M. Serological diagnosis of human leishmaniasis by ELISA (enzyme-linked immunosorbent assay). Parassitologia 1978, 20, 91–99. [Google Scholar]
- Pappas, M.G.; Hajkowski, R.; Hockmeyer, W.T. Dot enzyme-linked immunosorbent assay (Dot-ELISA): A micro technique for the rapid diagnosis of visceral leishmaniasis. J. Immunol. Methods 1983, 64, 205–214. [Google Scholar] [CrossRef]
- Jaffe, C.L.; Bennett, E.; Grimaldi, G., Jr.; McMahon-Pratt, D. Production and characterization of species-specific monoclonal antibodies against Leishmania donovani for immunodiagnosis. J. Immunol. 1984, 133, 440–447. [Google Scholar]
- Handman, E.; Mitchell, G.F.; Goding, J.W. Leishmania major: A very sensitive dot-blot ELISA for detection of parasites in cutaneous lesions. Mol. Biol. Med. 1987, 4, 377–383. [Google Scholar]
- Reed, S.G.; Shreffler, W.G.; Burns, J.M., Jr.; Scott, J.M.; Orge Mda, G.; Ghalib, H.W.; Siddig, M.; Badaro, R. An improved serodiagnostic procedure for visceral leishmaniasis. Am. J. Trop. Med. Hyg. 1990, 43, 632–639. [Google Scholar] [CrossRef]
- Sundar, S.; Pai, K.; Sahu, M.; Kumar, V.; Murray, H.W. Immunochromatographic strip-test detection of anti-K39 antibody in Indian visceral leishmaniasis. Ann. Trop. Med. Parasitol. 2002, 96, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Boelaert, M.; Bhattacharya, S.; Chappuis, F.; El Safi, S.H.; Hailu, A.; Mondal, D.; Rijal, S.; Sundar, S.; Wasunna, M.; Peeling, R.W. Evaluation of rapid diagnostic tests: Visceral leishmaniasis. Nat. Rev. Microbiol. 2007, 5, S30–S39. [Google Scholar] [CrossRef]
- Walton, B.C.; Shaw, J.J.; Lainson, R. Observations on the in vitro cultivation of Leishmania braziliensis. J. Parasitol. 1977, 63, 1118–1119. [Google Scholar] [CrossRef] [PubMed]
- Thakur, S.; Joshi, J.; Kaur, S. Leishmaniasis diagnosis: An update on the use of parasitological, immunological and molecular methods. J. Parasit. Dis. 2020, 44, 1–20. [Google Scholar] [CrossRef]
- Patrinos, G.P.; Ansorge, W.J. Molecular Diagnostic, 2nd ed.; Academic Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Uliana, S.R.; Ishikawa, E.; Stempliuk, V.A.; de Souza, A.; Shaw, J.J.; Floeter-Winter, L.M. Geographical distribution of neotropical Leishmania of the subgenus Leishmania analysed by ribosomal oligonucleotide probes. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 261–264. [Google Scholar] [CrossRef]
- Brandão-Filho, S.P.; Brito, M.E.; Carvalho, F.G.; Ishikawa, E.A.; Cupolillo, E.; Floeter-Winter, L.; Shaw, J.J. Wild and synanthropic hosts of Leishmania (Viannia) braziliensis in the endemic cutaneous leishmaniasis locality of Amaraji, Pernambuco State, Brazil. Trans. R. Soc. Trop. Med. Hyg. 2003, 97, 291–296. [Google Scholar] [CrossRef]
- Lopez, M.; Inga, R.; Cangalaya, M.; Echevarria, J.; Llanos-Cuentas, A.; Orrego, C.; Arevalo, J. Diagnosis of Leishmania using the polymerase chain reaction: A simplified procedure for field work. Am. J. Trop. Med. Hyg. 1993, 49, 348–356. [Google Scholar] [CrossRef]
- Disch, J.; Pedras, M.J.; Orsini, M.; Pirmez, C.; de Oliveira, M.C.; Castro, M.; Rabello, A. Leishmania (Viannia) subgenus kDNA amplification for the diagnosis of mucosal leishmaniasis. Diagn. Microbiol. Infect. Dis. 2005, 51, 185–190. [Google Scholar] [CrossRef]
- Uliana, S.R.; Affonso, M.H.; Camargo, E.P.; Floeter-Winter, L.M. Leishmania: Genus identification based on a specific sequence of the 18S ribosomal RNA sequence. Exp. Parasitol. 1991, 72, 157–163. [Google Scholar] [CrossRef]
- Uliana, S.R.; Nelson, K.; Beverley, S.M.; Camargo, E.P.; Floeter-Winter, L.M. Discrimination amongst Leishmania by polymerase chain reaction and hybridization with small subunit ribosomal DNA derived oligonucleotides. J. Eukaryot. Microbiol. 1994, 41, 324–330. [Google Scholar] [CrossRef]
- Castilho, T.M.; Shaw, J.J.; Floeter-Winter, L.M. New PCR assay using glucose-6-phosphate dehydrogenase for identification of Leishmania species. J. Clin. Microbiol. 2003, 41, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Garcia, L.; Kindt, A.; Bermudez, H.; Llanos-Cuentas, A.; De Doncker, S.; Arevalo, J.; Wilber Quispe Tintaya, K.; Dujardin, J.C. Culture-independent species typing of neotropical Leishmania for clinical validation of a PCR-based assay targeting heat shock protein 70 genes. J. Clin. Microbiol. 2004, 42, 2294–2297. [Google Scholar] [CrossRef] [PubMed]
- Montalvo, A.M.; Fraga, J.; Monzote, L.; Montano, I.; De Doncker, S.; Dujardin, J.C.; Van der Auwera, G. Heat-shock protein 70 PCR-RFLP: A universal simple tool for Leishmania species discrimination in the New and Old World. Parasitology 2010, 137, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Tellevik, M.G.; Muller, K.E.; Løkken, K.R.; Nerland, A.H. Detection of a broad range of Leishmania species and determination of parasite load of infected mouse by real-time PCR targeting the arginine permease gene AAP3. Acta Trop. 2014, 137, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Quispe Tintaya, K.W.; Ying, X.; Dedet, J.P.; Rijal, S.; De Bolle, X.; Dujardin, J.C. Antigen genes for molecular epidemiology of leishmaniasis: Polymorphism of cysteine proteinase B and surface metalloprotease glycoprotein 63 in the Leishmania donovani complex. J. Infect. Dis. 2004, 189, 1035–1043. [Google Scholar] [CrossRef][Green Version]
- Cupolillo, E.; Grimaldi Júnior, G.; Momen, H.; Beverley, S.M. Intergenic region typing (IRT): A rapid molecular approach to the characterization and evolution of Leishmania. Mol. Biochem. Parasitol. 1995, 73, 145–155. [Google Scholar] [CrossRef]
- Moreira, M.A.; Luvizotto, M.C.; Garcia, J.F.; Corbett, C.E.; Laurenti, M.D. Comparison of parasitological, immunological and molecular methods for the diagnosis of leishmaniasis in dogs with different clinical signs. Vet. Parasitol. 2007, 145, 245–252. [Google Scholar] [CrossRef]
- De Paiva-Cavalcanti, M.; de Morais, R.C.; Pessoa, E.S.R.; Trajano-Silva, L.A.; Gonçalves-de-Albuquerque Sda, C.; Tavares Dde, H.; Brelaz-de-Castro, M.C.; Silva Rde, F.; Pereira, V.R. Leishmaniases diagnosis: An update on the use of immunological and molecular tools. Cell Biosci. 2015, 5, 31. [Google Scholar] [CrossRef]
- Rodríguez, N.; Guzman, B.; Rodas, A.; Takiff, H.; Bloom, B.R.; Convit, J. Diagnosis of cutaneous leishmaniasis and species discrimination of parasites by PCR and hybridization. J. Clin. Microbiol. 1994, 32, 2246–2252. [Google Scholar] [CrossRef]
- Kumar, A.; Boggula, V.R.; Misra, P.; Sundar, S.; Shasany, A.K.; Dube, A. Amplified fragment length polymorphism (AFLP) analysis is useful for distinguishing Leishmania species of visceral and cutaneous forms. Acta Trop. 2010, 113, 202–206. [Google Scholar] [CrossRef]
- Mueller, U.G.; Wolfenbarger, L.L. AFLP genotyping and fingerprinting. Trends Ecol. Evol. 1999, 14, 389–394. [Google Scholar] [CrossRef]
- Schönian, G.; Schweynoch, C.; Zlateva, K.; Oskam, L.; Kroon, N.; Gräser, Y.; Presber, W. Identification and determination of the relationships of species and strains within the genus Leishmania using single primers in the polymerase chain reaction. Mol. Biochem. Parasitol. 1996, 77, 19–29. [Google Scholar] [CrossRef]
- Macedo, A.M.; Melo, M.N.; Gomes, R.F.; Pena, S.D. DNA fingerprints: A tool for identification and determination of the relationships between species and strains of Leishmania. Mol. Biochem. Parasitol. 1992, 53, 63–70. [Google Scholar] [CrossRef]
- Wang, J.Y.; Ha, Y.; Gao, C.H.; Wang, Y.; Yang, Y.T.; Chen, H.T. The prevalence of canine Leishmania infantum infection in western China detected by PCR and serological tests. Parasit. Vectors 2011, 4, 69. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, P.R.; Sethy, K.; Mohapatra, S.; Panda, D. Loop mediated isothermal amplification: An innovative gene amplification technique for animal diseases. Vet. World 2016, 9, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Avishek, K.; Sharma, V.; Negi, N.S.; Ramesh, V.; Salotra, P. Application of loop-mediated isothermal amplification assay for the sensitive and rapid diagnosis of visceral leishmaniasis and post-kala-azar dermal leishmaniasis. Diagn. Microbiol. Infect. Dis. 2013, 75, 390–395. [Google Scholar] [CrossRef]
- Savani, E.S.; de Oliveira Camargo, M.C.; de Carvalho, M.R.; Zampieri, R.A.; dos Santos, M.G.; D’Auria, S.R.; Shaw, J.J.; Floeter-Winter, L.M. The first record in the Americas of an autochthonous case of Leishmania (Leishmania) infantum chagasi in a domestic cat (Felix catus) from Cotia County, São Paulo State, Brazil. Vet. Parasitol. 2004, 120, 229–233. [Google Scholar] [CrossRef]
- Savani, E.S.; Nunes, V.L.; Galati, E.A.; Castilho, T.M.; Araujo, F.S.; Ilha, I.M.; Camargo, M.C.; D’Auria, S.R.; Floeter-Winter, L.M. Occurrence of co-infection by Leishmania (Leishmania) chagasi and Trypanosoma (Trypanozoon) evansi in a dog in the state of Mato Grosso do Sul, Brazil. Memorias do Instituto Oswaldo Cruz 2005, 100, 739–741. [Google Scholar] [CrossRef]
- Maruyama, S.R.; de Santana, A.K.M.; Takamiya, N.T.; Takahashi, T.Y.; Rogerio, L.A.; Oliveira, C.A.B.; Milanezi, C.M.; Trombela, V.A.; Cruz, A.K.; Jesus, A.R.; et al. Non-Leishmania Parasite in Fatal Visceral Leishmaniasis-Like Disease, Brazil. Emerg. Infect. Dis. 2019, 25, 2088–2092. [Google Scholar] [CrossRef] [PubMed]
- Boisseau-Garsaud, A.M.; Cales-Quist, D.; Desbois, N.; Jouannelle, J.; Jouannelle, A.; Pratlong, F.; Dedet, J.P. A new case of cutaneous infection by a presumed monoxenous trypanosomatid in the island of Martinique (French West Indies). Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 51–52. [Google Scholar] [CrossRef]
- Mortarino, M.; Franceschi, A.; Mancianti, F.; Bazzocchi, C.; Genchi, C.; Bandi, C. Quantitative PCR in the diagnosis of Leishmania. Parassitologia 2004, 46, 163–167. [Google Scholar] [PubMed]
- Castilho, T.M.; Camargo, L.M.; McMahon-Pratt, D.; Shaw, J.J.; Floeter-Winter, L.M. A real-time polymerase chain reaction assay for the identification and quantification of American Leishmania species on the basis of glucose-6-phosphate dehydrogenase. Am. J. Trop. Med. Hyg. 2008, 78, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Reed, G.H.; Kent, J.O.; Wittwer, C.T. High-resolution DNA melting analysis for simple and efficient molecular diagnostics. Pharmacogenomics 2007, 8, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, L.; Milon, G.; Prina, E. Rapid differentiation of Old World Leishmania species by LightCycler polymerase chain reaction and melting curve analysis. J. Microbiol. Methods 2002, 51, 295–299. [Google Scholar] [CrossRef]
- Pita-Pereira, D.; Lins, R.; Oliveira, M.P.; Lima, R.B.; Pereira, B.A.; Moreira, O.C.; Brazil, R.P.; Britto, C. SYBR Green-based real-time PCR targeting kinetoplast DNA can be used to discriminate between the main etiologic agents of Brazilian cutaneous and visceral leishmaniases. Parasit. Vectors 2012, 5, 15. [Google Scholar] [CrossRef]
- Ceccarelli, M.; Galluzzi, L.; Migliazzo, A.; Magnani, M. Detection and characterization of Leishmania (Leishmania) and Leishmania (Viannia) by SYBR green-based real-time PCR and high resolution melt analysis targeting kinetoplast minicircle DNA. PLoS ONE 2014, 9, e88845. [Google Scholar] [CrossRef]
- Hernández, C.; Alvarez, C.; González, C.; Ayala, M.S.; León, C.M.; Ramírez, J.D. Identification of six New World Leishmania species through the implementation of a High-Resolution Melting (HRM) genotyping assay. Parasit. Vectors 2014, 7, 501. [Google Scholar] [CrossRef]
- Zampieri, R.A.; Laranjeira-Silva, M.F.; Muxel, S.M.; Stocco de Lima, A.C.; Shaw, J.J.; Floeter-Winter, L.M. High Resolution Melting Analysis Targeting hsp70 as a Fast and Efficient Method for the Discrimination of Leishmania Species. PLoS Negl. Trop. Dis. 2016, 10, e0004485. [Google Scholar] [CrossRef]
- Müller, K.E.; Zampieri, R.A.; Aoki, J.I.; Muxel, S.M.; Nerland, A.H.; Floeter-Winter, L.M. Amino acid permease 3 (aap3) coding sequence as a target for Leishmania identification and diagnosis of leishmaniases using high resolution melting analysis. Parasit. Vectors 2018, 11, 421. [Google Scholar] [CrossRef]
- Grimaldi, G.; McMahon-Pratt, D. Monoclonal antibodies for the identification of New World Leishmania species. Memorias do Instituto Oswaldo Cruz 1996, 91, 37–42. [Google Scholar] [CrossRef][Green Version]
- Sogawa, K.; Watanabe, M.; Sato, K.; Segawa, S.; Ishii, C.; Miyabe, A.; Murata, S.; Saito, T.; Nomura, F. Use of the MALDI BioTyper system with MALDI-TOF mass spectrometry for rapid identification of microorganisms. Anal. Bioanal. Chem. 2011, 400, 1905–1911. [Google Scholar] [CrossRef] [PubMed]
- Cassagne, C.; Pratlong, F.; Jeddi, F.; Benikhlef, R.; Aoun, K.; Normand, A.C.; Faraut, F.; Bastien, P.; Piarroux, R. Identification of Leishmania at the species level with matrix-assisted laser desorption ionization time-of-flight mass spectrometry. Clin. Microbiol. Infect. 2014, 20, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Dey, A.; Sivakumar, R. Applications of molecular methods for Leishmania control. Expert Rev. Mol. Diagn. 2005, 5, 251–265. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.J. A partnership that worked: The Wellcome Trust and the Instituto Evandro Chagas and beyond. Rev. Pan-Amazônica de Saúde 2016, 7, 23–42. [Google Scholar] [CrossRef]
- Walton, B.C. Evaluation of chemotherapy of American leishmaniasis by the indirect fluorescent antibody test. Am. J. Trop. Med. Hyg. 1980, 29, 747–752. [Google Scholar] [CrossRef]
- Singh, O.P.; Sundar, S. Developments in Diagnosis of Visceral Leishmaniasis in the Elimination Era. J. Parasitol. Res. 2015, 2015, 239469. [Google Scholar] [CrossRef]
- Mohapatra, T.M.; Singh, D.P.; Sen, M.R.; Bharti, K.; Sundar, S. Compararative evaluation of rK9, rK26 and rK39 antigens in the serodiagnosis of Indian visceral leishmaniasis. J. Infect. Dev. Ctries. 2010, 4, 114–117. [Google Scholar] [CrossRef]
- Greiner, M.; Gardner, I.A. Application of diagnostic tests in veterinary epidemiologic studies. Prev. Vet. Med. 2000, 45, 43–59. [Google Scholar] [CrossRef]
- Pan-American Health Organization (PAHO). Plan of Action to Strengthen the Surveillance and Control of Leishmaniasis in the Americas 2017–2022. Available online: https://iris.paho.org/bitstream/handle/10665.2/34147/PlanactionLeish20172022-eng.pdf?sequence=5&isAllowed=y (accessed on 1 May 2020).
- Oleaga, A.; Zanet, S.; Espí, A.; Pegoraro de Macedo, M.R.; Gortázar, C.; Ferroglio, E. Leishmania in wolves in northern Spain: A spreading zoonosis evidenced by wildlife sanitary surveillance. Vet. Parasitol. 2018, 255, 26–31. [Google Scholar] [CrossRef]
- Ostad, M.; Shirian, S.; Pishro, F.; Abbasi, T.; Ai, A.; Azimi, F. Control of Cutaneous Leishmaniasis Using Geographic Information Systems from 2010 to 2014 in Khuzestan Province, Iran. PLoS ONE 2016, 11, e0159546. [Google Scholar] [CrossRef]
- Richini-Pereira, V.B.; Marson, P.M.; Hayasaka, E.Y.; Victoria, C.; da Silva, R.C.; Langoni, H. Molecular detection of Leishmania spp. in road-killed wild mammals in the Central Western area of the State of São Paulo, Brazil. J. Venom. Anim. Toxins. Incl. Trop. Dis. 2014, 20, 27. [Google Scholar] [CrossRef]
- Salahi-Moghaddam, A.; Mohebali, M.; Moshfae, A.; Habibi, M.; Zarei, Z. Ecological study and risk mapping of visceral leishmaniasis in an endemic area of Iran based on a geographical information systems approach. Geospat. Health 2010, 5, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Melo, S.N.; Teixeira-Neto, R.G.; Werneck, G.L.; Struchiner, C.J.; Ribeiro, R.A.N.; Sousa, L.R.; de Melo, M.O.G.; Carvalho Júnior, C.G.; Penaforte, K.M.; Manhani, M.N.; et al. Prevalence of visceral leishmaniasis in A population of free-roaming dogs as determined by multiple sampling efforts: A longitudinal study analyzing the effectiveness of euthanasia. Prev. Vet. Med. 2018, 161, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Bsrat, A.; Berhe, M.; Gadissa, E.; Taddele, H.; Tekle, Y.; Hagos, Y.; Abera, A.; G/micael, M.; Alemayhu, T.; Gugsa, G.; et al. Serological investigation of visceral Leishmania infection in human and its associated risk factors in Welkait District, Western Tigray, Ethiopia. Parasite Epidemiol. Control. 2018, 3, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Mody, R.M.; Lakhal-Naouar, I.; Sherwood, J.E.; Koles, N.L.; Shaw, D.; Bigley, D.P.; Co, E.A.; Copeland, N.K.; Jagodzinski, L.L.; Mukbel, R.M.; et al. Asymptomatic Visceral Leishmania infantum Infection in US Soldiers Deployed to Iraq. Clin. Infect. Dis. 2019, 68, 2036–2044. [Google Scholar] [CrossRef]
- Latif, A.A.; Nkabinde, B.; Peba, B.; Matthee, O.; Pienaar, R.; Josemans, A.; Marumo, D.; Labuschagne, K.; Abdelatief, N.A.; Krüger, A.; et al. Risk of establishment of canine leishmaniasis infection through the import of dogs into South Africa. Onderstepoort. J. Vet. Res. 2019, 86, e1–e11. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Clinical Manifestation | Reservoir Host | Country/Region | |
|---|---|---|---|---|
| Eurasia (Old World) | L. (L.) donovani | AVL, PKDL, CL | Human | VL: West and Central Asia, China, The Indian subcontinent, The Mediterranean Basin, East Africa; CL: The Mediterranean Basin; ML: North Africa; PKDL: The Indian subcontinent, East and North Africa |
| L. (L.) infantum | AVL, ZVL, CL | Human, Dog, Fox, Jackal, Badger, Rodent, Cat, Opossum | VL: Central and West Asia, China, The Mediterranean Basin, Africa; CL: The Mediterranean Basin, West Asia, China, West Africa | |
| L. (L.) major | ZCL | Rodent | West and Central Asia, The Indian subcontinent, The Mediterranean Basin, Africa | |
| L. (L.) tropica | ACL, ZCLAVL | Human, HyraxHuman | Central, South and West Asia, The Mediterranean Basin, East Africa West Asia | |
| L. (L.) killicki | CL | Unknown | The Mediterranean Basin | |
| L. (L.) aethiopica | ZCL, DCL, ML | Hyrax, Rodent | CL: East Africa (Ethiopia and Kenya); ML: Ethiopia | |
| L. (M.) orientalis | CL, DL, VL | Unknown | Thailand | |
| Americas (New World) | L. (L.) infantum chagasi | ZVL, CL | Dog, Cat, Fox, Opossum | South and Central America, Mexico |
| L. (L.) mexicana | ZCL, MCL, DCL | Rodent, Opossum | Americas | |
| L. (L.) pifanol | DCL | Unknown | Venezuela | |
| L. (L.) venezuelensis | CL | Unknown | Venezuela | |
| L. (L.) garnhami | ZCL | Unknown | Central America, Venezuela | |
| L. (L.) amazonensis | ZCL, DCL, CL | Rodent | South America | |
| L. (V.) braziliensis | ZCL, MCL, DL | Dog, Horse, Donkey, Mule, Rodent, Opossum | South and Central America, Mexico | |
| L. (L.) waltoni | DCL | Unknown | Dominican Republic | |
| L. (V.) guyanesis | ZCL, MCL | Sloth, Anteater, Opossum | South America | |
| L. (V.) panamensis | ZCL, MCL | Dog, Sloth, Opossum, Tamandua | South and Central America | |
| L. (V.) shawi | ZCL | Sloth, Primate | Brazil | |
| L. (V.) naiffi | ZCL | Armadillo | Brazil, French Guiana | |
| L. (V.) lainsoni | ZCL | Rodent | South America | |
| L. (V.) lindenbergi | ZCL | Unknown | Brazil | |
| L. (V.) peruviana | ZCL, MCL | Dog, Opossum, Rodent | Peru | |
| L. (M.) martiniquensis | CL | Unknown | French Guiana | |
| Endotrypanum colombiensis | ZCL | Sloth | Colombia, Venezuela, Panama |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, A.; Zampieri, R.A.; Shaw, J.J.; Floeter-Winter, L.M.; Laranjeira-Silva, M.F. One Health Approach to Leishmaniases: Understanding the Disease Dynamics through Diagnostic Tools. Pathogens 2020, 9, 809. https://doi.org/10.3390/pathogens9100809
Hong A, Zampieri RA, Shaw JJ, Floeter-Winter LM, Laranjeira-Silva MF. One Health Approach to Leishmaniases: Understanding the Disease Dynamics through Diagnostic Tools. Pathogens. 2020; 9(10):809. https://doi.org/10.3390/pathogens9100809
Chicago/Turabian StyleHong, Ahyun, Ricardo Andrade Zampieri, Jeffrey Jon Shaw, Lucile Maria Floeter-Winter, and Maria Fernanda Laranjeira-Silva. 2020. "One Health Approach to Leishmaniases: Understanding the Disease Dynamics through Diagnostic Tools" Pathogens 9, no. 10: 809. https://doi.org/10.3390/pathogens9100809
APA StyleHong, A., Zampieri, R. A., Shaw, J. J., Floeter-Winter, L. M., & Laranjeira-Silva, M. F. (2020). One Health Approach to Leishmaniases: Understanding the Disease Dynamics through Diagnostic Tools. Pathogens, 9(10), 809. https://doi.org/10.3390/pathogens9100809