Immune–Pathological Correlates of Disease Severity in New-World Kala-Azar: The Role of Parasite Load and Cytokine Profiles

 ,
, 
Abstract

1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Medical Data
2.3. DNA Isolation, Purity, and Standardization
2.4. Quantitative PCR
2.5. Plasma Cytokines
2.6. Statistical Analysis
3. Results
3.1. Study Population
3.2. Clinical Findings
3.3. Quantity of Parasite Load and Cytokines
3.4. Plasma and Bone Marrow L. infantum Loads
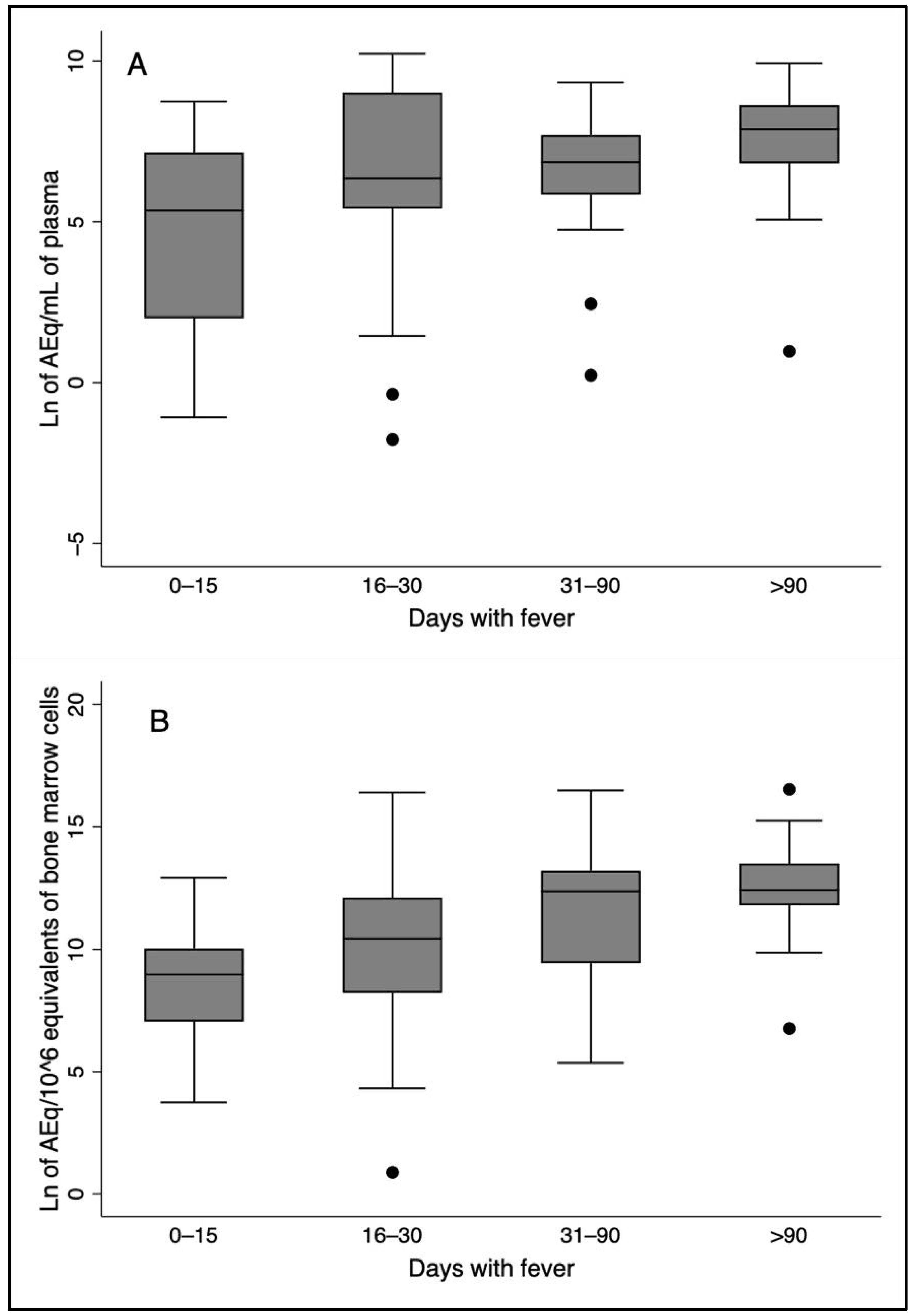
3.5. Time of Disease, Plasma and Bone Marrow L. infantum Loads, Plasma Cytokines, and Severity
3.6. Plasma and Bone Marrow L. infantum Loads, Age, Sex, HIV Infection, and Kala-Azar Severity
3.7. Plasma Cytokines, Age, Sex, HIV Infection, and Markers of Kala-Azar Severity
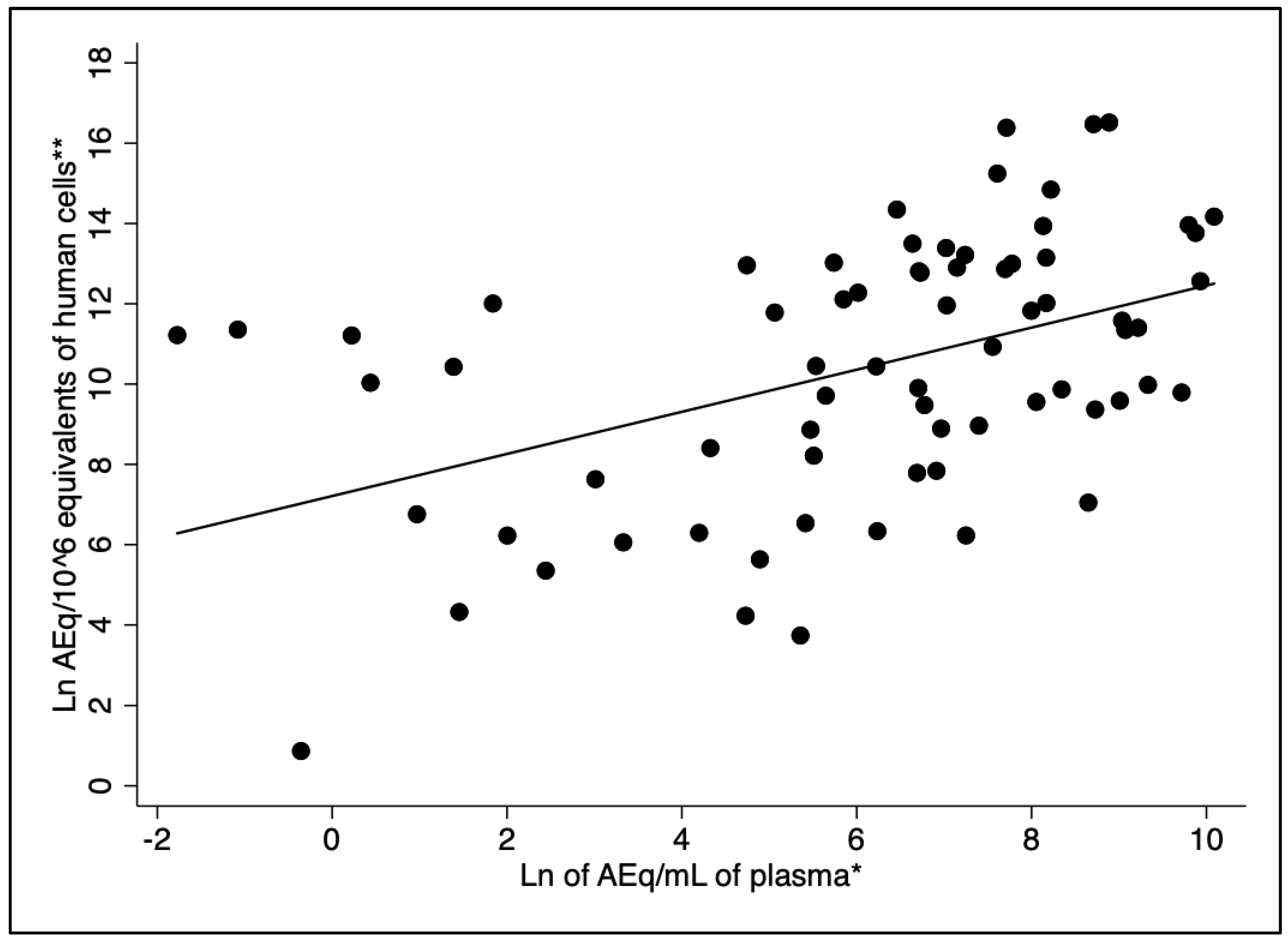
3.8. Regression Analysis Between L. infantum Load and Plasma Cytokines
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burza, S.; Croft, S.L.; Boelaert, M. Leishmaniasis. Lancet 2018, 392, 951–970. [Google Scholar] [CrossRef] [PubMed]
- Andrade, T.M.; Carvalho, E.M.; Rocha, H. Bacterial Infections in Patients with Visceral Leishmaniasis. J. Infect. Dis. 1990, 162, 1354–1359. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.H.N.; Werneck, G.L.; Costa, D.L.; Holanda, T.A.; Aguiar, G.B.; Carvalho, A.S.; Cavalcanti, J.C.; Santos, L.S. Is severe visceral leishmaniasis a systemic inflammatory response syndrome? A case control study. Rev. Soc. Bras. Med. Trop. 2010, 43, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, M.J.A.d.Q.; Cavalcanti, N.V.; Alves, J.G.B.; Fernandes Filho, M.J.C.; Correia, J.B. Risk Factors for Death in Children with Visceral Leishmaniasis. PLoS Negl. Trop. Dis. 2010, 4, e877. [Google Scholar] [CrossRef]
- Costa, D.L.; Rocha, R.L.; Chaves, E.d.B.F.; Batista, V.G.d.V.; Costa, H.L.; Costa, C.H.N. Predicting death from kala-azar: Construction, development, and validation of a score set and accompanying software. Rev. Soc. Bras. Med. Trop. 2016, 49, 728–740. [Google Scholar] [CrossRef]
- Akuffo, H.; Costa, C.; van Griensven, J.; Burza, S.; Moreno, J.; Herrero, M. New insights into leishmaniasis in the immunosuppressed. PLoS Negl. Trop. Dis. 2018, 12, e0006375. [Google Scholar] [CrossRef]
- Kurizky, P.S.; Marianelli, F.F.; Cesetti, M.V.; Damiani, G.; Sampaio, R.N.R.; Gonçalves, L.M.T.; Sousa, C.A.F.d.; Martins, S.S.; Vernal, S.; Mota, L.M.H.d.; et al. A comprehensive systematic review of leishmaniasis in patients undergoing drug-induced immunosuppression for the treatment of dermatological, rheumatological and gastroenterological diseases. Rev. Inst. Med. Trop. Sao Paulo 2020, 62, e28. [Google Scholar] [CrossRef]
- Rahim, S.; Karim, M.M. The Elimination Status of Visceral Leishmaniasis in Southeast Asia Region. Acta Parasitol. 2024, 69, 1704–1716. [Google Scholar] [CrossRef]
- da Rocha, I.C.M.; dos Santos, L.H.M.; Coura-Vital, W.; da Cunha, G.M.R.; Magalhães, F.d.C.; da Silva, T.A.M.; Morais, M.H.F.; Oliveira, E.; Reis, I.A.; Carneiro, M. Effectiveness of the Brazilian Visceral Leishmaniasis Surveillance and Control Programme in reducing the prevalence and incidence of Leishmania infantum infection. Parasit. Vectors 2018, 11, 586. [Google Scholar] [CrossRef]
- McCall, L.I.; Zhang, W.W.; Matlashewski, G. Determinants for the Development of Visceral Leishmaniasis Disease. PLoS Pathog. 2013, 9, e1003053. [Google Scholar] [CrossRef]
- Volpedo, G.; Pacheco-Fernandez, T.; Bhattacharya, P.; Oljuskin, T.; Dey, R.; Gannavaram, S.; Satoskar, A.R.; Nakhasi, H.L. Determinants of Innate Immunity in Visceral Leishmaniasis and Their Implication in Vaccine Development. Front. Immunol. 2021, 12, 748325. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, C.; Islam, N.A.K.; Barinberg, D.; Soulat, D.; Schleicher, U.; Rai, B. The immunomicrotope of Leishmania control and persistence. Trends Parasitol. 2024, 40, 788–804. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.C.; Zacarias, D.A.; Silva, V.C.; Rolão, N.; Costa, D.L.; Costa, C.H. Comparison of optical microscopy and quantitative polymerase chain reaction for estimating parasitaemia in patients with kala-azar and modelling infectiousness to the vector Lutzomyia longipalpis. Mem. Inst. Oswaldo Cruz 2016, 111, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, G.R.; Santos-Oliveira, J.R.; Silva-Freitas, M.L.; Honda, M.; Costa, D.L.; Da-Cruz, A.M.; Costa, C.H.N. Biomarkers of disease severity in patients with visceral leishmaniasis co-infected with HIV. Cytokine 2022, 149, 155747. [Google Scholar] [CrossRef]
- Al-Khalaifah, H.S. Major Molecular Factors Related to Leishmania Pathogenicity. Front. Immunol. 2022, 13, 847797. [Google Scholar] [CrossRef]
- Samant, M.; Sahu, U.; Pandey, S.C.; Khare, P. Role of Cytokines in Experimental and Human Visceral Leishmaniasis. Front. Cell. Infect. Microbiol. 2021, 11, 624009. [Google Scholar] [CrossRef]
- Costa, D.L.; Rocha, R.L.; Carvalho, R.M.A.; Lima-Neto, A.S.; Harhay, M.O.; Costa, C.H.N.; Barral-Neto, M.; Barral, A.P. Serum cytokines associated with severity and complications of kala-azar. Pathog. Glob. Health 2013, 107, 78–87. [Google Scholar] [CrossRef]
- Costa, C.H.N.; Chang, K.P.; Costa, D.L.; Cunha, F.V.M. From Infection to Death: An Overview of the Pathogenesis of Visceral Leishmaniasis. Pathogens 2023, 12, 969. [Google Scholar] [CrossRef]
- dos Santos, P.L.; de Oliveira, F.A.; Santos, M.L.B.; Cunha, L.C.S.; Lino, M.T.B.; de Oliveira, M.F.S.; Bomfim, M.O.M.; Silva, A.M.; de Moura, T.R.; de Jesus, A.R.; et al. The Severity of Visceral Leishmaniasis Correlates with Elevated Levels of Serum IL-6, IL-27 and sCD14. PLoS Negl. Trop. Dis. 2016, 10, e0004375. [Google Scholar] [CrossRef]
- Silva, J.M.; Zacarias, D.A.; de Figueirêdo, L.C.; Soares, M.R.A.; Ishikawa, E.A.Y.; Costa, D.L.; Costa, C.H.N. Bone Marrow Parasite Burden among Patients with New World Kala-Azar is Associated with Disease Severity. Am. Soc. Trop. Med. Hyg. 2014, 90, 621–626. [Google Scholar] [CrossRef]
- Zacarias, D.A.; Rolão, N.; de Pinho, F.A.; Sene, I.; Silva, J.C.; Pereira, T.C.; Costa, D.L.; Costa, C.H.N. Causes and consequences of higher Leishmania infantum burden in patients with kala-azar: A study of 625 patients. Trop. Med. Int. Health 2017, 22, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Grace, C.A.; Sousa Carvalho, K.S.; Sousa Lima, M.I.; Costa Silva, V.; Reis-Cunha, J.L.; Brune, M.J.; Forrester, S.; Pedrozo e Silva de Azevedo, C.d.M.; Costa, D.L.; Speed, D.; et al. Parasite Genotype Is a Major Predictor of Mortality from Visceral Leishmaniasis. MBio 2022, 13, e0206822. [Google Scholar] [CrossRef] [PubMed]
- Cota, G.; Erber, A.C.; Schernhammer, E.; Simões, T.C. Inequalities of visceral leishmaniasis case-fatality in Brazil: A multilevel modeling considering space, time, individual and contextual factors. PLoS Negl. Trop. Dis. 2021, 15, e0009567. [Google Scholar] [CrossRef] [PubMed]
- Kildey, K.; Rooks, K.; Weier, S.; Flower, R.L.; Dean, M.M. Effect of age, gender and mannose-binding lectin (MBL) status on the inflammatory profile in peripheral blood plasma of Australian blood donors. Hum. Immunol. 2014, 75, 973–979. [Google Scholar] [CrossRef]
- Porcino, G.N.; Carvalho, K.S.S.; Braz, D.C.; Costa Silva, V.; Costa, C.H.N.; de Miranda Santos, I.K.F. Evaluation of methods for detection of asymptomatic individuals infected with Leishmania infantum in the state of Piauí, Brazil. PLoS Negl. Trop. Dis. 2019, 13, e0007493. [Google Scholar] [CrossRef]
- Das, V.N.R.; Bimal, S.; Siddiqui, N.A.; Kumar, A.; Pandey, K.; Sinha, S.K.; Topno, R.K.; Mahentesh, V.; Singh, A.K.; Lal, C.S.; et al. Conversion of asymptomatic infection to symptomatic visceral leishmaniasis: A study of possible immunological markers. PLoS Negl. Trop. Dis. 2020, 14, e0008272. [Google Scholar] [CrossRef]
- Chakravarty, J.; Hasker, E.; Kansal, S.; Singh, O.P.; Malaviya, P.; Singh, A.K.; Chourasia, A.; Singh, T.; Sudarshan, M.; Singh, A.P.; et al. Determinants for progression from asymptomatic infection to symptomatic visceral leishmaniasis: A cohort study. PLoS Negl. Trop. Dis. 2019, 13, e0007216. [Google Scholar] [CrossRef]
- Virginia Batista Vieira, A.; Farias, P.C.S.; Silva Nunes Bezerra, G.; Xavier, A.T.; Sebastião Da Costa Lima Júnior, M.; Silva, E.D.D.; Barbosa Júnior, W.L.; Medeiros, Z.M. Evaluation of molecular techniques to visceral leishmaniasis detection in asymptomatic patients: A systematic review. Expert Rev. Mol. Diagn. 2021, 21, 493–504. [Google Scholar] [CrossRef]
- Harhay, M.O.; Olliaro, P.L.; Vaillant, M.; Chappuis, F.; Lima, M.A.; Ritmeijer, K.; Costa, C.H.; Costa, D.L.; Rijal, S.; Sundar, S.; et al. Who Is a Typical Patient with Visceral Leishmaniasis? Characterizing the Demographic and Nutritional Profile of Patients in Brazil, East Africa, and South Asia. Am. Soc. Trop. Med. Hyg. 2011, 84, 543–550. [Google Scholar] [CrossRef]
- Leite de Sousa-Gomes, M.; Romero, G.A.S.; Werneck, G.L. Visceral leishmaniasis and HIV/AIDS in Brazil: Are we aware enough? PLoS Negl. Trop. Dis. 2017, 11, e0005772. [Google Scholar] [CrossRef]
- Machado, C.A.L.; Sevá, A.d.P.; Silva, A.A.F.A.e.; Horta, M.C. Epidemiological profile and lethality of visceral leishmaniasis/human immunodeficiency virus co-infection in an endemic area in Northeast Brazil. Rev. Soc. Bras. Med. Trop. 2021, 54, e0795. [Google Scholar] [CrossRef] [PubMed]
- Hailu, A.; van der Poll, T.; Berhe, N.; Kager, P.A. Elevated plasma levels of interferon (IFN)-gamma, IFN-gamma inducing cytokines, and IFN-gamma inducible CXC chemokines in visceral leishmaniasis. Am. J. Trop. Med. Hyg. 2004, 71, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Peruhype-Magalhães, V.; Martins-Filho, O.A.; Prata, A.; Silva, L.D.A.; Rabello, A.; Teixeira-Carvalho, A.; Figueiredo, R.M.; Guimarães-Carvalho, S.F.; Ferrari, T.C.A.; Van Weyenbergh, J.; et al. Mixed inflammatory/regulatory cytokine profile marked by simultaneous raise of interferon-γ and interleukin-10 and low frequency of tumour necrosis factor-α+ monocytes are hallmarks of active human visceral Leishmaniasis due to Leishmania chagasi infection. Clin. Exp. Immunol. 2006, 146, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Ansari, N.A.; Saluja, S.; Salotra, P. Elevated levels of interferon-γ, interleukin-10, and interleukin-6 during active disease in Indian kala azar. Clin. Immunol. 2006, 119, 339–345. [Google Scholar] [CrossRef]
- van Dijk, N.J.; Carter, J.; Kiptanui, D.; Mens, P.F.; Schallig, H.D.F.H. A case–control study on risk factors for visceral leishmaniasis in West Pokot County, Kenya. Trop. Med. Int. Health 2024, 29, 904–912. [Google Scholar] [CrossRef]
- Decker, M.L.; Grobusch, M.P.; Ritz, N. Influence of Age and Other Factors on Cytokine Expression Profiles in Healthy Children—A Systematic Review. Front. Pediatr. 2017, 5, 255. [Google Scholar] [CrossRef]
- Ringleb, M.; Javelle, F.; Haunhorst, S.; Bloch, W.; Fennen, L.; Baumgart, S.; Drube, S.; Reuken, P.A.; Pletz, M.W.; Wagner, H.; et al. Beyond muscles: Investigating immunoregulatory myokines in acute resistance exercise—A systematic review and meta-analysis. FASEB J. 2024, 38, e23596. [Google Scholar] [CrossRef]
- Tasca, K.I.; Correa, C.R.; Caleffi, J.T.; Mendes, M.B.; Gatto, M.; Manfio, V.M.; de Camargo, C.C.; Tavares, F.C.; Biasin, M.; de Souza, L.d.R. Asymptomatic HIV People Present Different Profiles of sCD14, sRAGE, DNA Damage, and Vitamins, according to the Use of cART and CD4+ T Cell Restoration. J. Immunol. Res. 2018, 2018, 7531718. [Google Scholar] [CrossRef]
- Guedes, D.L.; Silva, E.D.d.; Castro, M.C.A.B.; Júnior, W.L.B.; Ibarra-Meneses, A.V.; Tsoumanis, A.; Adriaensen, W.; van Griensven, J.; Pereira, V.R.A.; Medeiros, Z.M.d. Comparison of serum cytokine levels in symptomatic and asymptomatic HIV-Leishmania coinfected individuals from a Brazilian visceral leishmaniasis endemic area. PLoS Negl. Trop. Dis. 2022, 16, e0010542. [Google Scholar] [CrossRef]
- Hunter, C.A.; Jones, S.A. IL-6 as a keystone cytokine in health and disease. Nat. Immunol. 2015, 16, 448–457. [Google Scholar] [CrossRef]
- Mary, C.; Faraut, F.; Lascombe, L.; Dumon, H. Quantification of Leishmania infantum DNA by a Real-Time PCR Assay with High Sensitivity. J. Clin. Microbiol. 2004, 42, 5249–5255. [Google Scholar] [CrossRef] [PubMed]
- de Araújo Albuquerque, L.P.; da Silva, A.M.; de Araújo Batista, F.M.; de Souza Sene, I.; Costa, D.L.; Costa, C.H.N. Influence of sex hormones on the immune response to leishmaniasis. Parasite Immunol. 2021, 43, e12874. [Google Scholar] [CrossRef] [PubMed]
- Murray, H.W.; Rubin, B.Y.; Rothermel, C.D. Killing of intracellular Leishmania donovani by lymphokine-stimulated human mononuclear phagocytes. Evidence that interferon-gamma is the activating lymphokine. J. Clin. Investig. 1983, 72, 1506–1510. [Google Scholar] [CrossRef] [PubMed]
- Reiner, N.E.; Ng, W.; Wilson, C.B.; McMaster, W.R.; Burchett, S.K. Modulation of in vitro monocyte cytokine responses to Leishmania donovani. Interferon-gamma prevents parasite-induced inhibition of interleukin 1 production and primes monocytes to respond to Leishmania by producing both tumor necrosis factor-alpha and in. J. Clin. Investig. 1990, 85, 1914–1924. [Google Scholar] [CrossRef]
- Heinzel, F.P.; Schoenhaut, D.S.; Rerko, R.M.; Rosser, L.E.; Gately, M.K. Recombinant interleukin 12 cures mice infected with Leishmania major. J. Exp. Med. 1993, 177, 1505–1509. [Google Scholar] [CrossRef]
- Murray, H.W.; Hariprashad, J. Interleukin 12 is effective treatment for an established systemic intracellular infection: Experimental visceral leishmaniasis. J. Exp. Med. 1995, 181, 387–391. [Google Scholar] [CrossRef]
- Ghalib, H.W.; Piuvezam, M.R.; Skeiky, Y.A.; Siddig, M.; Hashim, F.A.; El-Hassan, A.M.; Russo, D.M.; Reed, S.G. Interleukin 10 production correlates with pathology in human Leishmania donovani infections. J. Clin. Investig. 1993, 92, 324–329. [Google Scholar] [CrossRef]
- Bacellar, O.; Brodskyn, C.; Guerreiro, J.; Barral-Netto, M.; Costa, C.H.; Coffman, R.L.; Johnson, W.D.; Carvalho, E.M. Interleukin-12 Restores Interferon- Production and Cytotoxic Responses in Visceral Leishmaniasis. J. Infect. Dis. 1996, 173, 1515–1518. [Google Scholar] [CrossRef]
- Verma, S.; Kumar, R.; Katara, G.K.; Singh, L.C.; Negi, N.S.; Ramesh, V.; Salotra, P. Quantification of Parasite Load in Clinical Samples of Leishmaniasis Patients: IL-10 Level Correlates with Parasite Load in Visceral Leishmaniasis. PLoS ONE 2010, 5, e10107. [Google Scholar] [CrossRef]
- Bhattacharya, P.; Ghosh, S.; Ejazi, S.A.; Rahaman, M.; Pandey, K.; Ravi Das, V.N.; Das, P.; Goswami, R.P.; Saha, B.; Ali, N. Induction of IL-10 and TGFβ from CD4+CD25+FoxP3+ T Cells Correlates with Parasite Load in Indian Kala-azar Patients Infected with Leishmania donovani. PLoS Negl. Trop. Dis. 2016, 10, e0004422. [Google Scholar] [CrossRef]
- Teles, L.d.F.; Viana, A.G.; Cardoso, M.S.; Pinheiro, G.R.G.; Bento, G.A.; Lula, J.F.; Soares, T.d.C.M.; Fujiwara, R.T.; Carvalho, S.F.G.d. Evaluation of medullary cytokine expression and clinical and laboratory aspects in severe human visceral leishmaniasis. Parasite Immunol. 2021, 43, e12880. [Google Scholar] [CrossRef] [PubMed]
- Ansari, N.A.; Kumar, R.; Gautam, S.; Nylén, S.; Singh, O.P.; Sundar, S.; Sacks, D. IL-27 and IL-21 Are Associated with T Cell IL-10 Responses in Human Visceral Leishmaniasis. J. Immunol. 2011, 186, 3977–3985. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, E.M.; Badaró, R.; Reed, S.G.; Jones, T.C.; Johnson, W.D. Absence of gamma interferon and interleukin 2 production during active visceral leishmaniasis. J. Clin. Investig. 1985, 76, 2066–2069. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, E.M.; Bacellar, O.; Brownell, C.; Regis, T.; Coffman, R.L.; Reed, S.G. Restoration of IFN-gamma production and lymphocyte proliferation in visceral leishmaniasis. J. Immunol. 1994, 152, 5949–5956. [Google Scholar] [CrossRef]
- Karp, C.L.; El-Safi, S.H.; Wynn, T.A.; Satti, M.M.; Kordofani, A.M.; Hashim, F.A.; Hag-Ali, M.; Neva, F.A.; Nutman, T.B.; Sacks, D.L. In vivo cytokine profiles in patients with kala-azar. Marked elevation of both interleukin-10 and interferon-gamma. J. Clin. Investig. 1993, 91, 1644–1648. [Google Scholar] [CrossRef]
- Nylén, S.; Sacks, D. Interleukin-10 and the pathogenesis of human visceral leishmaniasis. Trends Immunol. 2007, 28, 378–384. [Google Scholar] [CrossRef]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef]
- Fabri, A.; Kandara, K.; Coudereau, R.; Gossez, M.; Abraham, P.; Monard, C.; Cour, M.; Rimmelé, T.; Argaud, L.; Monneret, G.; et al. Characterization of Circulating IL-10-Producing Cells in Septic Shock Patients: A Proof of Concept Study. Front. Immunol. 2021, 11, 615009. [Google Scholar] [CrossRef]
- D’Oliveira Júnior, A.; Costa, S.R.M.; Bispo Barbosa, A.; Orge Orge, M.d.L.G.; Carvalho, E.M. Asymptomatic Leishmania chagasi Infection in Relatives and Neighbors of Patients with Visceral Leishmaniasis. Mem. Inst. Oswaldo Cruz 1997, 92, 15–20. [Google Scholar] [CrossRef]
- Lima, S.; Braz, D.; Silva, V.; Farias, T.; Zacarias, D.A.; Silva, J.C.; Costa, C.H.N.; Costa, D.L. Biomarkers of the early response to treatment of visceral leishmaniasis: A prospective cohort study. Parasite Immunol. 2021, 43, e12797. [Google Scholar] [CrossRef]
- Saraiva, M.; Vieira, P.; O’Garra, A. Biology and therapeutic potential of interleukin-10. J. Exp. Med. 2020, 217, e20190418. [Google Scholar] [CrossRef] [PubMed]
- Munk, R.B.; Sugiyama, K.; Ghosh, P.; Sasaki, C.Y.; Rezanka, L.; Banerjee, K.; Takahashi, H.; Sen, R.; Longo, D.L. Antigen-Independent IFN-γ Production by Human Naïve CD4+ T Cells Activated by IL-12 Plus IL-18. PLoS ONE 2011, 6, e18553. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.G.; Cho, M.J.; Choi, J.M. Author Correction: Bystander CD4+ T cells: Crossroads between innate and adaptive immunity. Exp. Mol. Med. 2023, 55, 1275. [Google Scholar] [CrossRef] [PubMed]
- Reis-Cunha, J.L.; Grace, C.A.; Ahmed, S.; Harnqvist, S.E.; Lynch, C.M.; Boité, M.C.; Barcellos, G.; Lachaud, L.; Bastien, P.; Munt, H.; et al. The global dispersal of visceral leishmaniasis occurred within human history. bioRxiv 2024. [Google Scholar] [CrossRef]
- Zijlstra, E.E. The immunology of post-kala-azar dermal leishmaniasis (PKDL). Parasit. Vectors 2016, 9, 464. [Google Scholar] [CrossRef]
- Jarczak, D.; Nierhaus, A. Cytokine Storm—Definition, Causes, and Implications. Int. J. Mol. Sci. 2022, 23, 11740. [Google Scholar] [CrossRef]
- Miller, C.H.T.; Maher, S.G.; Young, H.A. Clinical Use of Interferon-γ. Ann. N.Y. Acad. Sci. 2009, 1182, 69–79. [Google Scholar] [CrossRef]
- Baessler, A.; Vignali, D.A.A. T Cell Exhaustion. Annu. Rev. Immunol. 2024, 42, 179–206. [Google Scholar] [CrossRef]
- Sonar, S.A.; Watanabe, M.; Nikolich, J.Ž. Disorganization of secondary lymphoid organs and dyscoordination of chemokine secretion as key contributors to immune aging. Semin. Immunol. 2023, 70, 101835. [Google Scholar] [CrossRef]
- Gautam, S.; Kumar, R.; Singh, N.; Singh, A.K.; Rai, M.; Sacks, D.; Sundar, S.; Nylén, S. CD8 T Cell Exhaustion in Human Visceral Leishmaniasis. J. Infect. Dis. 2014, 209, 290–299. [Google Scholar] [CrossRef]
- Silva-O’Hare, J.; de Oliveira, I.S.; Klevorn, T.; Almeida, V.A.; Oliveira, G.G.S.; Atta, A.M.; de Freitas, L.A.R.; Dos-Santos, W.L.C. Disruption of Splenic Lymphoid Tissue and Plasmacytosis in Canine Visceral Leishmaniasis: Changes in Homing and Survival of Plasma Cells. PLoS ONE 2016, 11, e0156733. [Google Scholar] [CrossRef] [PubMed]
- Hermida, M.d.-R.; de Melo, C.V.B.; Lima, I.d.S.; Oliveira, G.G.d.S.; Dos-Santos, W.L.C. Histological Disorganization of Spleen Compartments and Severe Visceral Leishmaniasis. Front. Cell. Infect. Microbiol. 2018, 8, 394. [Google Scholar] [CrossRef] [PubMed]
- de Souza, T.L.; da Silva, A.V.A.; Pereira, L.d.O.R.; Figueiredo, F.B.; Mendes Junior, A.A.V.; Menezes, R.C.; Mendes-da-Cruz, D.A.; Boité, M.C.; Cupolillo, E.; Porrozzi, R.; et al. Pro-Cellular Exhaustion Markers are Associated with Splenic Microarchitecture Disorganization and Parasite Load in Dogs with Visceral Leishmaniasis. Sci. Rep. 2019, 9, 12962. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Liu, L.; Gao, J.; Yang, Y.; Hu, C.; Guo, B.; Zhu, B. T lymphocytes maintain structure and function of fibroblastic reticular cells via lymphotoxin (LT)-B. BMC Immunol. 2014, 15, 33. [Google Scholar] [CrossRef]
- Montes de Oca, M.; de Labastida Rivera, F.; Winterford, C.; Frame, T.C.M.; Ng, S.S.; Amante, F.H.; Edwards, C.L.; Bukali, L.; Wang, Y.; Uzonna, J.E.; et al. IL-27 signalling regulates glycolysis in Th1 cells to limit immunopathology during infection. PLOS Pathog. 2020, 16, e1008994. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Characteristic | Number (%) | 95% CI 1 |
|---|---|---|
| Sex | ||
| Male | 42 (58.3) | 46.1; 69.85 |
| Female | 30 (41.7) | 30.2; 53.89 |
| Age groups (Years) | ||
| <2 | 17 (23.6) | 14.0; 35.0 |
| 2 < 4 | 5 (6.9) | 2.2; 15.4 |
| 4 < 15 | 22 (30.6) | 20.2; 42.5 |
| 15 < 40 | 22 (30.6) | 20.2; 42.5 |
| 40+ | 6 (8.3) | 3.1; 17.3 |
| HIV 2 (number, %) | 13 (18.6) | 10.3; 30.0 |
| Deaths (number, %) | 4 (5.6) | 1.5; 13.61 |
| Chance of death > 10% by Kala-Cal® | 25 (34.7) | 23.9; 46.9 |
| Hemorrhages or infections | 31 (43.7) | 31.9; 56.0 |
| Reported bleeding | 4 (5.6%) | 1.5; 13.6 |
| Detected bleeding | 15 (20.8) | 12.2; 32.0 |
| Sepsis | 10 (14.1) | 7.0; 24.4 |
| Any bacterial infection | 23 (31.9) | 21.4; 44.0 |
| Variables | Median | Interquartile Intervals | Mean | Reference Values (Median) | Kolmogorov–Smirnov Test (p-Value) |
|---|---|---|---|---|---|
| Plasma kDNA (AEq 1/mL) | 856.7 | 145.5–3527.9 | 3515.4 | 0 3 | 0.000 |
| Bone marrow kDNA (HCEq 2) | 55.7 | 3.6–4008 | 889.9 | 0 3 | 0.000 |
| IL-1β pg/mL | 0.9 | 0.2–2.1 | 2.0 | 0.18 (0–3.66) 4 | 0.000 |
| IL-6 pg/mL | 9.5 | 2.4–28.0 | 41.7 | 0 (0–0) 4 | 0.000 |
| IL-8 pg/mL | 26.2 | 9.8–145.5 | 146.9 | 0 (0–0) 4 | 0.000 |
| IL-10 pg/mL | 18.4 | 8.7–35 | 30.2 | 0 (0–0) 4 | 0.000 |
| IL-12 pg/mL | 1.2 | 0.0–2.5 | 1.8 | 0 (0–0) 4 | 0.000 |
| TNF-α pg/mL | 1.0 | 0.3–3.0 | 2.2 | 0 (0–0) 4 | 0.000 |
| TGF-β ng/mL | 23.6 | 11.2–42.4 | 39.6 | NA 5 | - |
| Markers of Severe Disease (Number of Patients) | Plasma Load (AEq) 1 Median, (Mean) | p-Value 2 | Bone Marrow Load (AEq/109HCEq) 3 Median, (Mean) | p-Value |
|---|---|---|---|---|
| Age (years) | ||||
| <15 (27) | 508 (2513) | 27 (519) | ||
| 15+ (44) | 2679 (5091) | <0.05 | 137 (1494) | <0.05 |
| Sex | ||||
| Female (30) | 329 (1580) | 28 (631) | ||
| Male (41) | 1452 (4898) | <0.05 | 90 (1079) | >0.05 |
| HIV | ||||
| Yes (13) | 4207 (7016) | 56 (2867) | ||
| No (57) | 511 (2768) | <0.01 | 35 (500) | >0.05 |
| Hospital outcome | ||||
| Death (4) | 11,826 (11,925) | 487 (605) | ||
| Survival (67) | 830 (3020) | >0.05 | 56 (907) | >0.05 |
| Chance of death > 10% according to Kala-Cal® | ||||
| >10% (25) | 3532 (6617) | 290 (2280) | ||
| <10% (46) | 311 (1866) | <0.001 | 20 (350) | <0.001 |
| Reported bleeding | ||||
| Yes (4) | 8855 (10,440) | 89 (406) | ||
| No (67) | 830 (3108) | >0.05 | 56 (919) | >0.05 |
| Detected bleeding | ||||
| Yes (15) | 823 (4427) | 214 (1406) | ||
| No (56) | 876 (3276) | >0.05 | 21 (752) | <0.05 |
| Sepsis | ||||
| Yes (10) | 888 (5619) | 247 (375) | ||
| No (60) | 1130 (3210) | >0.05 | 34 (988) | >0.05 |
| Any bacterial infection | ||||
| Yes (23) | 837 (3679) | 425 (466) | ||
| No (48) | 1059 (3439) | >0.05 | 65 (1093) | >0.05 |
| Variables (Number of Patients) | IL-1β Median, (Mean) | p-Value 1 | IL-6 Median, (Mean) | p-Value | IL-8 Median, (Mean) | p-Value | IL-10 Median, (Mean) | p-Value | IL-12 Median, (Mean | p-Value | TNF-α Median, (Mean) | p-Value | TGF-β Median, (Mean) | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Age (years) | ||||||||||||||
| <15 (27) | 1.1 (2.1) | 14.0 (49.3) | 28.8 (130.7) | 23.4 (36.4) | 1.4 (2.1) | 1.2 (2.5) | 23.1 (40.5) | |||||||
| 15+ (44) | 0.4 (1.9) | >0.05 | 7.8 (28.9) | >0.05 | 23.3 (174.2) | >0.05 | 12.8 (19.8) | <0.05 | 0.6 (1.3) | >0.05 | 0.9 (1.6) | >0.05 | 24.5 (38.0) | >0.05 |
| Sex | ||||||||||||||
| Female (30) | 0.9 (1.9) | 8.4 (55.9) | 25.6 (118.7) | 23.1 (32.6) | 1.0 (2.2) | 2.1 (3.1) | 13.9 (40.6) | |||||||
| Male (41) | 1.0 (2.0) | >0.05 | 13.6 (32.6) | >0.05 | 26.2 (164.8) | >0.05 | 13.6 (28.7) | >0.05 | 1.3 (1.6) | >0.05 | 0.9 (1.7) | >0.05 | 28.5 (38.9) | >0.05 |
| HIV-infection | ||||||||||||||
| Yes (12) | 1.3 (2.7) | 8.1 (13.0) | 46.7 (188.1) | 12.4 (14.3) | 1.2 (1.7) | 1.6 (1.9) | 16.5 (35.9) | |||||||
| No (57) | 0.9 (1.9) | >0.05 | 13.0 (48.8) | >0.05 | 25.4 (140.5) | >0.05 | 21.8 (34.2) | <0.05 | 1.3 (1.9) | >0.05 | 1.0 (2.3) | >0.05 | 24.05 (40.1) | >0.05 |
| Hospital outcome | ||||||||||||||
| Death (4) | 1.3 (1.3) | 24.7 (44.4) | 48.1 (173.9) | 12.0 (31.5) | 1.4 (1.8) | 1.2 (1.8) | 18.6 (20.4) | |||||||
| Survival (62) | 0.9 (2.1) | >0.05 | 9.1 (42.2) | >0.05 | 26.4 (147.5) | >0.05 | 18.7 (30.5) | >0.05 | 1.2 (1.8) | 0.61 | 1.0 (2.3) | >0.05 | 24.1 (20.4) | >0.05 |
| Chance of death 2 | ||||||||||||||
| >10% (23) | 1.0 (2.5) | 15.4 (44.3) | 65.2 (206.5) | 16.2 (27.8) | 1.1 (1.6) | 1.0 (2.3) | 24.5 (45.4) | |||||||
| <10% (43) | 0.9 (1.8) | >0.05 | 9.5 (44.3) | >0.05 | 24.6 (118.5) | >0.05 | 19.1 (32.0) | >0.05 | 1.3 (1.9) | >0.05 | 0.9 (2.2) | >0.05 | 22.3 (36.1) | >0.05 |
| Reported bleeding | ||||||||||||||
| Yes (3) | 1.6 (1.8) | 34.1 (63.7) | 78.6 (246.1) | 34.7 (51.3) | 2.0 (2.1) | 0.4 (1.8) | 15.7 (19.8) | |||||||
| No (63) | 0.9 (2.0) | >0.05 | 8.6 (41.2) | >0.05 | 24.9 (144.5) | >0.05 | 16.6 (29.5) | >0.05 | 1.2 (1.8) | >0.05 | 1.0 (2.3) | >0.05 | 23.6 (40.3) | >0.05 |
| Detected bleeding | ||||||||||||||
| Yes (13) | 0.9 (2.8) | 17.2 (24.9) | 88.5 (242.7) | 16.1 (24.1) | 2.0 (2.5) | 2.1 (2.2) | 42.1 (59.8) | |||||||
| No (53) | 1.0 (1.9) | >0.05 | 8.6 (46.6) | >0.05 | 24.6 (126.2) | >0.05 | 21.8 (32.1) | >0.05 | 1.2 (1.7) | >0.05 | 1.0 (2.3) | >0.05 | 20.9 (34.3) | >0.05 |
| Sepsis | ||||||||||||||
| Yes (10) | 1.6 (1.9) | 23.0 (137.3) | 110.2 (229.9) | 36.9 (37.3) | 1.9 (2.5) | 2.4 (2.5) | 18.3 (31.9) | |||||||
| No (56) | 0.8 (2.0) | >0.05 | 8.1 (25.3) | >0.05 | 24.7 (134.7) | >0.05 | 16.1 (29.3) | >0.05 | 0.9 (1.7) | <0.05 | 0.9 (2.2) | >0.05 | 25.7 (40.7) | >0.05 |
| Any bacterial infection | ||||||||||||||
| Yes (22) | 1.6 (2.4) | 25.2 (83.1) | 67.0 (203.9) | 30.5 (40.6) | 1.9 (2.1) | 2.4 (2.8) | 18.9 (33.0) | |||||||
| No (44) | 0.7 (1.8) | <0.05 | 7.2 (21.9) | <0.05 | 22.6 (121.8) | <0.05 | 16.0 (25.5) | <0.05 | 0.9 (1.7) | >0.05 | 0.8 (2.0) | <0.05 | 29.5 (42.5) | >0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sene, I.d.S.; Costa, D.L.; Zacarias, D.A.; dos Santos, J.C.; Ferreira, G.R.; Andrade, D.R.; Andrade, J.C.d.S.; Costa, C.H.N. Immune–Pathological Correlates of Disease Severity in New-World Kala-Azar: The Role of Parasite Load and Cytokine Profiles. Pathogens 2025, 14, 615. https://doi.org/10.3390/pathogens14070615
Sene IdS, Costa DL, Zacarias DA, dos Santos JC, Ferreira GR, Andrade DR, Andrade JCdS, Costa CHN. Immune–Pathological Correlates of Disease Severity in New-World Kala-Azar: The Role of Parasite Load and Cytokine Profiles. Pathogens. 2025; 14(7):615. https://doi.org/10.3390/pathogens14070615
Chicago/Turabian StyleSene, Ingridi de Souza, Dorcas Lamounier Costa, Daniele Alves Zacarias, Jailthon Carlos dos Santos, Gabriel Reis Ferreira, Daniela Rodrigues Andrade, Jorge Clarêncio de Sousa Andrade, and Carlos Henrique Nery Costa. 2025. "Immune–Pathological Correlates of Disease Severity in New-World Kala-Azar: The Role of Parasite Load and Cytokine Profiles" Pathogens 14, no. 7: 615. https://doi.org/10.3390/pathogens14070615
APA StyleSene, I. d. S., Costa, D. L., Zacarias, D. A., dos Santos, J. C., Ferreira, G. R., Andrade, D. R., Andrade, J. C. d. S., & Costa, C. H. N. (2025). Immune–Pathological Correlates of Disease Severity in New-World Kala-Azar: The Role of Parasite Load and Cytokine Profiles. Pathogens, 14(7), 615. https://doi.org/10.3390/pathogens14070615