


Complete Genome Sequences of Human Japanese Encephalitis Virus Genotype V Isolates in Korea Reveal Genotype-Specific Amino Acid Signatures


Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Diagnosis of Japanese Encephalitis
2.2. Specimens Included in the Study
2.3. Virus Isolation
2.4. Complete Genome Sequencing
2.5. Phylogenetic and Mutation Analyses
2.6. Structural Modeling and Visualization
3. Results
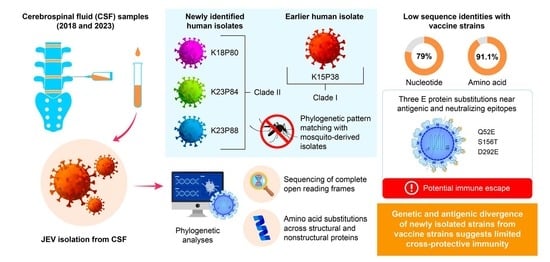
3.1. Phylogenetic Characterization and Clade Divergence of Human-Derived JEV GV Isolates in Korea
3.2. Genetic Divergence of Human JEV Genotype V Isolates from Vaccine and Other Genotypes
3.3. Distinct Amino Acid Substitutions and Molecular Signatures in Human-Derived JEV Genotype V Isolates
3.4. Molecular and Phylogenetic Continuity Between Human- and Mosquito-Derived JEV Genotype V Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| JEV | Japanese encephalitis virus |
| CSF | Cerebrospinal fluid |
| GV | Genotype V |
References
- Van den Eynde, C.; Sohier, C.; Matthijs, S.; De Regge, N. Japanese Encephalitis Virus Interaction with Mosquitoes: A Review of Vector Competence, Vector Capacity and Mosquito Immunity. Pathogens 2022, 11, 317. [Google Scholar] [CrossRef] [PubMed]
- Campbell, G.L.; Hills, S.L.; Fischer, M.; Jacobson, J.A.; Hoke, C.H.; Hombach, J.M.; Marfin, A.A.; Solomon, T.; Tsa, T.F.; Tsui, V.D.; et al. Estimated Global Incidence of Japanese Encephalitis: A Systematic Review. Bull. World Health Organ. 2011, 89, 766–774E. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Japanese Encephalitis. Fact Sheet. 2024. Available online: https://www.who.int/news-room/fact-sheets/detail/japanese-encephalitis (accessed on 3 September 2025).
- Cheng, Y.; Tran Minh, N.; Tran Minh, Q.; Khandelwal, S.; Clapham, H.E. Estimates of Japanese Encephalitis Mortality and Morbidity: A Systematic Review and Modeling Analysis. PLoS Neglected Trop. Dis. 2022, 16, e0010361. [Google Scholar] [CrossRef] [PubMed]
- Uchil, P.D.; Satchidanandam, V. Phylogenetic Analysis of Japanese Encephalitis Virus: Envelope Gene-Based Analysis Reveals a Fifth Genotype, Geographic Clustering, and Multiple Introductions of the Virus into the Indian Subcontinent. Am. J. Trop. Med. Hyg. 2001, 65, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Solomon, T.; Ni, H.; Beasley, D.W.; Ekkelenkamp, M.; Cardosa, M.J.; Barrett, A.D. Origin and Evolution of Japanese Encephalitis Virus in Southeast Asia. J. Virol. 2003, 77, 3091–3098. [Google Scholar] [CrossRef]
- Xia, Q.; Yang, Y.; Zhang, Y.; Zhou, L.; Ma, X.; Xiao, C.; Zhang, J.; Li, Z.; Liu, K.; Li, B.; et al. Shift in Dominant Genotypes of Japanese Encephalitis Virus and Its Impact on Current Vaccination Strategies. Front. Microbiol. 2023, 14, 1302101. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, J.S.; Williams, D.T.; van den Hurk, A.F.; Smith, D.W.; Currie, B.J. Japanese Encephalitis Virus: The Emergence of Genotype IV in Australia and Its Potential Endemicity. Viruses 2022, 14, 2480. [Google Scholar] [CrossRef] [PubMed]
- Li, M.-H.; Fu, S.-H.; Chen, W.-X.; Wang, H.-Y.; Guo, Y.-H.; Liu, Q.-Y.; Li, Y.-X.; Luo, H.-M.; Da, W.; Ji, D.Z.D.; et al. Genotype V Japanese Encephalitis Virus Is Emerging. PLoS Neglected Trop. Dis. 2011, 5, e1231. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.R.; Song, J.M.; Seo, S.U. Emerging Japanese Encephalitis Virus Genotype V in Republic of Korea. J. Microbiol. Biotechnol. 2022, 32, 955–959. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.H.; Jeong, Y.E.; Jo, J.E.; Shim, S.M.; Ryou, J.; Kim, K.C.; Lee, W.J.; Lee, J.Y. Genetic Characterization of Japanese Encephalitis Virus Genotype 5 Isolated from Patient, South Korea, 2015. Emerg. Infect. Dis. 2020, 26, 1002–1006. [Google Scholar] [CrossRef] [PubMed]
- Honjo, S.; Masuda, M.; Ishikawa, T. Effects of the Japanese Encephalitis Virus Genotype V-Derived Sub-Viral Particles on the Immunogenicity of the Vaccine Characterized by a Novel Virus-Like Particle-Based Assay. Vaccines 2019, 7, 81. [Google Scholar] [CrossRef]
- Lee, A.R.; Kim, W.J.; Choi, H.; Kim, S.H.; Hong, S.Y.; Shim, S.M.; Lee, H.I.; Song, J.M.; Kim, S.J.; Ishikawa, T.; et al. Genotype III-Based Japanese Encephalitis Vaccines Exhibit Diminished Neutralizing Response to Reemerging Genotype V. J. Infect. Dis. 2025, 231, 1281–1289. [Google Scholar] [CrossRef]
- Lee, A.-R.; Kim, S.-H.; Hong, S.-Y.; Lee, S.-H.; Oh, J.-S.; Lee, K.-Y.; Kim, S.-J.; Ishikawa, T.; Shim, S.-M.; Lee, H.-I.; et al. Characterization of Genotype V Japanese Encephalitis Virus Isolates from Republic of Korea. Emerg. Microbes Infect. 2024, 13, 2362392. [Google Scholar] [CrossRef] [PubMed]
- Luca, V.C.; AbiMansour, J.; Nelson, C.A.; Fremont, D.H. Crystal Structure of the Japanese Encephalitis Virus Envelope Protein. J. Virol. 2012, 86, 2337–2346. [Google Scholar] [CrossRef]
- Sun, Y.; Ding, H.; Zhao, F.; Yan, Q.; Li, Y.; Niu, X.; Zeng, W.; Wu, K.; Ling, B.; Fan, S.; et al. Genomic Characteristics and E Protein Bioinformatics Analysis of JEV Isolates from South China from 2011 to 2018. Vaccines 2022, 10, 1303. [Google Scholar] [CrossRef] [PubMed]
- Tajima, S.; Taniguchi, S.; Nakayama, E.; Maeki, T.; Inagaki, T.; Saijo, M.; Lim, C.K. Immunogenicity and Protective Ability of Genotype I-Based Recombinant Japanese Encephalitis Virus (JEV) with Attenuation Mutations in E Protein against Genotype V JEV. Vaccines 2021, 9, 1077. [Google Scholar] [CrossRef]
- Morita, K.; Tadano, M.; Nakaji, S.; Kosai, K.; Mathenge, E.G.M.; Pandey, B.D.; Hasebe, F.; Inoue, S.; Igarashi, A. Locus of a Virus Neutralization Epitope on the Japanese Encephalitis Virus Envelope Protein Determined by Use of Long PCR-Based Region-Specific Random Mutagenesis. Virology 2001, 287, 417–426. [Google Scholar] [CrossRef]
- Butrapet, S.; Childers, T.; Moss, K.J.; Erb, S.M.; Luy, B.E.; Calvert, A.E.; Blair, C.D.; Roehrig, J.T.; Huang, C.Y.-H. Amino Acid Changes within the E Protein Hinge Region That Affect Dengue Virus Type 2 Infectivity and Fusion. Virology 2011, 413, 118–127. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, P.; Cao, R.; Gu, J. Mutation of Putative N-Linked Glycosylation Sites in Japanese Encephalitis Virus Premembrane and Envelope Proteins Enhances Humoral Immunity in BALB/c Mice after DNA Vaccination. Virol. J. 2011, 8, 138. [Google Scholar] [CrossRef]
- Yun, S.-I.; Lee, Y.-M. Early Events in Japanese Encephalitis Virus Infection: Viral Entry. Pathogens 2018, 7, 68. [Google Scholar] [CrossRef] [PubMed]
- van den Elsen, K.; Chew, B.L.A.; Ho, J.S.; Luo, D. Flavivirus Nonstructural Proteins and Replication Complexes as Antiviral Drug Targets. Curr. Opin. Virol. 2023, 59, 101305. [Google Scholar] [CrossRef] [PubMed]
- Goh, J.Z.H.; De Hayr, L.; Khromykh, A.A.; Slonchak, A. The Flavivirus Non-Structural Protein 5 (NS5): Structure, Functions, and Targeting for Development of Vaccines and Therapeutics. Vaccines 2024, 12, 865. [Google Scholar] [CrossRef] [PubMed]
- Chanama, S.; Sukprasert, W.; Sa-ngasang, A.; A-nuegoonpipat, A.; Sangkitporn, S.; Kurane, I.; Anantapreecha, S. Detection of Japanese Encephalitis (JE) Virus-Specific IgM in Cerebrospinal Fluid and Serum Samples from JE Patients. Jpn. J. Infect. Dis. 2005, 58, 294–296. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Cheng, X.; Luo, M.; Shi, J. Japanese Encephalitis Virus: An Overview. J. Vector Borne Dis. 2025, 62, 16–23. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Common Substitutions (All Korean GV Isolates) | Clade I (K15P38) Specific | Clade II (K18P80, K23P84, K23P88) Specific | Functional Relevance (Predicted) |
|---|---|---|---|---|
| E (envelope) | Q52E, S156T, D292E | – | – | Antigenic sites; fusion domain; glycosylation-associated |
| NS1 | V94S, S105A, D177N | – | – | Replication complex formation; vascular permeability |
| NS2a | S96R, K187R | – | – | Membrane association; immune evasion |
| NS3 | V175I, Q249P, R269K, K304R, M586T | F31 | L31 | Protease/helicase activity; RNA replication |
| NS4b | S15N, R84K | – | – | Membrane interaction; innate immunity modulation |
| NS5a | K26R, A587V | – | K269R, M330I | RNA polymerase activity; replication fidelity |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, S.-R.; Lee, Y.-J.; Han, M.G.; Kim, H.M. Complete Genome Sequences of Human Japanese Encephalitis Virus Genotype V Isolates in Korea Reveal Genotype-Specific Amino Acid Signatures. Pathogens 2025, 14, 1279. https://doi.org/10.3390/pathogens14121279
Cho S-R, Lee Y-J, Han MG, Kim HM. Complete Genome Sequences of Human Japanese Encephalitis Virus Genotype V Isolates in Korea Reveal Genotype-Specific Amino Acid Signatures. Pathogens. 2025; 14(12):1279. https://doi.org/10.3390/pathogens14121279
Chicago/Turabian StyleCho, Seung-Rye, Ye-Ji Lee, Myung Guk Han, and Heui Man Kim. 2025. "Complete Genome Sequences of Human Japanese Encephalitis Virus Genotype V Isolates in Korea Reveal Genotype-Specific Amino Acid Signatures" Pathogens 14, no. 12: 1279. https://doi.org/10.3390/pathogens14121279
APA StyleCho, S.-R., Lee, Y.-J., Han, M. G., & Kim, H. M. (2025). Complete Genome Sequences of Human Japanese Encephalitis Virus Genotype V Isolates in Korea Reveal Genotype-Specific Amino Acid Signatures. Pathogens, 14(12), 1279. https://doi.org/10.3390/pathogens14121279