


Orientia tsutsugamushi Infection in Wild Small Mammals in Western Yunnan Province, China

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Setting and Sampling Period
2.2. Small Mammals Collection and DNA Extraction
2.3. NPCR Amplification and Sequencing
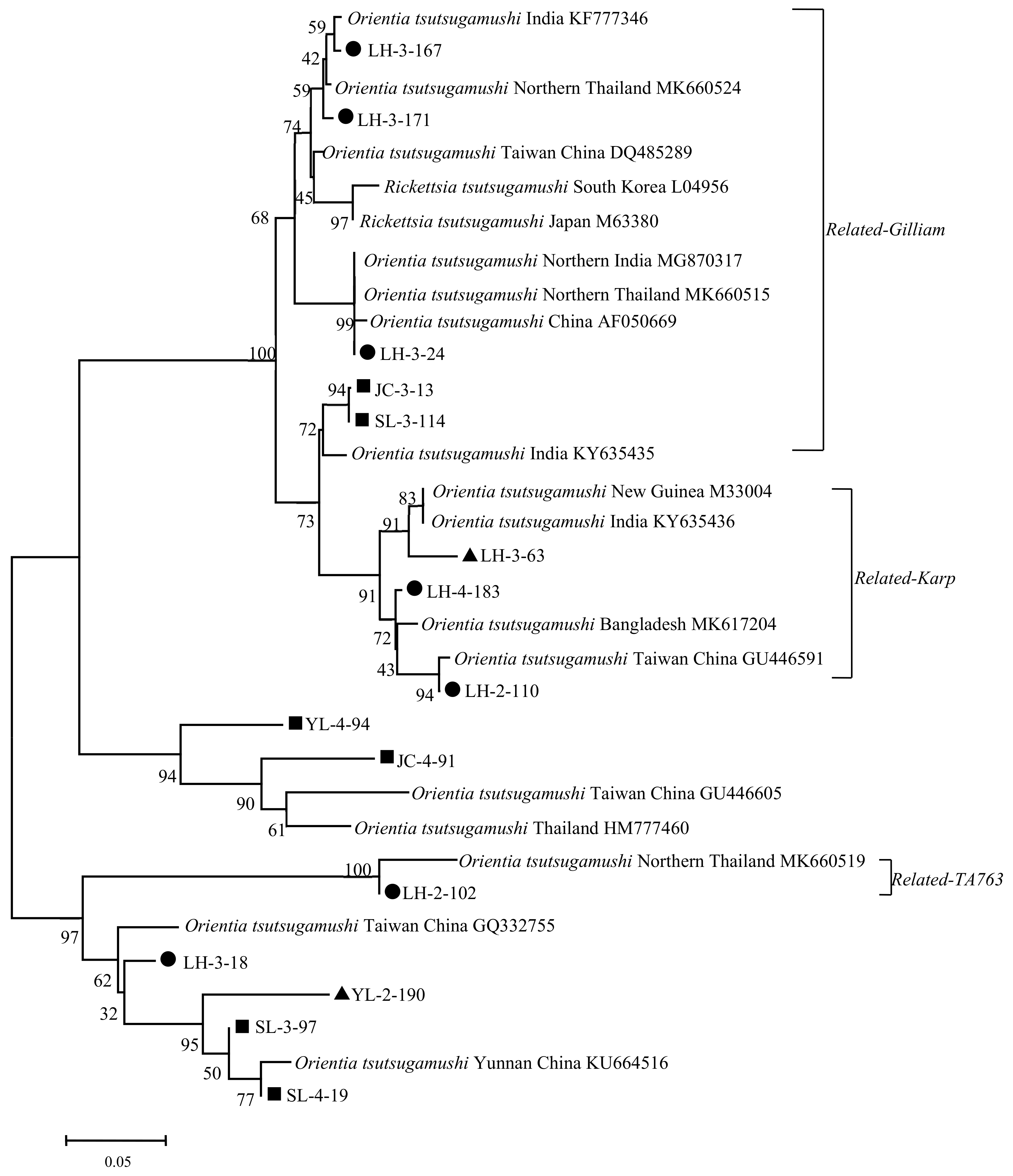
2.4. Phylogenetic Analysis
2.5. Statistical Analysis
3. Results
3.1. Species of Wild Small Mammals and Ot Infection
3.2. Phylogenetic Analysis Based on the 56-kDa TSA Gene of Ot
3.3. Associated Environmental and Seasonal Factors for Ot Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, J.; Richards, A.L. Scrub Typhus: No Longer Restricted to the Tsutsugamushi Triangle. Trop. Med. Infect. Dis. 2018, 3, 11. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Walker, D.H.; Jupiter, D.; Melby, P.C.; Arcari, C.M. A review of the global epidemiology of scrub typhus. PLoS Negl. Trop. Dis. 2017, 11, e0006062. [Google Scholar] [CrossRef]
- Abarca, K.; Martínez-Valdebenito, C.; Angulo, J.; Jiang, J.; Farris, C.M.; Richards, A.L.; Acosta-Jamett, G.; Weitzel, T. Molecular Description of a Novel Orientia Species Causing Scrub Typhus in Chile. Emerg. Infect. Dis. 2020, 26, 2148–2156. [Google Scholar] [CrossRef] [PubMed]
- Cosson, J.F.; Galan, M.; Bard, E.; Razzauti, M.; Bernard, M.; Morand, S.; Brouat, C.; Dalecky, A.; Bâ, K.; Charbonnel, N.; et al. Detection of Orientia sp. DNA in rodents from Asia, West Africa and Europe. Parasites Vectors 2015, 8, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.-Y.; Wang, L.-Y.; Ding, F.; Hu, W.-B.; Magalhaes, R.J.S.; Sun, H.-L.; Liu, Y.-X.; Liu, Q.-Y.; Huang, L.-Y.; Clements, A.C.A.; et al. Scrub Typhus in Mainland China, 2006–2012: The Need for Targeted Public Health Interventions. PLoS Neglected Trop. Dis. 2013, 7, e2493. [Google Scholar] [CrossRef]
- De, W.; Jing, K.; Huan, Z.; Qiong, Z.H.; Monagin, C.; Min, Z.J.; Ping, H.; Wen, K.C.; Yan, L.J. Scrub Typhus, a Disease with Increasing Threat in Guangdong, China. PLoS ONE 2015, 10, e0113968. [Google Scholar] [CrossRef]
- Kim, Y.B.; Choi, M.B.; Kwon, O. Bimonthly surveillance of wild rodents and chigger mites in urban parks in Daegu from 2018 to 2019. Entomol. Res. 2020, 50, 609–615. [Google Scholar] [CrossRef]
- Akhunji, B.; Bhate, R.; Pansare, N.; Chaudhari, S.P.; Khan, W.; Kurkure, N.V.; Kolte, S.W.; Barbuddhe, S.B. Distribution of Orientia tsutsugamushi in rodents and mites collected from Central India. Environ. Monit. Assess. 2019, 191, 82. [Google Scholar] [CrossRef]
- Elliott, I.; Pearson, I.; Dahal, P.; Thomas, N.V.; Roberts, T.; Newton, P.N. Scrub typhus ecology: A systematic review of Orientia in vectors and hosts. Parasites Vectors 2019, 12, 36. [Google Scholar] [CrossRef]
- Park, J.W.; Jung, S.H.; Ha, D.R.; Park, D.W.; Lee, J.M.; Kim, S.H.; Jeong, H.J.; Kim, N.M.; Gill, B.C.; Lee, J.Y.; et al. Molecular Epidemiology of an Orientia tsutsugamushi Gene Encoding a 56-kDa Type-Specific Antigen in Chiggers, Small Mammals, and Patients from the Southwest Region of Korea. Am. J. Trop. Med. Hyg. 2018, 98, 616–624. [Google Scholar] [CrossRef]
- Coleman, R.E.; Sangjun, N.; Frances, S.P.; Lerdthusnee, K.; Tanskul, P.; Monkanna, T.; Kollars, T.M.; Inlao, I.; Watcharapichat, P.; Khlaimanee, N.; et al. Occurrence of Orientia tsutsugamushi in Small Mammals from Thailand. Am. J. Trop. Med. Hyg. 2003, 69, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Hotta, K.; Pham, H.T.; Hoang, H.T.; Trang, T.C.; Vu, T.N.; Ung, T.T.; Shimizu, K.; Arikawa, J.; Yamada, A.; Nguyen, H.T.; et al. Prevalence and Phylogenetic Analysis of Orientia tsutsugamushi in Small Mammals in Hanoi, Vietnam. Vector Borne Zoonotic Dis. 2016, 16, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Latif, A.; Liu, B.-Y.; Chen, Z.; Sun, Y.; Shi, Y.-L.; Zong, J.; Li, J.-J.; Ren, C.-P.; Zhang, X.-C.; Liu, X.-N.; et al. Orientia tsutsugamushi infection in rodents in Anhui Province of China. Infect. Genet. Evol. 2017, 56, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhao, Z.-T.; Wang, X.-J.; Li, Z.; Ding, L.; Ding, S.-J.; Yang, H.-L. Genetic Variants of Orientia tsutsugamushiin Domestic Rodents, Northern China. Emerg. Infect. Dis. 2013, 19, 1135–1137. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-X.; Jia, N.; Xing, Y.-B.; Suo, J.-J.; Du, M.-M.; Jia, N.; Gao, Y.; Xie, L.-J.; Liu, B.-W.; Ren, S.-W. Consistency of the Key Genotypes of Orientia tsutsugamushi in Scrub Typhus Patients, Rodents, and Chiggers from a New Endemic Focus of Northern China. Cell Biochem. Biophys. 2013, 67, 1461–1466. [Google Scholar] [CrossRef] [PubMed]
- Ya, H.X.; Dong, W.X.; Wei, P.F.; Zhou, T.H.; Zhou, J.H.; Zhang, Y.Z.; Fan, R. Investigation on scrub typhus in Yongshan county, Yunnan Province, China. Chin. J. Zoonoses. 2018, 34, 255–259. [Google Scholar] [CrossRef]
- Li, L.L.; Zuo, S.Q.; Du, C.H.; Peng, H.H.; Zhao, Q.M.; Dai, K.; Zhang, J.S. Investigation of several bacterial pathogens on the wild small mammals in Gongshan area, Yunnan province. Acta Parasitol. Med. Entomol. Sin. 2018, 25, 32. [Google Scholar]
- Ya, H.X.; Zhang, Y.Z.; Wang, J.L. Investigation of Orientia tsutsugamushi infection in rodents and human in Yunnan. Dis. Surveil. 2016, 31, 145–148. [Google Scholar]
- Yin, J.-X.; Luo, Y.-Y.; Zhao, Q.-F.; Wei, Z.-F.; Hong, R.-D.; Liu, Z.-X.; Hong, M.; Xu, D.-D.; Wang, M.-D.; Zhou, Y.; et al. Comparative Analysis of Small Mammal and Parasitic Flea Composition in Yulong and Jianchuan Plague Foci in Yunnan Province, China: A Cross-Sectional Study. Biomed. J. Sci. Tech. Res. 2021, 37, 29901–29911. [Google Scholar] [CrossRef]
- Olson, N.D.; Morrow, J.B. DNA extract characterization process for microbial detection methods development and validation. BMC Res. Notes 2012, 5, 668. [Google Scholar] [CrossRef]
- Hall, B.G. Building Phylogenetic Trees from Molecular Data with MEGA. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Wei, Y.; Huang, Y.; Li, X.; Ma, Y.; Tao, X.; Wu, X.; Yang, Z. Climate variability, animal reservoir and transmission of scrub typhus in Southern China. PLoS Negl. Trop. Dis. 2017, 11, e0005447. [Google Scholar] [CrossRef] [PubMed]
- Ya, H.X.; Wang, J.L.; Zhong, Y.H. Investigation on rickettsiae natural infection in mice and vector arthropods in partial areas of Yunnan. Chin. J. Dis. Control. Pre. 2019, 23, 196–200. [Google Scholar] [CrossRef]
- Cao, M.; Che, L.; Zhang, J.; Hu, J.; Srinivas, S.; Xu, R.; Guo, H.; Zhang, Y.; Wang, C.; Feng, Y. Determination of Scrub typhus Suggests a New Epidemic Focus in the Anhui Province of China. Sci. Rep. 2016, 6, 20737. [Google Scholar] [CrossRef]
- Jun-Ke, J.; Dong, L.; Zheng-Da, G.; Zhi-Hou, G.; Hong-Yu, L.; Shou-Ying, K.; Xing-De, D.; Xi-Guang, F.; Mei, H.; Li-Yun, Z. Flea species diversity, spatial distribution and the relations to human plague in Yunnan Southwest moutain farmlan. Acta Parasitol. Med. Entomol. Sin. 2021, 28, 165–183. [Google Scholar]
- Yin, J.-X.; Cheng, X.-O.; Luo, Y.-Y.; Zhao, Q.-F.; Wei, Z.-F.; Xu, D.-D.; Wang, M.-D.; Zhou, Y.; Wang, X.-F.; Liu, Z.-X. The relationship between fleas and small mammals in households of the Western Yunnan Province, China. Sci. Rep. 2020, 10, 19. [Google Scholar] [CrossRef]
- Kuo, C.-C.; Lee, P.-L.; Chen, C.-H.; Wang, H.-C. Surveillance of potential hosts and vectors of scrub typhus in Taiwan. Parasites Vectors 2015, 8, 11. [Google Scholar] [CrossRef]
- Lu, M.; Li, F.; Liao, Y.; Shen, J.-J.; Xu, J.-M.; Chen, Y.-Z.; Li, J.-H.; Holmes, E.C.; Zhang, Y.-Z. Epidemiology and Diversity of Rickettsiales Bacteria in Humans and Animals in Jiangsu and Jiangxi provinces, China. Sci. Rep. 2019, 9, 19. [Google Scholar] [CrossRef]
- Park, J.W.; Yu, D.S.; Lee, G.S.; Seo, J.J.; Chung, J.K.; Lee, J.I. Epidemiological Characteristics of Rodents and Chiggers with Orientia tsutsugamushi in the Republic of Korea. Korean J. Parasitol. 2020, 58, 559–564. [Google Scholar] [CrossRef]
- Kinoshita, H.; Arima, Y.; Shigematsu, M.; Sunagawa, T.; Saijo, M.; Oishi, K.; Ando, S. Descriptive epidemiology of rickettsial infections in Japan: Scrub typhus and Japanese spotted fever, 2007–2016. Int. J. Infect. Dis. 2021, 105, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Yang, Z.; Dong, Z.; Wang, M. Meteorological factors and risk of scrub typhus in Guangzhou, southern China, 2006–2012. BMC Infect. Dis. 2014, 14, 139. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-C.; Qian, Q.; Magalhaes, R.J.S.; Han, Z.-H.; Hu, W.-B.; Haque, U.; Weppelmann, T.A.; Wang, Y.; Liu, Y.-X.; Li, X.-L.; et al. Spatiotemporal Dynamics of Scrub Typhus Transmission in Mainland China, 2006-2014. PLoS Neglected Trop. Dis. 2016, 10, e0004875. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.-R.; Tsai, H.-P.; Tsui, P.-Y.; Weng, M.-H.; Kuo, M.-D.; Lin, H.-C.; Chen, K.-C.; Ji, D.-D.; Chu, D.-M.; Liu, W.-T. Genetic Typing, Based on the 56-Kilodalton Type-Specific Antigen Gene, of Orientia tsutsugamushi Strains Isolated from Chiggers Collected from Wild-Caught Rodents in Taiwan. Appl. Environ. Microbiol. 2011, 77, 3398–3405. [Google Scholar] [CrossRef]


 Positive samples from Lianghe;
Positive samples from Lianghe;  Positive samples from Jianchuan;
Positive samples from Jianchuan;  Positive samples from Yulong.
Positive samples from Lianghe; Positive samples from Jianchuan; Positive samples from Yulong.
Positive samples from Yulong.
Positive samples from Lianghe; Positive samples from Jianchuan; Positive samples from Yulong. Positive Samples from Lianghe; Positive Samples from Jianchuan; Positive Samples from Yulong.
Positive Samples from Lianghe; Positive Samples from Jianchuan; Positive Samples from Yulong.
Positive Samples from Lianghe; Positive Samples from Jianchuan; Positive Samples from Yulong.
Positive Samples from Lianghe; Positive Samples from Jianchuan; Positive Samples from Yulong.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Primer Sequence | Amplified Fragments |
|---|---|---|
| Out Primer | ||
| a | 5’- TACATTAGCTGCGGGTATGACA-3’ | 306~339 bp |
| b | 5’- CCAGCATAATTCTTCAACCAAG-3’ | |
| Inner Primer | ||
| a’ | 5’- GAGCAGAGCTAGGTGTTATGTA-3’ | 150~168 bp |
| b’ | 5’- TAGGCATTATAGTAGGCTGAGG-3’ |
| Species | Lianghe County | Jianchuan County | Yulong County | Total |
|---|---|---|---|---|
| Apodemus chevrieri | 0 | 2/479 (0.42) | 0/305 | 2/784 (0.26) |
| Apodemus draco | 0 | 3/118 (2.54) | 3/158 (1.90) | 6/276 (2.17) |
| Apodemus latronum | 0 | 0/22 | 1/104 (0.96) | 1/126 (0.79) |
| Rattus nitidus | 0/12 | 0 | 0/1 | 0/13 |
| Rattus norvegicus | 0 | 0/4 | 0 | 0/4 |
| Rattus rattus/sladeni | 9/122 (7.38) | 0/1 | 0 | 9/123 (7.32) |
| Rattus tanezumi | 5/182 (2.75) | 0/2 | 0/2 | 5/186 (2.69) |
| Niviventer confucianus | 0/2 | 0/9 | 1/44 (2.27) | 1/55 (1.82) |
| Niviventer fulvescens | 1/62 (1.61) | 0 | 0 | 1/62 (1.61) |
| Niviventer andersoni | 0/2 | 1/6 (16.67) | 0/3 | 1/11 (9.09) |
| Micromys minutes | 0 | 0/6 | 0 | 0/6 |
| Mus pahari | 0/73 | 0 | 0 | 0/73 |
| Vernaya fulva | 0 | 0 | 0/1 | 0/1 |
| Bandicota indica | 0/1 | 0 | 0 | 0/1 |
| Berylmys bowersi | 0/9 | 0 | 0 | 0/9 |
| Eothenomys miletus | 0/7 | 15/338 (4.44) | 0/140 | 15/485 (3.09) |
| Eothenomys proditor | 0 | 0 | 0/56 | 0/56 |
| Eothenomys Eleusis | 0/17 | 0 | 0 | 0/17 |
| Eothenomys melanogaster | 0 | 0/2 | 0 | 0/2 |
| Dremomys pernyi | 0 | 0/1 | 0/20 | 0/21 |
| Sciurotamias forresti | 0 | 0 | 0/1 | 0/1 |
| Collosciurus erythraeus | 0 | 0 | 0/1 | 0/1 |
| Suncus murinus | 1/46 (2.17) | 3 | 0 | 1/49 (2.04) |
| Sorex minutes | 0 | 0 | 0/2 | 0/2 |
| Crocidura dracula | 0/3 | 0/3 | 0/6 | 0/12 |
| Soriculus leucops | 0 | 0 | 0/4 | 0/4 |
| Anourosorex squamipes | 1/31 (3.23) | 0/1 | 0 | 1/32 (3.13) |
| Crocidura attenuate | 0/6 | 1/12 (8.33) | 0/6 | 1/24 (4.17) |
| Hylomys suillus | 1/51 (1.96) | 0 | 0 | 1/51 (1.96) |
| Tupaia belangeri | 0/4 | 0/24 | 0/23 | 0/51 |
| Total | 18/630 (2.86) | 22/1031 (2.13) | 5/877 (0.57) | 45/2538 (1.77) |
| Factors | Sample | Positive Sample | Infection Rate (%) | p |
|---|---|---|---|---|
| County | <0.05 | |||
| Lianghe | 630 | 18 | 2.86 | |
| Jianchuan | 1031 | 22 | 2.13 | |
| Yulong | 877 | 5 | 0.57 | |
| Season | 0.057 | |||
| Spring | 537 | 3 | 0.56 | |
| Summer | 548 | 8 | 1.46 | |
| Autumn | 806 | 19 | 2.36 | |
| Winter | 647 | 15 | 2.32 | |
| Landscape | 0.802 * | |||
| Woodland | 1661 | 27 | 1.63 | |
| Cultivation | 341 | 7 | 2.05 | |
| Scrub | 70 | 1 | 1.43 | |
| Cultivation & Woodland | 273 | 5 | 1.83 | |
| Cultivation & Scrub | 193 | 5 | 2.59 | |
| Species | <0.01 * | |||
| A. chevrieri | 784 | 2 | 0.26 | |
| E. miletus | 485 | 15 | 3.09 | |
| A. draco | 276 | 6 | 2.17 | |
| Other | 993 | 22 | 2.22 | |
| Sex | 0.906 | |||
| Male | 1169 | 20 | 1.71 | |
| Female | 1354 | 24 | 1.77 |
| County | Elevation (m) | Sample | Positive Sample | Infection Rate (%) | p |
|---|---|---|---|---|---|
| Lianghe | 0.009 * | ||||
| 1000~1200 | 175 | 0 | 0 | ||
| 1200~1400 | 206 | 10 | 4.85 | ||
| 1400~1600 | 156 | 6 | 3.85 | ||
| ≥1600 | 93 | 2 | 2.15 | ||
| Jiangchuan | 0.822 * | ||||
| 2250~2450 | 154 | 2 | 1.3 | ||
| 2450~2650 | 646 | 15 | 2.32 | ||
| ≥2650 | 231 | 5 | 2.16 | ||
| Yulong | 0.013 * | ||||
| 2400~2600 | 81 | 1 | 1.23 | ||
| 2600~2800 | 115 | 1 | 0.87 | ||
| 2800~3000 | 169 | 3 | 1.78 | ||
| ≥3000 | 512 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Y.-Y.; Liu, S.-T.; He, Q.-N.; Hong, R.-D.; Zhu, J.-J.; Ai, Z.-Q.; Yin, J.-X. Orientia tsutsugamushi Infection in Wild Small Mammals in Western Yunnan Province, China. Pathogens 2023, 12, 128. https://doi.org/10.3390/pathogens12010128
Luo Y-Y, Liu S-T, He Q-N, Hong R-D, Zhu J-J, Ai Z-Q, Yin J-X. Orientia tsutsugamushi Infection in Wild Small Mammals in Western Yunnan Province, China. Pathogens. 2023; 12(1):128. https://doi.org/10.3390/pathogens12010128
Chicago/Turabian StyleLuo, Yun-Yan, Si-Tong Liu, Qi-Nan He, Ru-Dan Hong, Jun-Jie Zhu, Zhi-Qiong Ai, and Jia-Xiang Yin. 2023. "Orientia tsutsugamushi Infection in Wild Small Mammals in Western Yunnan Province, China" Pathogens 12, no. 1: 128. https://doi.org/10.3390/pathogens12010128
APA StyleLuo, Y.-Y., Liu, S.-T., He, Q.-N., Hong, R.-D., Zhu, J.-J., Ai, Z.-Q., & Yin, J.-X. (2023). Orientia tsutsugamushi Infection in Wild Small Mammals in Western Yunnan Province, China. Pathogens, 12(1), 128. https://doi.org/10.3390/pathogens12010128