
Prevalence, Genetic Diversity and Factors Associated with Distribution of Listeria monocytogenes and Other Listeria spp. in Cattle Farms in Latvia

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Prevalence of Listeria spp. in the Farm Environment
2.2. Molecular Serotyping, Clonal Complexes (CCs) and Genetic Diversity of Listeria Species Isolated from the Farm Environment
2.3. Factors Associated with L. monocytogenes within-Farm Prevalence
3. Discussion
4. Materials and Methods
4.1. Sampling
4.2. Microbiological Testing and Confirmation of Listeria spp.
4.3. Whole Genome Sequencing
4.4. Questionnaire
4.5. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Quereda, J.J.; Leclercq, A.; Moura, A.; Vales, G.; Gómez-Martín, A.; García-Muñoz, A.; Thouvenot, P.; Tessaud-Rita, N.; Bracq-Dieye, H.; Lecuit, M. Listeria valentina sp. nov., isolated from a water trough and the faeces of healthy sheep. Int. J. Syst. Evol. Microbiol. 2020, 70, 5868–5879. [Google Scholar] [CrossRef]
- Engelbrecht, F.; Dominguez-Bernal, G.; Hess, J.; Dickneite, C.; Greiffenberg, L.; Lampidis, R.; Raffalsbauer, D.; Daniels, J.J.D.; Kreft, J.; Kaufmann, S.H.E.; et al. A novel PrfA-regulated chromosomal locus, which is specific for Listeria ivanovii, encodes two small, secreted internalins and contributes to virulence in mice. Mol. Microbiol. 1998, 30, 405–417. [Google Scholar] [CrossRef]
- Doganay, M. Listeriosis: Clinical presentation. FEMS Immunol. Med. Microbiol. 2003, 35, 173–175. [Google Scholar] [CrossRef]
- Bērziņš, A.; Terentjeva, M.; Korkeala, H. Prevalence and genetic diversity of Listeria monocytogenes in vacuum-packaged ready-to-eat meat products at retail markets in Latvia. J. Food. Prot. 2009, 72, 1283–1287. [Google Scholar] [CrossRef]
- Desai, A.N.; Anyoha, A.; Madoff, L.C.; Lassmann, B. Changing epidemiology of Listeria monocytogenes outbreaks, sporadic cases, and recalls globally: A review of ProMED reports from 1996 to 2018. Int. J. Infect. Dis. 2019, 84, 48–53. [Google Scholar] [CrossRef] [Green Version]
- Koskar, J.; Kramarenko, T.; Meremäe, K.; Kuningas, M.; Sõgel, J.; Mäesaar, M.; Anton, D.; Lillenberg, M.; Roasto, M. Prevalence and numbers of Listeria monocytogenes in various ready-to-eat foods over a 5-year period in Estonia. J. Food. Prot. 2019, 82, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Stea, E.C.; Purdue, L.M.; Jamieson, R.C.; Yost, C.K.; Hansen, L.T. Comparison of the prevalence and diversities of Listeria species and Listeria monocytogenes in an Urban and a Rural Agricultural Watershed. Appl. Environ. Microbiol. 2015, 81, 3812–3822. [Google Scholar] [CrossRef] [Green Version]
- Castro, H.; Jaakkonen, A.; Hakkinen, M.; Korkeala, H.; Lindström, M. Occurrence, persistence, and contamination routes of Listeria monocytogenes genotypes on three Finnish dairy cattle farms: A Longitudinal study. Appl. Environ. Microbiol. 2018, 84, e02000-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oevermann, A.; Zurbriggen, A.; Vandevelde, M. Rhombencephalitis caused by Listeria monocytogenes in humans and ruminants: A zoonosis on the rise? Interdiscip. Perspect. Infect. Dis. 2010, 2010, 632513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitman, K.J.; Bono, J.L.; Clawson, M.L.; Loy, J.D.; Sosilevac, J.M.; Arthur, T.M.; Ondrak, J.D. Genomic-based identification of environmental and clinical Listeria monocytogenes strains associated with an abortion outbreak in beef heifers. BMC Vet. Res. 2020, 16, 70. [Google Scholar] [CrossRef] [PubMed]
- Hellström, S.; Kiviniemi, K.; Autio, T.; Korkeala, H. Listeria monocytogenes is common in wild birds in Helsinki region and genotypes are frequently similar with those found along the food chain. J. Appl. Microbiol. 2008, 104, 883–888. [Google Scholar] [CrossRef]
- Queiroz, O.C.M.; Ogunade, I.M.; Weinberg, Z.; Adesogan, A.T. Silage review: Foodborne pathogens in silage and their mitigation by silage additives. J. Dairy Sci. 2018, 101, 4132–4142. [Google Scholar] [CrossRef]
- Mohammed, H.O.; Stipetic, K.; McDonough, P.L.; Ginzalez, R.N.; Nydam, D.V.; Atwill, E.R. Identification of potential on-farm sources of Listeria monocytogenes in herds of dairy cattle. Am. J. Vet. Res. 2009, 70, 383–388. [Google Scholar] [CrossRef]
- Nightingale, K.K.; Schukken, Y.H.; Nightingale, C.R.; Fortes, E.D.; Ho, A.J.; Her, Z.; Grohn, Y.T.; McDonough, P.L.; Wiedmann, M. Ecology and transmission of Listeria monocytogenes infecting ruminants and in the farm environment. Appl. Environ. Microbiol. 2004, 70, 4458–4467. [Google Scholar] [CrossRef] [Green Version]
- Ho, A.J.; Ivanek, R.; Gröhn, Y.T.; Nightingale, K.K.; Wiedmann, M. Listeria monocytogenes fecal shedding in dairy cattle shows high levels of day-to-day variation and includes outbreaks and sporadic cases of shedding of specific L. monocytogenes subtypes. Prev. Vet. Med. 2007, 80, 287–305. [Google Scholar] [CrossRef] [PubMed]
- Esteban, J.I.; Oporto, B.; Aduriz, G.; Juste, R.A.; Hurtado, A. Faecal shedding and strain diversity of Listeria monocytogenes in healthy ruminants and swine in Northern Spain. BMC Vet. Res. 2009, 5, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, P.R.; Lomonaco, S.; Bottero, M.T.; Dalmasso, A.; Dondo, A.; Grattarola, C.; Zuccon, F.; Iulini, B.; Knabel, S.J.; Capucchio, M.T.; et al. Ruminant rhombencephalitis-associated Listeria monocytogenes strains constitute a genetically homogeneous group related to human outbreak strains. Appl. Environ. Microbiol. 2013, 79, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Haley, B.J.; Sonnier, J.; Schukken, Y.H.; Karns, J.S.; Van Kessel, J.A. Diversity of Listeria monocytogenes within a U.S. dairy herd, 2004–2010. Foodborne Pathog. Dis. 2015, 12, 844–850. [Google Scholar] [CrossRef]
- Moura, A.; Criscuolo, A.; Pouseele, H.; Maury, M.M.; Leclercq, A.; Tarr, C.; Björkman, J.T.; Dallman, T.; Reimer, A.; Enouf, V.; et al. Whole-genome-based population biology and epidemiological surveillance of Listeria monocytogeens. Nat. Microbiol. 2016, 2, 16185. [Google Scholar] [CrossRef] [PubMed]
- Maury, M.M.; Tsai, Y.H.; Charlier, C.; Touchom, M.; Chenal-Francisque, V.; Leclercq, A.; Criscuolo, A.; Gaultier, C.; Roussel, S.; Brisabois, A.; et al. Uncovering Listeria monocytogenes hypervirulence by harnessing its biodiversity. Nat. Genet. 2016, 48, 308–313. [Google Scholar] [CrossRef] [Green Version]
- Dreyer, M.; Aguilar-Bultet, L.; Rupp, S.; Guldimann, C.; Stephan, R.; Schock, A.; Otter, A.; Schupbach, G.; Brisse, S.; Lecuit, M.; et al. Listeria monocytogenes sequence type 1 is predominant in ruminant rhombencephalitis. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Papic, B.; Golob, M.; Kušar, D.; Pate, M.; Zdovc, I. Source tracking on a dairy farm reveals a high occurrence of subclinical mastitis due to hypervirulent Listeria monocytogenes clonal complexes. J. Appl. Microbiol. 2019, 127, 1349–1361. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, S.; Lopez-Alonso, V.; Rodriguez, P.; Martinez-Suarez, V. The connection between persistent, disinfectant-resistant Listeria monocytogenes strains from two geographically separate Iberian pork processing plants: Evidence from comparative genome analysis. Appl. Environ. Microbiol. 2016, 82, 308–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabal, A.; Pietzka, A.; Huhulescu, S.; Allelberger, F.; Ruppitsch, W.; Schmid, D. Isolate-based surveillance of Listeria monocytogenes by Whole Genome Sequencing in Austria. Front. Microbiol. 2019, 10, 2282. [Google Scholar] [CrossRef]
- Bandelj, P.; Jamnikar-Ciglenecki, U.; Ocepek, M.; Blagus, R.; Vengust, M. Risk factors associated with fecal shedding of Listeria monocytogenes by dairy cows and calves. J. Vet. Intern. Med. 2018, 32, 1773–1779. [Google Scholar] [CrossRef] [PubMed]
- Borucki, M.K.; Gay, C.C.; Reynolds, J.; McElwain, K.L.; Kim, S.H.; Call, D.R.; Knowles, D.P. Genetic diversity of Listeria monocytogenes strains from a high-prevalence dairy farm. Appl. Environ. Microbiol. 2005, 71, 5893–5899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, E.; O’Mahony, T.; Clancy, M.; Dempsey, R.; O’Brien, M.; Jordan, K. Listeria monocytogenes in the Irish dairy farm environment. J. Food Prot. 2009, 72, 1450–1456. [Google Scholar] [CrossRef] [PubMed]
- Murinda, S.E.; Nguyen, L.T.; Nam, H.M.; Almeida, R.A.; Headrick, S.J.; Oliver, S.P. Detection of sorbitol-negative and sorbitol-positive Shiga toxin-producing Escherichia coli, Listeria monocytogenes, Campylobacter jejuni, and Salmonella spp. in dairy farm environmental samples. Foodborne Pathog. Dis. 2004, 1, 97–104. [Google Scholar] [CrossRef]
- Vilar, M.J.; Yus, E.; Sanjuán, M.L.; Diéguez, F.J.; Rodríguez-Otero, J.L. Prevalence of and risk factors for Listeria species on dairy farms. J. Dairy Sci. 2007, 90, 5083–5088. [Google Scholar] [CrossRef]
- Nucera, D.M.; Grassi, M.A.; Morra, P.; Piano, S.; Tabacco, E.; Borreani, G. Detection, identification, and typing of Listeria species from baled silages fed to dairy cows. J. Dairy Sci. 2016, 99, 6121–6133. [Google Scholar] [CrossRef]
- McDonald, P.; Henderson, N.; Heron, S. The Biochemistry of Silage, 2nd ed.; Chalcombe Publications: Bucks, UK, 1991; p. 340. [Google Scholar]
- Kim, S.W.; Haendiges, J.; Keller, E.N.; Myers, R.; Kim, A.; Lombard, J.E.; Karns, J.S.; Van Kessel, J.A.S.; Haley, B.J. Genetic diversity and virulence of Listeria monocytogenes recovered from bulk tank milk, milk filters, and milking equipment from dairies in the United States (2002 to 2014). PLoS ONE 2018, 13, e0197053. [Google Scholar] [CrossRef]
- Chen, Y.; Gonzalez-Escalona, N.; Hammack, T.S.; Allard, M.W.; Strain, E.A.; Brown, E.W. Core genome multilocus sequence typing for identification of globally distributed clonal groups and differentiations of outbreaks of strains of Listeria monocytogenes. Appl. Environ. Microbiol. 2016, 82, 6258–6272. [Google Scholar] [CrossRef] [Green Version]
- Knudsen, G.M.; Nielsen, J.B.; Marvig, R.L.; Ng, R.L.; Worning, P.; Westh, H.; Gram, L. Genome-wide-analyses of Listeria monocytogenes from food-processing plants reveal clonal diversity and date the emergence of persisting sequence types. Environ. Microbiol. Rep. 2017, 9, 428–440. [Google Scholar] [CrossRef] [Green Version]
- Rychli, K.; Stresl, B.; Szakmary-Brändle, A.; Strauß, A.; Wagner, M.; Schoder, D. Listeria monocytogenes isolated from illegally imported food products into the European Union harbor different virulence factor variants. Genes 2018, 9, 428. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Cheng, J.; Zhang, J.; Chen, Y.; Zeng, H.; Xue, L.; Lei, T.; Pang, R.; Wu, S.; Wu, H.; et al. Isolation, potential virulence, and population diversity of Listeria monocytogenes from meat and meat products in China. Front. Microbiol. 2019, 10, 946. [Google Scholar] [CrossRef] [Green Version]
- Knabel, S.J.; Reimer, A.; Verghese, B.; Lok, M.; Ziegler, J.; Farber, J.; Pagotto, F.; Graham, M.; Nadon, C.A.; Gilmour, M.W. Sequence typing confirms that a predominant Listeria monocytogenes clone caused human listeriosis cases and outbreaks in Canada from 1988 to 2010. J. Clin. Microbiol. 2012, 50, 1748–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuch, A.; Goc, A.; Belkiewicz, K.; Filipello, V.; Ronkiewicz, P.; Golebiewska, A.; Wrobel, I.; Kiedrowska, M.; Wasko, I.; Hryniewicz, W.; et al. Molecular diversity and antimicrobial susceptibility of Listeria monocytogenes isolates from invasive infections in Poland (1997–2013). Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Raschle, S.; Stephan, R.; Stevens, M.J.A.; Cernela, N.; Zurfluh, K.; Muchaamba, F.; Nüesch-Inderbinen, M. Environmental dissemination of pahogenic Listeria monocytogenes in flowing surface waters in Switzerland. Sci. Rep. 2021, 11, 1–11. [Google Scholar] [CrossRef]
- Papić, B.; Pate, M.; Félix, B.; Kušar, D. Genetic diversity of Listeria monocytogenes strains in ruminant abortion and rhomenchephalitis cases in comparison with the natural environment. BMC Microbiol. 2019, 19, 299. [Google Scholar] [CrossRef] [Green Version]
- Maury, M.M.; Bracq-Dieye, H.; Huang, L.; Vales, G.; Lavina, M.; Thouvenot, P.; Disson, O.; Leclercq, A.; Brisse, S.; Lecuit, M. Hypervirulent Listeria monocytogenes clones’ adaption to mammalian gut accounts for their association with dairy products. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Hassan, L.; Guard, C.L.; Mohammed, H.O. Feeding practices associated with the presence of Listeria monocytogenes: A case control study in New York state diaries. Dairy Food Environ. Sanit. 2002, 22, 326–332. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prjibelski, A.; Antipov, D.; Meleshko, D.; Lapidus, A.; Korobeynikov, A. Using SPAdes de novo assembler. Curr. Protoc. Bioinform. 2020, 70, e102. [Google Scholar] [CrossRef]
- Ragon, M.; Wirth, T.; Hollandt, F.; Lavenir, R.; Lecuit, M.; Le Monnier, A.; Brisse, S. A new perspective on Listeria monocytogenes evolution. PLoS Pathog. 2008, 4, e1000146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruppitsch, W.; Pietzka, A.; Prior, K.; Bletz, S.; Fernandez, H.L.; Allerberger, F.; Harmsen, D.; Mellmann, A. Defining and evaluating a core genome multilocus sequence typing scheme for whole-genome sequence-based typing of Listeria monocytogenes. J. Clin. Microbiol. 2015, 53, 2869–2876. [Google Scholar] [CrossRef] [Green Version]
- Jünemann, S.; Sedlazeck, F.J.; Prior, K.; Albersmeier, A.; John, U.; Kalinowski, J.; Mellmann, A.; Goesmann, A.; von Haeseler, A.; Stoye, J.; et al. Updating benchtop sequencing performance comparison. Nat. Biotechnol. 2013, 31, 294–296. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 15 April 2021).
- Bürkner, P.C. brms: An R package for Bayesian multilevel models using Stan. J. Stat. Softw. 2017, 80, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R.V. Emmeans: Estimated Marginal Means, aka Least-Squares Means. R Package Version 1.5.5-1. 2021. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 20 April 2021).
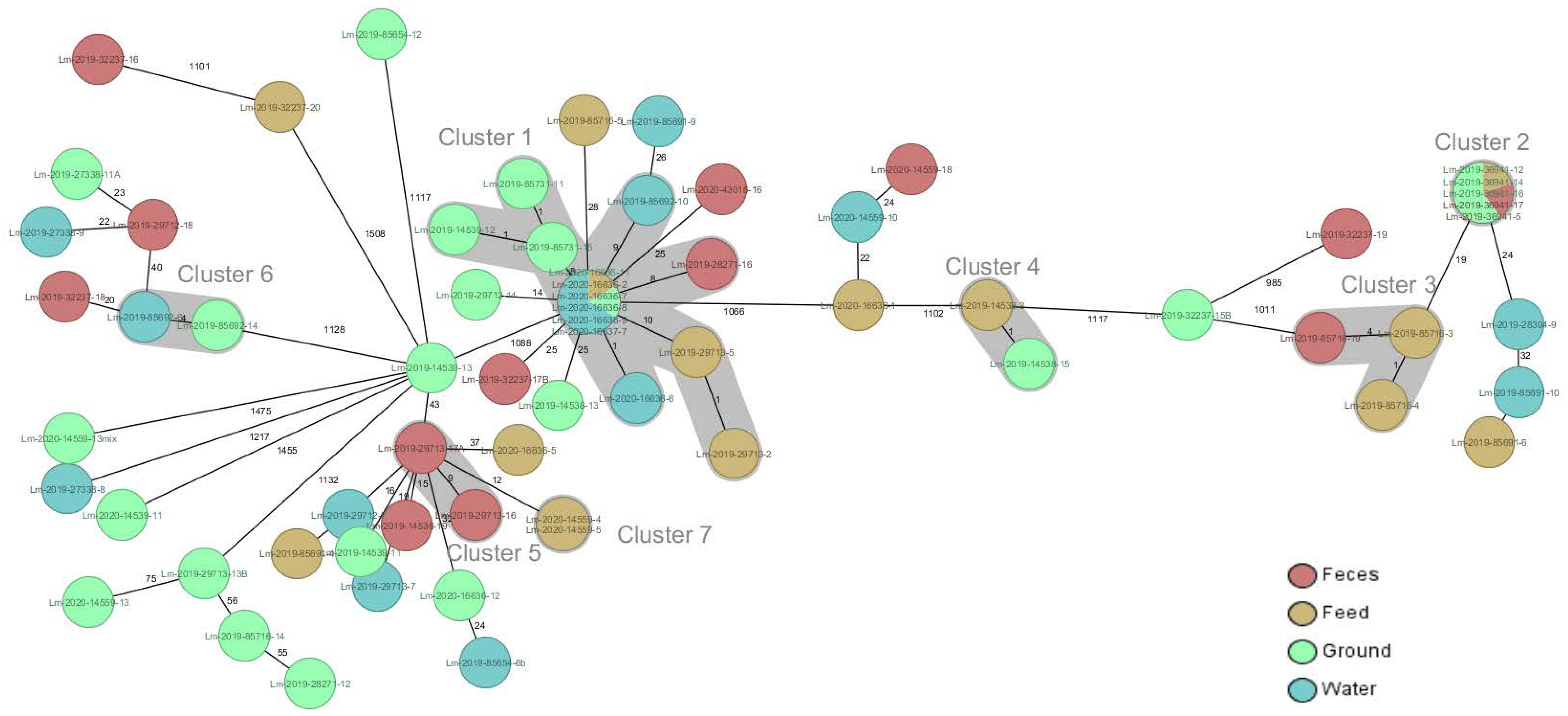

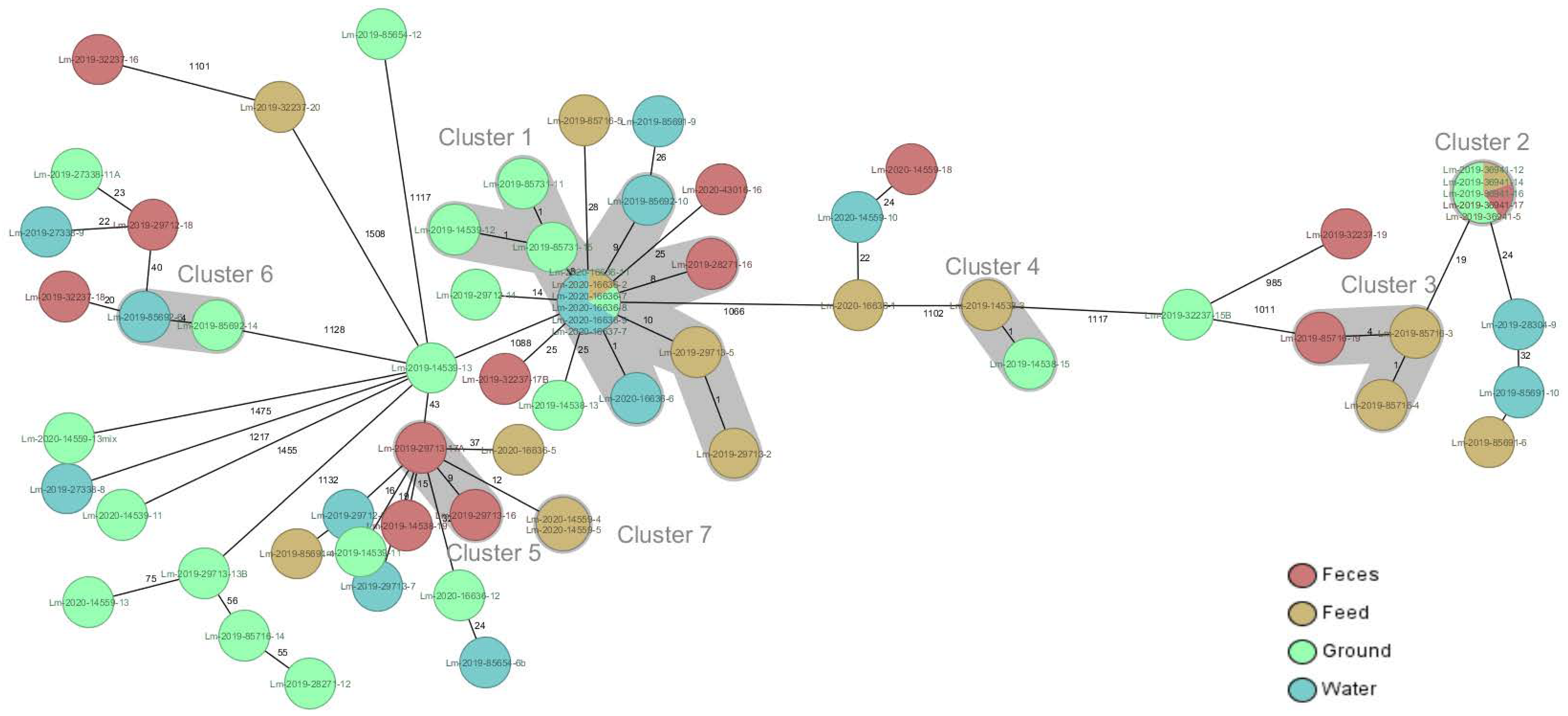{kind=link}
| Listeria Species | Sample | |||||||
|---|---|---|---|---|---|---|---|---|
| Soil (n = 133) | Feed (n = 141) | Water (n = 136) | Feces (n = 111) | |||||
| No. of Positive Samples (%) | 95% CI | No. of Positive Samples (%) | 95% CI | No. of Positive Samples (%) | 95% CI | No. of Positive Samples (%) | 95% CI | |
| L. monocytogenes | 25 (18.8) | 11.9–25.6 | 21 (14.9) | 9.5–21.9 | 26 (19.1) | 12.9–26.7 | 28 (25.2) | 17.5–34.4 |
| L. innocua a | 72 (54.1) | 45.3–62.8 | 51 (36.2) | 28.3–44.7 | 53 (38.9) | 30.7–47.7 | 37 (33.2) | 24.7–42.9 |
| L. seeligeri | 14 (10.5) | 5.8–17.0 | 11 (7.8) | 3.9–13.5 | 11 (8.1) | 4.1–14.0 | 17 (15.3) | 9.2–23.4 |
| L. fleishmanii | 1 (0.8) | 0.0–4.1 | 1 (0.8) | 0.0–3.9 | 0 (0) | 0 | 0 (0) | 0 |
| L. welshimeri | 0 (0) | 0.0–2.7 | 1 (0.8) | 0.0–3.9 | 2 (1.5) | 1.8–5.2 | 0 (0) | 0 |
| L. ivanovii | 0 (0) | 0.0–2.7 | 0 (0) | 0 | 2 (1.5) | 1.8–5.2 | 3 (2.7) | 0.6–7.7 |
| Total | 93 (69.9) | 61.3–77.6 | 71 (50.4) | 41.8–58.9 | 78 (57.4) | 48.6–65.8 | 65 (58.6) | 48.8–67.8 |
| L. monocytogenes | L. innocua | L. seeligeri | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Feed | No. of Samples | No. of Listeria Positive Samples (%) | No. of Positive Samples (%) | Serogroups | ST a | CC b | Mean log cfu/g | No. Positive Samples (%) | Cfu/g | No. of Positive Samples (%) | Cfu/g |
| TMR c | 5 | 3 (60) | 1 (20) | ND | - | - | 1.48 | 2 (40) | 4.48 | 0 (0) | - |
| By-products d | 4 | 2 (50) | 0 (0) | - | - | - | - | 2 (50) | 2.22 | 0 (0) | - |
| Pastures | 3 | 1 (33) | 0 (0) | - | - | - | - | 1 (33) | 2.04 | 0 (0) | - |
| Feed in feed trough | 34 | 28 (82) | 10 (29) | IIa, IVb | 18, 20, 37, 194, 451 | 5.15 | 22 (65) | 5.22 | 2 (6) | 5.00 | |
| Hay | 21 | 12 (57) | 6 (29) | IIa | 18, 37 | 18, 37 | 4.37 | 6 (29) | 2.2 | 4 (19) | 1.83 |
| Grains and flour e | 23 | 14 (61) | 2 (9) | IIa | 451 | 11 | 2.31 | 11 (48) | 3.74 | 1 (4) | 4.00 |
| Silage | 50 | 11 (22) | 2 (4) | ND | ND | ND | 4.65 | 7 (14) | 5.77 | 4 (8) | 6.06 |
| Feeding Table | 4 | 2 (50) | 0 (0) | - | - | - | - | 2 (50) | 3.54 | 0 (0) | - |
| Silage pit | 5 | 0 (0) | 0 (0) | - | - | - | - | 0 (0) | - | 0 (0) | - |
| Heap | 2 | 0 (0) | 0 (0) | - | - | - | - | 0 (0) | - | 0 (0) | |
| Haylage bales | 19 | 8 (42) | 1 (5) | - | - | - | 2.6 | 4 (50) | 6.00 | 4 (50) | 6.06 |
| Trench | 9 | 0 (0) | 0 (0) | - | - | - | - | 0 (0) | - | 0 (0) | - |
| Tunnel | 1 | 0 (0) | 0 (0) | - | - | - | - | 0 (0) | - | 0 (0) | - |
| Other | 9 | 1 (11) | 0 (0) | - | - | - | - | 1 (11) | 5.15 | 0 (0) | - |
| Straw | 1 | 0 (0) | 0 (0) | - | - | - | - | 0 (0) | - | 0 (0) | - |
| Total | 141 | 71 (50) | 21 (15) | 3.46 f | 51 (36) | 4.04 f | 15 (11) | 4.59 f | |||
| Sample | No. of Samples | Listeria spp. | L. monocytogenes | L. innocua | L. seeligeri | |||
|---|---|---|---|---|---|---|---|---|
| No. of Positive Sample (%) | Serogroup | ST | CC | No. of Positive Samples (%) | ||||
| Soil | ||||||||
| Near to pond | 3 | 1 (33) | 1 (33) | IIa | 37 | 37 | 0 (0) | 1 (33) |
| Near to farm | 7 | 4 (57) | 3 (43) | IIa | 7, 37 | 7, 37 | 3 (43) | 0 (0) |
| Near to manure a | 15 | 14 (93) | 3 (20) | IIa | 451 | 11 | 10 (67) | 3 (20) |
| Manure | 26 | 19 (73) | 5 (19) | IIa | 7 | 7 | 15 (58) | 2 (8) |
| Pasture | 21 | 14 (67) | 4 (19) | IIa, | 18, 37, 1085 | 18, 37, 1085 | 12 (46) | 1 (4) |
| Near feed storage | 22 | 15 (68) | 2 (9) | IIa | 8, 403 | 8, 403 | 13 (59) | 3 (14) |
| Bedding at farm | 25 | 16 (64) | 5 (20) | IIa | 18, 20, 21, 37 | 18, 20, 21, 37 | 11 (44) | 3 (12) |
| Straw and bedding in storage | 14 | 8 (57) | 1 (7) | IIa | 7 | 7 | 7 (50) | 1 (7) |
| Feed a | ||||||||
| Feeding table | 59 | 40 (68) | 14 (24) | IIa, IVb | 31,518, 20, 37, 194, 451 | 11, 18, 20, 27 | 5 (8) | |
| Pasture | 3 | 1 (33) | 0 (0) | - | - | - | 1 (33) | 0 (0) |
| Storage | 79 | 29 (37) | 7 (9) | IIa | 18, 37, 451 | 11, 18, 371 | 18 (23) | 6 (8) |
| Water b | ||||||||
| Farm | ||||||||
| -drinking bowl | 60 | 45 (75) | 13 (22) | IIa | 8, 37, 451, 1482 | 8, 11, 37, 1482 | 36 (60) | 5 (8) |
| -drinking trough | 32 | 22 (69) | 9 (28) | IIa | 8, 37, 451, 1482 | 8, 14, 37, 1482 | 13 (41) | 3 (9) |
| -other | 15 | 0 (0) | 0 (0) | 0 | - | - | 0 (0) | 0 (0) |
| Pasture | ||||||||
| -drinking trough | 10 | 3 (33) | 1 (10) | ND | - | - | 2 (20) | 0 (0) |
| -water body | 7 | 7 (100) | 3 (43) | IIa | 18, 37 | 18,37 | 1 (14) | 2 (28) |
| Other | 8 | 1 (12) | 0 (0) | - | - | - | 1 (12) | 0 (0) |
| Feces | 111 | 65 (59) | 28 (25) | IIa, IIc, IVb | 4, 8, 9, 18, 29, 37, 451 | 4, 8, 9, 11, 18, 29, 37 | 37 (33) | 17 (15) |
| Serogroup | ST/CC | Faeces | Feed | Soil | Water | Total |
|---|---|---|---|---|---|---|
| No. of Isolates | ||||||
| IIa | ST7/CC7 | 0 | 0 | 4 | 0 | 4 |
| ST8/CC8 | 2 | 0 | 2 | 2 | 6 | |
| ST451/CC11 | 4 | 4 | 2 | 3 | 13 | |
| ST399/CC14 | 0 | 0 | 0 | 1 | 1 | |
| ST18/CC18 | 2 | 4 | 3 | 1 | 11 | |
| ST20/CC20 | 0 | 1 | 1 | 0 | 2 | |
| ST21/CC21 | 0 | 0 | 1 | 0 | 1 | |
| ST29/CC29 | 1 | 0 | 0 | 1 | 2 | |
| ST37/CC37 | 2 | 4 | 7 | 7 | 20 | |
| ST403/CC403 | 0 | 0 | 1 | 0 | 1 | |
| ST1482/CC1482 | 0 | 0 | 0 | 2 | 2 | |
| ST1085/CC1085 | 0 | 0 | 1 | 0 | 1 | |
| IIc | ST9/CC9 | 1 | 0 | 0 | 0 | 0 |
| IVb | ST4/CC4 | 1 | 0 | 0 | 0 | 0 |
| ST194/CC315 | 0 | 1 | 0 | 0 | 0 | |
| Clonal Complex | Case Farm | Control Farm | Total |
|---|---|---|---|
| No. of Isolates | |||
| CC4 | 1 | 0 | 1 |
| CC7 | 2 | 2 | 4 |
| CC8 | 6 | 0 | 6 |
| CC9 | 1 | 0 | 1 |
| CC11 | 5 | 8 | 13 |
| CC14 | 1 | 0 | 1 |
| CC18 | 8 | 3 | 11 |
| CC20 | 0 | 2 | 2 |
| CC21 | 1 | 0 | 1 |
| CC29 | 0 | 2 | 2 |
| CC37 | 7 | 13 | 20 |
| CC315 | 1 | 0 | 1 |
| CC403 | 0 | 1 | 1 |
| CC1085 | 0 | 1 | 1 |
| CC1482 | 0 | 1 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terentjeva, M.; Šteingolde, Ž.; Meistere, I.; Elferts, D.; Avsejenko, J.; Streikiša, M.; Gradovska, S.; Alksne, L.; Ķibilds, J.; Bērziņš, A. Prevalence, Genetic Diversity and Factors Associated with Distribution of Listeria monocytogenes and Other Listeria spp. in Cattle Farms in Latvia. Pathogens 2021, 10, 851. https://doi.org/10.3390/pathogens10070851
Terentjeva M, Šteingolde Ž, Meistere I, Elferts D, Avsejenko J, Streikiša M, Gradovska S, Alksne L, Ķibilds J, Bērziņš A. Prevalence, Genetic Diversity and Factors Associated with Distribution of Listeria monocytogenes and Other Listeria spp. in Cattle Farms in Latvia. Pathogens. 2021; 10(7):851. https://doi.org/10.3390/pathogens10070851
Chicago/Turabian StyleTerentjeva, Margarita, Žanete Šteingolde, Irēna Meistere, Didzis Elferts, Jeļena Avsejenko, Madara Streikiša, Silva Gradovska, Laura Alksne, Juris Ķibilds, and Aivars Bērziņš. 2021. "Prevalence, Genetic Diversity and Factors Associated with Distribution of Listeria monocytogenes and Other Listeria spp. in Cattle Farms in Latvia" Pathogens 10, no. 7: 851. https://doi.org/10.3390/pathogens10070851
APA StyleTerentjeva, M., Šteingolde, Ž., Meistere, I., Elferts, D., Avsejenko, J., Streikiša, M., Gradovska, S., Alksne, L., Ķibilds, J., & Bērziņš, A. (2021). Prevalence, Genetic Diversity and Factors Associated with Distribution of Listeria monocytogenes and Other Listeria spp. in Cattle Farms in Latvia. Pathogens, 10(7), 851. https://doi.org/10.3390/pathogens10070851