1. Introduction
A major impediment to mass-producing insect predators in quantities that are necessary for augmentative biological control is encouraging females to oviposit their full potential of eggs when reared on non-prey foods [
1,
2]. In addition, chemical cues that guide females to suitable oviposition sites are often connected to natural prey [
3,
4]. In artificial rearing systems, many of these cues are completely lacking. Consequently, females decide to withhold some of their eggs or choose to lay them on other substrates, often of synthetic origin. Some commonly used substrates are tissue paper, filter paper, and aluminum sheets [
5,
6,
7]. In the absence of any oviposition substrates, females might opt to lay their eggs on the underside of the lid or sidewalls of the rearing container, vial, or Petri dish [
8,
9].
Very few plant-derived natural products are known to stimulate the oviposition in predators including ladybirds. An extract that was prepared from the leaves and wood of European barberry (
Berberis vulgaris L.) purportedly stimulated oviposition in the ladybird beetles,
Adalia bipunctata (L.) and
Coccinella septempunctata L., when sprayed on the leaves of sour cherry (
Prunus cerasus L.), which is not a preferred plant for oviposition [
10]. Wood panels, wood extracts, and several fractions from eastern redcedar (
Juniperus virginiana L.) stimulated oviposition in four ladybirds,
A.
bipunctata,
Coccinella transversoguttata Faldermann,
Cycloneda munda (Say), and
Coleomegilla maculata (DeGeer) [
11,
12]. Polyphenols in redcedar wood were thought to be the source of the compounds stimulating oviposition [
11,
12]. In our recent study, several fractions (containing polyphenols) that were isolated from redcedar heartwood stimulated oviposition by
C.
maculata in cage bioassays; some of the active fractions were bioflavonoids, such as taxifolin, quercetin, and naringenin (to a lesser extent) [
13]. Additional bioassays using synthetic compounds confirmed the strong, stimulatory effect of taxifolin and quercetin on oviposition by
C.
maculata. It was also found that quercetin had no negative effect on egg hatch rate [
13].
Quercetin is a flavonol with the chemical formula of C
15H
10O
7 and IUPAC name 2-(3,4-dihydroxyphenyl)-3,5,7-trihydroxychromen-4-one, and molecular weight of 302.238 g/mol (
https://pubchem.ncbi.nlm.nih.gov). It has been discovered in numerous plant species and is one of several bioflavonoids responsible for the color of flower petals, attracting pollinating insects to flowers (and pollen), and some herbivorous insects to their host plants [
14,
15,
16,
17]. It is also known to stimulate oviposition in monarch butterflies [
18]. Quercetin has repellent and insecticidal activity against some herbivorous insects, including aphids [
19].
In this study, we further investigate the effects of quercetin on oviposition by C. maculata. We tested the hypothesis that quercetin stimulates oviposition by C. maculata in the presence of conspecifics and a tissue substrate. The key objectives were to evaluate the effects of quercetin on (1) oviposition behavior and (2) female positioning in cages in the presence or absence of a preferred oviposition substrate, i.e., tissue paper.
2. Materials and Methods
2.1. Insect Colonies and Food Sources
The C. maculata stock colony originated from adults provided by USDA colleagues in Beltsville, MD and Brookings, SD, USA. All of the life stages were reared separately in plastic containers in a climate room (24 °C, 50–60% RH, and 16 h photophase, year-round). Immatures were kept in Petri dishes (2.5 cm high, 9.0 cm diam). Adult mating pairs were kept in plastic cages (10 cm high, 8.0 cm diam) with screened lids and crumbled facial tissue paper (Kleenex®, Kimberly-Clark Corp., Neenah, WI, USA) as an oviposition substrate. Egg clutches were harvested from tissue paper for experiments. At the time that we commenced this study, we had maintained this stock colony for 33 consecutive generations without introducing any wild-type individuals from the field. Adults and larvae were fed an excess of frozen-fresh eggs of the Mediterranean flour moth, Ephestia kuehniella Zeller (Lepidoptera: Pyralidae), twice a week. A microcentrifuge tube containing distilled water (stoppered with a cotton wad) was present in Petri dishes at all times. Similarly, a cotton wad that was moistened with distilled water was provided in rearing cages at all times.
2.2. Quercetin Source
We purchased “synthetic” quercetin (greater than 98.0% pure powder) from MedChem Express LLC (Princeton, NJ, USA). We assumed that the behavioral and ovipositional responses of C. maculata would not differ between synthetic and natural sources of quercetin.
2.3. Quercetin and Oviposition Behavior in Solitary Cages
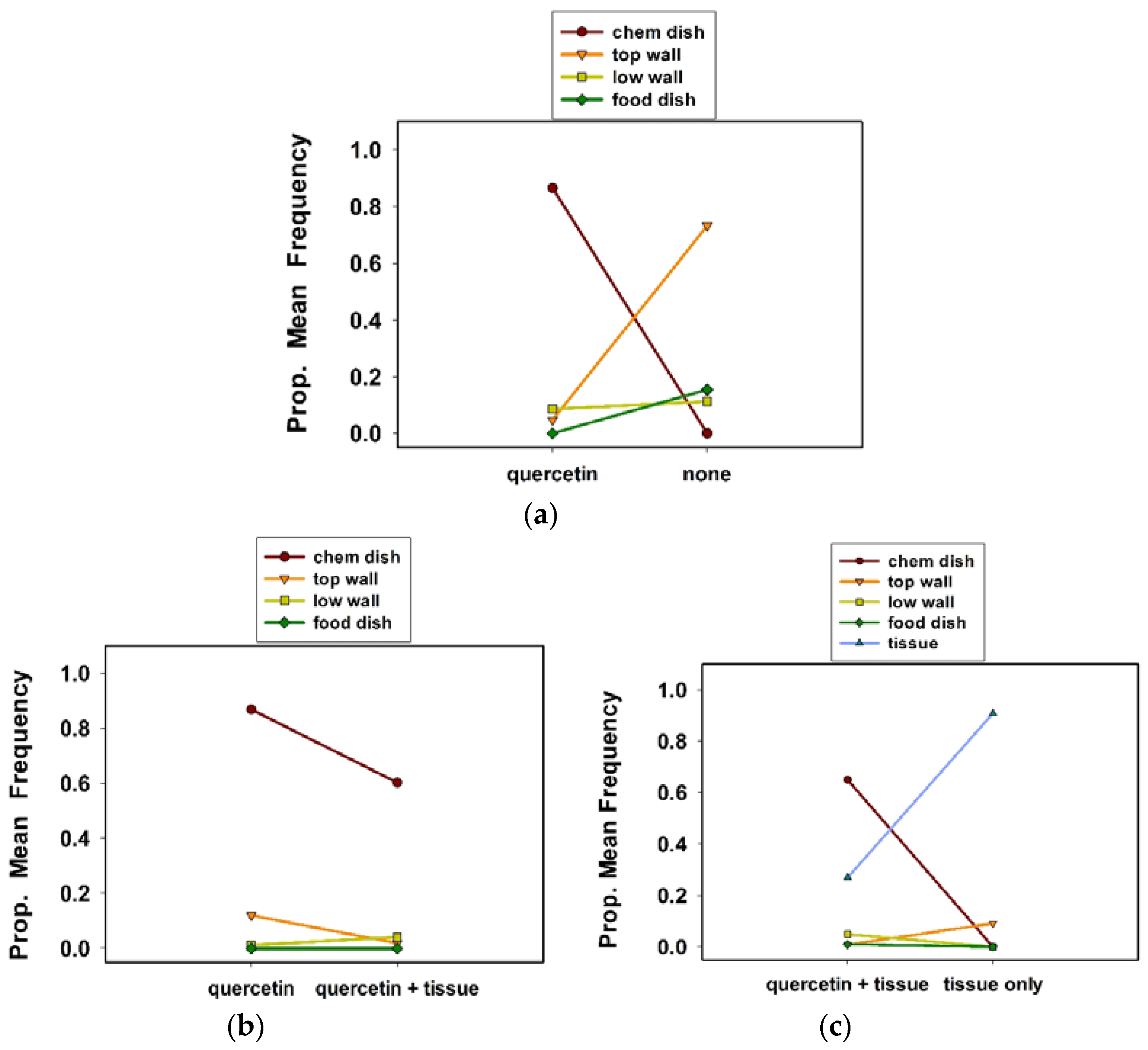
Our three initial experiments tested the ovipositional responses of individual females to quercetin in 500 mL “solitary” cages (7 × 11 cm, height, diam; see
Figure 1). Treatments included quercetin vs. none (experiment 1), quercetin vs. quercetin plus a tissue substrate (experiment 2), and quercetin plus a tissue substrate vs. a tissue substrate only (experiment 3). The tissue substrate was the same paper and from the same company, as used in our stock colony. Seven replicate cages were used for each treatment, for a total of 14 cages for each experiment. We measured oviposition site preference, egg clutch number per female, and eggs per clutch per female.
For each of the three experiments, we harvested approximately 100 one-month-old adults (combined sexes) from the same generation (33rd to 35th generations) from our stock colony, and then divided them into two plastic containers (9.5 cm high, 11 cm diam) for 2–3 days to ensure mating. Next, we isolated females in a Petri dish with a cotton wad moistened with distilled water, but no food, for 24 h in a growth chamber, to standardize hunger levels amongst females destined for bioassays. Hungry beetles are known to lay fewer eggs than well-fed ones [
20].
Females were randomly selected from the plastic containers and then placed separately into the plastic solitary cages, one female per cage. All of the cages were provisioned with an excess of factitious food, i.e., E. kuehniella moth eggs in a tiny Petri dish (1.0 × 3.5 cm, height, diam) and distilled water in a microcentrifuge tube at the base of the cage. All of the cages contained another tiny Petri dish (a chemical dish, 1.0 × 3.5 cm) containing quercetin powder (1 mg) in the appropriate cage; the chemical dish was empty in cages not containing any quercetin.
Preliminary observations revealed that 1 mg of quercetin powder in the tiny dish were sufficient to illicit oviposition behavior of a single female. No other concentrations were used in these three experiments. Quercetin powder occasionally adhered to the body parts of females and their egg clutches in the dish. When this occurred, we added more quercetin to the chemical dish to ensure that approximately 1 mg was in the dish during the course of the experiments.
Twice a day, once in the morning and again in the afternoon, we recorded the oviposition site preferences of females by noting the location of egg clutches inside the cages. Oviposition sites included the chemical dish, food dish, cage wall (lower wall, top wall), and tissue substrate (see experiment 3). We also counted the number of egg clutches per cage, then removed the egg clutches from the cages, and counted the number of eggs per clutch, using a stereo zoom microscope. We continued this procedure for 12 consecutive days for the three experiments.
The solitary cages were maintained in a growth chamber (24 °C, 60% RH, 16 h photophase), but removed twice a day to record oviposition data. Adult females were fed every other day, using the same food source (E. kuehniella eggs), as used in the stock colony. Old food and waste (fecal material) were discarded every other day.
2.4. Quercetin, Oviposition, and Female Locale in Communal Cages
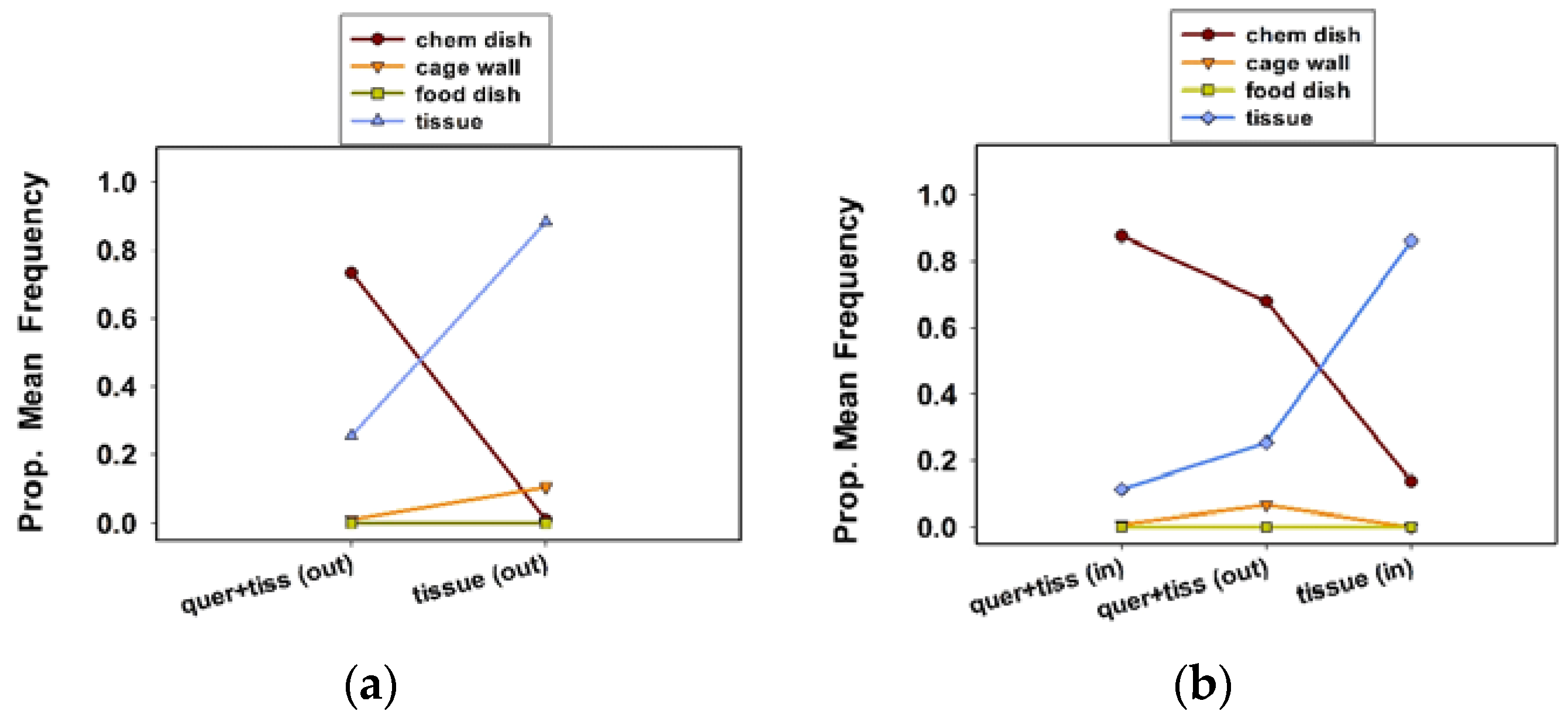
Our final two experiments tested the ovipositional responses of grouped females to quercetin in 1 L “communal” cages (
Figure 2). Cage dimensions were 13 cm × 11 cm (height, diam). A novel 4-unit oviposition chamber (
Figure 2) was used in these two experiments, to reduce female-female bodily contact, restrict the chemical dishes (with or without quercetin) to one locality, and increase the amount of quercetin in cages, to accommodate grouped females. The oviposition chamber consisted of four, stacked units (2 cm high, 6 cm diam each), with a chemical dish (1 cm high, 3.5 cm diam) in the center. Three holes of the same diameter were bored in the walls of each unit in order to provide easy access to the chemical dish. The tissue substrate was the same paper (and from the same company) as used in our stock colony. When inside the oviposition chamber, the tissue was three equal-sized, folded pieces (1.2 cm high, 4.0 cm diam), encircling the chemical dish (
Figure 2c). When outside the chamber, the lightly crumpled tissue substrate (20 cm × 20 cm sheet) occupied approximately 25% of the cage interior (
Figure 2d).
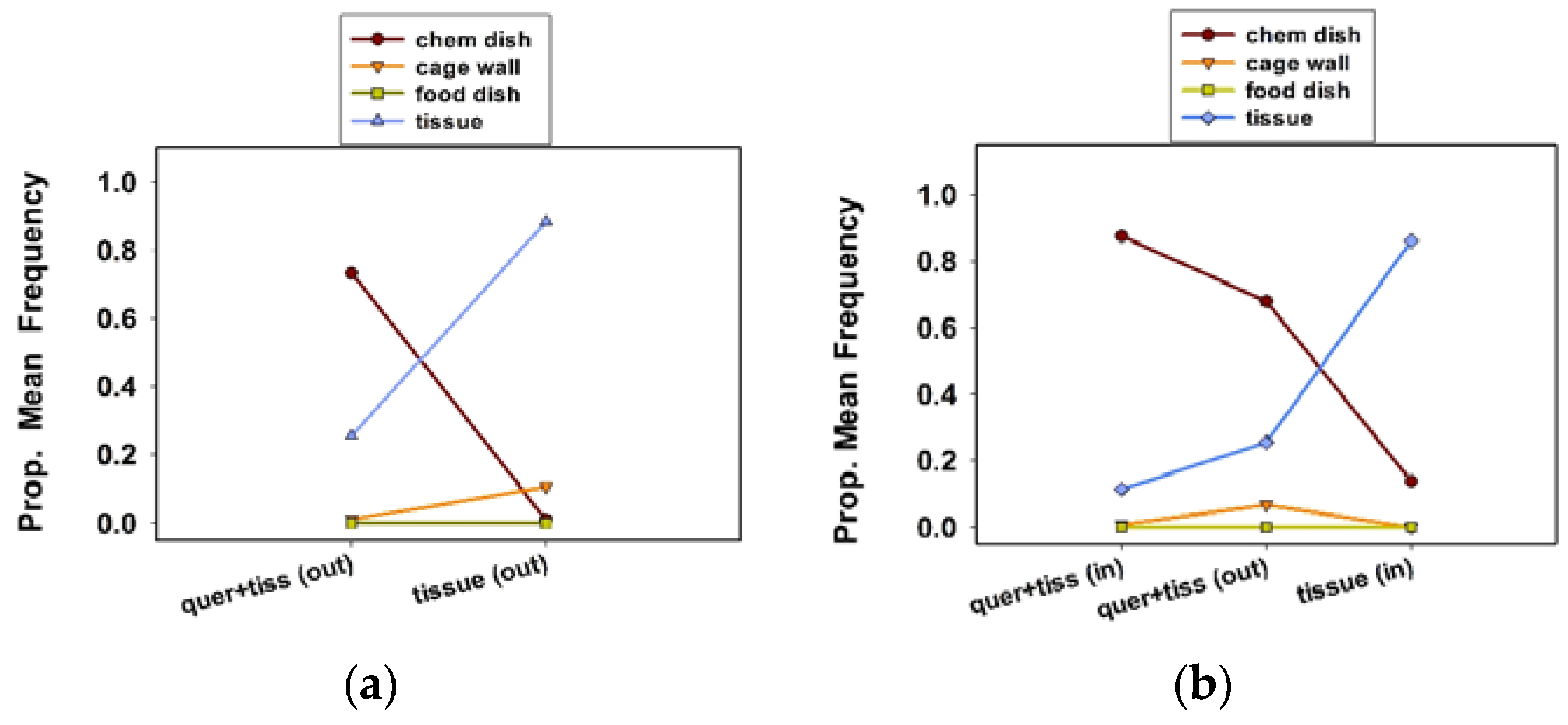
In experiment 4, two replicate 1 L cages were used for both treatments, for a total of four cages and 40 females. In experiment 5, two replicate 1 L cages were used for each of the three treatments, for a total of six cages and 60 females. We measured the oviposition site preference, egg clutch number per female, and eggs per clutch per female. Ten females were randomly assigned to each treatment cage. Treatments included quercetin plus tissue substrate (outside the chamber) vs. tissue substrate only (outside the chamber) in experiment 4, and quercetin plus tissue (in the chamber) vs. quercetin plus tissue substrate (outside), and tissue substrate only (inside the chamber) in experiment 5.
As described in the previous section, 100 one-month-old adults (mixed sexes) of the same generation were harvested from the stock colony and established in large containers to ensure mating, before the mated females were separated in Petri dishes (with water only) for 24 h to standardize hunger levels. As mentioned in the previous section, females were randomly assigned to cages that were provisioned with an excess of factitious food (
E.
kuehniella eggs in a tiny Petri dish), and distilled water in a microcentrifuge tube at the base of the cage. In experiments 4 and 5, a 4-unit oviposition chamber was used (see
Figure 2). Each chamber held a stack of four smaller Petri dishes, with each containing a tiny chemical dish. The four chemical dishes contained quercetin powder (2 mg) in the appropriate cage; the chemical dishes were empty in cages not containing any quercetin.
Based on preliminary assessments, the 2 mg of quercetin powder in the four stacked dishes was sufficient to elicit oviposition behavior of the 10 females that had easy access to the quercetin through entry/exit holes bored into the sides of each of the four units of the chamber. This was the only concentration used in these two experiments.
Twice a day, once in the morning and again in the afternoon, we recorded the oviposition site preferences of females by noting the location of egg clutches inside the cages. Oviposition sites included the chemical dish, food dish, cage wall (lower and top wall combined), and tissue substrate. As in the previous section, we counted the number of clutches per cage, removed them from the cages, and counted the eggs per clutch, using a stereo zoom microscope. We continued this procedure for 12 consecutive days for these final two experiments.
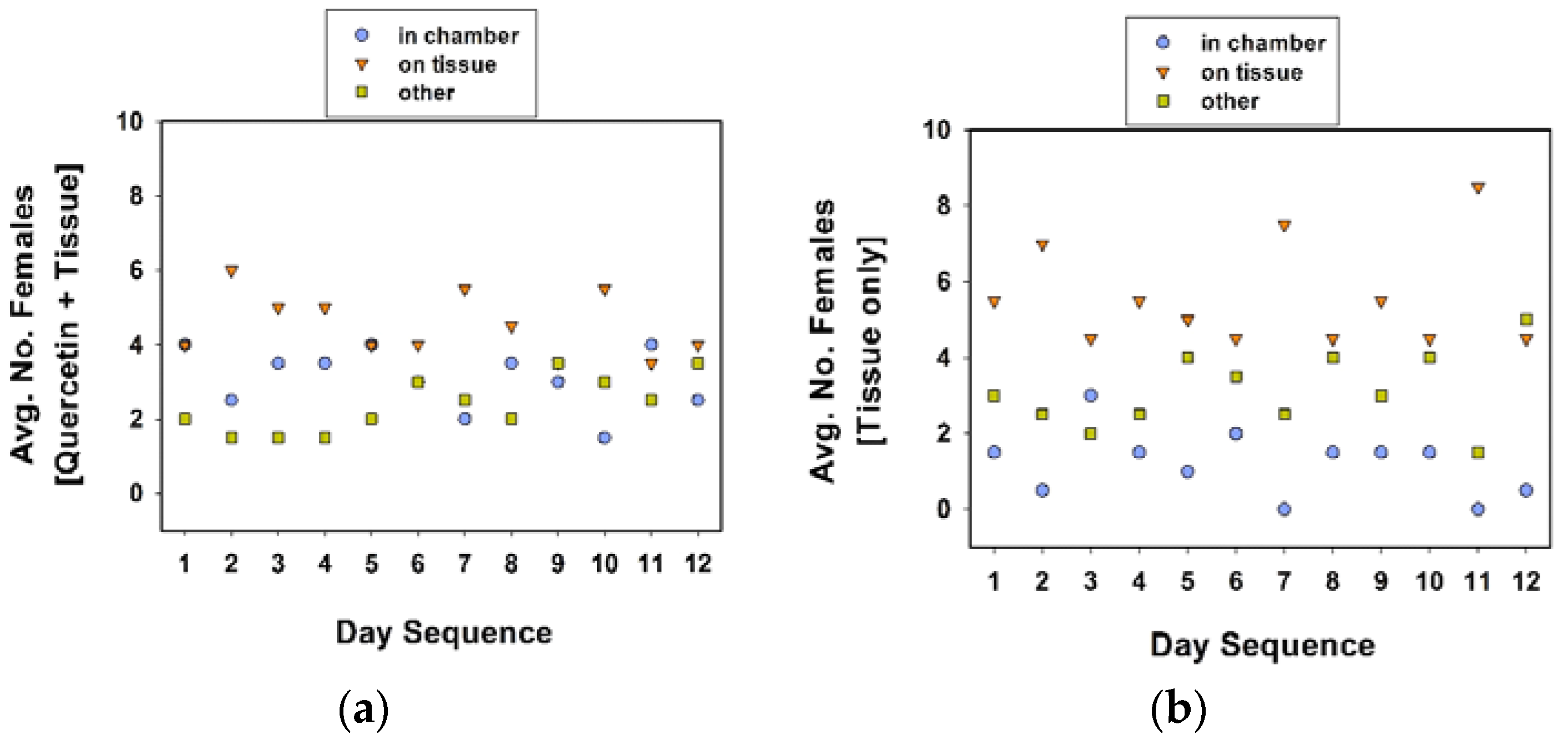
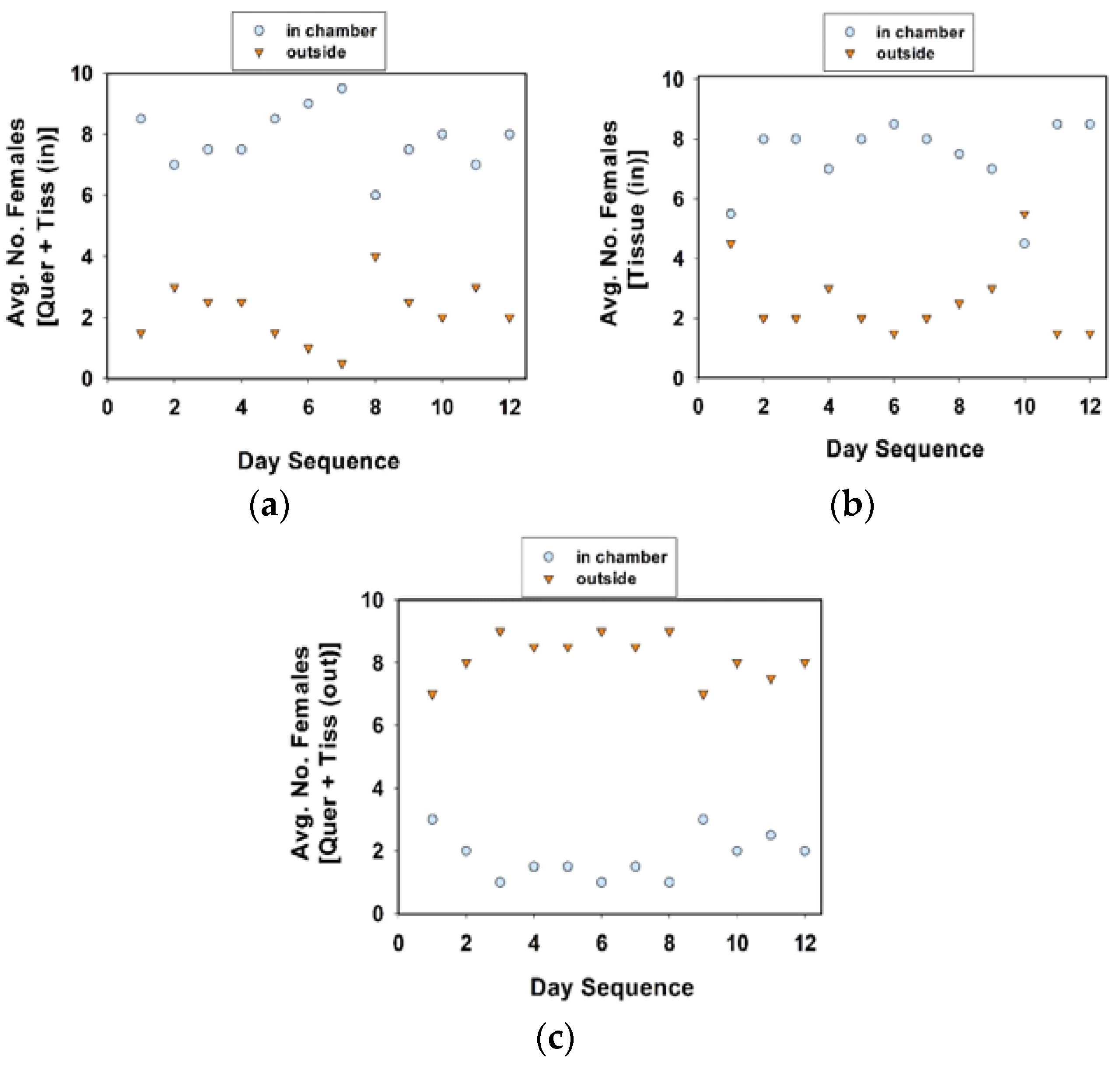
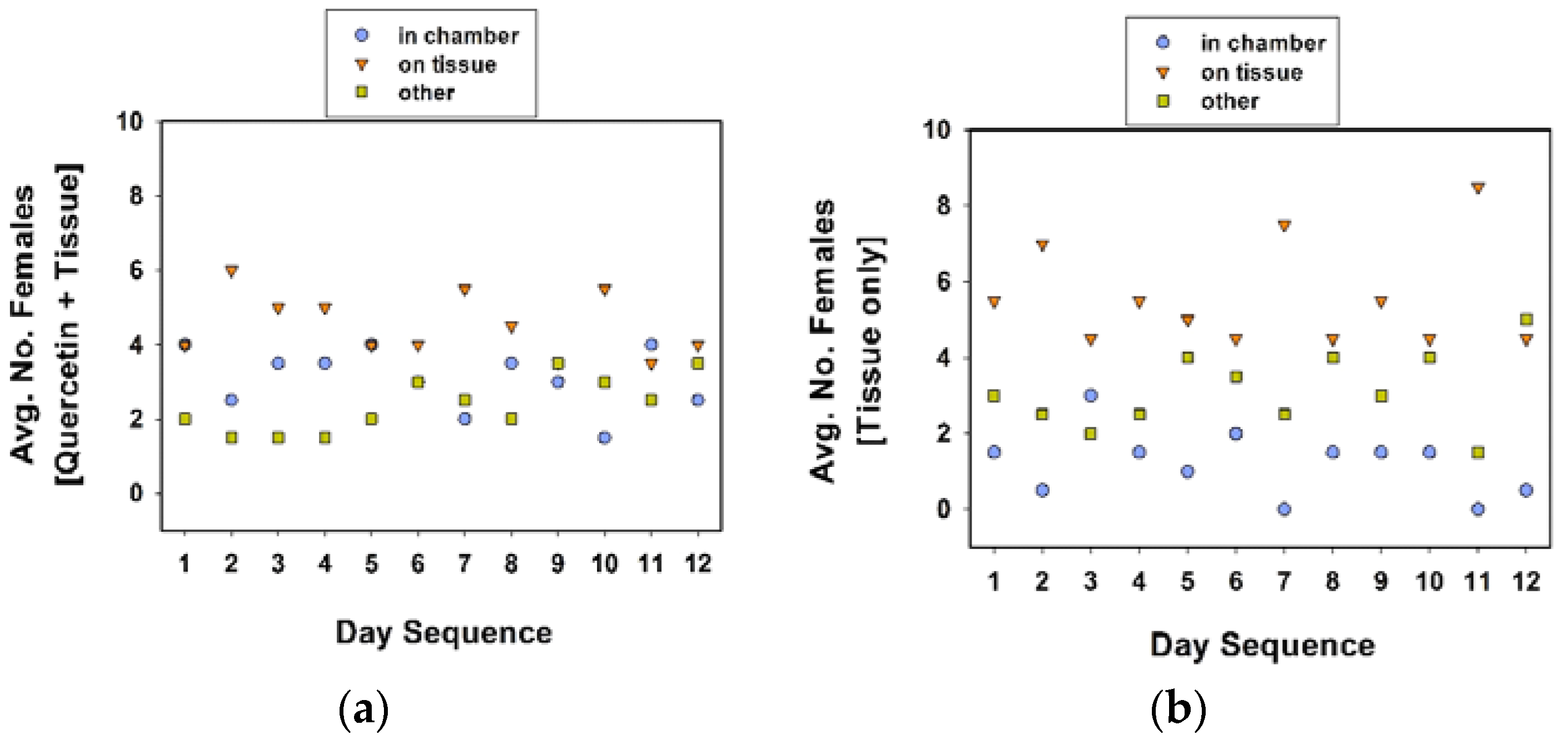
As in the previous section, the cages were maintained in a growth chamber (24 °C, 60% RH, 16 h photophase), but removed twice a day to record oviposition data. The location of females in each cage was recorded twice a day, once in the morning (at approximately 09:00 h), and in the afternoon (at approximately 16:00 h). For the ease of recording, and reduce the disturbance to females while checking the locations of the 10 females in each treatment cage, we designated distinct locations. In experiment 4, distinct locations were (1) inside the chamber, (2) on tissue substrate, and (3) others, combined. In experiment 5, the distinct locations within the cage were (1) inside the chamber, and (2) outside the chamber.
Adult females were fed every other day, using E. kuehniella eggs, as used in the stock colony. Old food and waste (fecal material) were discarded every other day.
2.5. Statistical Analysis
Data were analyzed following a completely randomized design. A two-way analysis of variance (ANOVA), using the Standard Least Squares procedure with F statistic, tested the effects of treatment and clutch site preferences of females in solitary and communal cage bioassays. A Student’s t-test and 1-way ANOVA were used to test for significance of treatments on egg clutch number (and egg number per clutch), irrespective of clutch site. A two-way ANOVA was also used to test for significance of treatments on female location in communal cages. Treatment means were significantly different following the t-test or ANOVA, if p < 0.05. When necessary, a Tukey’s (Tukey-Kramer HSD) test was used as a multiple comparison procedure after the ANOVA. JMP® 12.0.1 (2012, SAS Institute Inc., in Cary, NC, USA) and SigmaStat 3.0.1 interfaced through SigmaPlot 12 (Systat Software Inc., in Richmond, CA, USA) software assisted with the analysis of data.
4. Discussion
In cost-effective mass rearing systems, which are designed to increase the production of progeny for augmentative biological control, oftentimes natural prey are not used [
1,
2]. Aphids are the preferred prey for many ladybirds. But, aphids are difficult or impossible to rear in a cost-effective manner, because live plants (hosts) must be supplied as a food source. Production of plants for aphids is labor intensive and not practical for long-term production of ladybird beetles. Consequently, we have been experimenting with alternative food sources in lieu of aphids in our ladybird production system. A disadvantage of using non-aphid foods is that chemical cues that stimulate oviposition are often not present. As a result, chemical stimulants are necessary to boost the oviposition of ladybirds fed alternative foods or artificial diets. Natural compounds or molecules from a plant source rather than animal source (i.e., aphid tissues or aphid by-products, e.g., honeydew) would be ideal. This leads to our rationale for conducting this research. We have been testing various bioflavonoids as stimulants for oviposition in ladybirds.
In a recent investigation, it was discovered for the first time that three bioflavonoids, quercetin, taxifolin, and naringenin (to a lesser extent), in bioassays in 250 mL cages, stimulated oviposition by
C.
maculata [
13]. We have expanded our knowledge of the relationship between quercetin and this ladybird in this study. Our observation that females preferentially oviposited in a chemical dish containing quercetin, at the base of 500 mL solitary cages (housing one female), rather than on the cage walls or on tissue paper, confirms the attraction to and the stimulatory effect of this compound on females in a larger cage. Notably, in the presence of a tissue substrate, which occupied a much larger surface area than the chemical dish, females showed a preference for ovipositing in the chemical dish, suggesting that quercetin was more attractive than tissue paper.
The observation that quercetin occasionally increased the number of egg clutches per female in 500 mL solitary cages was expected on the basis of previous research. For example, quercetin and naringenin stimulated slightly more egg clutches per female in 250 mL cages, than in the control cages—lacking any bioflavonoid [
13]. The mechanism of action that triggers the physiological response in females to oviposit in close proximity to quercetin and to occasionally produce more egg clutches is unknown. Although not tested, doubling the cage size did not apparently affect the clutch number (0.82 clutches per female per day in 250 mL cage [
13]; 0.80–0.92 clutches per female per day in 500 mL cage, in this study).
Quercetin does not harm
C.
maculata females. In fact, we have observed females “tasting” small amounts of quercetin powder, suggesting that taste chemoreceptors on the mouthparts recognize this compound as being suitable for consumption. Undigested fragments of quercetin powder have been found in their faeces (ZW, unpublished observations). The nutritional value of quercetin powder to female reproduction is unknown. Whether females must taste, touch, or digest quercetin (or any other bioflavonoids) to stimulate oviposition or boost egg production needs further study. Quercetin powder often adheres to the surface of freshly-laid
C.
maculata eggs, but it does not reduce the hatch rate [
13]. Although not being tested in a rigorous experiment, hatching
C. maculata first instars were not affected negatively when contacting quercetin powder.
Research on the effects of quercetin on larvae of an herbivorous ladybird
Epilachnia paenulata (Germar), which feeds on curcubit leaves, proved that this compound had little or no effect of leaf consumption and the larval weight at concentrations from 0.1 to 50 μg/cm
2 in laboratory bioassays [
21]. Quercetin functions as a phagostimulant at low concentrations, but a feeding deterrent at high concentrations for
E.
paenulata.
Although quercetin can incite females to increase clutch number, females never produced more than two clutches in a 24 h time frame. Quercetin never incited females to increase egg number per clutch in this study. In fact, neither quercetin nor taxifolin increased egg number per clutch in solitary cages in a previous investigation [
13]. The simplest explanation for this observation is that females are limited in the number of eggs that they can produce over a given time period, because of the rate at which eggs mature in their ovaries. Other researchers have found that egg maturation rate was more dependent on a physiological response to food quality, ambient temperature, and ovariole size in ladybirds [
22].
To facilitate cost-effective and space-efficient mass rearing, grouping ovipositing adults in the same cage would be ideal. Commercial scale rearing could involve placing hundreds of ladybird adults (males and females) in the same cage, provisioned with an oviposition stimulant (such as quercetin) and some type of oviposition substrate. Quercetin would encourage females to oviposit in oviposition chambers, placed in a restricted region in these cages, and to facilitate rapid harvesting of egg clutches each day and limiting egg cannibalism, by females and/or males. Thus, our attempt to foster a situation where females were restricted to ovipositing in the company of conspecifics, yielded interesting results. Our observation that females preferred to oviposit in the chemical dishes inside a novel oviposition chamber in 1 L communal cages containing quercetin powder, in the presence or absence of a tissue substrate, further confirms the capacity of this compound to stimulate oviposition. We used tissue paper as an oviposition substrate in our rearing system (stock colony) for this ladybird beetle, indicating that females had exposure to this substrate type prior to placement in experiments. Tissue paper, filter paper, and paper hand toweling are often deployed as oviposition substrates for aphidophagous ladybird beetles in the laboratory [
6,
7,
23]. Nevertheless, quercetin appears to be a stronger oviposition stimulant than tissue paper, based on our cage bioassays.
The two- to three-fold decrease in egg clutch number per female per day in communal cages, in comparison to number per female per day in solitary cages (compare
Table 2 and
Table 4), suggests some female-female interference in the communal cages. Frequent bodily contact between females might discourage oviposition. Reluctance to oviposit in response to crowding of adults in a container, rather than lack of suitable prey (quality or quantity) has rarely been reported for adult ladybirds [
6]. Adults of the ladybird
Propylea dissecta (Mulsant) reduce their fecundity when female density (and the density of chemical tracks deposited by females) increases in small arenas, Petri dishes [
24]. Perhaps, females decide to oviposit, or not, depending on a certain threshold concentration of chemical(s) released by conspecifics. Chemical cues could signal the threat of cannibalism. Nonetheless, we did not detect any cannibalism of eggs by adult females in the cages in our experiments.
The observation that the number of egg clutches per female was slightly greater in the communal cages with quercetin plus tissue (in the chamber) suggests that the presence of quercetin and tissue in close proximity occasionally triggers a stronger ovipositional response than either quercetin or tissue alone. We observed females “resting” on tissue whether it was inside or outside of the oviposition chamber, suggesting that they are attracted to the tissue because of the presence of chemical cues that are emanating from the paper or because the tissue provides a more secure grip for their tarsi than the plastic surfaces of the cage. The placement of tissue in close proximity to quercetin in rearing cages (in a mass rearing operation) could be the best strategy to stimulate more localized oviposition and perhaps more egg production by C. maculata females.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}