

High-Humidity Incubation Improves Post-Microinjection Viability and Larval Performance of CRISPR/Cas9-Edited Helicoverpa armigera Embryos



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. sgRNA Design and Synthesis
2.3. Microinjection of Embryos
2.4. Measurement of Hatching Rate and 100-Egg Weight Under Experimental Conditions
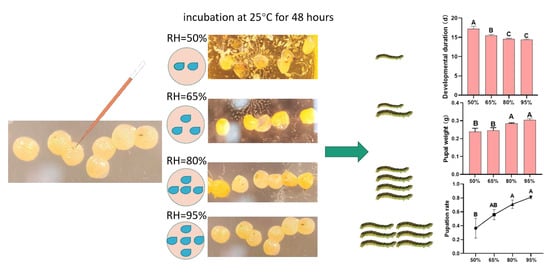
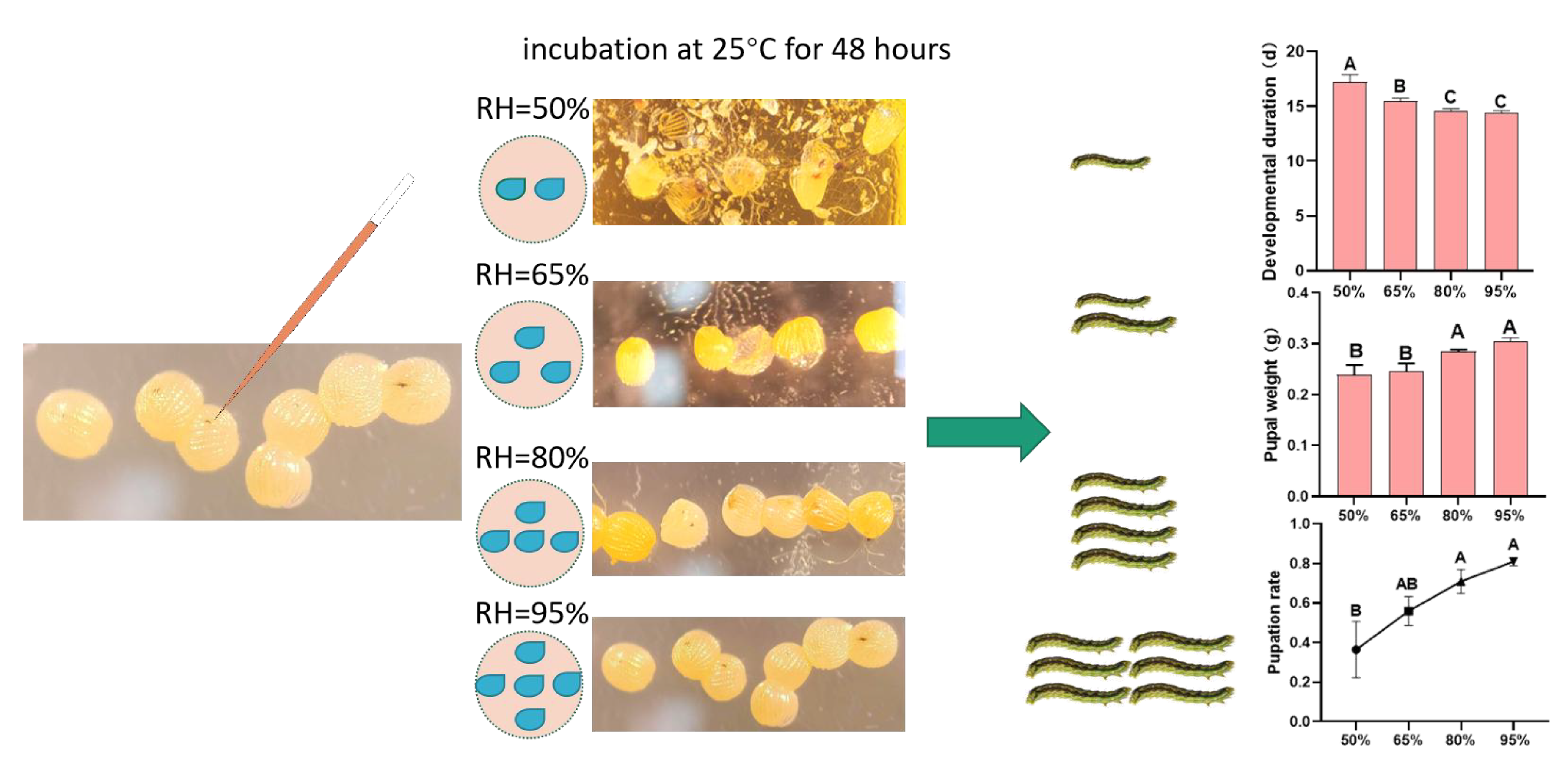
2.5. Observation and Measurement of Developmental Duration, Pupal Weight, and Pupation Rate
2.6. Determination of Gene Editing Efficiency in Hatched Larvae
2.7. Data Analysis
3. Results
3.1. Hatching Rate and 100-Egg Weight
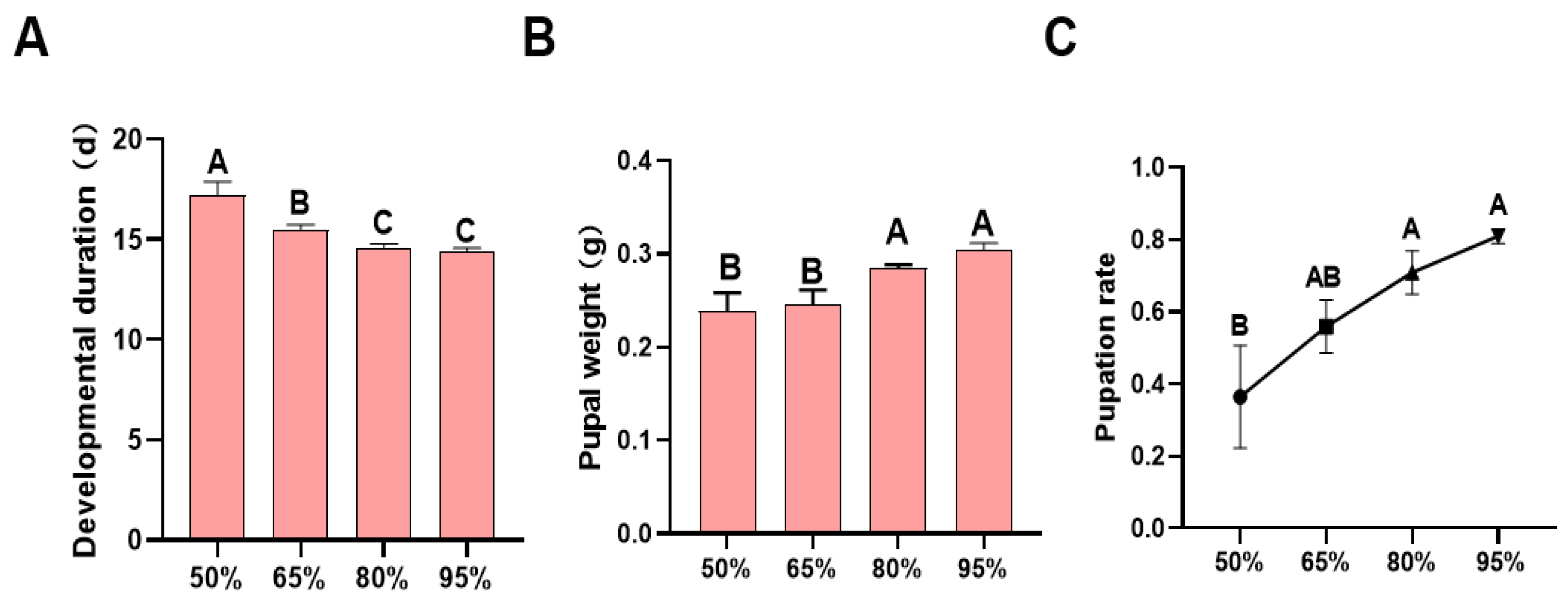
3.2. Developmental Duration, Pupal Weight, and Pupation Rate
3.3. Gene Editing Efficiency in Hatched Larvae
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- William, R.; O’Brochta, D.A. Applications of genome editing in insects. Curr. Opin. Insect Sci. 2016, 13, 43–54. [Google Scholar] [CrossRef]
- Gantz, V.M.; Akbari, O.S. Gene editing technologies and applications for insects. Curr. Opin. Insect Sci. 2018, 28, 66–72. [Google Scholar] [CrossRef]
- Taning, C.N.T.; Van Eynde, B.; Yu, N.; Ma, S.; Smagghe, G. CRISPR/Cas9 in insects: Applications, best practices and biosafety concerns. J. Insect Physiol. 2017, 98, 245–257. [Google Scholar] [CrossRef]
- Xu, J.; Xu, X.; Zhan, S.; Huang, Y. Genome editing in insects: Current status and challenges. Natl. Sci. Rev. 2019, 6, 399–401. [Google Scholar] [CrossRef]
- Zhang, L.; Reed, R.D. A Practical Guide to CRISPR/Cas9 Genome Editing in Lepidoptera. In Diversity and Evolution of Butterfly Wing Patterns; Sekimura, T., Nijhout, H., Eds.; Springer: Singapore, 2017. [Google Scholar]
- Li, X.; Fan, D.; Zhang, W.; Liu, G.; Zhang, L.; Zhao, L.; Fang, X.; Chen, L.; Dong, Y.; Chen, Y.; et al. Outbred genome sequencing and CRISPR/Cas9 gene editing in butterflies. Nat. Commun. 2015, 6, 8212. [Google Scholar] [CrossRef] [PubMed]
- Khadka, A.; Hodges, A.C.; Leppla, N.C.; Tillman, P.G. The effects of relative humidity on Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) egg hatch, nymph survival, and adult reproduction. Fla. Entomol. 2020, 103, 136–138. [Google Scholar] [CrossRef]
- Yang, W.Q.; Li, X.; Meng, F.X.; Liu, T.X.; Zhang, S.Z. Effects of temperature, relative humidity, and photoperiod on life history of Cotesia ruficrus (Hymenoptera: Braconidae), an indigenous parasitoid of Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Econ. Entomol. 2025, 118, 1133–1145. [Google Scholar] [CrossRef] [PubMed]
- Aslam, A.; Jafir, M.; Wajid Javed, M.; Shehzad, M.; Zubair Chaudhary, M.; Aftab, M.; Muhammad Jafir, C. Effect of temperature and relative humidity on development of Sitophilus oryzae L. (coleoptera: Curculionidae). J. Entomol. Zool. Stud. 2017, 5, 85–90. [Google Scholar]
- Ji, Y.T.; Xue, C.Z.; Wang, M.Q.; Xiang, M.; Li, P.; Li, Y.Y.; Mao, J.J.; Zhang, L.S. Effects of relative humidity on development of Picromerus lewisi Scott. Chin. J. Biol. Control. 2022, 38, 975–981. [Google Scholar]
- Deruytter, D.; Coudron, C.L.; Claeys, J. Transporting Tenebrio molitor Eggs: The Effect of Temperature, Humidity and Time on the Hatch Rate. Sustainability 2023, 15, 6231. [Google Scholar] [CrossRef]
- Prasad, K.V.H. Factors Affecting Growth and Metabolism of Insects. In Insect Ecology: Concepts to Management; Springer: Singapore, 2022. [Google Scholar]
- Griese, E.; Dicke, M.; Hilker, M.; Fatouros, N.E. Plant response to butterfly eggs: Inducibility, severity and success of egg-killing leaf necrosis depends on plant genotype and egg clustering. Sci. Rep. 2017, 7, 7316. [Google Scholar] [CrossRef]
- Urbanski, J.M.; Benoit, J.B.; Michaud, M.R.; Denlinger, D.L.; Armbruster, P. The molecular physiology of increased egg desiccation resistance during diapause in the invasive mosquito, Aedes albopictus. Proc. Biol. Sci. 2010, 277, 2683–2692. [Google Scholar] [CrossRef] [PubMed]
- Finch, G.; Nandyal, S.; Perretta, C.; Davies, B.; Rosendale, A.J.; Holmes, C.J.; Gantz, J.D.; Spacht, D.E.; Bailey, S.T.; Chen, X.; et al. Multi-level analysis of reproduction in an Antarctic midge identifies female and male accessory gland products that are altered by larval stress and impact progeny viability. Sci. Rep. 2020, 10, 19791. [Google Scholar] [CrossRef]
- Hilker, M.; Salem, H.; Fatouros, N.E. Adaptive plasticity of insect eggs in response to environmental challenges. Annu. Rev. Entomol. 2023, 68, 451–469. [Google Scholar] [CrossRef] [PubMed]
- Matsushima, R.; Yokoi, T. Eggs in faeces: Defensive mechanisms and effects of faecal coating by a water scavenger beetle, Coelostoma stultum (Coleopterea: Hydrophilidae). Sci. Nat. 2023, 110, 29. [Google Scholar] [CrossRef]
- Li, T.H.; Wang, X.; Desneux, N.; Wang, S.; Zang, L.S. Egg coverings in insects: Ecological adaptation to abiotic and biotic selective pressures. Biol. Rev. Camb. Philos. Soc. 2025, 100, 99–112. [Google Scholar] [CrossRef]
- Jacobs Chris, G.C.; Rezende Gustavo, L.; Lamers Gerda, E.M.; van der Zee, M. The extraembryonic serosa protects the insect egg against desiccation. Proc. Biol. Sci. 2013, 280, 20131082. [Google Scholar] [CrossRef] [PubMed]
- Vargas, H.C.; Farnesi, L.C.; Martins, A.J.; Valle, D.; Rezende, G.L. Serosal cuticle formation and distinct degrees of desiccation resistance in embryos of the mosquito vectors Aedes aegypti, Anopheles aquasalis and Culex quinquefasciatus. J. Insect Physiol. 2014, 62, 54–60. [Google Scholar] [CrossRef]
- Farnesi, L.C.; Vargas, H.C.M.; Valle, D.; Rezende, G.L. Darker eggs of mosquitoes resist more to dry conditions: Melanin enhances serosal cuticle contribution in egg resistance to desiccation in Aedes, Anopheles and Culex vectors. PLoS Negl. Trop. Dis. 2017, 11, e0006063. [Google Scholar] [CrossRef]
- Fischer, S.; De Majo, M.S.; Di Battista, C.; Campos, R.E. Effects of temperature and humidity on the survival and hatching response of diapausing and non-diapausing Aedes aegypti eggs. J. Insect Physiol. 2025, 161, 104726. [Google Scholar] [CrossRef]
- Xie, S.; Shen, B.; Zhang, C.; Huang, X.; Zhang, Y. sgRNAcas9: A software package for designing CRISPR sgRNA and evaluating potential off-target cleavage sites. PLoS ONE 2014, 9, e100448. [Google Scholar] [CrossRef]
- Jin, M.H.; Xiao, Y.T.; Cheng, Y.; Hu, J.; Xue, C.B.; Wu, K.M. Chromosomal deletions mediated by CRISPR/Cas9 in Helicoverpa armigera. Insect Sci. 2019, 26, 1029–1036. [Google Scholar] [CrossRef]
- Winston, P.W.; Bates, D.H. Saturated solutions for the control of humidity in biological research. Ecology 1960, 41, 232–237. [Google Scholar] [CrossRef]
- Fatouros, N.; Cusumano, A.; Bin, F.; Polaszek, A.; Van Lenteren, J. How to escape from insect egg parasitoids: A review of potential factors explaining parasitoid absence across the Insecta. Proc. R. Soc. B 2020, 287, 20200344. [Google Scholar] [CrossRef]
- Gillott, C. Entomology, 3rd ed.; Springer: Dordrecht, The Netherlands, 2005. [Google Scholar]
- Hans, K.R.; VanLaerhoven, S.L. Effects of relative humidity on egg hatching success of blow flies (Diptera: Calliphoridae). Can. Soc. Forensic Sci. 2024, 57, 215–223. [Google Scholar] [CrossRef]
- Mitchaothai, J.; Lertpatarakomol, R.; Trairatapiwan, T.; Lukkananukool, A. Influence of Incubation Temperature and Relative Humidity on the Egg Hatchability Pattern of Two-Spotted (Gryllus bimaculatus) and House (Acheta domesticus) Crickets. Animals 2024, 14, 2176. [Google Scholar] [CrossRef]
- Tang, J.; Cheng, Y.; Sappington, T.W.; Jiang, X.; Zhang, L.; Luo, L. Egg Hatch and Survival and Development of Beet Webworm (Lepidoptera: Crambidae) Larvae at Different Combinations of Temperature and Relative Humidity. J. Econ. Entomol. 2016, 109, 1603–1611. [Google Scholar] [CrossRef]
- Wiggleswort, V.B. The Principles of Insect Physiology, 7th ed.; Chapman and Hall: London, UK, 1972. [Google Scholar]
- Levi-Mourao, A.; Madeira, F.; Meseguer, R.; García, A.; Pons, X. Effects of Temperature and Relative Humidity on the Embryonic Development of Hypera postica Gyllenhal (Col.: Curculionidae). Insects 2021, 12, 250. [Google Scholar] [CrossRef]
- Fisher, J.J.; Rijal, J.P.; Zalom, F.G. Temperature and humidity interact to influence brown marmorated stink bug (Hemiptera: Pentatomidae), survival. Environ. Entomol. 2021, 50, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Ofuya, T.I.; Reichmuth, C. Effect of relative humidity on the susceptibility of Callosobruchus maculatus (Fabricius) (Coleoptera: Bruchidae) to two modified atmospheres. J. Stored Prod. Res. 2001, 38, 139–146. [Google Scholar] [CrossRef]
- Vargas, H.C.M.; Panfilio, K.A.; Roelofs, D.; Rezende, G.L. Increase in egg resistance to desiccation in springtails correlates with blastodermal cuticle formation: Eco-evolutionary implications for insect terrestrialization. J. Exp. Zool. B Mol. Dev. Evol. 2021, 336, 606–619. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Bui, M.; Akbari, O.S. Embryo Microinjection and Transplantation Technique for Nasonia vitripennis Genome Manipulation. J. Vis. Exp. 2017, 130, 56990. [Google Scholar]
- Salum, Y.M.; Yin, A.; Zaheer, U.; Liu, Y.; Guo, Y.; He, W. CRISPR/Cas9-Based Genome Editing of Fall Armyworm (Spodoptera frugiperda): Progress and Prospects. Biomolecules 2024, 14, 1074. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer ID | Sequence |
|---|---|
| sgRNA-TRPA1 | GGTGCCACAGCGCACGGTCCG |
| Primer ID | Sequence |
|---|---|
| D-TRPA1-F | ATGGGTGCATCGCTAGAG |
| D-TRPA1-R | AGGAAGCGAGGACTTGCG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, J.; Li, H.; Peng, Y.; Jin, M.; Mao, K.; Xiao, Y. High-Humidity Incubation Improves Post-Microinjection Viability and Larval Performance of CRISPR/Cas9-Edited Helicoverpa armigera Embryos. Insects 2025, 16, 1257. https://doi.org/10.3390/insects16121257
Zhu J, Li H, Peng Y, Jin M, Mao K, Xiao Y. High-Humidity Incubation Improves Post-Microinjection Viability and Larval Performance of CRISPR/Cas9-Edited Helicoverpa armigera Embryos. Insects. 2025; 16(12):1257. https://doi.org/10.3390/insects16121257
Chicago/Turabian StyleZhu, Jingyun, Hongran Li, Yan Peng, Minghui Jin, Kaikai Mao, and Yutao Xiao. 2025. "High-Humidity Incubation Improves Post-Microinjection Viability and Larval Performance of CRISPR/Cas9-Edited Helicoverpa armigera Embryos" Insects 16, no. 12: 1257. https://doi.org/10.3390/insects16121257
APA StyleZhu, J., Li, H., Peng, Y., Jin, M., Mao, K., & Xiao, Y. (2025). High-Humidity Incubation Improves Post-Microinjection Viability and Larval Performance of CRISPR/Cas9-Edited Helicoverpa armigera Embryos. Insects, 16(12), 1257. https://doi.org/10.3390/insects16121257