
The Morphological Diversity of Dragon Lacewing Larvae (Nevrorthidae, Neuroptera) Changed More over Geological Time Scales Than Anticipated


 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Documentation Methods
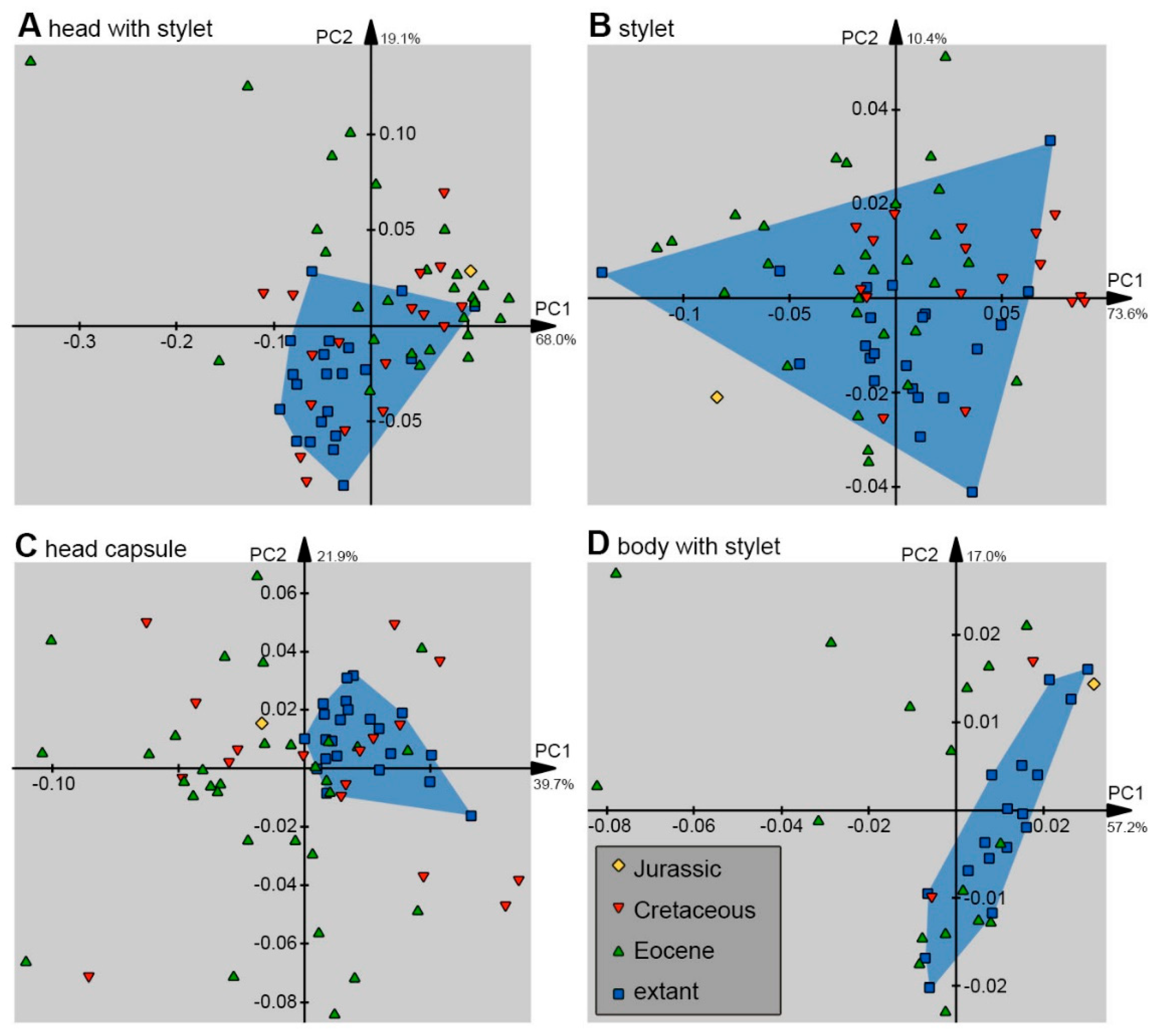
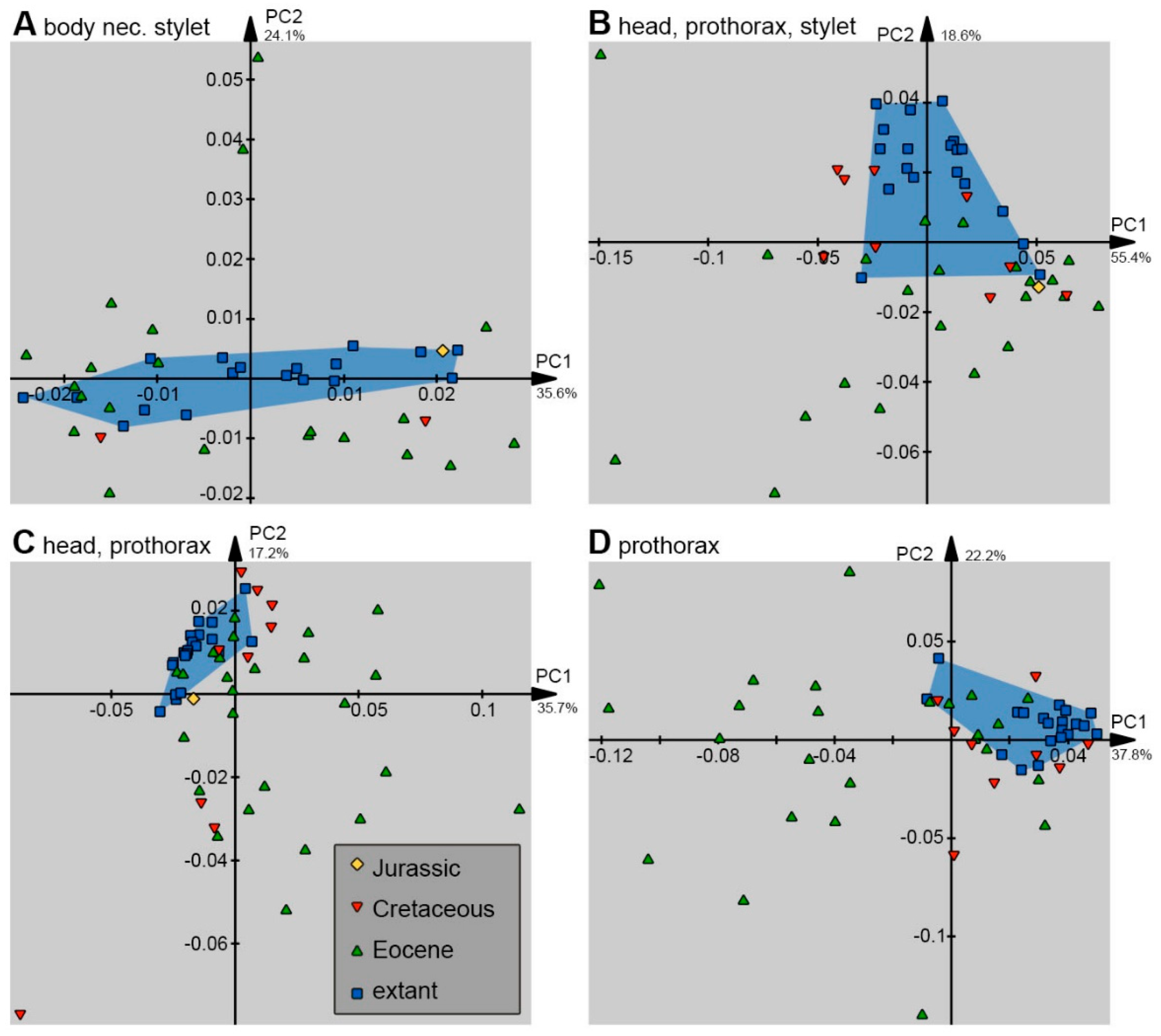
2.3. Shape Analysis
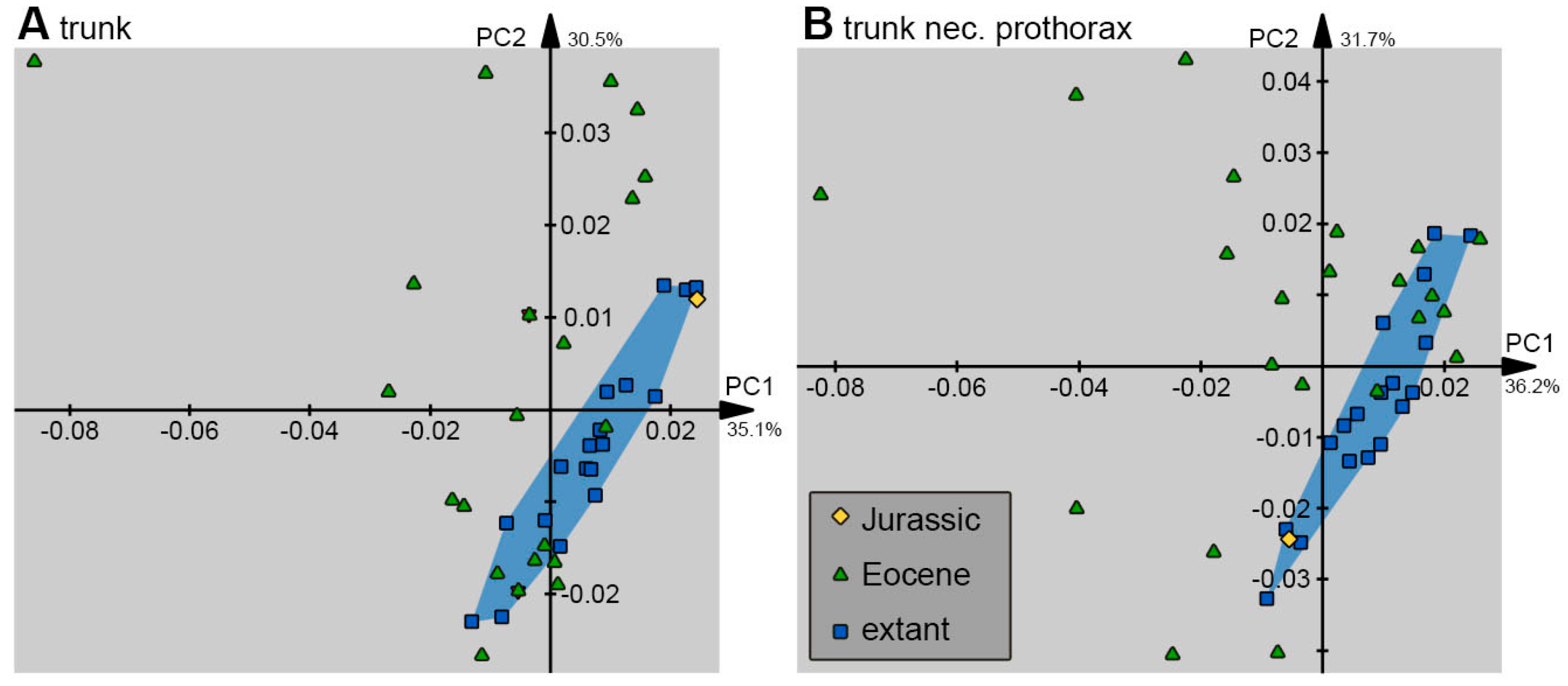
3. Results
3.1. Additional Larval Representatives of Nevrorthidae Depicted in the Literature
- (1)
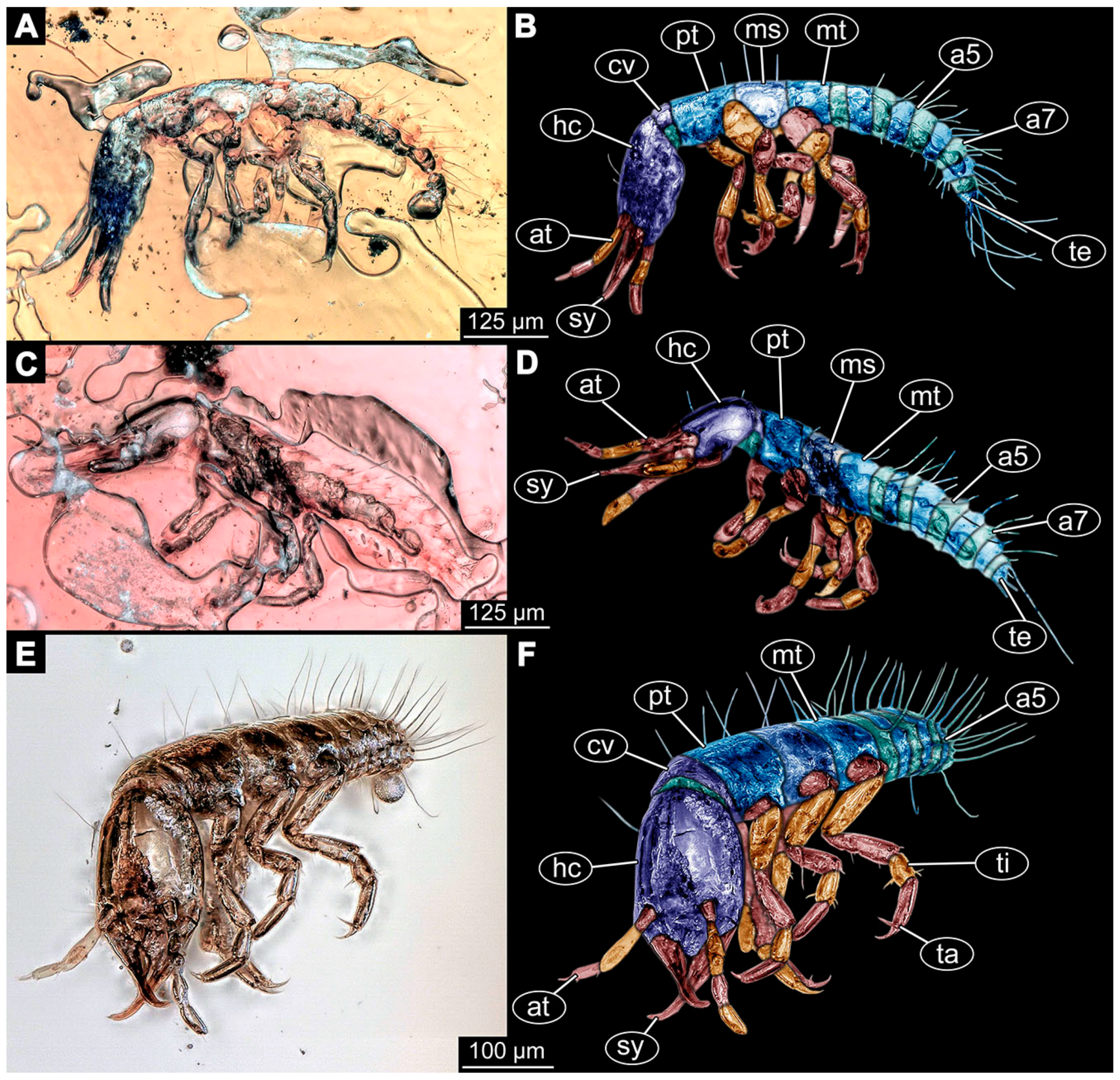
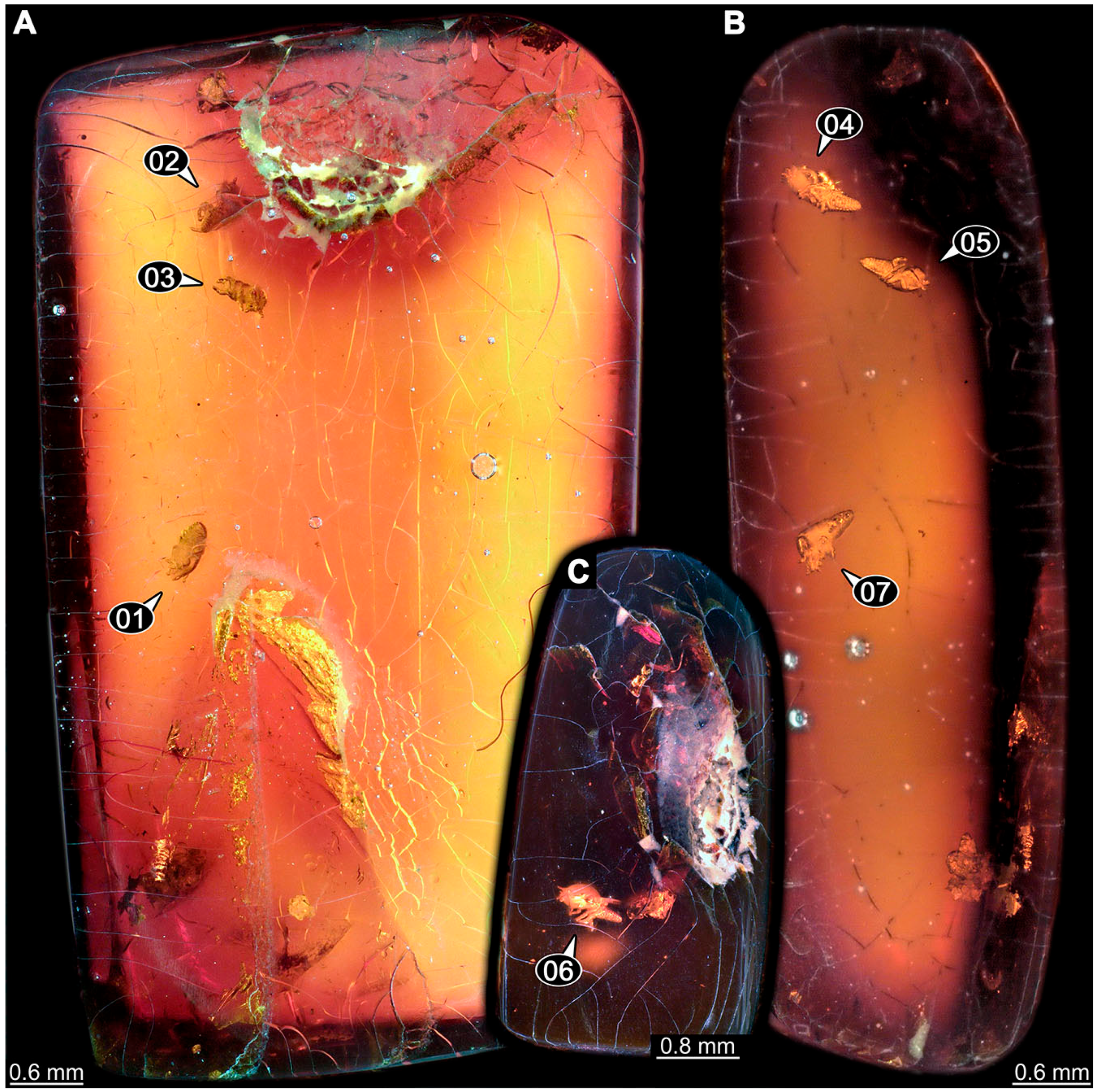
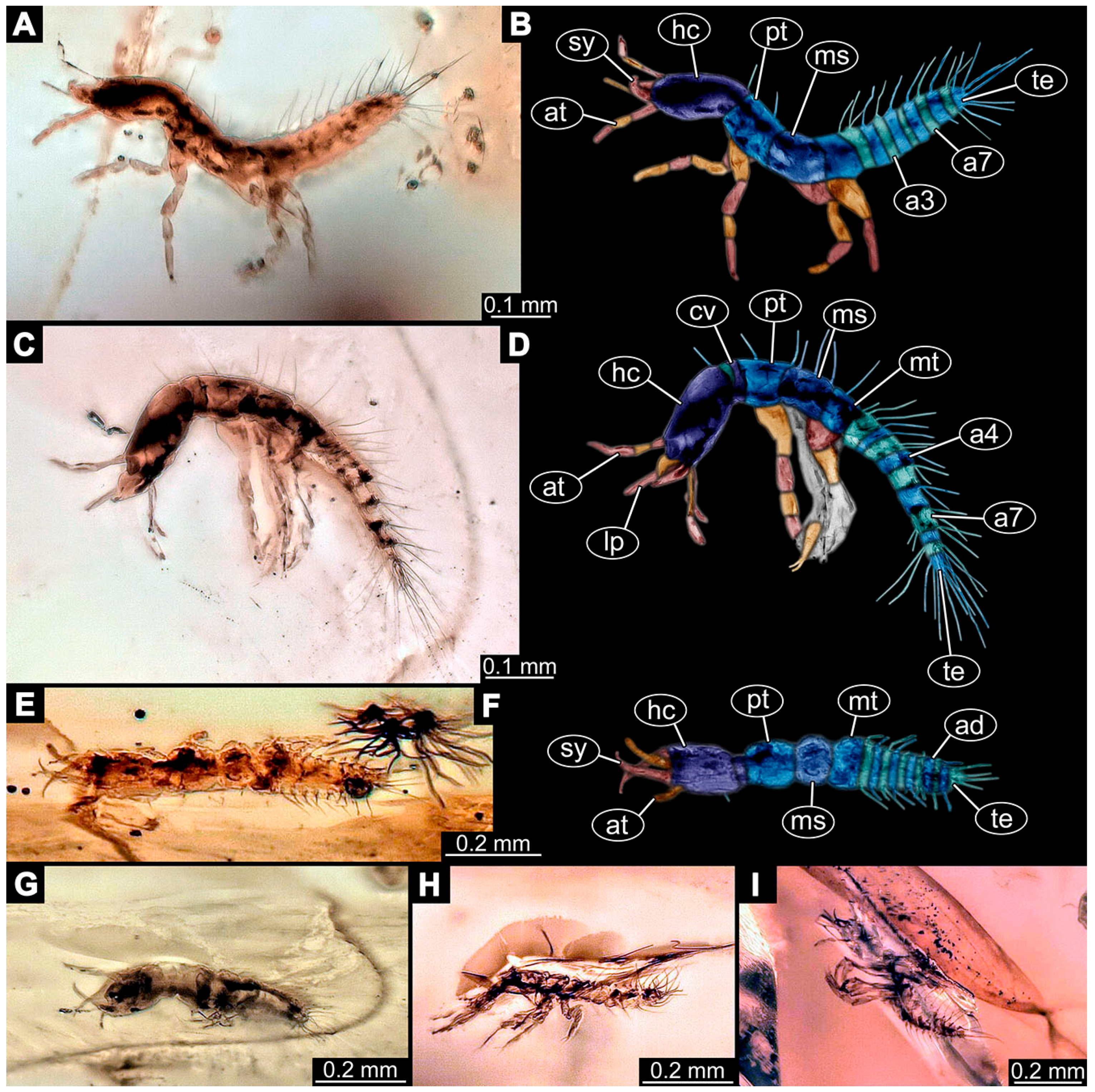
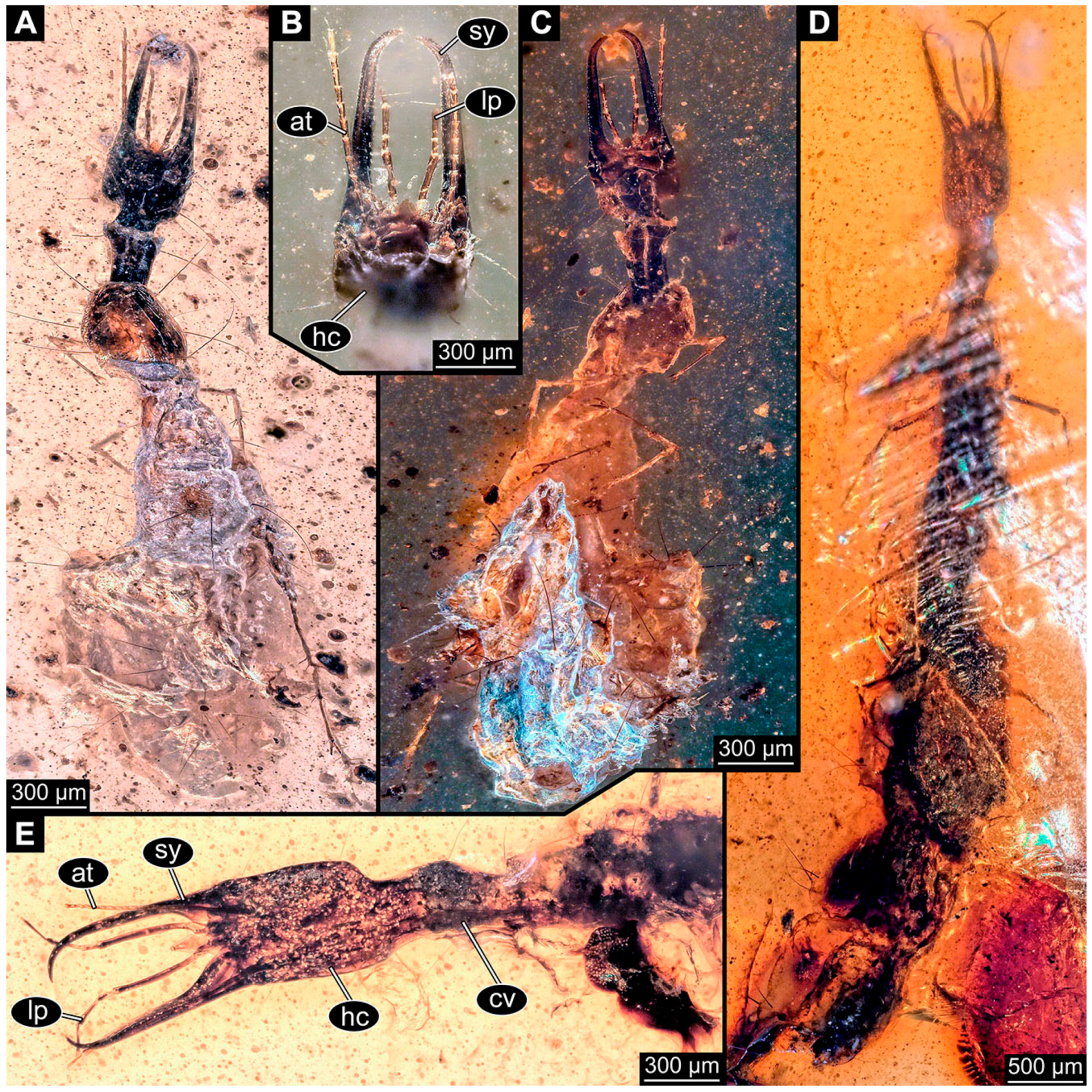
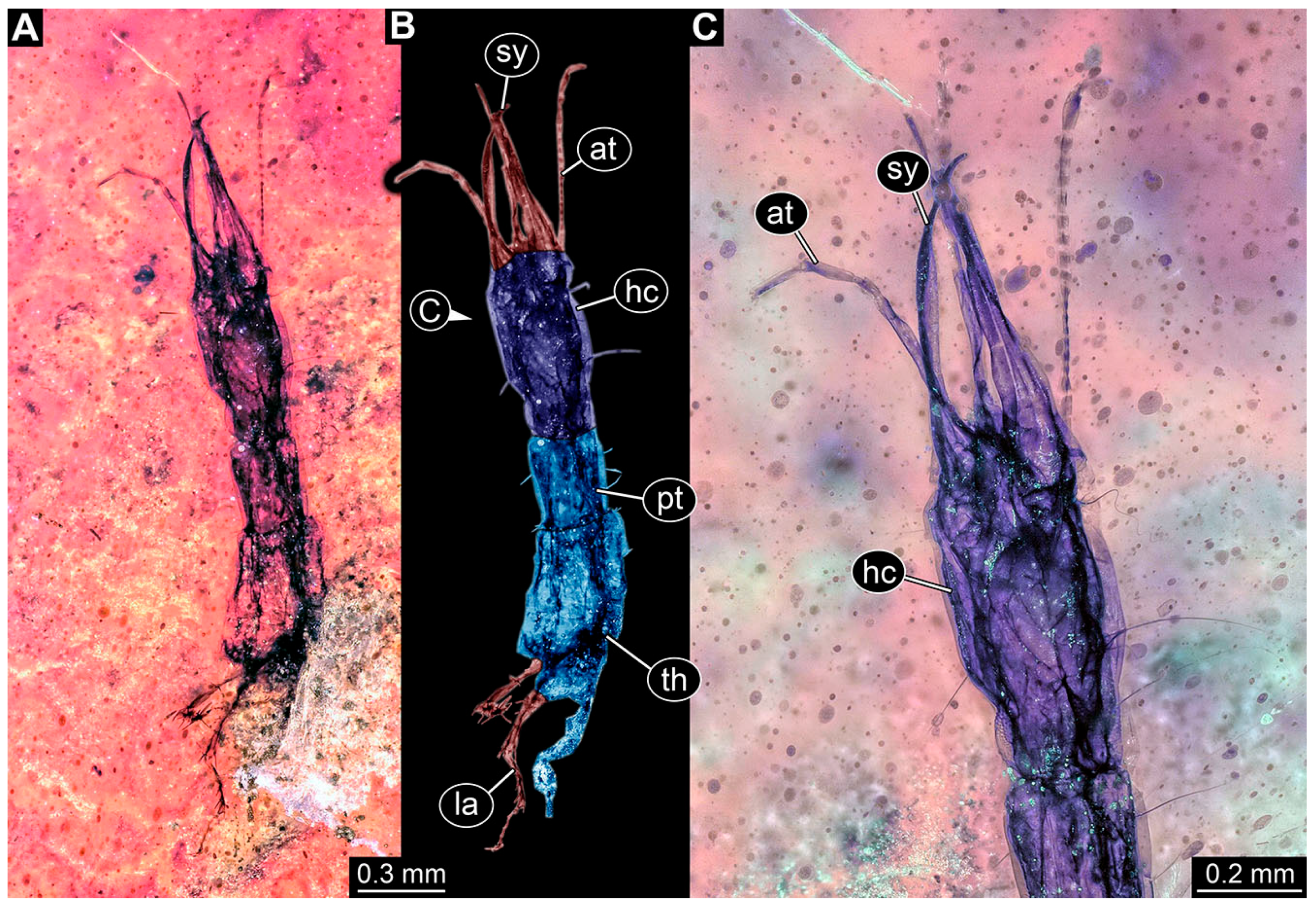
- Janzen [80] depicted a photomicrograph (fig. 261 p. 120; Figure 1A) of a dragon lacewing larva preserved in Baltic amber (specimen 6634). The image shows only the anterior region of the body in dorsal view. According to the figure legend, the specimen is part of the collection Glink. No indication of size was provided.
- (2)
- (3)
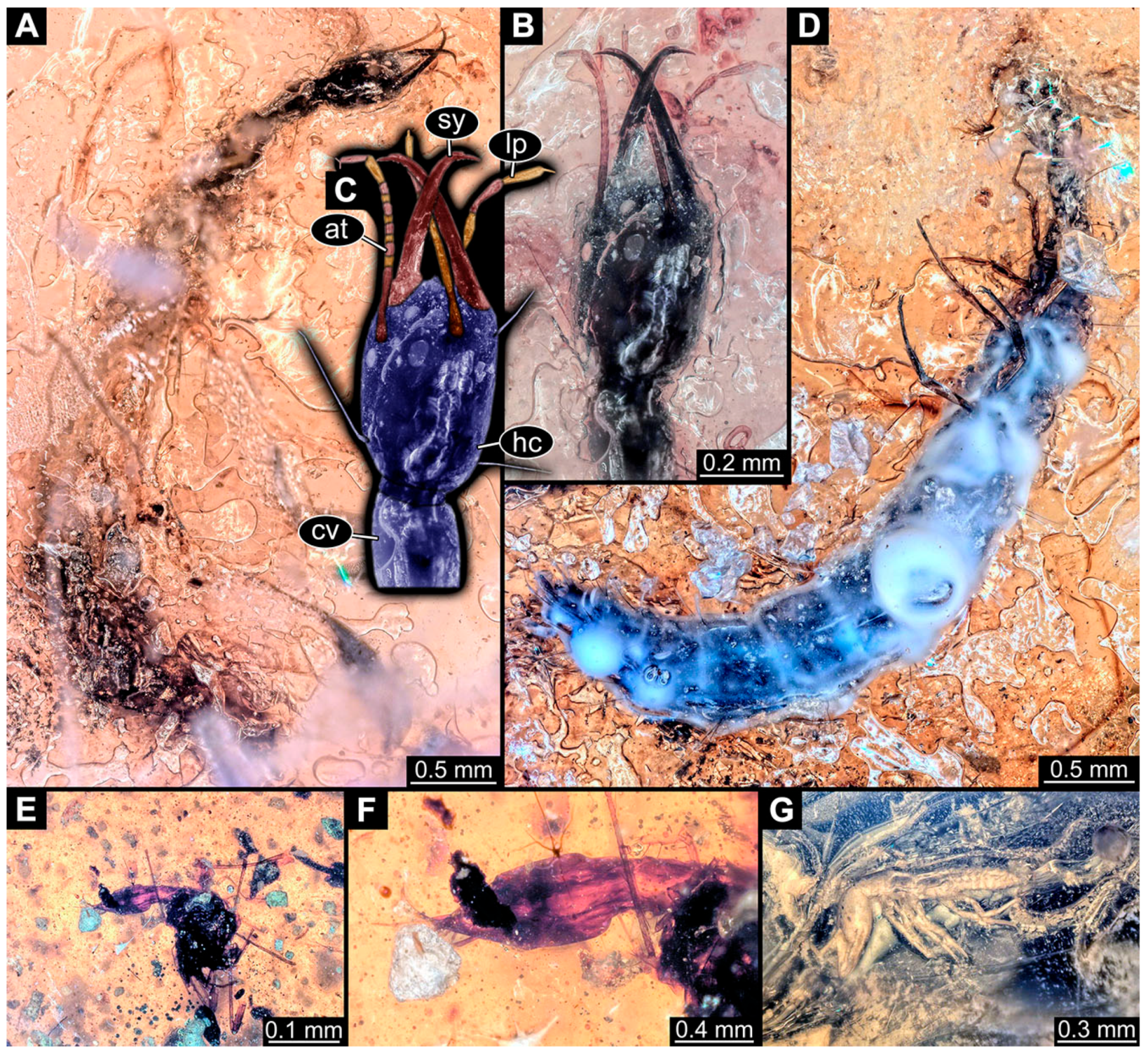
- Kobbert [82] depicted a photomicrograph (fig. T444 p. 108; Figure 1E) of a dragon lacewing larva (specimen 6638) preserved in Baltic amber, labelled as “Neurorthidae”. The image shows the animal in dorsal view. In addition, a close-up of the anterior body was provided (fig. T444 p. 109). No indication of size was provided.
- (4)
- Giacomino [83] depicted three photomicrographs (all on p. 118) of an extant larva (specimen 6639) of Nevrorthus fallax. Images include an overview in lateral view (fig. 1; Figure 1F), and the anterior head region in dorsal (fig. 2; Figure 1G) and ventral view (fig. 3). No indication of size was provided. According to the text, several specimens were available for study, but only a single one was depicted.
- (5)
- (6)
- (7)
- Du et al. [43] reported two fossil larvae of dragon lacewings (Girafficervix baii; fig. 1c, p. 3) from the Jurassic of China. The first specimen (6640) is the better preserved one (Figure 1H), almost complete in dorsal view. The second specimen (6641) is slightly less well preserved, but shows some details not available from the first specimen. Both together give a good impression of the overall morphology, which is unusual due to a long neck and an astonishing size of about 40 mm.
- (8)
- Websites are usually not considered a “good” source for scientific research. Yet, given the scarceness of appearances of larval representatives of Nevrorthidae in the literature, we decided to also include data from three websites hosted by organisations that are likely to guarantee a long-time availability. All specimens are from the extant fauna and were retrieved on 3 June 2021.
3.2. New Extant Larval Representatives of Nevrorthidae
- (9)
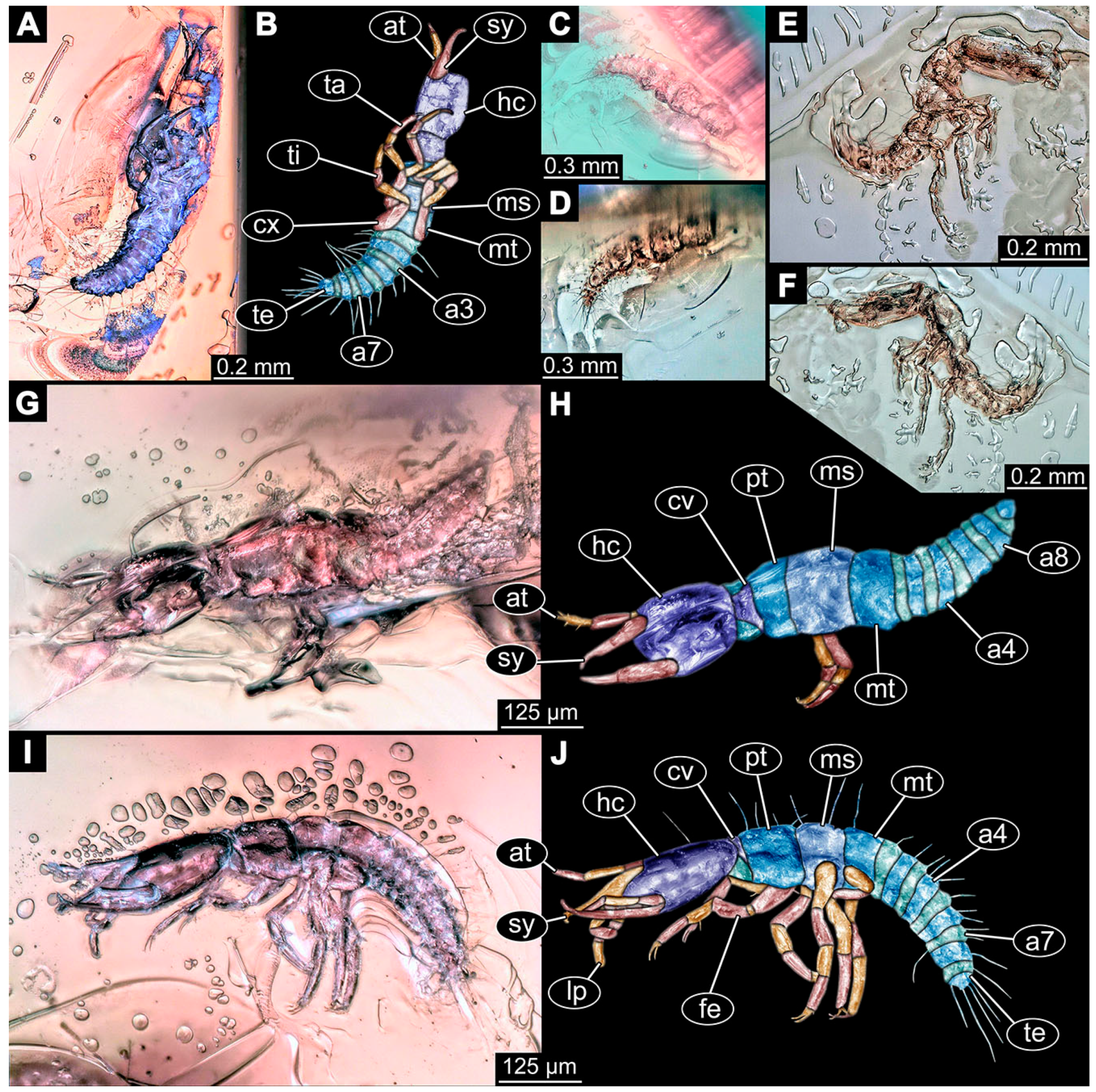
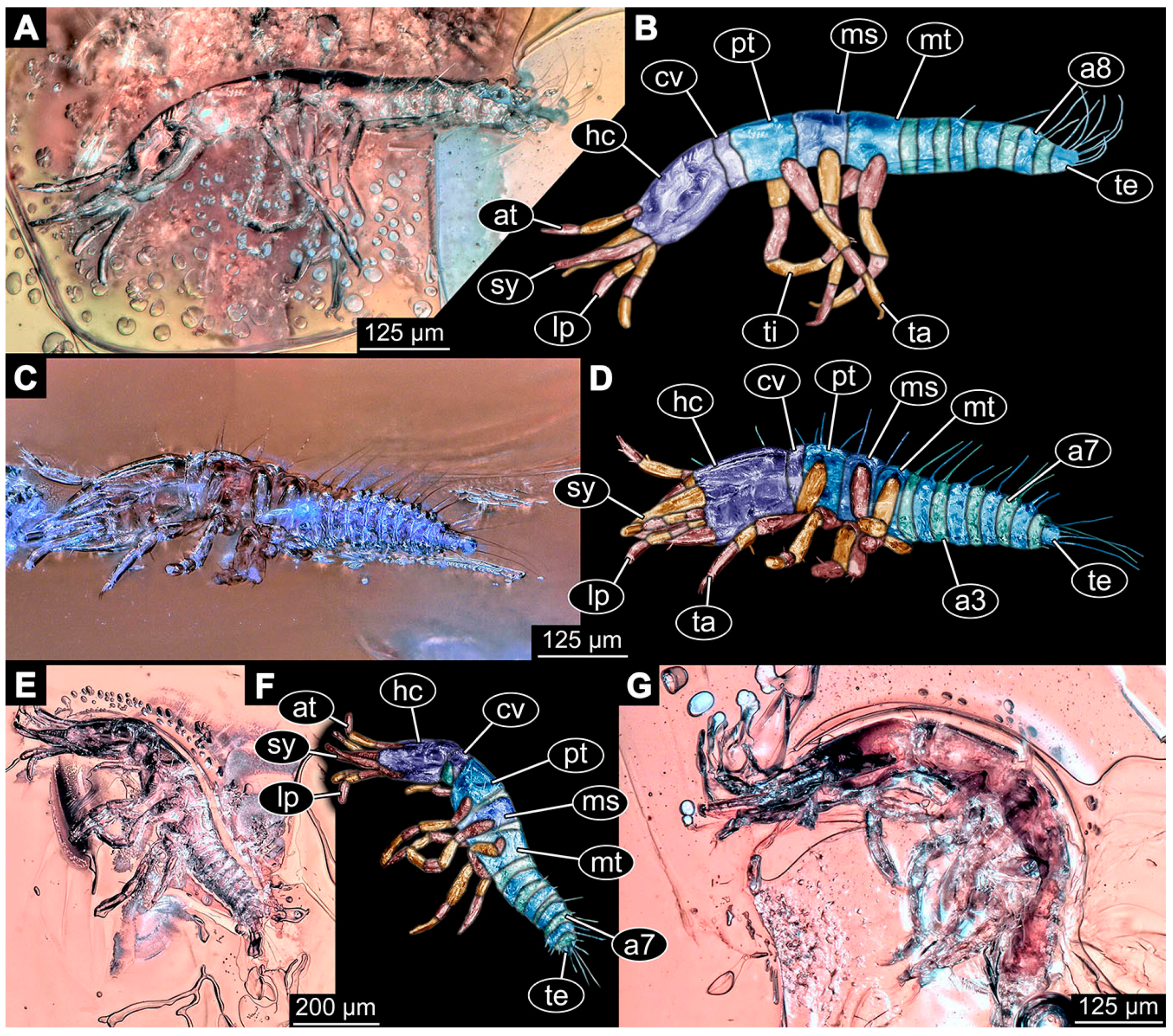
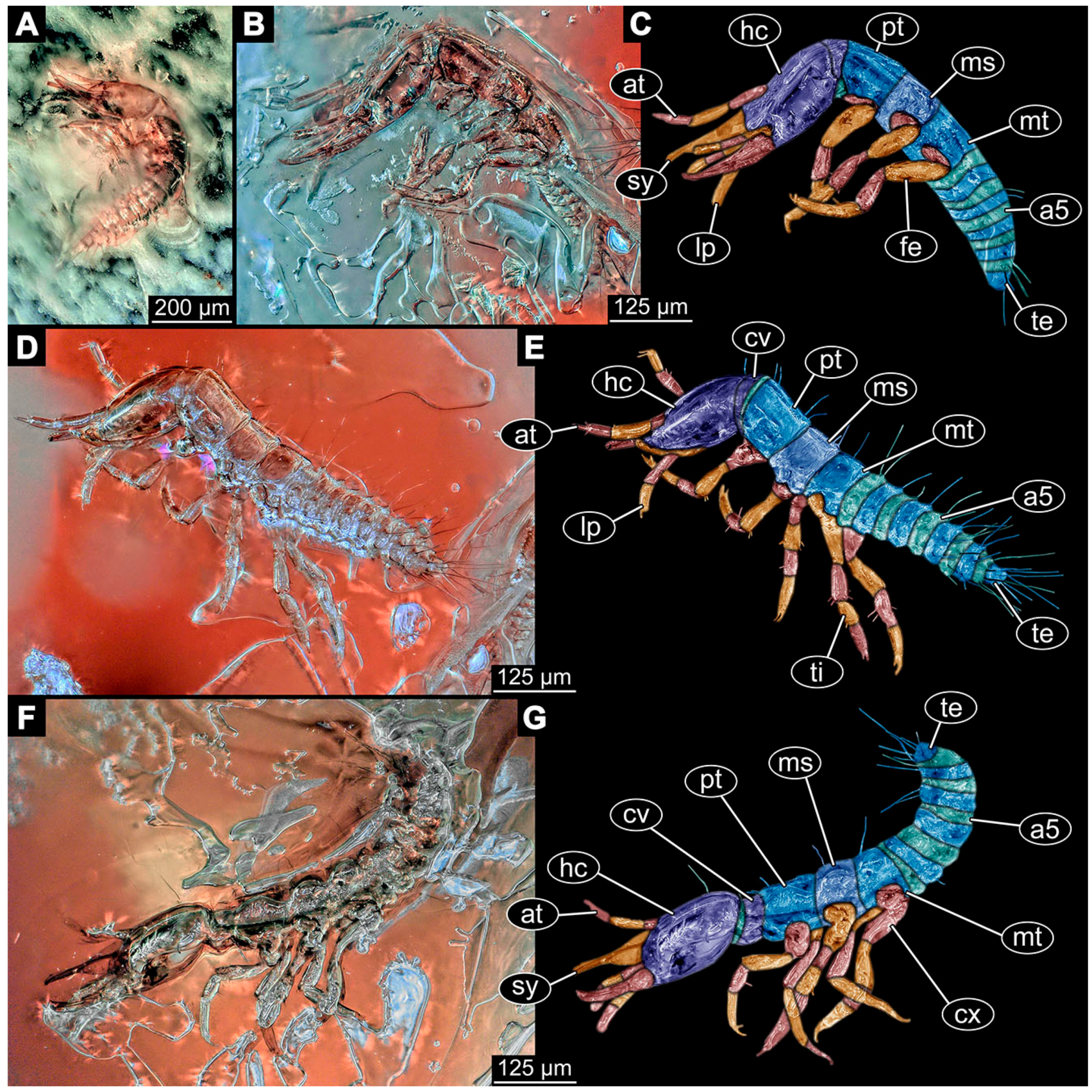
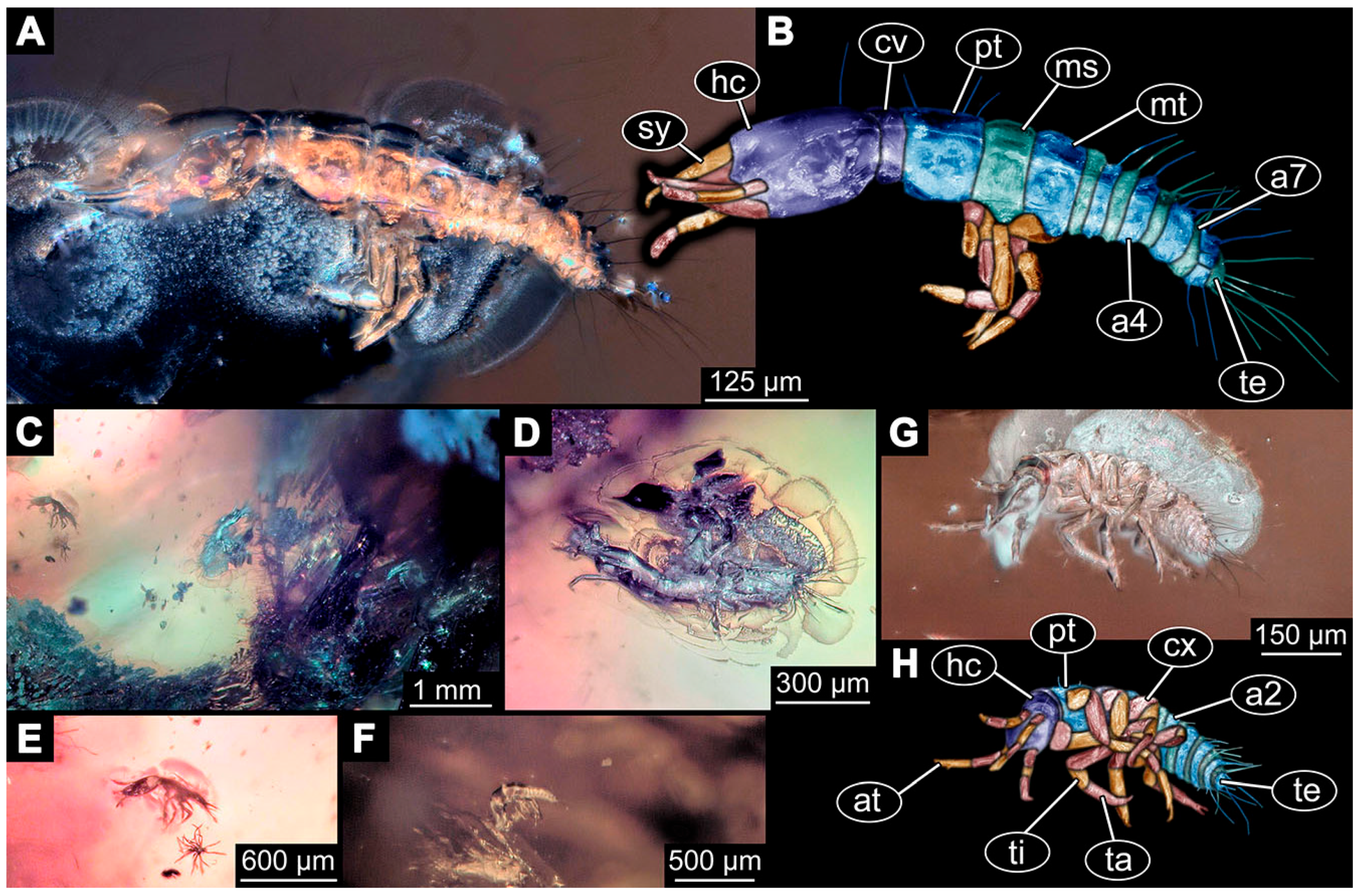
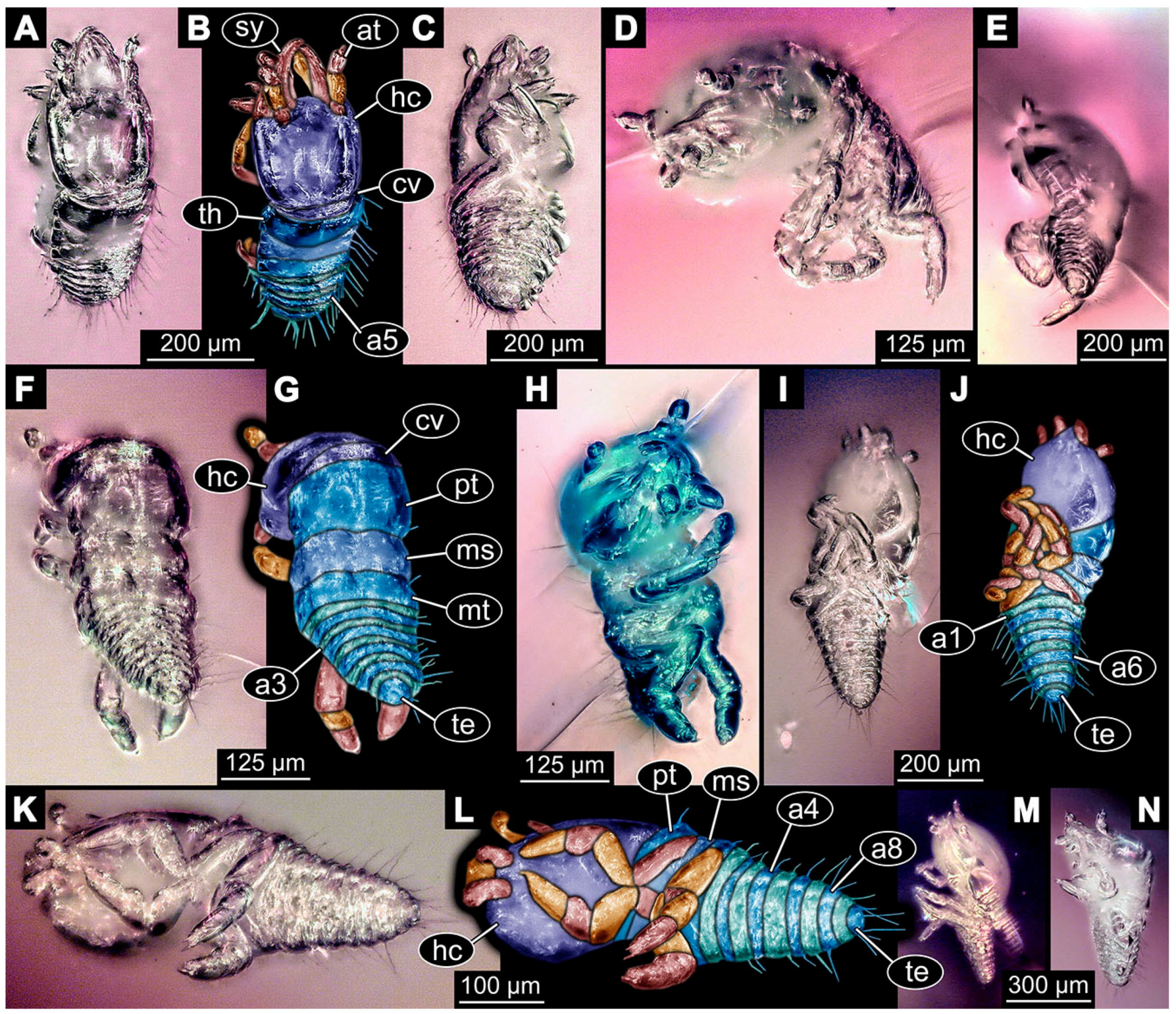
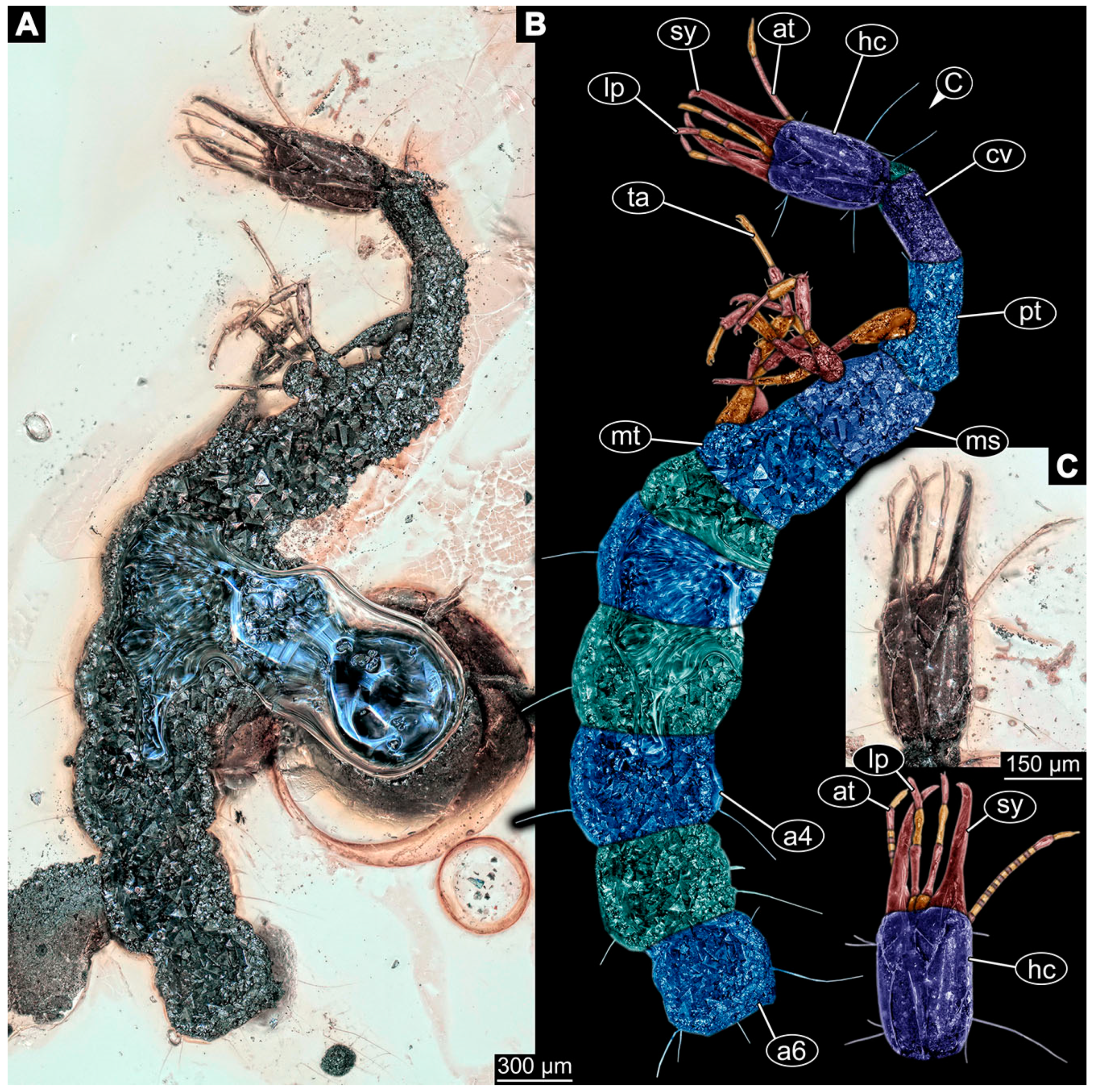
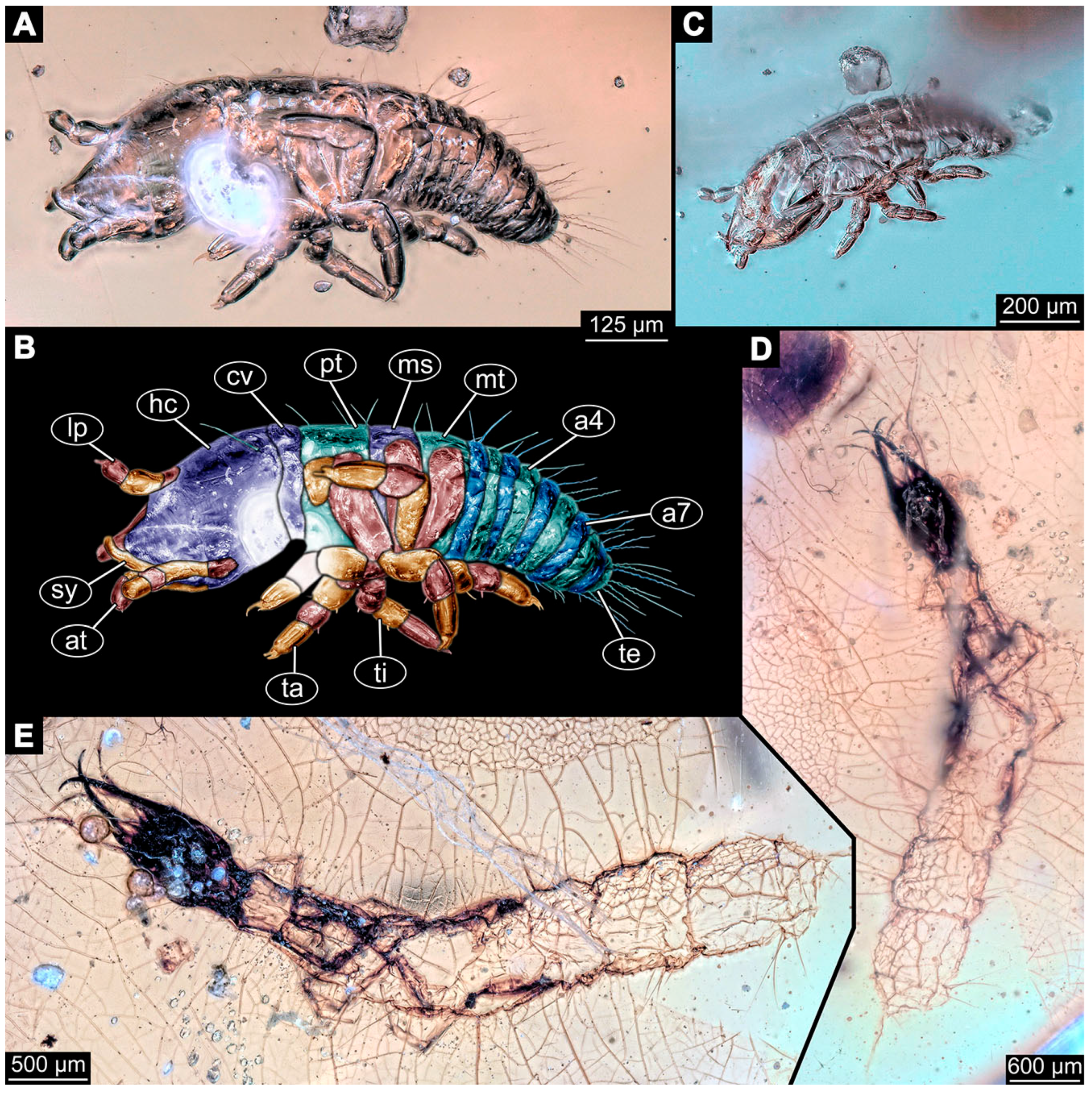
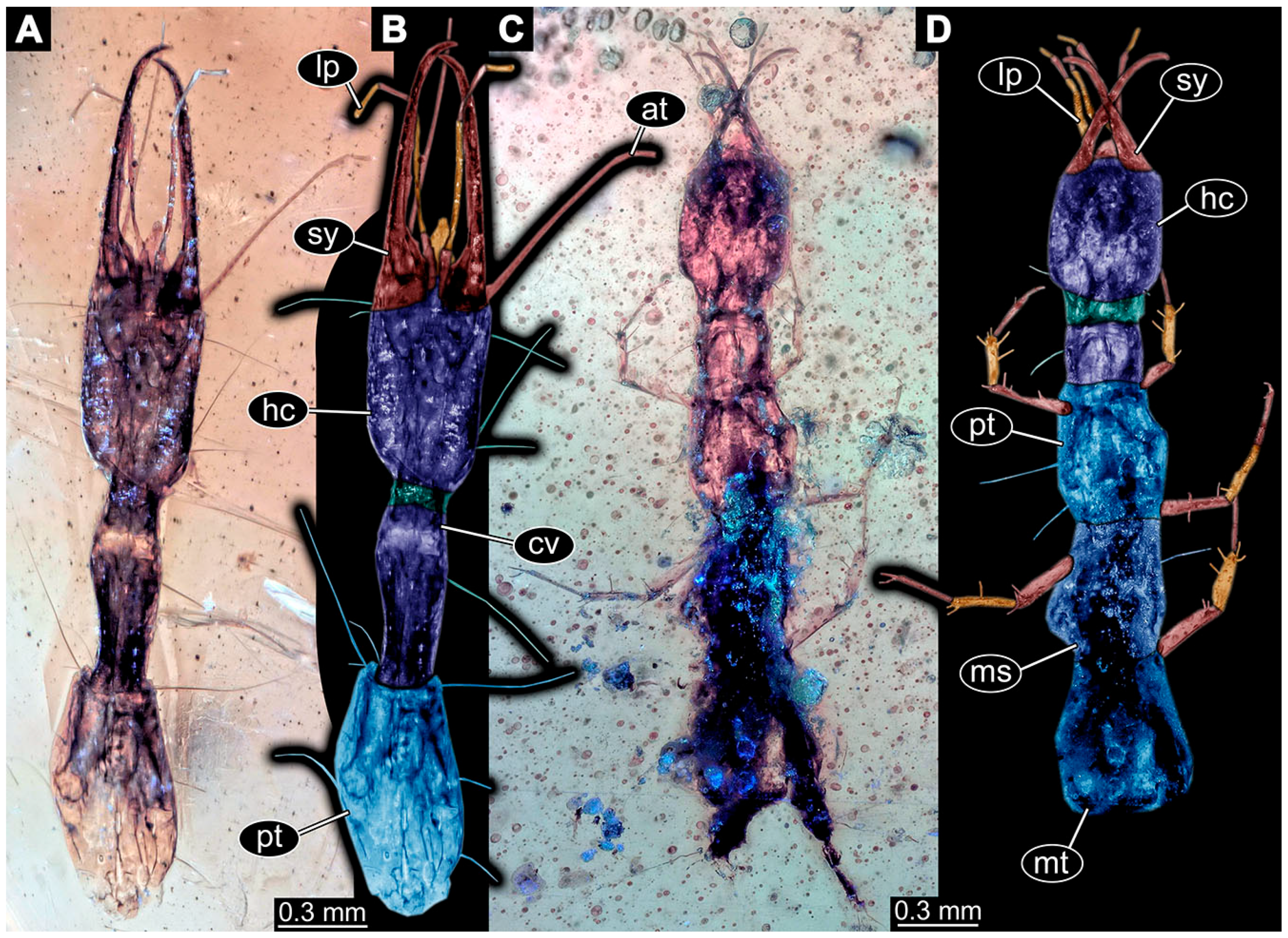
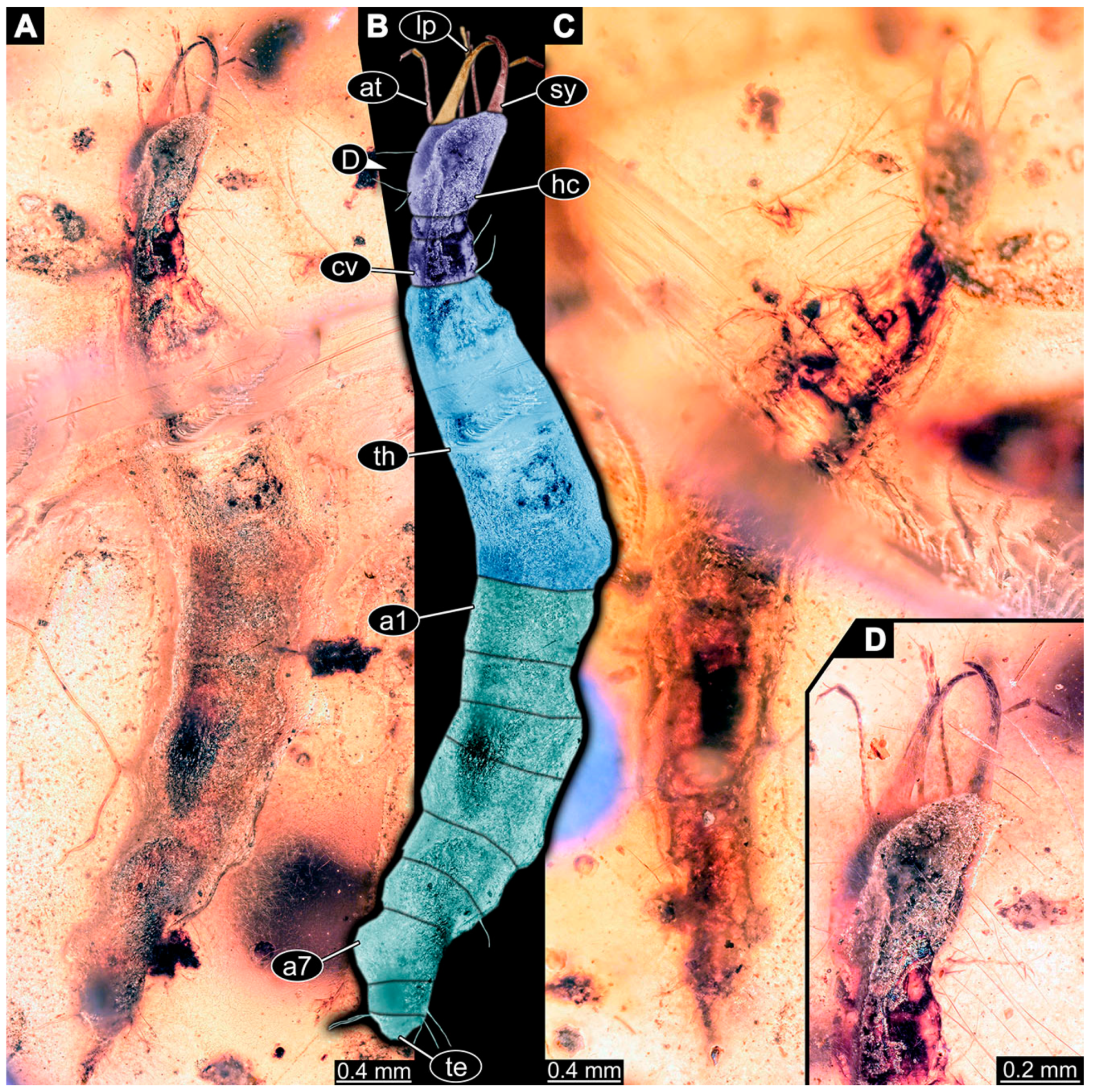
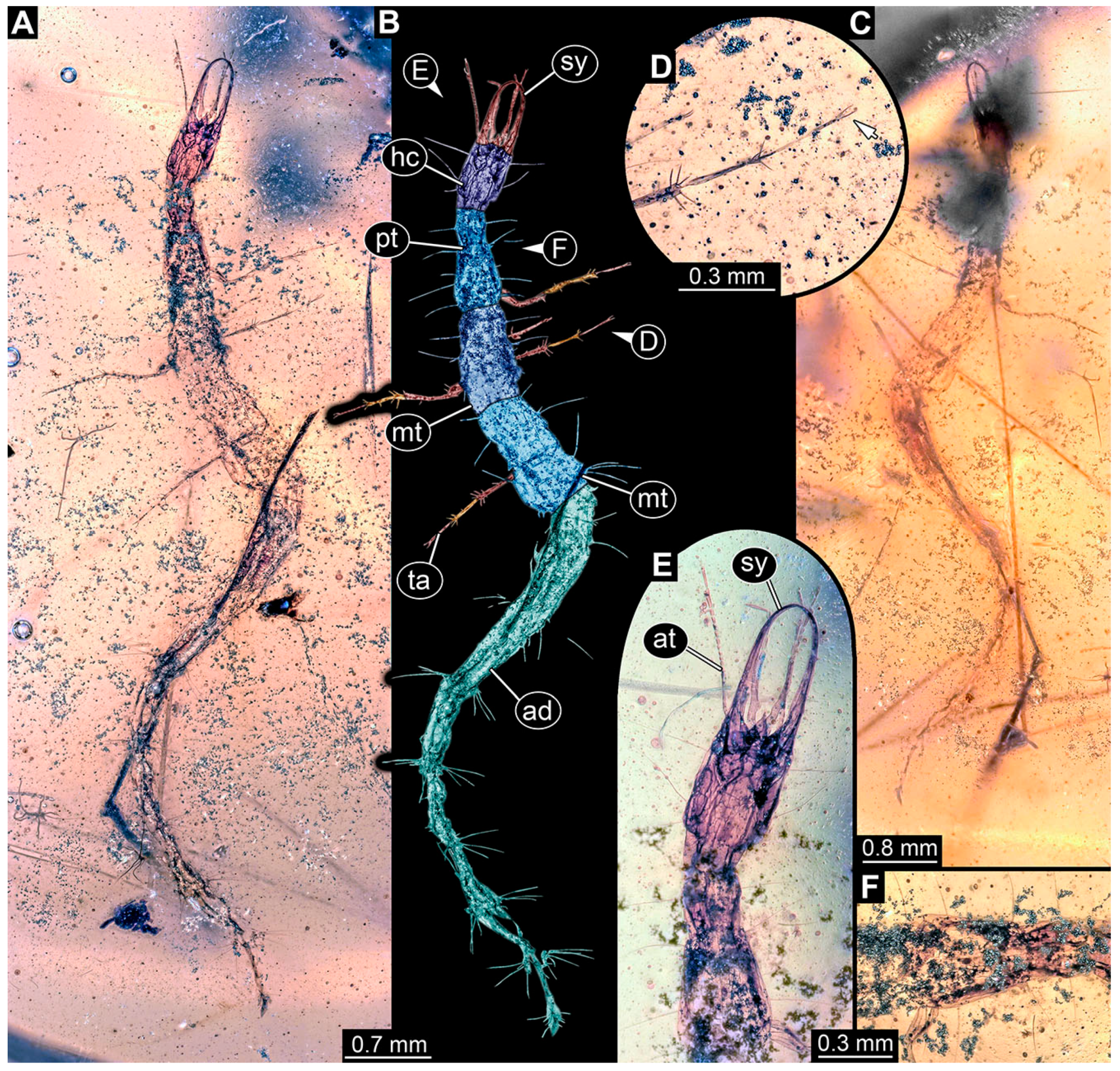
- Specimen 6645 (accession number nsw_epa_HUNT_04Williams_r_161299) is well accessible (Figure 2A); it likely represents a final larval stage. The specimen was collected during the NSW EPA-MRHI survey, HUNT-04, in the William River at Rocky Crossing, New South Wales, Australia (32°17′7″ S, 151°48′7″ E), on 16 December 1999. Head with antennae, stylets, and labial palps. Antennae are elongated and show subdivision into numerous elements (about 12). Stylets are proximally straight, distally more curved, tapering distally. Labial palps are slightly shorter than antennae, subdivided into at least four elements apparent (Figure 2E). Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 2A,F). Coxa and trochanter are short, not well accessible; femur tubular and elongated; tibia similar to femur, but slightly shorter; tarsus quite slender again and tapering distally with a pair of claws (Figure 2F). Thorax is subdivided into pro-, meso-, and metathorax, with a long cervix anteriorly. Abdomen with 11 units. The cervix and prothorax show a longitudinal groove (Figure 2A). The trunk segments posterior to the prothorax with a distinct pattern of dorsal sclerotisations. After the anterior third of the segment, a distinct fold is apparent, subdividing the dorsal region of each segment into two distinct subregions (Figure 2A). Prominent setae arise from the trunk appendage elements. Femur and tibia are distally with a set of three to five setae each. Trunk segments with prominent long setae. These arise close to the posterior end of each segment and appear to form a kind of ring around the segment.
- (10)
- Specimen 6646 (accession number nsw_epa_HUNT_04Williams_r_161299; in the same vial as specimen 6645) is well accessible (Figure 2B); it likely represents a final larval stage. The specimen was collected during the NSW EPA-MRHI survey, HUNT-04, in the William River at Rocky Crossing, New South Wales, Australia (32°17′7″ S, 151°48′7″ E), on 16 December 1999. Overall body outline is well accessible. Head with antennae, stylets, and labial palps (Figure 2G). Antennae are elongated, subdivided into numerous elements (about 11). Stylets are proximally straight, distally curved, tapering distally. Labial palps are subdivided into five elements (Figure 2G). Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 2B). Coxa is tubular and elongated; trochanter not accessible; femur similar to coxa, but thinner; tibia similar to femur, but a bit shorter; tarsus quite slender again and tapering distally with a pair of claws (Figure 2B). Thorax is subdivided into pro-, meso-, and metathorax with a long cervix anteriorly. Abdomen with 11 units. The cervix and prothorax show a longitudinal groove (Figure 2B). The trunk segments posterior to the prothorax with a distinct pattern of dorsal sclerotisations. After the anterior third of the segment, a distinct fold is apparent, subdividing the dorsal region of each segment into two distinct subregions (Figure 2B). Prominent setae arise from the trunk appendage elements. Femur and tibia are distally with a set of three to six setae each. Trunk segments with prominent long setae. These arise close to the posterior end of each segment and appear to form a kind of ring around the segment.
- (11)
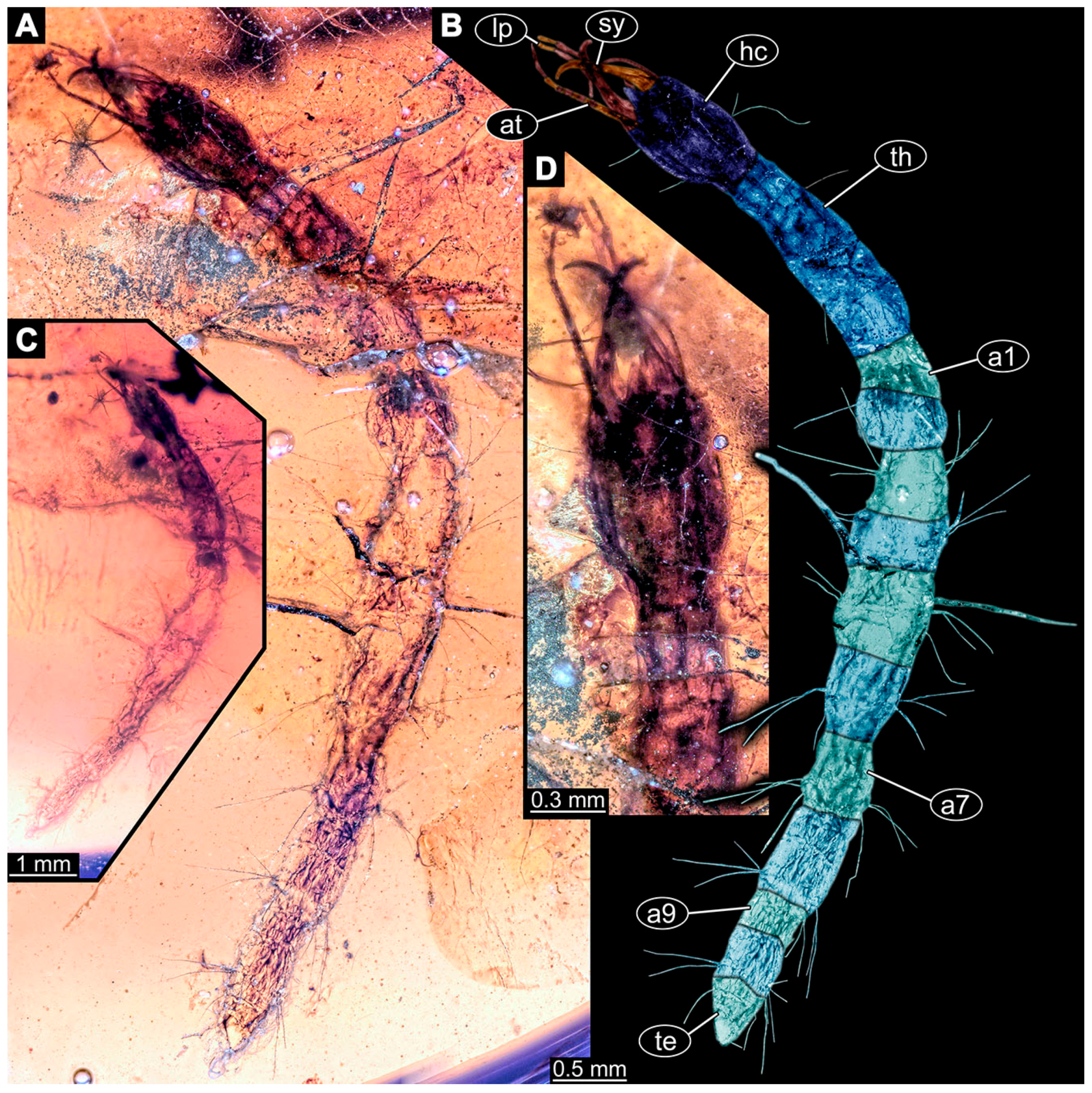
- Specimen 6647 (accession number nsw_epa_MANN16_batar_ck_220499) is well accessible (Figure 2C); it likely represents a penultimate larval stage. The specimen was collected during NSW EPA NSW EPA-MRHI survey, MANN-16, in the tributary of Batar Creek, New South Wales, Australia (31°40′57″ S, 152°40′7″ E), on 22 April 1999 by N. Waddell. Head with antennae, stylets, and labial palps. Antennae are elongated, subdivided into about six elements. Stylets are curved and tapering distally. Subdivision of the labial palps not well accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. Thorax is subdivided into pro-, meso-, and metathorax with a longer cervix anteriorly. Abdomen with 11 units. The cervix shows a longitudinal groove (Figure 2C). Trunk segments with prominent long setae. These arise close to the posterior end of each segment and appear to form a kind of ring around the segment.
- (12)
- Specimen 6648 (accession number nsw_epa_MANN16_batar_ck_220499; in the same vial as specimen 6647) is well accessible (Figure 2D); it likely represents a final larval stage. The specimen was collected during the NSW EPA NSW EPA-MRHI survey, MANN-16, in the tributary of Batar Creek, New South Wales, Australia (31°40′57″ S, 152°40′7″ E), on 22 April 1999 by N. Waddell. Head with antennae, stylets, and labial palps. Antennae are elongated, subdivided into several elements. Stylets proximally straight, distally more curved, tapering distally. Subdivision of the labial palps not well accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 2D) well preserved, but they are entangled with each other, partly obscuring subdivisions and details. Coxa is tubular and elongated; trochanter not well accessible; femur similar to coxa, but thinner; tibia similar to femur, but slightly shorter; tarsus quite slender again and tapering distally with a pair of claws. Thorax is subdivided into pro-, meso-, and metathorax with a longer cervix anteriorly. Abdomen with at least nine units. The cervix shows a longitudinal groove (Figure 2D). Trunk segments with prominent long setae. These arise close to the posterior end of each segment and appear to form a kind of ring around the segment.
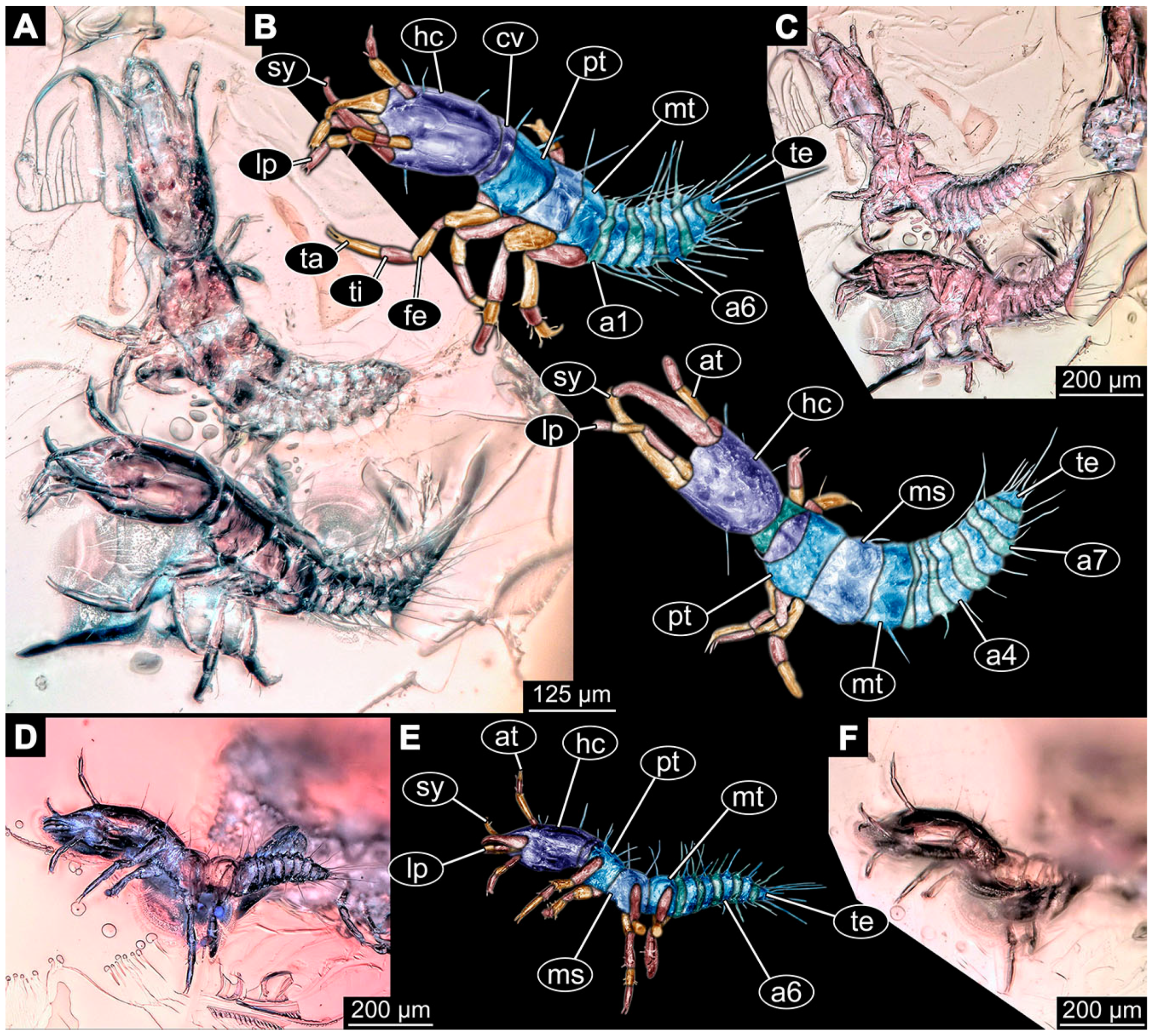
- (13)
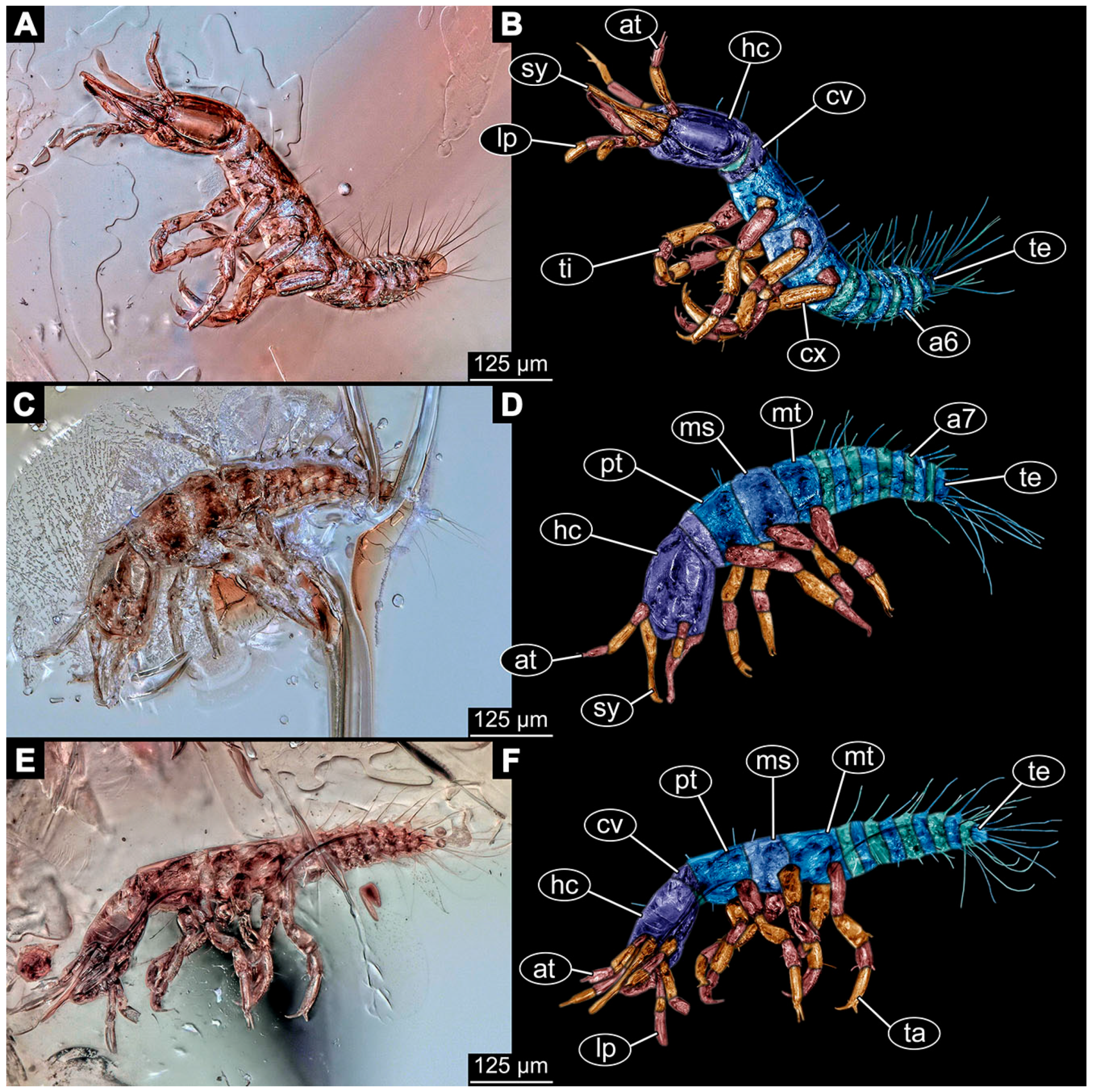
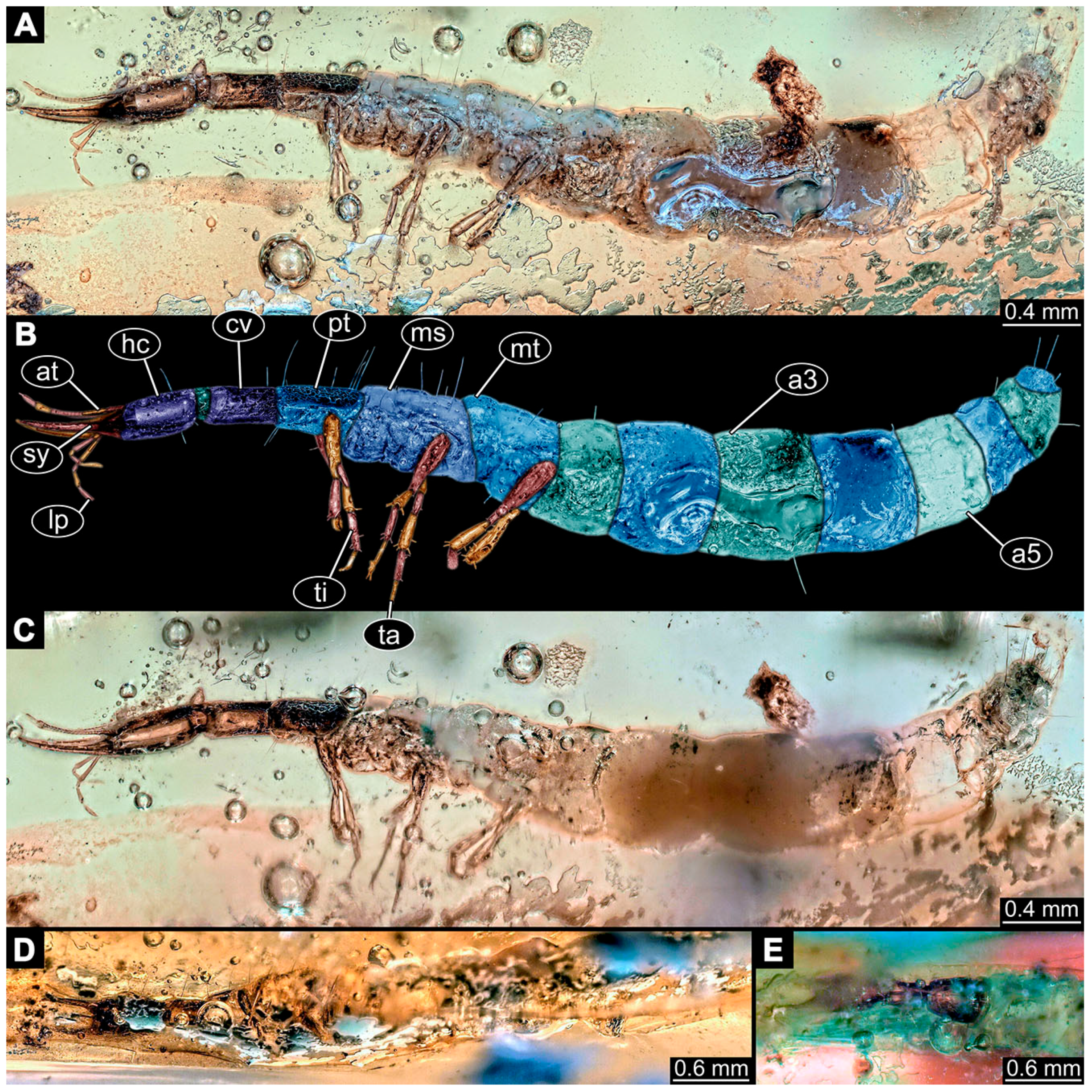
- Specimen 6649 (accession number nsw_epa_MANN16_batar_ck_220499; in the same vial as specimens 6647, 6648) is well accessible (Figure 3A); it likely represents a penultimate or final larval stage. The specimen was collected during the NSW EPA NSW EPA-MRHI survey, MANN-16, in the tributary of Batar Creek, New South Wales, Australia (31°40′57″ S, 152°40′7″ E), on 22 April 1999 by N. Waddell. Head with antennae, stylets, and labial palps. Antennae are elongated, subdivided into several elements (about 12). Stylets are proximally straight, distally more curved, tapering distally. Labial palps are subdivided into at least four elements, proximal region not well accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 3A), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. Thorax is subdivided into pro-, meso-, and metathorax, with a long cervix anteriorly. Abdomen with at least 10 units. The cervix and prothorax show a longitudinal groove (Figure 2B). The trunk segments posterior to the prothorax with a distinct pattern of dorsal sclerotisations. After the anterior third of the segment, a distinct fold is apparent, subdividing the dorsal region of each segment into two distinct subregions (Figure 3A). Prominent setae arise from the trunk appendage elements. Femur and tibia are distally with a set of three to four setae each. Trunk segments with prominent long setae. These arise close to the posterior end of each segment and appear to form a kind of ring around the segment.
- (14)
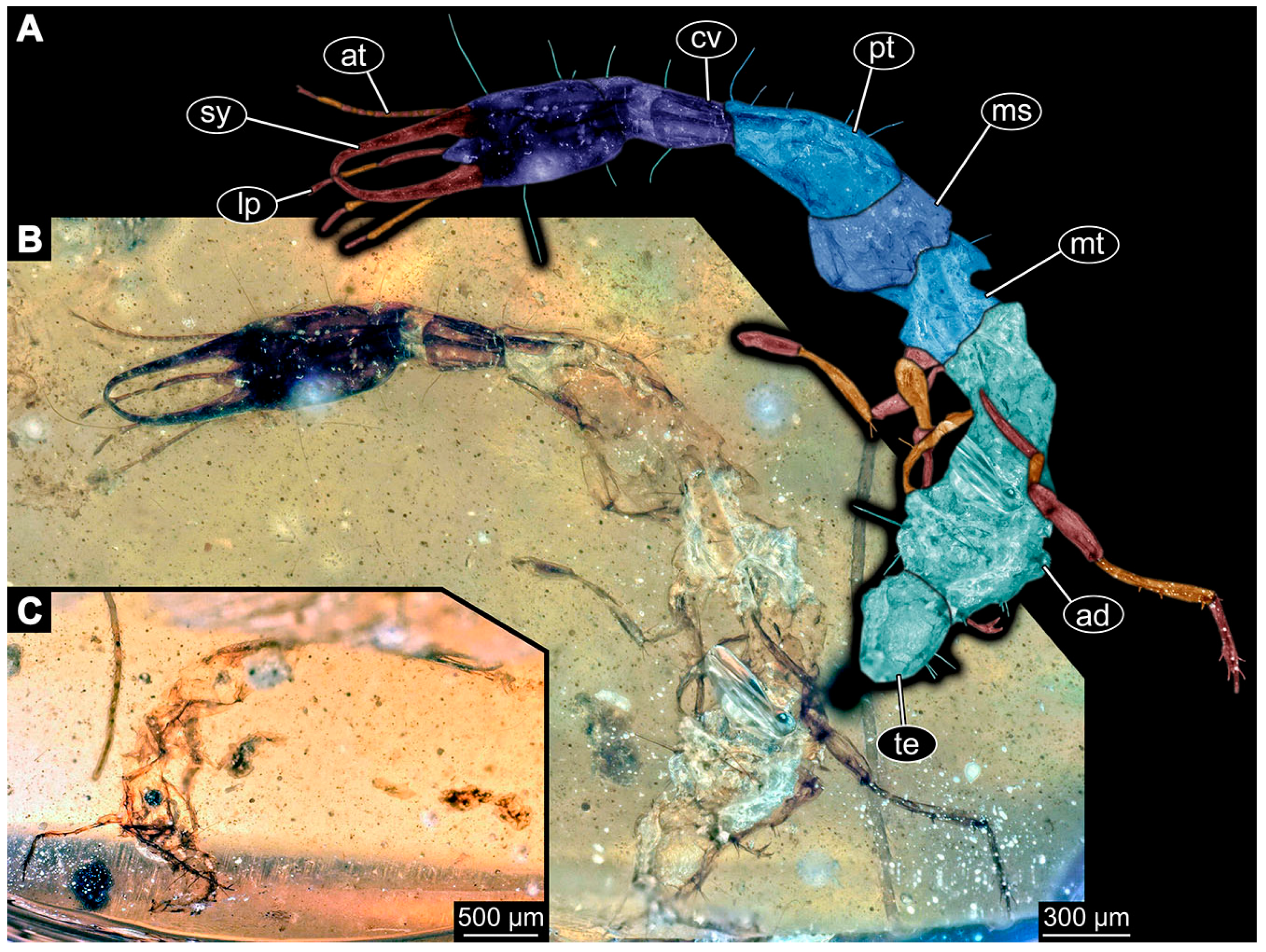
- Specimen 6650 (accession number nsw_epa_MANN16_batar_ck_220499; in the same vial as specimen 6647–6649) is well accessible (Figure 3B); it likely represents a final larval stage. The specimen was collected during the NSW EPA NSW EPA-MRHI survey, MANN-16, in the tributary of Batar Creek, New South Wales, Australia (31°40′57″S, 152°40′7″ E), on 22 April 1999 by N. Waddell. Head with antennae, stylets, and labial palps. Antennae are elongated, subdivided into several elements (about 14). Stylets are curved and tapering distally. Labial palps are subdivided into at least four elements apparent, proximal region not well accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 3B), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. Coxa is tubular and elongated; trochanter not accessible; femur similar to coxa, but shorter; tibia similar to femur, but a bit longer and wider; tarsus similar to tibia, but shorter and tapering distally with a pair of claws. Thorax is subdivided into pro-, meso-, and metathorax, with a long cervix anteriorly. Abdomen with at least 10 units. Prominent setae arise from the trunk appendage elements. Femur and tibia are distally with a set of four to six setae each. Few, less prominent setae also apparent further proximally, no clear pattern apparent. Trunk segments with prominent long setae. These arise close to the posterior end of each segment and appear to form a kind of ring around the segment.
- (15)
- Specimen 6651 (accession number nsw_epa_BEGA23_Paddys_crk_171199) is well accessible (Figure 3C); it likely represents a penultimate or final larval stage. The specimen was collected during the NSW EPA-MRHI survey, BEGA-23, in Paddys Creek, New South Wales, Australia (36°33′3″ S, 149°80′8″ E), on 16 December 1999 by Jamie Potts. Head with elongate antennae, stylets, and one labial palp left. Antennae are subdivided into several elements (about 13). Stylets are curved and tapering distally. Labial palps are subdivided into five elements, the other one is broken at the second segment (Figure 3E). Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus) are mainly covered by the trunk. Coxa and trochanter are not accessible; femur tubular and elongated; tibia similar to femur, but thinner; tarsus quite slender again and tapering distally with a pair of claws. Thorax is subdivided into pro-, meso-, and metathorax with a long cervix anteriorly. Abdomen with at least 10 units. The cervix and prothorax show a longitudinal groove. The trunk segments posterior to the prothorax with a distinct pattern of dorsal sclerotisations. After the anterior third of the segment, a distinct fold is apparent, subdividing the dorsal region of each segment into two distinct subregions (Figure 3C). Prominent setae arise from the trunk appendage elements. Femur and tibia are distally with a set of three to five setae each. Trunk segments with prominent long setae. These arise close to the posterior end of each segment and appear to form a kind of ring around the segment.
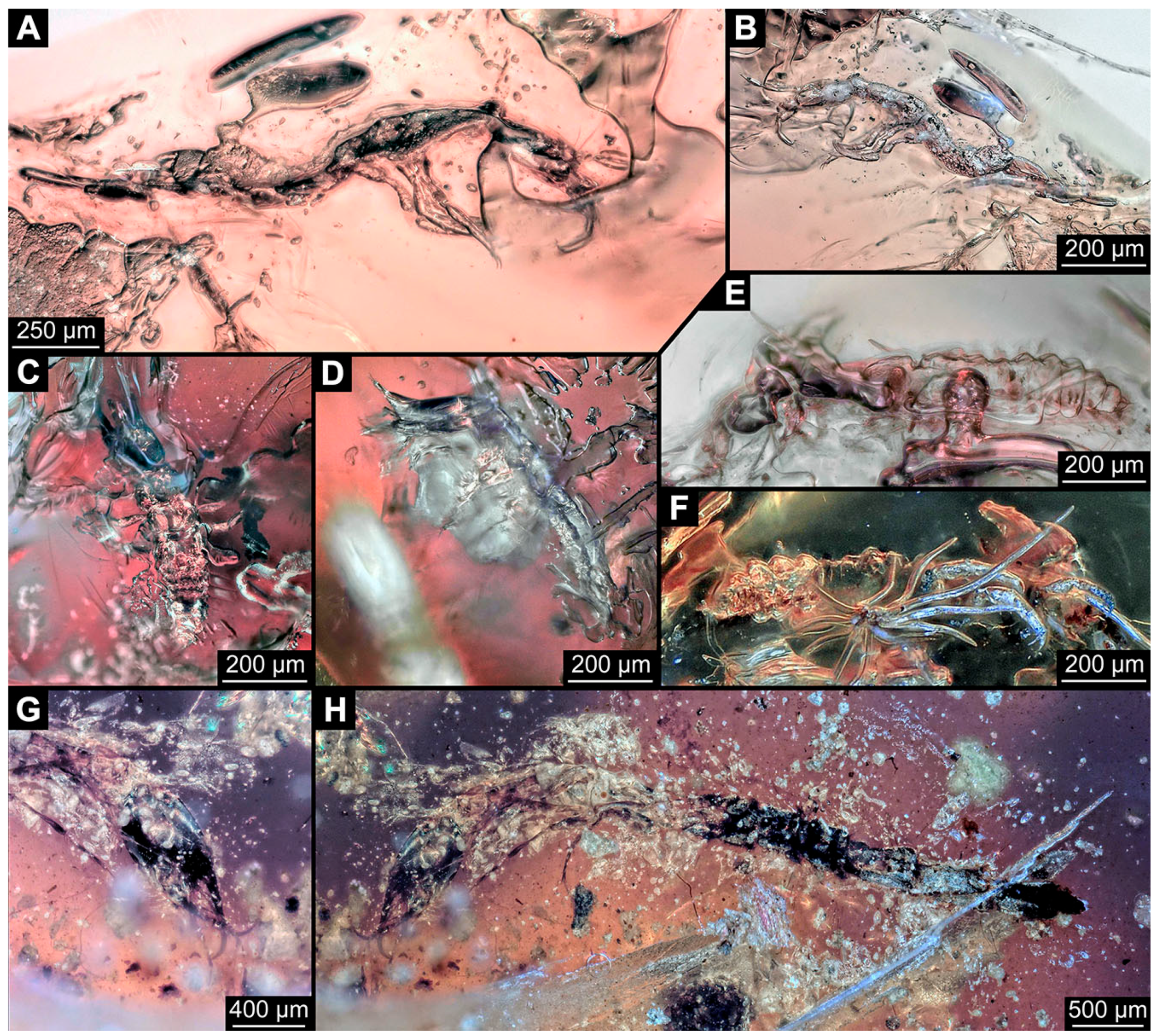
3.3. Additional Fossil Larval Representatives of Nevrorthidae in Baltic Amber
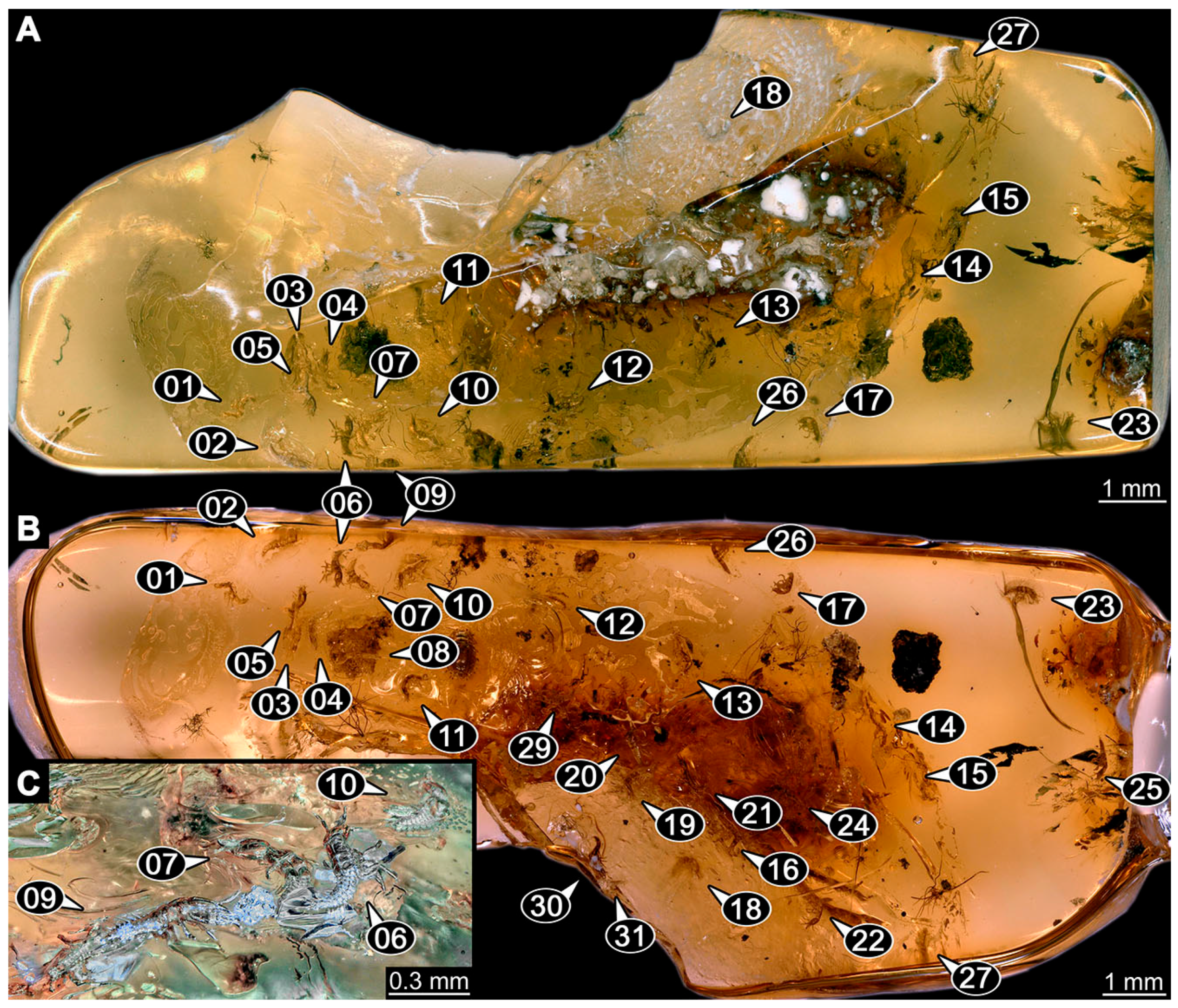
- (16)
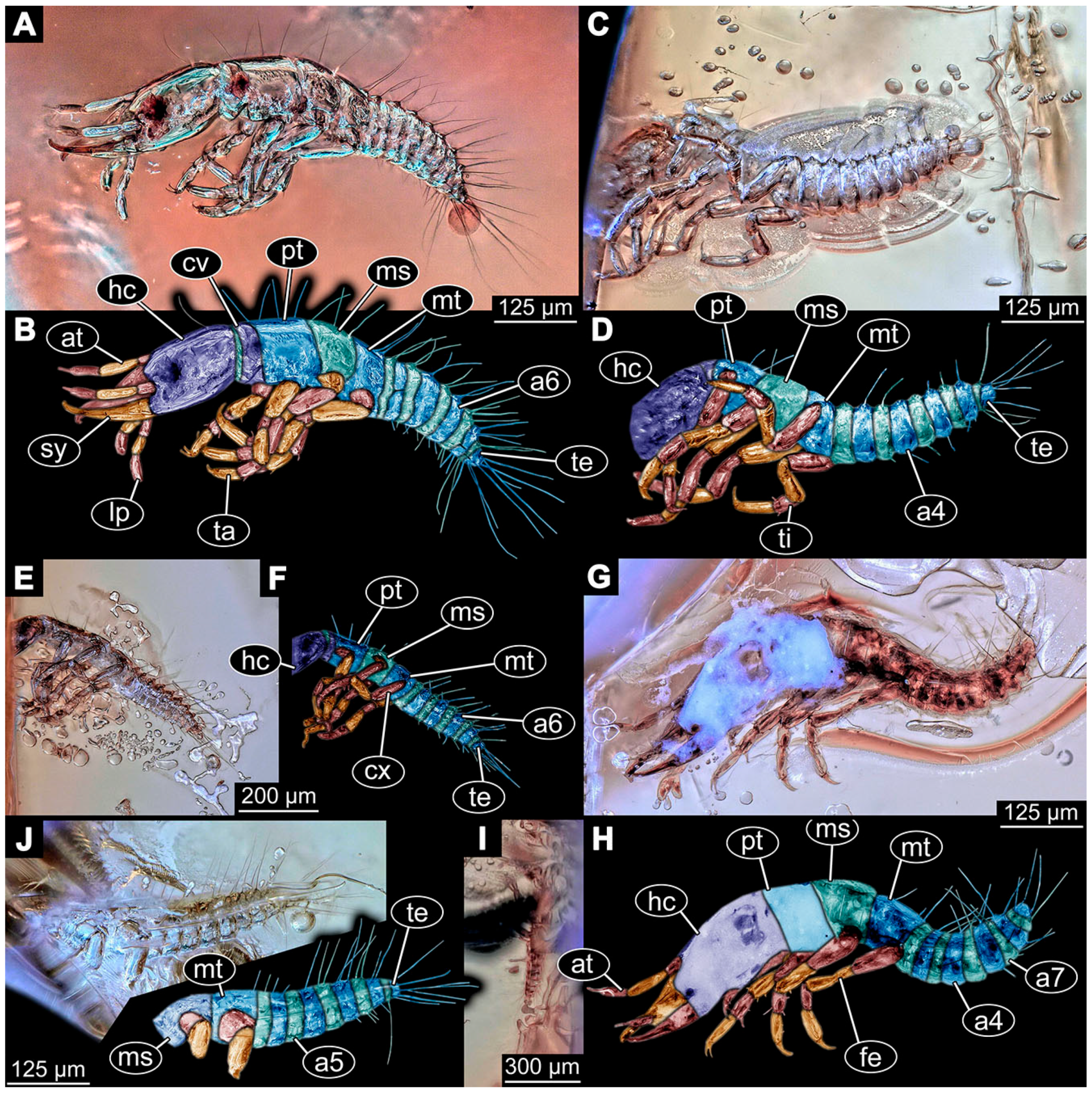
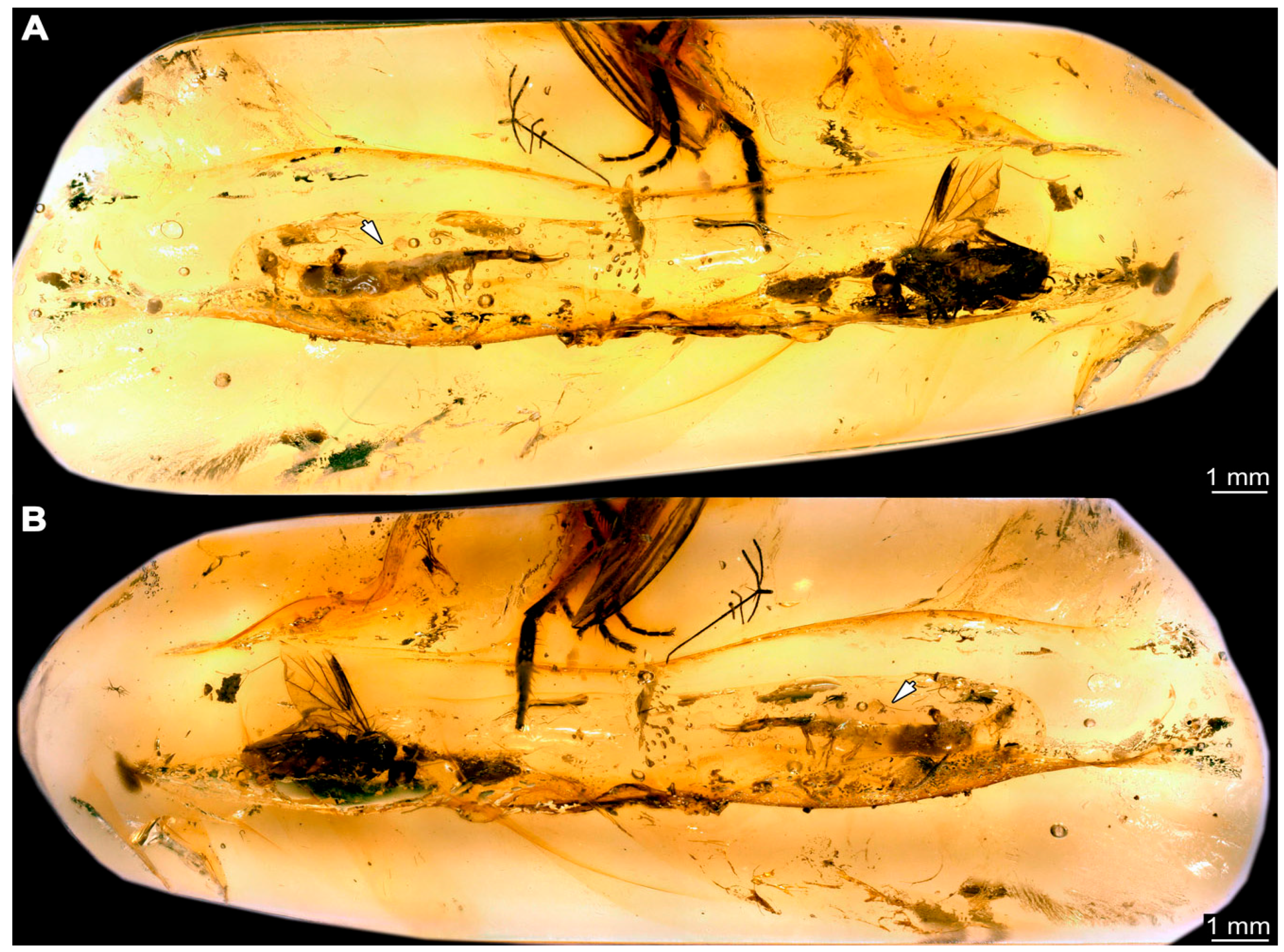
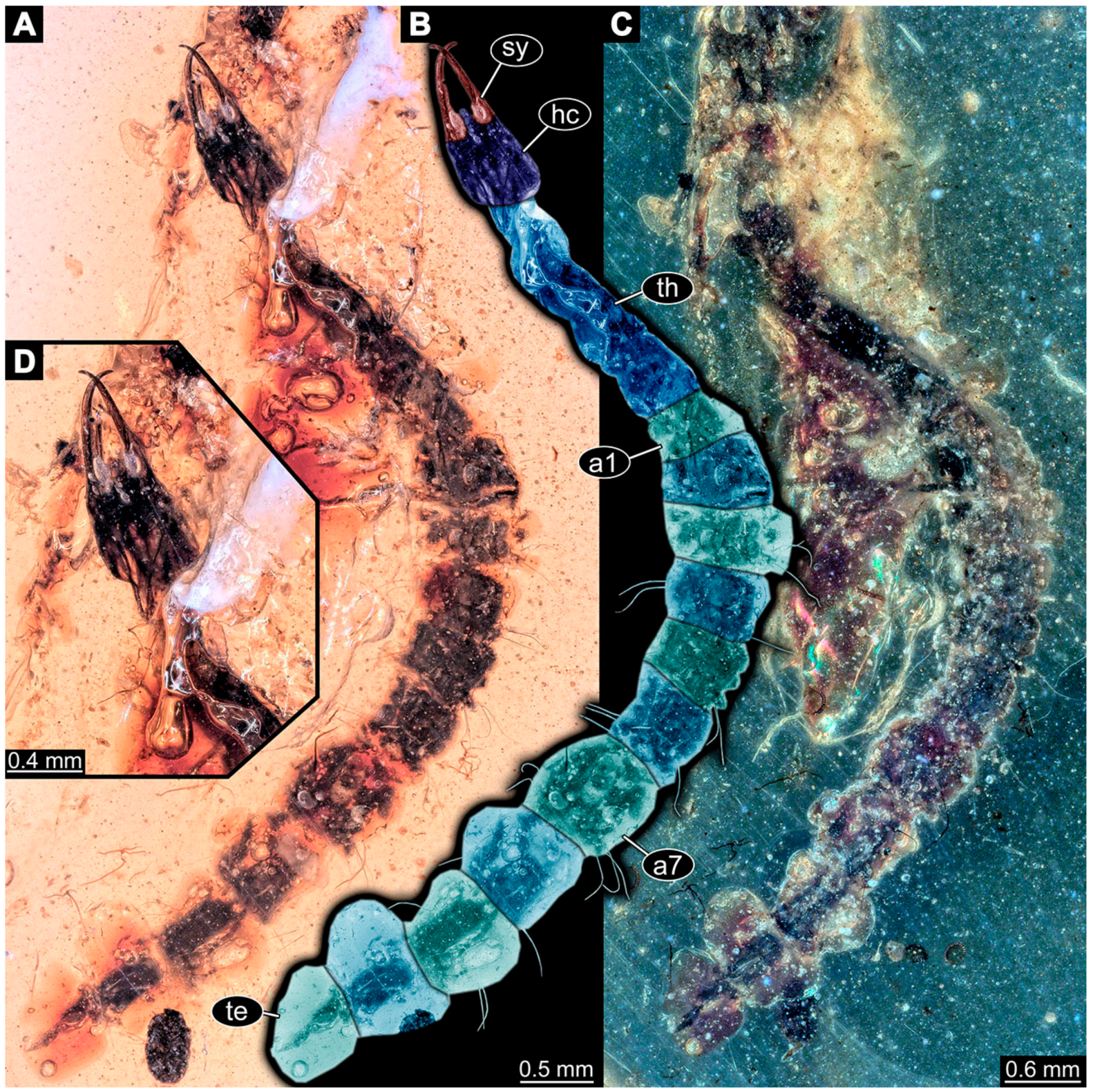
- Specimen 6652 (PED 0871 spec 1) is preserved together with numerous specimens in a single piece of Baltic amber (Figure 4). The specimen is accessible in lateral view from both sides of the body (Figure 5E,F). Other aspects are not accessible. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The details of the head (antennae, stylets, and labial palps) are not well accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus), but subdivisions not well visible. Subdivisions of the thorax and abdomen largely obscured by bubbles. A few elongate setae arise from the thorax and abdomen segments.
- (17)
- Specimen 6653 (PED 0871 spec 2) is accessible from the ventral side and less well from the dorsal side (Figure 5A–D). It is preserved in the same amber piece as specimen 6652. Well-accessible details include the abdomen segments with several setae and parts of the thorax appendages. Other aspects are not well accessible. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. Most of the anterior body is destroyed. Of the head appendages, only one stylet and a part of the antenna remain. Stylets are proximally straight, then they get more curved, tapering distally. Parts of trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 5B). Coxa is tubular and bellied; trochanter not visible; femur a bit thinner and elongated; tibia shorter and thicker than the femur; tarsus a bit longer than tibia and tapering distally. Also, the ten abdomen units with their subdivisions are well accessible. Numerous elongate setae arise from the abdomen segments; insertion areas obscured. They seem to arise in the middle of each segment.
- (18)
- Specimen 6654 (PED 0871 spec 3) is only accessible in dorsal view (Figure 5G,H). Other aspects are not accessible. It is preserved in the same amber piece as specimens 6652 and 6653. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. Head outline well apparent in dorsal view with a short cervix. Head with stylets and one stout antenna left. The rest of the head appendages are not accessible. Stylets taper distally; they are straight proximally broader, but sharply curved more distally. The antenna, subdivided into three elements, appears a bit thicker than at other specimens (Figure 5H). Most of the subdivisions of the thorax and trunk appendages are obscured. On the trunk no setae are visible.
- (19)
- Specimen 6655 (PED 0871 spec 4) is only accessible from the lateral left side (Figure 5I,J), other aspects are not accessible. It is preserved in the same amber piece as specimens 6652–6654. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The amber is rather clear and the overall outline is easily accessible. The specimen is a bit curved dorsally. The head outline with a short cervix is well apparent. Head with antennae, stylets, and labial palps. Subdivisions are a bit obscured by Verlumung. Antennae elongate slender, subdivided into three elements. The stylets are curved and tapering distally. They seem to be as long as the antennae. Labial palps are present, but mostly covered by the stylets. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 5J), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. Coxa is tubular and elongate; trochanter significantly shorter; femur shorter than the coxa and a bit thicker; tibia thinner and a bit shorter again; tarsus quite slender, elongate and longer than tibia, distally with a pair of claws. The thorax is clearly subdivided into pro-, meso-, and metathorax. Body carrying numerous elongate setae; insertion areas obscured. They seem to be longer at the trunk end.
- (20)
- Specimen 6656 (PED 0871 spec 5) is accessible in left and right lateral view of the body (Figure 6D–F). The distal body parts of the left view are obscured, other aspects are not accessible. The specimen is preserved in the same amber piece as specimens 6652–6655. It shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. Furthermore, it seems a bit smaller than the other specimens. Overall outline is well accessible from the lateral right side of the body. The head with a short cervix is seen in ventral view with antennae, stylets, and labial palps. Antennae elongate slender, subdivided into three elements (Figure 6F) with a few setae. The stylets are curved and tapering distally. Labial palps are slightly shorter than antennae, subdivided into at least three elements apparent, proximal region not accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 6E). Coxa is tubular and elongated; trochanter not accessible; femur tubular and elongated as well; tibia shorter and thicker than femur; tarsus quite slender, elongate and longer than tibia, distally with a pair of claws. The thorax is clearly subdivided into pro-, meso-, and metathorax. Body carrying numerous elongate setae; most insertion areas are obscured. They seem to be longer at the trunk end.
- (21)
- Specimen 6657 (PED 0871 spec 6) is accessible in dorsal and ventral view (Figure 6A lower, B upper, C lower). It is preserved in the same amber piece as specimens 6652–6656. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. Overall outline is well accessible from both sides. Head with stout antennae, stylets, and labial palps. Antennae are a bit thicker in shape than in other specimens and subdivided into three elements (Figure 6B, upper). The stylets are curved with a broad proximal region and tapering distally. Labial palps are slightly longer than antennae, subdivided into at least three elements apparent, proximal region not accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 6B, upper). Coxa is tubular and elongated; trochanter conical and shorter; femur tubular, shorter than coxa; tibia similar, but shorter; tarsus similar as well, but tapering distally and with a pair of claws. The thorax is clearly subdivided into pro-, meso-, and metathorax with a short cervix and the abdomen is divided into 10 units. All over the body, several setae arise. Most insertion areas are obscured by Verlumung, but at the abdomen, they seem to arise in the middle of each segment.
- (22)
- Specimen 6658 (PED 0871 spec 7) is accessible in dorsal and ventral view (Figure 6A upper B lower, C upper). It is preserved in the same amber piece as specimens 6652–6657. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. Overall outline is well accessible from both sides. The subdivisions of the head appendages (antennae, stylets, and labial palps) are largely obscured by Verlumung. Only one stout antenna, both stylets, and one labial palp remain. Antenna is elongated and subdivided into three elements. The stylets taper distally; they are straight and broad proximally, but sharply curved more distally. Labial palp is elongated and slightly longer than antenna, subdivided into four elements. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus), but proximal not well visible; distally with a pair of claws (Figure 6B, lower). The thorax is subdivided into pro-, meso-, and metathorax. Along the short cervix and thorax, a distinct fold is apparent. Abdomen slightly obscured by Verlumung, but the ten abdomen units still well accessible. A few setae arise from the body, most of them at the abdomen segments. They seem to arise in the middle of each segment.
- (23)
- Specimen 6659 (PED 0871 spec 8) is only accessible from left lateral side of the body (Figure 7A,B). It is preserved in the same amber piece as specimens 6652–6658. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. Overall outline is a bit obscured by tiny bubbles, but still accessible. Head with one antenna, both stylets, and labial palps. Antenna is elongated, quite slender, and tapering distally, subdivided into three elements. Stylets are elongate and tapering distally, but mostly cover each other. Labial palps are slightly shorter than antenna, subdivided into at least three elements apparent, proximal region not accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 7B), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. Coxa is tubular and elongated; trochanter not accessible; femur tubular and slightly shorter than coxa; tibia similar; tarsus quite slender, elongate and distally with a pair of claws. The thorax is clearly subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. Only at the trunk end are a few elongate setae visible. The insertion areas are obscured.
- (24)
- Specimen 6660 (PED 0871 spec 9) is only accessible from ventral view of the body (Figure 7C,D). It is preserved in the same amber piece as specimens 6652–6659. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The overall outline is well accessible. The head appendages (antennae, stylets, labial palps) and the abdomen units are a bit obscured by Verlumung. Stout antennae, with a few setae elongate slender; subdivision into at least three elements. Stylets with a broad proximal region tapering distally. The distal parts of the stylets are mostly covered by the labial palps, which are slightly shorter than the antennae and subdivided into four elements. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 7D). Only one leg is fully accessible. Coxa tubular, elongate; trochanter not accessible; femur tubular and similar to the coxa; tibia similar, but shorter than femur; tarsus similar, but longer than tibia, distally with a pair of claws. The thorax is clearly subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. In spite of Verlumung the subdivisions of the abdomen are well accessible. The ten abdomen units bear several setae, which seem to have their insertion areas at the middle of each unit.
- (25)
- Specimen 6661 (PED 0871 spec 10) is only accessible from the right lateral side of the body (Figure 7E,F). It is preserved in the same amber piece as specimens 6652–6660. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. Despite the Verlumung, the overall outline is well accessible. Head with antennae, stylets, and labial palps. Stout antennae subdivided into three elements. The stylets taper distally. They are straight proximally with a broad proximal region, but a bit curved more distally. Labial palps look similar to the antennae, subdivided into at least three elements apparent, proximal region not accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 7D), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. The thorax is clearly subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. Only a few setae arise at the ten abdomen units. The insertion areas are mostly obscured by Verlumung.
- (26)
- Specimen 6662 (PED 0871 spec 11) is only accessible from the left lateral side of the body (Figure 7G). It is preserved in the same amber piece as specimens 6652–6661. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The specimen is strongly curved ventrally, basically adopting a U-shaped posture. Overall body outline well accessible, yet most details, especially of the more distal body and the head are obscured by Verlumung, dark spots, and bubbles. Head with antennae, stylets, and labial palps, but details such as subdivision obscured. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. No setae or clear subdivision of the trunk are accessible.
- (27)
- Specimen 6663 (PED 0871 spec 12) is only accessible from the lateral right side of the body (Figure 8A,B). It is preserved in the same amber piece as specimens 6652–6662. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The specimen is a bit curved ventrally, basically adopting a U-shaped posture. Overall body outline well accessible. Head with stout antennae and stylets. Antennae are elongated and subdivided into three elements, tapering distally. The labial palps are totally covered by the head and not accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 8B), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. Most accessible are pairs of claws, which are located at the distal end of the tarsi. The thorax is clearly subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. Numerous elongate setae arise from the ten abdomen units, they seem to be longer at the trunk end.
- (28)
- Specimen 6664 (PED 0871 spec 13) is only accessible from the lateral right side of the body (Figure 8C,D). It is preserved in the same amber piece as specimens 6652–6663. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The overall outline is more difficult to identify. Large parts of the body are obscured by bubbles. Head with very stout antennae, stylets, and labial palps. Antennae are elongated and tapering distally. Stylets are straight with a broad proximal region and tapering distally as well. The labial palps are mostly not accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 8D), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. The thorax is subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. Numerous elongate setae arise from the ten abdomen units.
- (29)
- Specimen 6668 (PED 0871 spec 17) is only accessible from the lateral right side of the body (Figure 8E,F). It is preserved in the same amber piece as specimens 6652–6664. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. At this part the amber is very clear, hence the overall body outline of the specimen is well accessible. Head with antennae, stylets, and labial palps. Stout antennae are elongated and show subdivision into three to four elements. Stylets taper distally. They are straight proximally, but sharply curved more distally. The labial palps are covered by the head and not accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 8F). Coxa appears conical; trochanter tubular, elongate; femur similar, but shorter; tibia similar as well, but shorter than femur; tarsus similar too, but taper distally with a pair of claws at the end. The thorax is subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. Prominent setae arise from the trunk appendage elements. Femur and tibia are distally with a set of three setae each. Few, less prominent setae also apparent further proximally, no clear pattern apparent. Trunk segments with prominent long setae. These arise in the middle of each segment. The abdomen is subdivided into at least six units apparent, posterior region not accessible.
- (30)
- Specimen 6669 (PED 0871 spec 18) is only accessible from the lateral right side of the body (Figure 9A). It is preserved in the same amber piece as specimens 6652–6664 and 6668. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The specimen is a bit curved ventrally, basically adopting a U-shaped posture. The overall body outline is partly difficult to identify. Head with antennae, stylets, and labial palps, but the details, such as subdivision are obscured. Trunk appendages are strongly verlumt, only vague outlines become apparent. Despite the Verlumung, the thorax shows a subdivision into pro-, meso-, and metathorax. A few setae arise from the trunk, yet due to the Verlumung, their insertion areas are obscured.
- (31)
- Specimen 6670 (PED 0871 spec 19) is only accessible from the lateral left side of the body (Figure 9B,C). It is preserved in the same amber piece as specimens 6652–6664, 6668, and 6669. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The outline of the specimen is accessible. Head with stout antennae, stylets, and labial palps. Antennae elongate and subdivide into three elements. Stylets taper distally. They are straight proximally with a broad proximal region, but curved more distally. The labial palps are mostly covered by the head, subdivided into at least three elements apparent, proximal region not accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 9C), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. The thorax is subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. Only a few setae arise from the ten abdomen units.
- (32)
- Specimen 6671 (PED 0871 spec 20) is only accessible from the lateral right side of the body (Figure 9D,E). It is preserved in the same amber piece as specimens 6652–6664 and 6668–6670. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The overall body outline is clearly accessible. Head with antennae, stylets, and labial palps. Stout antennae taper distally with a few setae. The stylets are mostly covered in this position by the head capsule, as well as the labial palps. Labial palps are subdivided into at least two elements apparent, proximal region not accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 9E). The thorax is subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. Numerous elongate setae arise from the trunk.
- (33)
- Specimen 6672 (PED 0871 spec 21) is only accessible from the lateral left side of the body (Figure 9F,G). It is preserved in the same amber piece as specimens 6652–6664 and 6668–6671. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The overall body outline is well accessible. Head with one antenna (left), stylets, and labial palps. Antennae elongate and subdivided into three elements. Stylets taper distally. They are straight proximally with a broad proximal region, but curved more distally. The labial palps are mostly covered by the stylets, subdivided into at least three elements apparent, proximal region not accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 9G), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. Most accessible are the pairs of claws, which are located at the distal end of the tarsi. The thorax is clearly subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. A few elongate setae arise on the trunk.
- (34)
- Specimen 6673 (PED 0871 spec 22) is only accessible from the lateral right side of the body (Figure 10A,B). It is preserved in the same amber piece as specimens 6652–6664 and 6668–6672. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The amber is very clear and the overall body outline is well accessible. Head with stout antennae, stylets, and labial palps. Antennae subdivided into three elements with a few setae apparent. Stylets with a broad proximal region tapering distally. Labial palps are subdivided into four elements. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 10B), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. Coxa appears elongate and tubular; trochanter similar, but way shorter; femur similar to coxa, but a bit shorter; tibia similar and shorter as well; tarsus similar too, but a bit longer than tibia and taper distally with a pair of claws at the end. The thorax is subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. Prominent setae arise from the trunk appendage elements and all over the trunk. At the trunk end, the setae are longer. The insertion areas seem to be in the middle of each abdomen segment.
- (35)
- Specimen 6674 (PED 0871 spec 23) is only accessible from the lateral right side of the body (Figure 10C,D). It is preserved in the same amber piece as specimens 6652–6664 and 6668–6673. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The overall body outline is partly obscured by bubbles, but still accessible. Head with antennae, stylets, and labial palps. One of the antennae is broken, the other one is elongated and subdivided into three elements. Stylets tapering distally. The labial palps are not accessible in this view. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 10D). The thorax is subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. Numerous elongate setae arise from the ten abdomen units. The insertion areas seem to be in the middle of each abdomen segment.
- (36)
- Specimen 6676 (PED 0871 spec 25) is only accessible from the lateral right side of the body (Figure 10E,F). It is preserved in the same amber piece as specimens 6652–6664 and 6668–6674. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. Overall body outline well accessible. Head with stout antennae, stylets, and labial palps. Antennae subdivided into three elements. Stylets tapering distally. Labial palps look similar to the antennae, subdivided into at least three elements apparent, proximal region not accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 10F), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. Coxa and femur are both tubular and elongated; tibia similar, but shorter than femur; tarsus similar, but a bit longer than tibia, distally with a pair of claws. The thorax is clearly subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. The abdomen is subdivided into 10 units with several setae. They seem to have their insertion areas in the middle of each unit and are longer at the trunk end. Also, the thorax appendages bear several setae.
- (37)
- Specimen 6677 (PED 0871 spec 26) is only partly accessible from the lateral left side of the body (Figure 11E,F). It is preserved in the same amber piece as specimens 6652–6664, 6668–6674 and 6676. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The body outline is well accessible, except for the head with the appendages. The anterior part of the head is outside the edge of the amber. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 11F), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. The thorax is clearly subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. Abdomen subdivided into 10 units. Trunk with several setae.
- (38)
- Specimen 6678 (PED 0871 spec 27) is only partly accessible from the lateral left side of the body (Figure 11C,D). It is preserved in the same amber piece as specimens 6652–6664, 6668–6674, 6676 and 6777. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The body outline is well accessible, except for the head with the appendages. The anterior part of the head outside the edge of the amber. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 11D). Coxa and femur are both tubular and elongated; tibia similar, but shorter than femur; tarsus similar, but a bit longer than tibia; distally one claw remains. The thorax is subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. The abdomen is subdivided into 10 units. Several setae arise at the middle of each unit.
- (39)
- Specimen 6679 (PED 0871 spec 28) is only accessible from the dorsal side of the body (Figure 11I) and sits very deep within the amber piece. As the amber piece contains multiple inclusions it is not possible to grind it further down. It is preserved in the same amber piece as specimens 6652–6664, 6668–6674, and 6676–6678. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. Most parts of the head, thorax, and trunk appendages are covered by other inclusions. At the abdomen, several setae arise.
- (40)
- Specimen 6680 (PED 0871 spec 29) is only accessible from the lateral right side of the body (Figure 11A,B). It is preserved in the same amber piece as specimens 6652–6664, 6668–6674, and 6676–6679. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The amber is very clear, hence the overall body outline of the specimen is well accessible. Head with antennae, stylets, and labial palps. Stout antennae are elongated and show subdivision into three elements. Stylets with a broad proximal region, curved and tapering distally. Labial palps are slightly shorter than antennae, subdivided into at least three elements apparent, proximal region not accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 11B), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. Coxa is tubular and elongated; trochanter conical and short; femur similar to coxa, but shorter; tibia similar as well, but shorter than femur; tarsus tubular as well, but a bit longer than tibia and tapering distally with a pair of claws. The thorax is clearly subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. Abdomen subdivided into 10 units. Numerous setae arise from the body. They seem to have their insertion areas in the middle of each unit and get longer at the trunk end. Also, the thorax appendages bear several setae.
- (41)
- Specimen 6681 (PED 0871 spec 30) is only accessible from the lateral left side of the body (Figure 11G,H). It is preserved in the same amber piece as specimens 6652–6664, 6668–6674, and 6676–6680. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. As the amber piece contains multiple inclusions most parts of the head and thorax are obscured. Head with one antenna, stylets, and labial palps. The stout antenna is subdivided into three elements. Stylets with a broad proximal region, slightly curved and tapering distally. The labial palps are mostly covered by the stylets. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 11H), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. Several setae arise from the ten abdomen units. Most insertion areas are obscured by Verlumung.
- (42)
- Specimen 6682 (PED 0871 spec 31) is only partly accessible from the lateral left side of the body (Figure 11J upper and lower). It is preserved in the same amber piece as specimens 6652–6664, 6668–6674, and 6676–6681. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The body outline is well accessible, except for the head and the trunk appendages. The anterior part of the specimen is outside the edge of the amber. The abdomen is subdivided into 10 units and bears several setae.
- (43)
- Specimen 6665 (PED 0871 spec 14) is accessible from the lateral left and right side (Figure 12A,B) and sits very deep within the amber piece. It is preserved in the same amber piece as specimens 6652–6664, 6668–6674, and 6676–6682. As the amber piece contains multiple inclusions it is not possible to grind it further down. Slightly more accessible parts include the anterior region of the specimen and the trunk appendages distal with the pair of claws. Other aspects are not accessible. Most of the posterior body is obscured by various inclusions.
- (44)
- Specimen 6666 (PED 0871 spec 15) is accessible from the lateral left and right side (Figure 12E,F) and sits very deep within the amber piece. It is preserved in the same amber piece as specimens 6652–6665, 6668–6674, and 6676–6682. As the amber piece contains multiple inclusions it is not possible to grind it further down. The most accessible parts are the distal end of one trunk appendage and a few units of the abdomen. Other aspects are not accessible. The remaining parts of the body are obscured by various inclusions.
- (45)
- Specimen 6667 (PED 0871 spec 16) is only accessible from the dorsal side (Figure 12C) and sits very deep within the amber piece. It is preserved in the same amber piece as specimens 6652–6666, 6668–6674, and 6676–6682. As the amber piece contains multiple inclusions it is not possible to grind it further down. The most accessible parts are the thorax segments and a few units of the anterior abdomen. Other aspects are not well accessible. The remaining parts of the body are obscured by various inclusions.
- (46)
- Specimen 6675 (PED 0871 spec 24) is only accessible from the lateral left side (Figure 12D) and sits very deep within the amber piece. It is preserved in the same amber piece as specimens 6652–6674 and 6676–6682. As the amber piece contains multiple inclusions it is not possible to grind it further down. The slightly more accessible part is the head with appendages. Other aspects are not well accessible. The remaining parts of the body are mostly obscured by various inclusions. Nevertheless, the specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, especially the posterior trunk, but appears much less slender, the segments of the abdomen are extremely short.
- (47)
- Specimen 6683 (PED 1373 spec 1) is only accessible from the lateral right side of the body (Figure 13A,B). The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The body outline is well accessible. Head with one elongate antenna, both stylets, and one labial palp (left). Antenna subdivided into three elements. Stylets with a broad proximal region tapering distally. Labial palp is subdivided into at least two elements apparent, proximal region not accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 13B), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. The thorax is subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. Several setae arise from the abdomen.
- (48)
- Specimen 6684 (PED 1373 spec 2) is only accessible from the lateral left side of the body (Figure 13E) and sits very deep within the amber piece. Hence, the specimen is not fully in focus. It is preserved in the same amber piece as specimen 6683. Subdivisions of the body are not visible. Nevertheless, the specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. Head with one elongate antenna, both stylets, and one labial palp (left). Stylets are curved and tapering distally. Labial palp is mainly obscured by the head. Trunk appendages are out of focus and entangled with each other. Abdomen with few setae.
- (49)
- Specimen 6685 (PED 1373 spec 3) is only accessible from the lateral right side of the body (Figure 13F) and sits very deep within the amber piece. Hence, the specimen is not fully in focus. It is preserved in the same amber piece as specimens 6683 and 6684. Subdivisions of the body are not well visible. Some subdivisions of the thorax and abdomen are slightly accessible. Nevertheless, the specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. Head with appendages not accessible. Trunk appendages are out of focus and entangled with each other, partly obscuring subdivisions. The thorax is subdivided into pro-, meso-, and metathorax. The abdomen has no visible setae.
- (50)
- Specimen 6686 (PED 1373 spec 4) is only accessible from the lateral right side of the body (Figure 13D) and sits very deep within the amber piece. It is preserved in the same amber piece as specimens 6683–6685. As the amber piece contains multiple inclusions it is not possible to grind it further down. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The slightly better accessible parts are the head, parts of the trunk appendages and several setae, which are arising from the abdomen. From the head appendages, only one elongate antenna is accessible, subdivided into three elements. The rest of the body is mainly obscured by various inclusions.
- (51)
- Specimen 6687 (PED 1373 spec 5) is only accessible from the ventral side of the body (Figure 13G,H). It is preserved in the same amber piece as specimen 6683–6686. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The overall body outline is well accessible. Head with antennae and labial palps. The stylets are obscured by Verlumung. Antennae and labial palps are subdivided into at least four elements apparent, proximal region not accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 13H), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. Tarsus distally with a pair of claws. A few setae arise from the abdomen.
- (52)
- Specimen 6688 (NHMD 115214 spec 1) is accessible from the dorsal and ventral side of the body (Figure 14 and Figure 15A–C). The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The amber is very clear, hence the overall body outline of the specimen is well accessible. Head with short and stout antennae, stylets, and labial palps. Antennae subdivided into three elements. Stylets are curved and tapering distally. Labial palps are mainly obscured by the stylets. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 15C), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. Also, they are largely obscured by Verlumung. The thorax is subdivided into pro-, meso-, and metathorax. Several setae arise from the trunk. The insertion areas are obscured by Verlumung.
- (53)
- Specimen 6689 (NHMD 115214 spec 2) is accessible from the dorsal and ventral side of the body (Figure 14 and Figure 15F–H). It is preserved in the same amber piece as specimen 6688. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The overall body outline is well accessible. Head with stout antennae, stylets, and labial palps (Figure 15H). Antennae subdivided into at least two elements, proximal region not accessible. Stylets and labial palps are mainly obscured by Verlumung. Only the tip of the stylet and the distal element of the labial palps are visible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 15H); these seem thicker in comparison to other specimens. The thorax is subdivided into pro-, meso-, and metathorax with a short cervix anteriorly (Figure 15G). Few setae arise from the trunk.
- (54)
- Specimen 6690 (NHMD 115214 spec 3) is accessible from the lateral right and ventral side of the body (Figure 14 and Figure 15D,E). It is preserved in the same amber piece as specimens 6688 and 6689. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The body outline is well accessible, except for the head with appendages and the thorax. The anterior part of body is largely obscured by Verlumung. Head with stout antennae, stylets, and labial palps (Figure 15D). Antennae subdivided into at least two elements apparent, proximal region not accessible. Stylets and labial palps are mostly obscured by Verlumung. Only one stylet and the distal element of the labial palps are visible. Stylet tapering distally. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. A few setae arise from the abdomen. The insertion areas are obscured by Verlumung.
- (55)
- Specimen 6691 (NHMD 115214 spec 4) is only accessible from the ventral side of the body (Figure 14 and Figure 15I,J). It is preserved in the same amber piece as specimens 6688–6690. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The body outline is well accessible, except for the head with appendages. The head is largely obscured by Verlumung. Only the distal elements of the antennae and labial palps are visible. Stylets are entirely obscured. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 15J), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. The thorax is subdivided into pro-, meso-, and metathorax. Abdomen units with several setae. They seem to have their insertion areas in the middle of each abdomen unit.
- (56)
- Specimen 6692 (NHMD 115214 spec 5) is only accessible from the ventral side of the body (Figure 14 and Figure 15K,L). It is preserved in the same amber piece as specimens 6688–6691. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The amber is very clear, hence the overall body outline of the specimen is accessible, except for parts of the head with appendages that are obscured by Verlumung. Only the distal elements of one antenna and both labial palps are visible. Stylets are entirely obscured. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 15L). Tarsus distally with a pair of claws. Abdomen units with several setae. They seem to have their insertion areas in the middle of each abdomen unit.
- (57)
- Specimen 6693 (NHMD 115214 spec 6) is only accessible from the ventral side of the body (Figure 14 and Figure 15N). It is preserved in the same amber piece as specimens 6688–6692. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The anterior part of the specimen is largely obscured by Verlumung, hence the overall body outline is partly difficult to access. Head with stout antennae, stylets, and labial palps (Figure 15N). Antennae subdivided into at least two elements, proximal region not accessible. Stylets and labial palps are mainly obscured by Verlumung. Only distal element of the labial palps is visible. Trunk appendages are mainly obscured. The most accessible part is the tarsus distally with a pair of claws. The abdomen shows a subdivision into several units. Abdomen with few setae.
- (58)
- Specimen 6694 (NHMD 115214 spec 7) is only accessible from the ventral side of the body (Figure 14 and Figure 15M) and sits very deep within the amber piece. Hence, the specimen is not fully in focus. It is preserved in the same amber piece as specimens 6688–6693. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The anterior part of the specimen is largely obscured by Verlumung, hence the overall body outline is partly difficult to access. Head with antennae, stylets, and labial palps (Figure 15M). Only the distal elements of the antennae, both labial palps, and the tip of the stylets are accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus) are mostly out of focus and entangled with each other, partly obscuring subdivisions and details. Thorax and setae are not accessible. The abdomen is subdivided into several units.
- (59)
- Specimen 6695 (CCHH 1124-4b spec 1) is only accessible from the lateral right side of the body (Figure 16 and Figure 17A,B). The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The specimen is a bit curved dorsally, basically adopting a U-shaped posture. Overall body outline well accessible. Head with antennae and stylets. Antennae are elongated and subdivided into three elements. Stylets are curved and tapering distally. Labial palps are not accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 17B). Coxa is tubular and elongated; trochanter not accessible; femur similar to coxa, but shorter; tibia similar, but shorter than femur; tarsus similar and taper distally with a pair of claws at the end. The thorax is subdivided into pro-, meso-, and metathorax. Numerous elongate setae arise from the abdomen. The insertion areas seem to be in the middle of each abdomen unit.
- (60)
- Specimen 6696 (CCHH 1124-4b spec 2) is only accessible from the lateral left side of the body (Figure 16 and Figure 17C,D). It is preserved in the same amber piece as specimen 6695. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The specimen is extremely curved ventrally, basically adopting a U-shaped posture. Overall body outline well accessible. Head with antennae, stylets, and labial palps. Antennae are elongated and subdivided into three elements. Stylets are not well accessible, because of the position of the head, but tapering distally. The labial palps are mostly covered by the stylets. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 17D) are entangled with each other, partly obscuring subdivisions and details. The thorax is subdivided into pro-, meso-, and metathorax, with a short cervix anteriorly. Numerous elongate setae arise from the trunk. They seem to be longer at the trunk end.
- (61)
- Specimen 6697 (CCHH 1124-4b spec 3) is only accessible from the dorsal side of the body (Figure 16 and Figure 17E,F). It is preserved in the same amber piece as specimens 6695 and 6696. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. Despite several inclusions, the overall body outline is well accessible, except for the head appendages. Head with elongate antennae and stylets. Subdivision of the antennae are not accessible. Stylets proximally straight, but sharply curved more distally and tapering at the end. Labial palps and trunk appendages are not accessible. The thorax is subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. Few elongate setae arise from the abdomen.
- (62)
- Specimen 6698 (CCHH 1124-4b spec 4) is only accessible from the dorsal side of the body (Figure 16 and Figure 17G) and sits very deep within the amber piece. Hence, the specimen is not fully in focus. As the amber piece contains multiple inclusions it is not possible to grind it further down. It is preserved in the same amber piece as specimens 6695–6697. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. Overall body outline accessible. Head with elongate antennae and stylets. Stylets proximally straight, but sharply curved more distally and tapering at the end. Labial palps and trunk appendages are not well accessible. The thorax is subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. Several setae arise from the trunk.
- (63)
- Specimen 6699 (CCHH 1124-4b spec 5) is only accessible from the lateral right side of the body (Figure 16 and Figure 17H) and sits very deep within the amber piece. Hence, the specimen is not fully in focus. As the amber piece contains multiple inclusions it is not possible to grind it further down. It is preserved in the same amber piece as specimens 6695–6698. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. Overall outline is difficult to access. The slightly better accessible parts of the specimen are the abdomen with several setae and the trunk appendages distally with a pair of claws.
- (64)
- Specimen 6700 (CCHH 1124-4b spec 6) is only accessible from the lateral right side of the body (Figure 16 and Figure 17I) and sits very deep within the amber piece. Hence, the specimen is out of focus. As the amber piece contains multiple inclusions it is not possible to grind it further down. It is preserved in the same amber piece as specimens 6695–6699. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. Overall outline is difficult to access. The slightly better accessible parts of the specimen are the trunk appendages and the abdomen with several setae.
- (65)
- Specimen 6701 (CCHH 1270-4 spec 1) is only accessible from the lateral left side of the body (Figure 18 and Figure 19A–C). The specimen shows the overall slender morphology known for older larvae of Nevrorthidae (Figure 19A,B). Overall outline is well accessible, yet a few details of the anterior part of the abdomen are obscured by a large bubble, and from the sixth segment of the abdomen the posterior part of the specimen is missing. The entire specimen, except for the head and parts of the trunk appendages, appears covered with pyrite. Head with antennae, stylets, and labial palps. Antenna is elongated and shows subdivision into numerous elements (about 18). Stylets taper distally; they are straight and broad proximally, but sharply curved more distally. Labial palps are shorter than antennae, subdivided into three elements (Figure 19C). Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 19B), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. Well preserved are the pairs of claws distally at the tarsi. The thorax is subdivided into pro-, meso-, and metathorax with a longer cervix anteriorly. Prominent setae arise from the appendage elements, exact arrangement is difficult to assess. Trunk segments with prominent long setae; insertion areas obscured.
- (66)
- Specimen 6702 (CCHH 1270-4 spec 2) is well accessible from the lateral left and right side (Figure 18 and Figure 20A–C) and less well from the dorsal side of the body (Figure 20D,E). It is preserved in the same amber piece as specimen 6701. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae. Overall body outline well accessible. Head with antennae, stylets, and labial palps. Antenna is elongated and shows subdivision into numerous elements (about 13). Stylets are curved and taper distally. Labial palps are slightly shorter than antennae, subdivided into four elements (Figure 20B). Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 20B). Coxa is tubular and elongated; trochanter not accessible; femur similar to coxa, but shorter; tibia similar to femur, but thinner; tarsus quite slender again and tapering distally with a pair of claws. The thorax is subdivided into pro-, meso-, and metathorax with a longer cervix anteriorly. Abdomen with at least eight units. The cervix and prothorax show a longitudinal groove (Figure 20E). The two thorax segments posterior to the prothorax with a distinct pattern of dorsal sclerotisations. After the anterior third of the segment, a distinct fold is apparent, subdividing the dorsal region of each segment into two distinct subregions (Figure 20B). Prominent setae arise from the trunk appendage elements. Femur and tibia are distally with a set of three setae each. Few, less prominent setae also apparent further proximally, no clear pattern apparent. Also, a few setae arise from the trunk; insertion areas are obscured.
- (67)
- Specimen 6703 (CCHH 1387-1) is well accessible from the ventral, dorsal, and lateral right side (Figure 21A–D). The specimen shows the overall slender morphology known for older larvae of Nevrorthidae. Overall body outline well accessible. Head with antennae, stylets, and labial palps. Antenna is elongated and shows subdivision into numerous elements (about 15; Figure 21E). Stylets taper distally; they are straight proximally, but sharply curved more distally. Labial palps are shorter than antennae, subdivided into four elements (Figure 21A). Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 21A,B). Coxa is tubular and thickened; trochanter tubular, elongated, and short; femur similar to trochanter, but longer and thicker; tibia similar to femur, but shorter and thinner; tarsus quite slender and tapering distally with a pair of claws. The thorax is subdivided into pro-, meso-, and metathorax with a longer cervix anteriorly. The cervix and prothorax have a longitudinal groove (Figure 21E). The trunk segments posterior to the prothorax show a distinct pattern of dorsal sclerotisation. After the anterior third of the segment, a distinct fold is apparent, subdividing the dorsal region of each segment into two distinct subregions (Figure 21B,C). Prominent setae arise from the trunk appendage elements. Femur and tibia are distally with a set of three setae each. Trunk segments with prominent long setae. These arise close to the posterior end of each segment and appear to form a kind of ring around the segment. At the abdomen units appear to be six setae dorsally and four ventrally.
- (68)
- Specimen 6704 (SMF Be 2192) is only accessible from the lateral left side of the body (Figure 22G) and sits very deep within the amber piece. Hence, the specimen is not fully in focus. As the amber piece contains multiple inclusions it is not possible to grind it further down. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The overall body outline is accessible. Head with antennae, stylets, and labial palps. Further details of the head appendages are mainly obscured. Stylets taper distally. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 22G) are entangled with each other, partly obscuring subdivisions and details. The thorax is subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. Several setae arise from the trunk.
- (69)
- Specimen 6705 (PED 0792) is accessible from the lateral right and left side (Figure 22A–D) and sits very deep within the amber piece. As the amber piece contains multiple inclusions it is not possible to grind it further down. Additionally, the head is accessible from the dorsal side (Figure 22B). The specimen shows the overall slender morphology known for older larvae of Nevrorthidae. The overall body outline is accessible, yet most details, especially of the trunk are obscured by Verlumung and numerous bubbles. Head with antennae, stylets, and labial palps. Antenna is elongated and shows subdivision into numerous elements (about 11; Figure 22C). Stylets taper distally; they are straight proximally, but sharply curved more distally. Labial palps are slightly shorter than antennae, subdivided into at least four elements (Figure 21A), proximal region is not accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 22D). Proximal region with coxa and trochanter mainly obscured by Verlumung. Femur tubular and elongated; tibia similar to femur, but shorter; tarsus quite slender and, but longer than tibia, tapering distally with a pair of claws. Prominent setae arise from the trunk appendage elements. Femur and tibia are distally with a set of three setae each. Also, several setae arise from the trunk; insertions are obscured.
- (70)
- Specimen 6706 (PED 1379) is well accessible from the ventral and dorsal side (Figure 23A–C). The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the segments of the abdomen are extremely short. The amber is very clear and the overall outline is well accessible. Head with short and stout antennae, stylets, and labial palps. Antennae are subdivided into three elements. Stylets are curved and tapering distally. Labial palps are subdivided into four elements (Figure 23B). Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus), well preserved, but they are entangled with each other, partly obscuring subdivisions and details. Coxa tubular thickened and elongated; trochanter not accessible; femur similar to coxa, but shorter; tibia similar to femur, but shorter again; tarsus similar to tibia, but a bit longer and tapering distally with a pair of claws. The thorax is subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. Abdomen with 10 units. Prominent setae arise from the trunk appendage elements and several elongate setae arise from the trunk.
3.4. Additional Fossil Larval Representatives of Nevrorthidae in Kachin amber
- (71)
- Specimen 6707 (BUB 3703) is only accessible from the dorsal side of the body (Figure 24A–C). The specimen shows the overall slender morphology known for older larvae of Nevrorthidae. The overall body outline is partly accessible, except for the labial palps, the trunk appendages, and the thorax subdivisions. From the head, only antennae and stylets are accessible. Antennae are short and elongated. Stylets are curved and tapering distally. Several elongate setae arise from the abdomen; insertion areas are obscured.
- (72)
- Specimen 6708 (PED 0259) is only accessible from the dorsal side of the body (Figure 25A,B). The specimen shows the overall slender morphology known for older larvae of Nevrorthidae. Of this specimen, only the head with appendages, the cervix, and prothorax are preserved. Head with antennae, stylets, and labial palps. Antenna is elongated and shows subdivision into numerous elements (about 21; Figure 25A). Stylets taper distally; they are straight proximally, but sharply curved more distally. Labial palps are slightly shorter than antennae, subdivided into four elements (Figure 25B). The long cervix and prothorax show a longitudinal groove (Figure 25A). A few elongated setae arise from the trunk.
- (73)
- Specimen 6709 (PED 0327) is only accessible from the dorsal side of the body (Figure 25C,D). The specimen shows the overall slender morphology known for older larvae of Nevrorthidae. Of this specimen, the entire abdomen is missing. Head with antennae, stylets, and labial palps. Antenna is elongated and subdivided into at least two elements, proximal region is not accessible. The same applies to the labial palps, but there are three elements apparent. Stylets taper distally; they are straight proximally, but curved more distally. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus). Coxa and trochanter are not accessible from the dorsal side of the body. Femur tubular and elongated; tibia similar to femur, but shorter; tarsus quite slender and tapering distally with a pair of claws. The thorax is subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. A few elongated setae arise from the trunk.
- (74)
- Specimen 6710 (PED 0632) is accessible from the ventral and dorsal side (Figure 26A–C). The specimen shows the overall slender morphology known for older larvae of Nevrorthidae. Of this specimen, the entire abdomen is missing and the posterior part of the thorax is mainly destroyed and obscured by Verlumung. Head with antennae, stylets, and labial palps. Antenna is elongated and shows subdivision into numerous elements (about 12; Figure 26B). Stylets taper distally; they are straight proximally, but sharply curved more distally. Labial palps are slightly shorter than antennae, subdivided into four elements (Figure 26B). Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus), but mostly destroyed or obscured. Most accessible are the tarsi distal with a pair of claws. Subdivision of the thorax is obscured by Verlumung. This specimen has a long cervix and several elongate setae arise from the trunk.
- (75)
- Specimen 6711 (PED 0663) is only accessible from the ventral side of the body (Figure 26D,E) and sits very deep within the amber piece. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae. The most accessible parts of the specimen are the head with appendages and the long cervix. Head with antennae, stylets, and labial palps. Antenna is elongated and shows subdivision into numerous elements (Figure 26E). Stylets taper distally; they are straight proximally, but sharply curved more distally. Labial palps are slightly longer than antennae, subdivided into four elements (Figure 26E). Trunk appendages are mostly obscured, as well as the subdivisions of the thorax and anterior part of the abdomen. Few setae arise from the trunk insertion areas obscured.
- (76)
- Specimen 6712 (PED 1338) is only accessible from the dorsal side of the body (Figure 27A–C) and sits very deep within the amber piece. As the amber piece contains multiple inclusions it is not possible to grind it further down. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae. The overall body outline is accessible. Most accessible parts of the specimen are the head with appendages and the short cervix. Head with antennae, stylets, and labial palps. Antenna is elongated and shows subdivision into numerous elements (Figure 27D). Stylets are curved and taper distally. Only the distal two elements of the labial palps are apparent, the proximal region is obscured by the stylets. Despite the ventral view of the specimen the trunk appendages are not well accessible. Subdivision of the thorax is obscured by other inclusions. Trunk with few elongate setae.
- (77)
- Specimen 6713 (PED 2001) is only accessible from the dorsal side of the body (Figure 22E,F) and sits very deep within the amber piece. As the amber piece contains multiple inclusions it is not possible to grind it further down. The better accessible part of the specimen is the head with appendages. The rest of the body is obscured by a large black inclusion. Head with antennae, stylets, and labial palps. Antennae and labial palps are mainly obscured by inclusions. The stylets taper distally; they are straight proximally, but curved more distally. Only few fragments of the trunk appendages with a few setae are visible.
- (78)
- Specimen 6714 (PED 2447) is accessible from the dorsal and ventral sides of the body (Figure 28A–D). The specimen shows the overall slender morphology known for older larvae of Nevrorthidae. Overall body outline well accessible, except for the trunk appendages and details of the thorax, which are obscured by various inclusions. Head with antennae, stylets, and labial palps. Antenna is elongated and shows subdivision into numerous elements (about 11, Figure 28B). Stylets taper distally; they are straight proximally, but curved more distally. Only the distal two elements of the labial palps are apparent, the proximal region is obscured by the stylets. Trunk segments with prominent long setae. These arise close to the posterior end of each segment and appear to form a kind of ring around the segment.
- (79)
- Specimen 6715 (PED 2622) is accessible from the dorsal side of the body (Figure 29A,B) and less well from the ventral side (Figure 29C). The specimen shows the overall slender morphology known for older larvae of Nevrorthidae in the anterior region, but especially the posterior trunk appears much less slender, the abdomen is extremely short. Of this specimen, the entire abdomen and the trunk appendages are deformed. Head with antennae, stylets, and labial palps. Antenna is elongated and shows subdivision into numerous elements (about 19, Figure 29A). Stylets taper distally; they are straight proximally, but sharply curved more distally. Labial palps are subdivided into at least three elements apparent, proximal region not accessible. The thorax is subdivided into pro-, meso-, and metathorax with a longer cervix anteriorly. Few elongated setae arise from the trunk.
- (80)
- Specimen 6716 (PED 2662) is accessible from the ventral and dorsal sides (Figure 23D,E). The specimen shows the overall slender morphology known for older larvae of Nevrorthidae. The overall outline is only accessible until the anterior part of the abdomen. The rest of the body is missing. Head with antennae, stylets, and labial palps. Subdivisions of the antennae are obscured. Stylets taper distally; they are straight proximally, but curved more distally. Labial palps are subdivided into four elements (Figure 23D). Trunk appendages are not well accessible and entangled with each other. Several setae arise from the trunk.
- (81)
- Specimen 6717 (PED 2669) is accessible from the lateral right side of the body (Figure 12H) and the dorsal side of the head (Figure 12G). The slightly more accessible part is the head and parts of the trunk with their appendages. Other aspects are not well accessible. The remaining parts of the body are mostly obscured by various inclusions. Nevertheless, the specimen shows the overall slender morphology known for older larvae of Nevrorthidae. At the head are only the stylets and labial palps left. The stylets are curved and tapering distally. Labial palps seem to be longer than the stylets. Subdivisions are mainly obscured. A few setae arise from the body.
- (82)
- Specimen 6718 (PED 2744) is only accessible from the dorsal side of the body (Figure 30A–C). The posterior body is missing. The specimen shows the overall slender morphology known for older larvae of Nevrorthidae. Outline of the head with appendages and anterior part of the thorax is well accessible. Head with antennae, stylets, and labial palps. Antenna is elongated and shows subdivision into numerous elements (about 19, Figure 30B). Stylets are curved and taper distally. Labial palps are subdivided into at least four elements apparent, proximal region not accessible. Few setae arise from the body.
- (83)
- Specimen 6719 (PED 2786) is well accessible from the dorsal side of the body (Figure 31A,B) and less well from the ventral side (Figure 31C). The specimen shows the overall slender morphology known for older larvae of Nevrorthidae. The abdomen of this specimen is quite slender and deformed at the end. Overall body outline is well accessible. Head with antennae, stylets, and labial palps. Antenna is elongated and shows subdivision into numerous elements (about 13, Figure 31B). Stylets are curved and taper distally. Labial palps are subdivided into at least four elements apparent. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 31B,D). Coxa and trochanter are not accessible in this view. Femur quite slender, elongated, and tubular; tibia similar, but shorter; tarsus similar to tibia, but shorter and thinner again, tapering distally with a pair of claws. The thorax is subdivided into pro-, meso-, and metathorax with a longer cervix anteriorly. Prominent setae arise from the trunk appendage elements. Femur and tibia are distally with a set of four or five setae each. Few, less prominent setae are also apparent further proximally, no clear pattern apparent. Trunk segments with prominent long setae. These arise close to the posterior end of each segment and appear to form a kind of ring around the segment.
- (84)
- Specimen 6720 (Weiterschan BuB 24) is only accessible from lateral right side of the body (Figure 32A–C). The specimen shows the overall slender morphology known for older larvae of Nevrorthidae. Overall body outline is well accessible. Head with antennae, stylets, and labial palps. Antenna is elongated and shows subdivision into numerous elements (about 14, Figure 31B). Stylets taper distally; they are straight proximally, but slightly curved more distally. Labial palps are subdivided into at least three elements apparent, proximal region is not accessible. Trunk appendages with five major elements (coxa, trochanter, femur, tibia, tarsus; Figure 32B,D). Coxa is tubular and elongated; trochanter not accessible in this view; femur quite slender and longer than coxa; tibia similar to femur, but shorter; tarsus similar to tibia, but longer and thinner again, tapering distally with a pair of claws. The thorax is subdivided into pro-, meso-, and metathorax with a short cervix anteriorly. Prominent setae arise from the trunk appendage elements. Femur and tibia are distally with a set of three or four setae each. Trunk segments with a few setae.
3.5. Shape Analysis
4. Discussion
4.1. Identity of the New Specimens
4.2. The Assumed Stasis of Dragon Lacewing Larvae
4.3. Loss of Diversity in Other Groups of Lacewings
4.4. A Brief Look on Palaeoecology
5. Outlook
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef]
- Lister, B.C.; Garcia, A. Climate-driven declines in arthropod abundance restructure a rainforest food web. Proc. Natl. Acad. Sci. USA 2018, 115, E10397–E10406. [Google Scholar] [CrossRef] [PubMed]
- Seibold, S.; Gossner, M.M.; Simons, N.K.; Blüthgen, N.; Müller, J.; Ambarlı, D.; Ammer, C.; Bauhus, J.; Fischer, M.; Habel, J.C.; et al. Arthropod decline in grasslands and forests is associated with landscape-level drivers. Nature 2019, 574, 671–674. [Google Scholar] [CrossRef] [PubMed]
- Jähnig, S.C.; Baranov, V.; Altermatt, F.; Cranston, P.; Friedrichs-Manthey, M.; Geist, J.; He, F.; Heino, J.; Hering, D.; Hölker, F.; et al. Revisiting global trends in freshwater insect biodiversity. Wiley Interdisc. Rev. Water 2021, 8, e1506. [Google Scholar] [CrossRef]
- Van Klink, R.; Bowler, D.E.; Gongalsky, K.B.; Swengel, A.B.; Gentile, A.; Chase, J.M. Meta-analysis reveals declines in terrestrial but increases in freshwater insect abundances. Science 2020, 368, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Van Klink, R.; Bowler, D.E.; Gongalsky, K.B.; Chase, J.M. Long-term abundance trends of insect taxa are only weakly correlated. Biol. Lett. 2022, 18, 20210554. [Google Scholar] [CrossRef]
- Haug, C.; Braig, F.; Haug, J.T. Quantitative analysis of lacewing larvae over more than 100 million years reveals a complex pattern of loss of morphological diversity. Sci. Rep. 2023, 13, 6127. [Google Scholar] [CrossRef]
- MacLeod, E.G. The Neuroptera of the Baltic Amber. I. Ascalaphidae, Nymphidae, and Psychopsidae. Psyche 1970, 77, 045459. [Google Scholar] [CrossRef]
- Perrichot, V. Environnements Paraliques à Ambre et à Végétaux du Crétacé Nord-Aquitain (Charentes, Sud-Ouest de la France). Ph.D. Thesis, Université Rennes: Rennes, France, 2003; Unpublished. [Google Scholar]
- Engel, M.S.; Grimaldi, D.A. Diverse Neuropterida in Cretaceous amber, with particular reference to the paleofauna of Myanmar (Insecta). Nova Suppl. Entomol. 2008, 20, 1–86. [Google Scholar]
- Perrichot, V.; Neraudeau, D.; Tafforeau, P. Charentese amber. In Biodiversity of Fossils in Amber from the Major World Deposits; Penney, D., Ed.; Siri Scientific Press: Manchester, UK, 2010; pp. 192–207. [Google Scholar]
- Pérez-de la Fuente, R.; Delclòs, X.; Peñalver, E.; Speranza, M.; Wierzchos, J.; Ascaso, C.; Engel, M.S. Early evolution and ecology of camouflage in insects. Proc. Natl. Acad. Sci. USA 2012, 109, 21414–21419. [Google Scholar] [CrossRef]
- Pérez-de la Fuente, R.; Delclòs, X.; Peñalver, E.; Engel, M.S. A defensive behavior and plant-insect interaction in Early Cretaceous amber–the case of the immature lacewing Hallucinochrysa diogenesi. Arthropod Struct. Dev. 2016, 45, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Pérez-de la Fuente, R.; Peñalver, E.; Azar, D.; Engel, M.S. A soil-carrying lacewing larva in Early Cretaceous Lebanese amber. Sci. Rep. 2018, 8, 16663. [Google Scholar] [CrossRef] [PubMed]
- Pérez-de la Fuente, R.; Engel, M.S.; Azar, D.; Peñalver, E. The hatching mechanism of 130-million-year-old insects: An association of neonates, egg shells and egg bursters in Lebanese amber. Palaeontology 2019, 62, 547–559. [Google Scholar] [CrossRef]
- Pérez-de la Fuente, R.; Engel, M.S.; Delclòs, X.; Peñalver, E. Straight-jawed lacewing larvae (Neuroptera) from Lower Cretaceous Spanish amber, with an account on the known amber diversity of neuropterid immatures. Cretac. Res. 2020, 106, 104200. [Google Scholar] [CrossRef]
- Wedmann, S.; Makarkin, V.N.; Weiterschan, T.; Hörnschemeyer, T. First fossil larvae of Berothidae (Neuroptera) from Baltic amber, with notes on the biology and termitophily of the family. Zootaxa 2013, 3716, 236–258. [Google Scholar] [CrossRef] [PubMed]
- Xia, F.; Yang, G.; Zhang, Q.; Shi, G.; Wang, B. Amber: Life Through Time and Space; Science Press: Beijing, China, 2015; p. 197. [Google Scholar]
- Engel, M.S. Two new genera of Cretaceous dustywings in amber from northern Myanmar (Neuroptera: Coniopterygidae). Novit. Paleoentomol. 2016, 17, 1–16. [Google Scholar] [CrossRef]
- Wang, B.; Xia, F.; Engel, M.S.; Perrichot, V.; Shi, G.; Zhang, H.; Chen, J.; Jarzembowski, E.A.; Wappler, T.; Rust, J. Debris-carrying camouflage among diverse lineages of Cretaceous insects. Sci. Adv. 2016, 2, e1501918. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, W.; Winterton, S.L.; Breitkreuz, L.C.; Engel, M.S. Early morphological specialization for insect-spider associations in Mesozoic lacewings. Curr. Biol. 2016, 26, 1590–1594. [Google Scholar] [CrossRef]
- Liu, X.; Shi, G.; Xia, F.; Lu, X.; Wang, B.; Engel, M.S. Liverwort mimesis in a Cretaceous lacewing larva. Curr. Biol. 2018, 28, 1475–1481.e1. [Google Scholar] [CrossRef]
- Zhang, W.W. Frozen Dimensions. The Fossil Insects and Other Invertebrates in Amber; Chongqing University Press: Chongqing, China, 2017. [Google Scholar]
- Badano, D.; Engel, M.S.; Basso, A.; Wang, B.; Cerretti, P. Diverse Cretaceous larvae reveal the evolutionary and behavioural history of antlions and lacewings. Nature Comm. 2018, 9, 3257. [Google Scholar] [CrossRef]
- Makarkin, V.N. Re-description of Grammapsychops lebedevi Martynova, 1954 (Neuroptera: Psychopsidae) with notes on the Late Cretaceous psychopsoids. Zootaxa 2018, 4524, 581–594. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Liu, H.; Jarzembowski, E.A. High morphological disparity of neuropteran larvae during the Cretaceous revealed by a new large species. Geol. Mag. 2022, 159, 954–962. [Google Scholar] [CrossRef]
- MacLeod, E.G. A Comparative Morphological Study of the Head Capsule and Cervix of Larval Neuroptera (Insecta). Ph.D. Thesis, Harvard University, Cambridge, MA, USA, 1964. [Google Scholar]
- Zimmermann, D.; Randolf, S.; Aspöck, U. From chewing to sucking via phylogeny–From sucking to chewing via ontogeny: Mouthparts of Neuroptera. In Insect Mouthparts, Zoological Monographs 5; Krenn, H.W., Ed.; Springer: Berlin, Germany, 2019; pp. 361–385. [Google Scholar]
- Haug, G.T.; Haug, C.; van der Wal, S.; Müller, P.; Haug, J.T. Split-footed lacewings declined over time: Indications from the morphological diversity of their antlion-like larvae. PalZ 2022, 96, 29–50. [Google Scholar] [CrossRef]
- Haug, C.; Posada Zuluaga, V.; Zippel, A.; Braig, F.; Müller, P.; Gröhn, C.; Weiterschan, T.; Wunderlich, J.; Haug, G.T.; Haug, J.T. The morphological diversity of antlion larvae and their closest relatives over 100 million years. Insects 2022, 13, 587. [Google Scholar] [CrossRef]
- Hassenbach, C.; Buchner, L.; Haug, G.T.; Haug, C.; Haug, J.T. An expanded view on the morphological diversity of long-nosed antlion larvae further supports a decline of silky lacewings in the past 100 million years. Insects 2023, 14, 170. [Google Scholar] [CrossRef]
- Aspöck, U.; Aspöck, H. Kamelhälse, Schlammfliegen, Ameisenlöwen. Wer sind sie? (Insecta: Neuropterida: Raphidioptera, Megaloptera, Neuroptera). Stapfia 1999, 60, 1–34. [Google Scholar]
- Aspöck, U.; Aspöck, H. Phylogenetic relevance of the genital sclerites of Neuropterida (Insecta: Holometabola). Syst. Entomol. 2008, 33, 97–127. [Google Scholar] [CrossRef]
- Aspöck, U. Phylogeny of the Neuropterida (Insecta: Holometabola). Zool. Scr. 2002, 31, 51–55. [Google Scholar] [CrossRef]
- Aspöck, U.; Plant, J.D.; Nemeschkal, H.L. Cladistic analysis of Neuroptera and their systematic position within Neuropterida (Insecta: Holometabola: Neuropterida: Neuroptera). Syst. Entomol. 2001, 26, 73–86. [Google Scholar] [CrossRef]
- Aspöck, U.; Haring, E.; Aspöck, H. The phylogeny of the Neuropterida: Long lasting and current controversies and challenges (Insecta: Endopterygota). Arthropod Syst. Phyl. 2012, 70, 119–129. [Google Scholar] [CrossRef]
- Haring, E.; Aspöck, U. Phylogeny of the Neuropterida: A first molecular approach. Syst. Entomol. 2004, 29, 415–430. [Google Scholar] [CrossRef]
- Winterton, S.L.; Hardy, N.B.; Wiegmann, B.M. On wings of lace: Phylogeny and Bayesian divergence time estimates of Neuropterida (Insecta) based on morphological and molecular data. Syst. Entomol. 2010, 35, 349–378. [Google Scholar] [CrossRef]
- Winterton, S.L.; Lemmon, A.R.; Gillung, J.P.; Garzon, I.J.; Badano, D.; Bakkes, D.K.; Breitkreuz, L.C.V.; Engel, M.S.; Lemmon, E.M.; Liu, X. Evolution of lacewings and allied orders using anchored phylogenomics (Neuroptera, Megaloptera, Raphidioptera). Syst. Entomol. 2018, 43, 330–354. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.; Garzón-Orduña, I.J.; Winterton, S.L.; Yan, Y.; Aspöck, U.; Aspöck, H.; Yang, D. Mitochondrial phylogenomics illuminates the evolutionary history of Neuropterida. Cladistics 2017, 33, 617–636. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.S.; Winterton, S.L.; Breitkreuz, L.C.V. Phylogeny and evolution of Neuropterida: Where have wings of lace taken us? Ann. Rev. Entomol. 2018, 63, 531–551. [Google Scholar] [CrossRef] [PubMed]
- Vasilikopoulos, A.; Misof, B.; Meusemann, K.; Lieberz, D.; Flouri, T.; Beutel, R.G.; Niehuis, O.; Wappler, T.; Rust, J.; Peters, R.S.; et al. An integrative phylogenomic approach to elucidate the evolutionary history and divergence times of Neuropterida (Insecta: Holometabola). BMC Evol. Biol. 2020, 20, 64. [Google Scholar]
- Du, X.; Niu, K.; Bao, T. Giant Jurassic dragon lacewing larvae with lacustrine palaeoecology represent the oldest fossil record of larval neuropterans. Proc. R. Soc. B 2023, 290, 20222500. [Google Scholar] [CrossRef]
- Baranov, V.; Hoffeins, C.; Hoffeins, H.W.; Haug, J.T. Reaching across the ocean of time: A midge morphotype from the Cretaceous of Gondwana found in the Eocene Baltic amber. Palaeontol. Electron. 2019, 22, art.38A. [Google Scholar] [CrossRef]
- Makarkin, V.N.; Perkovsky, E.E. Rophalis relicta HAGEN (Neuroptera, Nevrorthidae) in the late Eocene Rovno amber, with a discussion of the taxonomic status of the genus. Denisia 2009, 26, 137–144. [Google Scholar]
- Wichard, W. Family Nevrorthidae (Insecta, Neuroptera) in mid-Cretaceous Burmese amber. Palaeodiv 2017, 10, 1–6. [Google Scholar] [CrossRef]
- Wichard, W.; Gröhn, C.; Seredszus, F. Aquatic Insects in Baltic Amber; Kessel: Remagen, Germany, 2009; p. 336. [Google Scholar]
- Aspöck, U.; Aspöck, H.; Liu, X. The Nevrorthidae, mistaken at all times: Phylogeny and review of present knowledge (Holometabola, Neuropterida, Neuroptera). Dt. Entomol. Z. 2017, 64, 77–110. [Google Scholar] [CrossRef][Green Version]
- Wichard, W. Overview and descriptions of Nevrorthidae in Baltic amber (Insecta, Neuroptera). Palaeodiv 2016, 9, 95–111. [Google Scholar] [CrossRef][Green Version]
- Haug, J.T.; Baranov, V.; Schädel, M.; Müller, P.; Gröhn, C.; Haug, C. Challenges for understanding lacewings: How to deal with the incomplete data from extant and fossil larvae of Nevrorthidae? (Neuroptera). Fragm. Entomol. 2020, 52, 137–167. [Google Scholar] [CrossRef]
- Takahashi, R. Uncommon neuropterous larva. Dobutsugaku Zasshi 1942, 54, 439–441. [Google Scholar]
- Zwick, P. Beschreibung der aquatischen Larve von Neurorthus fallax (Rambur) und Errichtung der neuen Planipennierfamilie Neurorthidae fam. nov. Gewässer Abwässer 1967, 44, 65–86. [Google Scholar]
- Riek, E.F. Neuroptera (lacewings). In The Insects of Australia; Melbourne University Press: Melbourne, VIC, Australia, 1970; pp. 472–494. [Google Scholar]
- Gepp, J. Erforschungsstand der Neuropteren. Larven der Erde (mit einem Schlüssel zur Larvaldiagnose der Familien, einer Übersicht von 340 beschriebenen Larven und 600 Literaturzitaten). In Progress in Worlds Neuropterology. In Proceedings of the First International Symposium on Neuropterology, Graz, Austria, 22–26 September 1980; pp. 183–239. [Google Scholar]
- Malicky, H. Ein Beitrag zur Autökologie und Bionomie der aquatischen Netzflüglergattung Neurorthus (Insecta, Neuroptera, Neurorthidae). Archiv Hydrobiol. 1984, 101, 231–246. [Google Scholar]
- New, T.R. Planipennia, Lacewings. Handbuch der Zoologie, Vol. 4 (Arthropoda: Insecta), Part 30; Walter de Gruyter: Berlin, Germany, 1989; p. 132. [Google Scholar]
- New, T.R. Neuroptera (lacewings). In The Insects of Australia, 2nd ed.; Naumann, I.D., Ed.; Melbourne University Press: Melbourne, VIC, Australia, 1991; Volume 1, pp. 525–542. [Google Scholar]
- New, T.R. The lacewings (Insecta, Neuroptera) of Tasmania. Pap. Proc. R. Soc. Tasman. 1992, 126, 29–45. [Google Scholar] [CrossRef]
- New, T.R. Insecta: Neuropteroidea. In Freshwater Invertebrates of the Malaysian Region; Yule, C.M., Yong, H.S., Eds.; Academy of Sciences Malaysia: Kualalumpur, Malaysia, 2004; pp. 491–500. [Google Scholar]
- Wichard, W.; Arens, W.; Eisenbeis, G.; Schwoerbel, J. Atlas zur Biologie der Wasserinsekten; Gustav Fischer: Stuttgart, Germany, 1995; p. 339. [Google Scholar]
- Weitschat, W.; Wichard, W. Atlas der Pflanzen und Tiere im Baltischen Bernstein; Friedrich Pfeil: Munich, Germany, 1998. [Google Scholar]
- Weitschat, W.; Wichard, W. Atlas of Plants and Animals in Baltic Amber; Friedrich Pfeil: Munich, Germany, 2002; p. 256. [Google Scholar]
- Tauber, C.A.; Tauber, M.J.; Albuquerque, G.S. Neuroptera (lacewings, antlions). In Encyclopedia of Insects; Resh, V.H., Cardé, R.T., Eds.; Academic Press: Cambridge, MA, USA, 2003; pp. 695–707. [Google Scholar]
- Grimaldi, D.; Engel, M.S. Evolution of the Insects; Cambridge University Press: Cambridge, UK, 2005; p. 755. [Google Scholar]
- Aspöck, U.; Aspöck, H. Verbliebene Vielfalt vergangener Blüte. Zur Evolution, Phylogenie und Biodiversität der Neuropterida (Insecta: Endopterygota). Denisia 2007, 20, 451–516. [Google Scholar]
- Aspöck, U.; Aspöck, H. Erobern Nevrorthidae Mitteleuropa? Eine biogeographische Provokation (Neuroptera: Neuropterida). Denisia 2010, 29, 25–35. [Google Scholar]
- Beutel, R.G.; Friedrich, F.; Aspöck, U. The larval head of Nevrorthidae and the phylogeny of Neuroptera (Insecta). Zool. J. Linn. Soc. 2010, 158, 533–562. [Google Scholar] [CrossRef]
- Gavira, O.; Sánchez, S.; Carrasco, P.; Ripoll, J.; Solís, S. Presence of the family Nevrorthidae (Neuroptera) in the Iberian Peninsula. Bol. Soc. Entomol. Aragon. 2012, 51, 217–220. [Google Scholar]
- Monserrat, V.J.; Gavira, O. A new European species of Nevrorthus in the Iberian Peninsula (Insecta, Neuropterida). Zootaxa 2014, 3796, 349–360. [Google Scholar] [CrossRef]
- Markovič, V.; Ilič, M.; Rakovič, M.; Paunovič, M. New Record of Nevrorthidae (Neuroptera; Neuropterida) from Serbia. SCUP 2016, 1, 163–168. [Google Scholar]
- Wichard, W.; Weitschat, W. Wasserinsekten im Bernstein: Eine paläobiologische Studie (Vol. 4); Löbbecke-Museum und Aquazoo: Düsseldorf, Germany, 1996; p. 121. [Google Scholar]
- Grimaldi, D.A.; Engel, M.S.; Nascimbene, P.C. Fossiliferous Cretaceous amber from Myanmar (Burma): Its rediscovery, biotic diversity, and paleontological significance. Amer. Mus. Nov. 2002, 3361, 1–71. [Google Scholar] [CrossRef]
- Gröhn, C. Einschlüsse im Baltischen Bernstein; Verlag Wachholtz: Kiel, Germany, 2015; p. 424. [Google Scholar]
- Lu, X.; Xia, F.; Wang, B.; Aspöck, U.; Liu, X. Taxonomic notes on Cretarophalis patrickmuelleri Wichard, 2017 (Insecta: Neuroptera: Nevrorthidae) from the mid-Cretaceous of Myanmar, and its phylogenetic significance. Zootaxa 2018, 4370, 591–600. [Google Scholar] [CrossRef]
- Cruickshank, R.D.; Ko, K. Geology of an amber locality in the Hukawng Valley, northern Myanmar. J. Asian Earth Sci. 2003, 21, 441–455. [Google Scholar] [CrossRef]
- Shi, G.; Grimaldi, D.A.; Harlow, G.E.; Wang, J.; Wang, J.; Yang, M.; Lei, W.; Li, Q.; Li, X. Age constraint on Burmese amber based on U–Pb dating of zircons. Cretac. Res. 2012, 37, 155–163. [Google Scholar] [CrossRef]
- Yu, T.; Kelly, R.; Mu, L.; Ross, A.; Kennedy, J.; Broly, P.; Xia, F.; Zhang, H.; Wang, B.; Dilcher, D. An ammonite trapped in Burmese amber. Proc. Natl. Acad. Sci. USA 2019, 116, 11345–11350. [Google Scholar] [CrossRef]
- Iwata, H.; Ukai, Y. SHAPE: A computer program package for quantitative evaluation of biological shapes based on elliptic Fourier descriptors. J. Hered. 2002, 93, 384–385. [Google Scholar] [CrossRef]
- Braig, F.; Haug, J.T.; Schädel, M.; Haug, C. A new thylacocephalan crustacean from the Upper Jurassic lithographic limestones of southern Germany and the diversity of Thylacocephala. Palaeodiv 2019, 12, 69–87. [Google Scholar] [CrossRef][Green Version]
- Janzen, J.W. Arthropods in Baltic Amber; Ampyx Verlag: Halle, Germany, 2002; p. 167. [Google Scholar]
- Scheven, J. Bernstein-Einschlüsse: Eine untergegangene Welt bezeugt die Schöpfung. Erinnerungen an die Welt vor der Sintflut; Kuratorium Lebendige Vorwelt e.V.: Hofheim, Germany, 2004; p. 160. [Google Scholar]
- Kobbert, M.J. Bernstein-Fenster in die Urzeit; Planet. Poster Editions: Göttingen, Germany, 2005; p. 244. [Google Scholar]
- Giacomino, M. Contribution à la connaissance des Neuropterida de Corse (Raphidioptera et Neuroptera). Micromus angulatus (Stephens, 1836), Microminae nouveau pour la faune de Corse (Neuroptera Hemerobiidae). L’Entomologiste 2013, 69, 117–120. [Google Scholar]
- Kobbert, M.J. Wunderwelt Bernstein: Faszinierende Fossilien in 3-D; WBG (Wissenschaftliche Buchgesellschaft): Darmstadt, Germany, 2013. [Google Scholar]
- Beutel, R.G.; Friedrich, F.; Yang, X.K.; Ge, S.Q. Insect Morphology and Phylogeny: A Textbook for Students of Entomology; Walter de Gruyter: Berlin, Germany, 2014; p. 516. [Google Scholar]
- Larsson, S.G. Baltic Amber: A Palaeobiological Study; Scandinavian Science Press: Klampenborg, Denmark, 1978. [Google Scholar]
- Haug, G.T.; Haug, C.; Pazinato, P.G.; Braig, F.; Perrichot, V.; Gröhn, C.; Müller, P.; Haug, J.T. The decline of silky lacewings and morphological diversity of long-nosed antlion larvae through time. Palaeont. Electron. 2020, 23, a39. [Google Scholar] [CrossRef]
- Badano, D.; Fratini, M.; Maugeri, L.; Palermo, F.; Pieroni, N.; Cedola, A.; Haug, J.T.; Weiterschan, T.; Velten, J.; Mei, M.; et al. X-ray microtomography and phylogenomics provide insights into the morphology and evolution of an enigmatic Mesozoic insect larva. Syst. Entomol. 2021, 46, 672–684. [Google Scholar] [CrossRef]
- Haug, J.T.; Haug, C. New details of the enigmatic 100 million years old antlion-like larvae of Ankyloleon (Myrmeleontiformia, Neuroptera). Europ. J. Taxon 2023, accepted. [Google Scholar]
- Haug, J.T.; Haug, G.T.; Zippel, A.; van der Wal, S.; Müller, P.; Gröhn, C.; Wunderlich, J.; Hoffeins, C.; Hoffeins, H.-W.; Haug, C. Changes in the morphological diversity of larvae of lance lacewings, mantis lacewings and their closer relatives over 100 million years. Insects 2021, 12, 860. [Google Scholar] [CrossRef]
- Haug, G.T.; Baranov, V.; Wizen, G.; Pazinato, P.G.; Müller, P.; Haug, C.; Haug, J.T. The morphological diversity of long-necked lacewing larvae (Neuroptera: Myrmeleontiformia). Bull. Geosci. 2021, 96, 431–457. [Google Scholar] [CrossRef]
- Haug, J.T.; van der Wal, S.; Gröhn, C.; Hoffeins, C.; Hoffeins, H.-W.; Haug, C. Diversity and fossil record of larvae of three groups of lacewings with unusual ecology and functional morphology: Ithonidae, Coniopterygidae and Sisyridae. Palaeont. Electron. 2022, 25, a14. [Google Scholar] [CrossRef]
- Haug, J.T.; Linhart, S.; Haug, G.T.; Gröhn, C.; Hoffeins, C.; Hoffeins, H.-W.; Müller, P.; Weiterschan, T.; Wunderlich, J.; Haug, C. The diversity of aphidlion-like larvae over the last 130 million years. Insects 2022, 13, 336. [Google Scholar] [CrossRef]
- Culler, L.E.; Ohba, S.-y.; Crumrine, P. Predator-prey interactions of dytiscids. In Ecology, Systematics, and the Natural History of Predaceous Diving Beetles (Coleoptera: Dytiscidae); Yee, D.A., Ed.; Springer: Dordrecht, The Netherlands, 2014; pp. 363–386. [Google Scholar]
- Haug, J.T.; Kiesmüller, C.; Haug, G.T.; Haug, C.; Hörnig, M.K. A fossil aphidlion preserved together with its prey in 40 million-year-old Baltic amber. Palaeobiodiv. Palaeoenv. 2023, 103, 155–163. [Google Scholar] [CrossRef]



































Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mengel, L.; Linhart, S.; Haug, G.T.; Weiterschan, T.; Müller, P.; Hoffeins, C.; Hoffeins, H.-W.; Baranov, V.; Haug, C.; Haug, J.T. The Morphological Diversity of Dragon Lacewing Larvae (Nevrorthidae, Neuroptera) Changed More over Geological Time Scales Than Anticipated. Insects 2023, 14, 749. https://doi.org/10.3390/insects14090749
Mengel L, Linhart S, Haug GT, Weiterschan T, Müller P, Hoffeins C, Hoffeins H-W, Baranov V, Haug C, Haug JT. The Morphological Diversity of Dragon Lacewing Larvae (Nevrorthidae, Neuroptera) Changed More over Geological Time Scales Than Anticipated. Insects. 2023; 14(9):749. https://doi.org/10.3390/insects14090749
Chicago/Turabian StyleMengel, Laura, Simon Linhart, Gideon T. Haug, Thomas Weiterschan, Patrick Müller, Christel Hoffeins, Hans-Werner Hoffeins, Viktor Baranov, Carolin Haug, and Joachim T. Haug. 2023. "The Morphological Diversity of Dragon Lacewing Larvae (Nevrorthidae, Neuroptera) Changed More over Geological Time Scales Than Anticipated" Insects 14, no. 9: 749. https://doi.org/10.3390/insects14090749
APA StyleMengel, L., Linhart, S., Haug, G. T., Weiterschan, T., Müller, P., Hoffeins, C., Hoffeins, H.-W., Baranov, V., Haug, C., & Haug, J. T. (2023). The Morphological Diversity of Dragon Lacewing Larvae (Nevrorthidae, Neuroptera) Changed More over Geological Time Scales Than Anticipated. Insects, 14(9), 749. https://doi.org/10.3390/insects14090749