Simple Summary
Soft fruit is an economically important global fruit crop. Drosophila suzukii is a significant pest of soft fruit, including grape bayberry. In China, Trichopria drosophilae is the dominant pupal parasitoid of D. suzukii. In order to control the population of D. suzukii more effectively and reduce the use of pesticides, researchers have investigated the potential role of T. drosophilae in the biological control of D. suzukii in China. In this paper, we investigated the functional response and control potential of T. drosophilae on D. suzukii pupae at different temperatures. Our aim was to expand the range of control applications for T. drosophilae. We investigated the functional response and interference effect of T. drosophilae and found that T. drosophilae showed good levels of parasitic ability on D. suzukii pupae in the laboratory environment. At temperatures of 23 °C to 35 °C, T. drosophilae exhibited a type II functional response on D. suzukii pupae.
Abstract
Researchers have previously investigated the role of Trichopria drosophilae as a pupal parasitoid in the biological control of Drosophila suzukii in China. Here, we investigated the ability of T. drosophilae to parasitize D. suzukii pupae at different temperatures. To do this, we evaluated the functional response of T. drosophilae to D. suzukii pupae at different temperatures and investigated the specific effects of density on parasitism. The results show that the parasitic functional response of T. drosophilae under different high-temperature stimuli is Holling type II. After processing at 29 °C, the instantaneous search rate was 1.1611; the theoretical maximum parasitic value was 20.88 at 31 °C. The parasitic efficiency decreased with increasing stimulation temperature, as the host pupa density increased from 5 to 25, and the strongest search effect occurred at 0.87 at 27 °C. The searching effect of T. drosophilae at each temperature fell gradually with an increase in prey density from 5 to 25. At 31 °C, the theoretical parasitic maximum of T. drosophilae reached a maximum of 20.88 pupae. At this temperature, when a pair of T. drosophilae was placed in a pupa density of 50, its actual total number of parasites was 18.60.
1. Introduction
The spotted wing Drosophila, Drosophila suzukii [Matsumura] (Diptera: Drosophilidae) was first discovered in 1916 in a strawberry orchard in the Yamanashi Prefectureof Japan [1]. D. suzukii was first detected in California, USA, in 2008. In Europe, D. suzukii was detected in Italy and Spain in 2009 and has spread rapidly; as of 2018, D. suzukii has spread globally to Mexico and Slovenia [2]. The host plants of D. suzukii are extensive; thus, the effect of this pest can cause damage to many economically important fruits, including Myrica rubra, Cerasus pseudocerasus, Vitis vinifera, and Rubus corchorifolius in China. Studies have shown that D. suzukii not only feeds on fruits with damaged skin or fruit that has dropped to the ground, but it also causes damage to mature or almost mature fresh fruit with soft skins. A female can directly insert her ovipositor into relatively mature or mature fresh fruit with soft skin and lay eggs directly into the pulp. The larvae subsequently feed on the pulp and then move their feeding point to the surrounding tissue, thus causing rapid decay and softening. This accelerates decay, renders the fruit inedible, and causes huge economic losses in the fruit industry [3]. According to statistics, in 2009, the damage rate of D. suzukii to cherry crops in California, USA, reached 20%, and the damage caused by D. suzukii to blueberries, raspberries, and strawberries reached 40%, 70%, and 80%, respectively [4,5]. Worryingly, the potential economic losses caused by D. suzukii continue to rise globally [6].
At present, D. suzukii is controlled mainly by chemical agents. However, the oviposition habit of D. suzukii and its resistance to chemical control measures resulting from the extensive and widespread use of chemical agents have led to a reduction in the efficacy of chemical control [7]. Based on the unique style of oviposition habit and the damaging characteristics of D. suzukii, the use of parasitic natural enemies to control D. suzukii is particularly promising [8]. In recent years, many countries have begun to strengthen their research on the biological control of D. suzukii in an attempt to make use of effective natural enemy resources and other control means to achieve sustainable development and control the occurrence of pests. Giving full play to the natural control effect of parasitic natural enemies on D. suzukii is the most economical and effective method to control D. suzukii. As one of the earliest countries in Asia to discover D. suzukii, China has identified more parasitic natural enemy insect resources because of its highly diverse natural geographic conditions. Of these resources, Trichopria drosophilae is one of the most effective parasitic wasps [9].
T. drosophilae belongs to the Hymenoptera: Diapriidae [10] and is an active parasite of D suzukii pupae. T. drosophilae has been shown to lay eggs directly into the host after anesthesia [11]. Many studies have proved that this ubiquitous parasitoid can successfully parasitize the pupae of D. suzukii in Asia, Europe, and North America [12]; furthermore, the parasitization rates of D. suzukii by T. drosophilae are high under laboratory conditions, and rearing in large numbers is feasible. In the south of Italy, Rossi Stacconi and other scholars used the local T. drosophilae population to control D. suzukii in cherry orchards; this practice significantly reduced the degree of insect damage in the orchards, which led to a 34% reduction in the damage caused by fruit insects. Collectively, these results demonstrated that the local T. drosophilae population could be used as a biological control method to control the economic losses caused by D. suzukii through field release combined with other pest control methods [13].
Studying the reproductive attributes that make parasitoids effective pest control agents, as well as the potential insect host–parasitoid interactions and behavioral ecology of parasitoids, is important for the successful application of parasitoids in biological control [14]. The reproductive success of an individual parasitoid depends on a combination of host and parasitoid interactions; in principle, it can be integrated into a host–parasitoid model to describe the temporal and spatial dynamics of host–parasitoid interactions. This type of host–parasitoid model is driven by the specific type of functional response, defined as the number of attacked hosts or prey consumed within a limited time interval in relation to host/prey density [15]. A simulation model related to host–parasitoid population dynamics is currently being developed to analyze the efficacy of T. drosophilae for the biological control of D. suzukii and to recommend further strategies for biological control.
An important component of this model is the functional response that describes the relationship between host density and the number of attacks carried out per time unit by an individual female parasitoid [16]. According to the shape of the response curve, the functional response is divided into type I (a linear increase), type II (a monotonic decelerating increase), type III (a sigmoidal increase), or type IV (dome shaped) [17]. Kacar et al. found that the type of parasitic functional response of T. drosophilae to D. suzukii and D. melanogaster pupae is a linear function [16]. Furthermore, parasitoid life history characteristics are known to change in response to temperature, and humidity can also alter these characteristics, but to a lesser extent, except under extreme conditions [18]. Similarly, the search and attack rates of parasitoids are known to be affected mainly by temperature [19]. Wu [20] treated Trichogramma nigra at 30, 33, 36, and 39 °C and four handling times of 2, 4, 6, and 8 h respectively. Analysis showed that a temperature of 30 °C increased the parasitic effect of Trichogramma pintoi. However, with an increase in temperature stimuli from 30 °C to 39 °C, the handling time was prolonged and the parasitic effect of T. pintoi was significantly inhibited; this may be related to the heat tolerance of this species. In addition, high temperature may change the functional response type of a parasitoid to the host. For example, Xie et al. [21] found that the parasitic functional response of two strains of Trichogramma dendrolimi to rice moth eggs changed from type III to type II with increasing temperature.
In the present study, we evaluated the functional response of T. drosophilae females to different densities (5, 10, 15, 20, and 25 host pupae/box) of D. suzukii after different temperature stimuli (23, 27, 29, 31, 33, and 35 °C). The purpose of this study was to clarify the type of parasitic functional response of T. drosophilae under the influence of temperature and to screen the optimal temperature and the ratio of parasitic wasps to host in the process of T. drosophilae expanding reproduction.
2. Materials and Methods
2.1. Insect Cultures
Colonies of Drosophila suzukii were derived from specimens emerging from Myrica rubra collected in Dianwei village, Kunming (102°9′ E, 24°8′ N), Yunnan Province, Southwest China, during the summer of 2016. The Trichopria drosophilae colony was derived from specimens emerging from infested fruit collected from the same location in Kunming in 2016. The stock culture of D. suzukii was held in incubators with a temperature of 25 ± 1 °C, a photoperiod of 16 h:8 h light/dark (L:D), and a relative humidity (RH) of 70 ± 5%. The colonies of D. suzukii were reared on a maize flour-based artificial diet in 50 mL plastic centrifuge tubes (height: 12 cm; diameter: 3 cm) [22]. The artificial diet contained 1000 mL of water, 15 g/L of yeast, and 21 g/L of agar, supplemented with 90 g/L of sucrose, 180 g/L of semolina flour, and 15 g/L of raisins [23]. Twice a week, adult flies were inserted into plastic tubes containing 25 mL of the diet, closed at the top by a fine mesh, and maintained for 2–3 days to encourage oviposition. T. drosophilae colonies were reared on D. suzukii. The population was maintained on a diet infested by D. suzukii larvae and pupae. Adult parasitoids were removed after two weeks just before the new generation emerged. Adults were fed a 10% honey solution in a soaked cotton ball [24].
Temperature and relative humidity inside all climate cabinets were logged at 30 min intervals by a COS-03 Temp/RH system (Shandong Renke Control Technology Co., Ltd., Jinan, China).
2.2. Functional Response at Constant Parasitoid Density and Varying Host Densities
To assess the functional response of T. drosophilae host density, we used a round transparent plastic food box (10 cm in height and diameter), created a square hole (4 cm × 4 cm) above the box, and used a 200-mesh screen as a vent. Second instar D. suzukii pupae were inserted into the boxes at densities of 5, 10, 15, 20, and 25 host pupae/box. Then, we took a pair of male and female T. drosophilae, placed them into an artificial climate room, and added 10% honey water. The temperatures were set to 23 ± 1 °C, 25 ± 1 °C, 27 ± 1 °C, 29 ± 1 °C, 31 ± 1 °C, 33 ± 1 °C, 35 ± 1 °C; the RH was 70 ± 5%; and the photoperiod (L:D) was set to 12:12. After 12 h of stimulation, we placed the T. drosophilae into plastic boxes with different densities of D. suzukii pupae at 23 °C. Twenty-four hours later, we removed the parasitoids and hosts after parasitism and raised them until the parasitoids emerged. Then, we recorded the number of eclosed pupae and dissected the hosts without parasitoids to observe whether they had been parasitized [25].
2.3. Mutual Interference Response of Parasitic Wasps
In order to understand the effect of intraspecific interference between parasitoids on the production of progeny, a certain number of host pupae were exposed to different parasitoid densities. We used a round transparent plastic food box (10 cm high and 10 cm in diameter). Then, we opened a 4 cm × 4 cm square hole above the box and created a 200-mesh screen as a ventilation hole. Two-day-old pupae of D. suzukii were then inserted at a density of 50 host pupae/box. After 24 h of eclosion, the T. drosophilae was taken and stimulated for 12 h in artificial climate boxes at different temperatures (23 ± 1 °C, 27 ± 1 °C, 29 ± 1 °C, 31 ± 1 °C, 33 ± 1 °C, 35 ± 1 °C). T. drosophilae wasps (1, 2, 3, 4, and 5 pairs) at different densities were respectively placed into the plastic boxes containing D. suzukii pupae at 23 °C. Twenty-four hours later, the parasitoids were removed and the pupae of D. suzukii were raised until parasitoids emerged [26]. The dead hosts were dissected to observe whether they had been parasitized [27]. The emergence of parasitoids and D. suzukii was observed and recorded. Five replicates were set for each density treatment. For each experiment, the RH was 70 ± 5%, and the photoperiod (L:D) was 12:12.
2.4. Data Analysis
The type of predation functional response was determined according to logistic regression analysis between prey density and captured food [28]. The shape (type) of functional response was determined by the maximum likelihood estimation of the parameters of a cubic logistic regression model between the proportion of D. suzukii pupae attacked and host pupae density. The response type was determined by the sign of the linear term coefficient of the model. A negative or positive coefficient was considered to be indicative of type II or type III responses, respectively. IBM SPSS Statistics 20.0 and Origin 2017 software were used for statistical calculation and mapping; after the normal distribution test, ANOVA and the LSD method were used for multiple comparative analysis.
The following formula was used [29]:
where N0 represents the initial number of pupae and Ne represents the number of pupae that were parasitized. P0, P1, P2, and P3 represent constant, first, second, and third power coefficients. If P1 = 0, then the number of parasitic events rose linearly with increased pupae density; this was considered a Holling I type response. If P1 < 0 and the number of parasitic events increased at a decreasing rate with increasing pupae density, then this represented a stationary state and was considered a Holling II type response. If P1 > 0, then the number of parasitic events changed in a sigmoidal way as a function of the density of pupae; this was considered to belong to a Holling III type response [30,31].
The model for the Holling Ⅱ disc equation adjusted to the parasitization data was as follows [32]:
where a represents the instantaneous attack rate, Th represents the handling time, Na represents the daily parasitic amount, N represents the prey density and T represents the test time (1 d).
The search effect (S) proposed by Holling (1959) [17] was fitted with the host density model, as follows:
a, Th, and N were the same as for the Holling Ⅱ equation.
In order to estimate the parameters of the parasitic functional response of T. drosophilae in a population, the following modification was made to the random parasitoid equation to include the potential interference among parasitic wasps [33]:
where A represents the parasitized number, P represents the parasitoid density, Q represents the quest constant, and m represents the mutual interference constant. The nonlinear least squares regression of the number of parasitized host pupae and parasitoid density was used to estimate the parameters of this functional response modified type II model.
3. Results
3.1. Functional Response at Constant Parasitoid Density and Varying Host Density after Stimulation at Different Temperatures
According to the logistic regression analysis of the proportion of Drosophila suzukii pupae parasitized in relation to the density of pupae, the estimated coefficient of the linear term was always negative. The P1 values of the control temperature and five treatments at different temperatures were all less than 0, thus indicating that the parasitic functional response type of T. drosophilae under different high-temperature stimuli was Holling type II (Table 1). Under different temperature conditions, the Pseudo-R2 of each index was very close to 1. This indicated that parasitism increased with host density to an upper asymptote representing the maximum daily per capita reproductive capacity of 1-day-old T. drosophilae females (Figure 1).

Table 1.
Verification of the parasitic model for Trichopria drosophilae at different temperatures (23–35 °C).
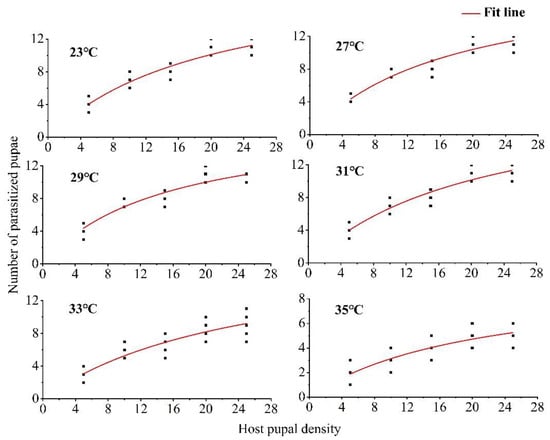
Figure 1.
Functional responses of Trichopria drosophilae to Drosophila suzukii pupae at different temperatures.
The instantaneous search rate first increased and then decreased with increasing temperature stimuli. After treatment at 29 °C, the instantaneous search rate reached a maximum of 1.1611, and then decreased with increasing temperature; the lowest instantaneous search rate of 0.4652 was recorded at 35 °C. Within the ranges of 23–29 °C and 31–35 °C, the handling time increased with increasing temperature. The theoretical maximum parasitic value was 20.88 at 31 °C and 9.59 at 35 °C. The parasitic efficiency decreased with increasing temperature, reaching a maximum of 21.73 at 27 °C and a minimum of 4.46 at 35 °C (Table 2).
Table 2.
Parasitic functional response formulae of Trichopria drosophilae to Drosophila suzukii pupa at different temperatures.
The searching effect of T. drosophilae at each temperature gradually fell with increasing prey density from 5 to 25 (Figure 2).
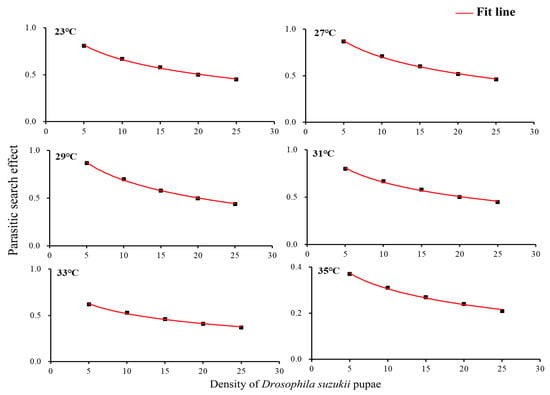
Figure 2.
Searching effect of Trichopria drosophilae for different densities of Drosophila suzukii pupae.
Furthermore, with the same density of D. suzukii pupae, the search effect of T. drosophilae increased as the temperature decreased. At a pupal density of 5, the search effect of T. drosophila stimulated at 27 °C, 29 °C, and 31 °C was significantly higher than at 33 °C and 35 °C (F5,24 = 5.11, p = 0.0001) (Figure 2).
3.2. Mutual Interference at Constant Host Density and Varying Parasitoid Densities
In addition, we investigated whether parasitic wasps exhibited different parasitism levels at the same host density, and there were significant differences between treatment groups under different stimulation temperatures (Table 3). When the density of T. drosophilae was one pair, the parasitism of D. suzukii pupae after stimulation at 35 °C was 9.0; this was significantly lower than other treatments and controls (F5,24 = 62.21, p = 0.0001). When the density of T. drosophila was two pairs, the parasitism of D. suzukii pupae when stimulated at 31 °C was 18.6; this was significantly higher than other treatment groups (F5,24 = 80.72, p = 0.0001). When the density of T. drosophila was three pairs and four pairs, the parasitism of D. suzukii pupae when stimulated at 29 °C and 31 °C was significantly higher than that of other treatment groups (three pairs: F5,24 = 39.64, p = 0.0001, four pairs: F5,24 = 7.68, p = 0.0001). When the density of T. drosophilae was five pairs, the parasitism of D. suzukii pupae when stimulated at 33 °C and 35 °C was significantly lower than that of other treatment groups (F5,24 = 8.33, p = 0.0001).
Table 3.
Parasitism rate of Drosophila suzukii pupa and self-interference response.
The average number of D. suzukii pupa parasitized (i.e., the number of parasitoid progeny produced) per single T. drosophilae female decreased as the stimulation temperature increased; this reduction in the number of pupa parasitized indicated mutual interference among parasitoids (Table 3). Within each temperature treatment group, the parasitism of one pair of parasitic wasps was significantly higher than that of the other treatments (23 °C: F4,20 = 97.05, p = 0.0001, 27 °C: F4,20 = 97.05, p = 0.0001, 29 °C: F4,20 = 91.45, p = 0.0001, 31 °C: F4,20 = 88.87, p = 0.0001, 33 °C: F4,20 = 94.12, p = 0.0001, 35 °C: F4,20 = 72.68, p = 0.0001).
The Hassell–Varley interference model was used to fit the parasitic effects of T. drosophilae under different temperature stimuli on its own density interference.
This model can reflect the self-interference of T. drosophilae during parasitism. The search coefficient Q showed a trend to first increase but then decrease with increasing temperature, reaching a maximum of 19.88 at 31 °C, and the mutual interference coefficient m reached a maximum of 0.8923 at 35 °C (Table 3). This indicated that within this temperature range, as the temperature increased, the search ability and mutual interference of T. drosophilae also increased. However, when the temperature exceeded 31 °C, the interference exceeded its increased search ability.
4. Discussion
4.1. Functional Response after Stimulation at Different Temperatures
Functional response is an important indicator for evaluating the parasitism or predation efficiency of the natural enemies of pests [34]. Since Solomon [35] first proposed the functional response method, parasitic functional response has become an important standard for measuring the effect of parasitic wasps in pest control [36]. The functional response type is a basic element that is used to understand the relationship between host density and the number of hosts parasitized [37]. Although laboratory studies on functional responses have little similarity to field contact hosts, they provide useful information relating to the potential efficacy of biological control agents. The rational use of functional response models for parasitic natural enemy insects can significantly improve the efficiency of using parasitic natural enemy insects to control pests in the field [38]. Our present results indicate that within the temperature range of the experiment, the parasitic functional response type of Trichopria drosophilae to Drosophila suzukii pupae conforms to the Holling II functional model. The parasitic effect increased with increasing host density, although the increase in parasitic effect slowed down when the host density increased to 20. This is consistent with the Holling II results of the study conducted by Fang et al. (2019) [31] on the functional response of T. drosophilae at 23 °C and also shares the same type of parasitic wasp functional response with Leptopolina japonica [39]. This may be related to the limited number of mature eggs per unit time of the parasitoid [40]. However, this is inconsistent with the results of Kacar et al., who found that the type of parasitic functional response of T. drosophilae to D. suzukii and D. melanogaster pupae is a linear function [16]. Their T. drosophilae was collected in the wild in California, USA. Our T. drosophilae collection is similar to that of Yunnan Province in China, which may be due to the inconsistency of parasitic functional response types caused by different geographical populations.
We found that the instantaneous attack rate of T. drosophilae was the highest at 29 °C and the lowest at 35 °C. Within the ranges of 23–29 °C and 31–35 °C, the handling time increased with increasing temperature. This is consistent with the pattern discovered by Li et al. [41] in their study of the effect of temperature on the functional response of Spalangia endius to the pupae of Bactrocera curubitae. These authors found that adverse environments promoted the reproductive potential of parasitic wasps to produce offspring. The reduction in handling time within the range of 29–31 °C is inconsistent with the findings of Chen et al. [42] relating to the functional response of Encarsia guadeloupae Viggiani on Aleurodicus dispersus Russell; this may be related to the physiological indicators of energy metabolism substances and physiological enzyme activity in the body at different temperatures. Several studies have evaluated the effects of temperature on the energetic efficiency of parasitic wasps: some studies found that the energetic efficiency of parasitic wasps decreased with increasing temperature [43], although a reverse relationship was reported by Vasseur and McCann [44]. At the same host density, the parasitism efficiency of T. drosophilae was the highest at 27 °C and the lowest at 35 °C; this was consistent with the findings reported by Wang et al. [45] after investigating the parasitic functional response of Parasitalis tenuis to Spodoptera frugiperda. In the present study, the parasitic efficiency of the parasitoids first increased and then decreased with increasing temperature, thus indicating that temperature was an important factor affecting the parasitic efficiency of T. drosophilae against D. suzukii pupae. When breeding T. drosophilae indoors, it is necessary to control the environmental temperature efficiently. At a temperature of 31 °C and a pupal density of 50, more pupae could be parasitized, and when releasing parasitic wasps into the field, it is necessary to choose a suitable environmental temperature and measure the density of D. suzukii in the field to maximize their parasitic potential.
4.2. The Search Effect and Interference Effect of Trichopria drosophilae
Our analysis showed that the search effect decreased as host density increased but increased with increasing temperature within the test temperature range; the highest search effect was recorded at 29 °C. Simulation results relating to the T. drosophila self-interference effect model showed that when the parasitic space and host density were fixed, the number of hosts that could be parasitized by female wasps did not increase proportionally with the increase in the parasitic wasp’s own density, thus indicating that interference effects exist among T. drosophilae individuals. Furthermore, within a certain spatial range, the interference effect increased with increasing parasitic wasp density. Therefore, in the breeding and field application of parasitic wasps, the number of parasitic wasps should be reasonably determined according to the size of the space and host density, so as to reduce the interference between parasitic wasps and reduce the cost while ensuring a high parasitic rate [46]. The density of T. drosophilae itself will also have an obvious impact on its parasitic effect. The results showed that T. drosophilae had an interference effect on D. suzukii pupae. The average parasitic effect of T. drosophilae decreased as its own density increased. The cause of intraspecific interference by T. drosophilae may be related to competition among individuals due to high density [47]. The intraspecific competition of species can be divided into two forms [48]: competition and sharing competition. When T. drosophilae parasitized D. suzukii pupae, this involved sharing competition and all parasitized wasps had equal opportunities to approach their prey. When the number of D. suzukii pupae was fixed, the competition intensity of T. drosophilae increased with increasing density; this caused an increase in intraspecific interference.
In conclusion, under laboratory conditions, in the temperature range of 23 °C to 35 °C, T. drosophilae showed a type II functional response. At 31 °C, the theoretical parasitic maximum of T. drosophilae reached a maximum, and expanding the reproductive effect is the best in this environment. When the temperature is 31 °C, the parasitoid:host pupa = 1:50, and the theoretical release effect is the best.
5. Conclusions
Under laboratory conditions, T. drosophilae showed good ability to parasitize D. suzukii pupae. In fact, in the temperature range of 23 °C to 35 °C, T. drosophilae showed a type II functional response. At 31 °C, the theoretical parasitic maximum of T. drosophilae reached a maximum, and expanding the reproductive effect is the best in this environment. When the temperature is 31 °C, the parasitoid:host pupa = 1:50, and the theoretical release effect is the best.
Author Contributions
Conceptualization, J.Z.; Methodology, J.Z.; Software, J.Z.; Validation, J.Z.; Formal analysis, Y.T.; Investigation, Y.T.; Resources, Q.C.; Data curation, Q.C.; Writing—original draft, Q.C.; Writing—review and editing, Q.C.; Visualization, G.C.; Supervision, G.C.; Project administration, X.Z.; Funding acquisition, X.Z. All authors have read and agreed to the published version of the manuscript.
Funding
The present research work was supported by the Key Agricultural Joint Project of Basic Research in Yunnan Province (202101BD070001-015), Reserve Talents of Young and Middle-aged Academic and Technological Leaders in Yunnan Province (202105AC160071), the Young Top Talents of the “High-level Talents Training Support Program in Yunnan Province” (YNWRQNBJ2020291).
Data Availability Statement
The datasets analyzed in the present study are available from the corresponding authors on reasonable request.
Acknowledgments
We thank Lingmei Li and Xiaoyan Liao (Yunnan Agricultural University) for helping us to collect the Drosophila suzukii and Trichopria drosophilae populations in the field. We thank Guizhen Qian and Yijuan Meng (Yunnan Agricultural University) for helping to feed the experimental population.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Doris, Y.; Zalom, F.G.; Hamby, K.A. Host status and fruit odor response of Drosophila suzukii (Diptera: Drosophilidae) to figs and mulberries. J. Econ. Entomol. 2013, 106, 1932–1937. [Google Scholar]
- Kruit, W.A.; Wertheim, B.; Beukeboom, L.W. Artificial selection for nonreproductive host killing in a native parasitoid on the invasive pest, Drosophila suzukii. Evol. Appl. 2021, 14, 1993–2011. [Google Scholar]
- Elsensohn, J.E.; Schal, C.; Burrack, H.J. Plasticity in oviposition site selection behavior in Drosophila suzukii (Diptera: Drosophilidae) in relation to adult density and host distribution and quality. J. Econ. Entomol. 2021, 114, 1517–1522. [Google Scholar] [CrossRef] [PubMed]
- Goodhue, R.E.; Bolda, M.; Farnsworth, D.; Williams, J.C.; Zalom, F.G. Spotted wing drosophila infestation of California strawberries and raspberries: Economic analysis of potential revenue losses and control costs. Pest Manag. Sci. 2011, 67, 1396–1402. [Google Scholar] [CrossRef] [PubMed]
- Bolda, M.P.; Goodhue, R.E.; Zalom, F.G. Spotted wing drosophila: Potential economic impact of a newly established pest. Agric. Resour. Econ. Update 2010, 13, 5–8. [Google Scholar]
- Wiman, N.G.; Walton, V.; Dalton, D.T.; Gianfranco, A.; Hannah, J.B.; Joanna, C.C.; Kent, M.D.; Alberto, G.; Betsey, M.; Samantha, T.; et al. Integrating temperature-dependent life table data into a matrix projection model for Drosophila suzukii population estimation. PLoS ONE 2014, 9, e106909. [Google Scholar] [CrossRef] [PubMed]
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.S.; Dong, C.; Kent, M.D.; Patricia, G.; Andrew, P.G.; Kim, A.H.; William, D.H.; et al. Invasion biology of spotted wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Swoboda, B.K.A.; Mcphie, D.R.; Burrack, H.J. Reproductive Status of Drosophila suzukii (Diptera: Drosophilidae) females influences attraction to fermentation-based baits and ripe fruits. J. Econ. Entomol. 2017, 110, 1648–1652. [Google Scholar] [CrossRef] [PubMed]
- Yi, C.D. Studies Basic Research on the Biology, Rapid Reproduction and Cold Storage of Trichopria drosophilae (Hymenoptera: Diapriidae), one of Parasitoids of Drosophila suzukii Matsumura. Master’s Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2019. [Google Scholar]
- Yi, C.; Cai, P.; Lin, J.; Liu, X.; Ao, G.; Zhang, Q.; Xia, H.; Yang, J.; Ji, Q. Life History and Host Preference of Trichopria drosophilae from Southern China, one of the effective pupal parasitoids on the Drosophila species. Insects 2020, 11, 103. [Google Scholar] [CrossRef]
- Stacconi, M.R.; Grassi, A.; Ioriatti, C.; Anfora, G. Augmentative releases of Trichopria drosophilae for the suppression of early season Drosophila suzukii populations. BioControl 2019, 64, 9–19. [Google Scholar] [CrossRef]
- Wang, X.G.; Nance, A.H.; Jones, J.M.L.; Kim, A.H.; Kent, M.D. Aspects of the biology and reproductive strategy of two Asian larval parasitoids evaluated for classical biological control of Drosophila suzukii. Biol. Control 2018, 121, 58–65. [Google Scholar] [CrossRef]
- Häussling, B.J.M.; Lienenlüke, J.; Stökl, J. The preference of Trichopria drosophilae for pupae of Drosophila suzukii is independent of host size. Sci. Rep. 2021, 11, 995. [Google Scholar] [CrossRef] [PubMed]
- Parra, J.R.P.; Coelho, A., Jr. Insect Rearing Techniques for Biological Control Programs, a Component of Sustainable Agriculture in Brazil. Insects 2022, 13, 105. [Google Scholar] [CrossRef] [PubMed]
- Wiedenmann, R.N. Functional response of the parasite Cotesia flavipes (Hymenoptera: Braconidae) at low densities of the host Diatraea saccharalis (Lepidoptera: Pyralidae). Environ. Entomol. 1993, 22, 849–858. [Google Scholar] [CrossRef]
- Kaçar, G.; Wang, X.G.; Biondi, A.; Kent, M.D. Linear functional response by two pupal Drosophila parasitoids foraging within single or multiple patch environments. PLoS ONE 2017, 12, e0183525. [Google Scholar] [CrossRef] [PubMed]
- Holling, C.S. Some characteristics of simple types of predation and parasitism. Can. Entomol. 1959, 91, 385–398. [Google Scholar] [CrossRef]
- Hance, T.; van Baaren, J.; Vernon, P.; Boivin, G. Impact of extreme temperatures on parasitoids in a climate change perspective. Annu. Rev. Entomol. 2007, 52, 107–126. [Google Scholar] [CrossRef]
- Romo, C.M.; Tylianakis, J.M. Elevated temperature and drought interact to reduce parasitoid effectiveness in suppressing hosts. PLoS ONE 2013, 8, e58136. [Google Scholar] [CrossRef] [PubMed]
- Wu, J. Effects of High Temperature Stimulate on the Growth, Development and Parasitic Behavior of Trichogramma chrysalis. Master’s Thesis, Shihezi University, Xinjiang, China, 2008. [Google Scholar]
- Xie, L.; Dong, H.; Qian, H.T.; Yan, J.J.; Cong, B. Parasitic functional response of Trichogramma dendrolimi parthenogenetic strain and hermaphroditic strain to rice moth eggs at different temperatures. Acta Insectae Sin. 2013, 56, 263–269. [Google Scholar]
- Wu, J.; Liao, T.l.; Sun, P.; Shi, Z.H.; Chen, J.H. Bionomics of Drosophila suzukii Matsumura,1931 (Diptera: Drosophilidae). Plant Quar. 2013, 27, 36–41. [Google Scholar]
- Amiresmaeili, N.; Jucker, C.; Savoldelli, S.; Lupi, D. Understanding Trichopria drosophilae performance in laboratory conditions. Bull. Insectology 2018, 71, 251–256. [Google Scholar]
- Wang, C.Q.; He, L.; Hu, X.; Liu, X.L.; Yang, Z.Z.; Gu, X.S. Parasitic behavior observation and artificial breeding conditions of Trichopria drosophilae. Tianjin Agric. Sci. 2022, 28, 59–64+69. [Google Scholar]
- Hu, H.; Meng, L.; Li, B.P. Functional responses of Meteorus pulchricornis (Hymenoptera: Braconidae) to Spodoptera litura larvae at different instars. Chin. J. Biol. Control 2015, 31, 176–180. [Google Scholar]
- Liu, S.; Wang, X.; Shi, Z.; Gebremeskel, F.B. The biology of Diadromus collaris (Hymenoptera: Ichneumonidae), a pupal parasitoid of Plutella xylostella (Lepidoptera: Plutellidae), and its interactions with Oomyzus sokolowskii (Hymenoptera: Eulophidae). Bull. Entomol. Res. 2001, 91, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Desneux, N.; Barta, R.J.; Hoelmer, K.A.; Hopper, K.R.; Heimpel, G.E. Multifaceted determinants of host specificity in an aphid parasitoid. Oecologia 2009, 160, 387–398. [Google Scholar] [CrossRef]
- Hoque, M.F.; Islam, M.W.; Khalequzzaman, M. Functional response of Phytoseiulus persimilis Athias-Henriot to Tetranychus urticae Koch: Effects of prey life stages and temperature. Univ. J. Zool. Rajshahi Univ. 2012, 29, 1–8. [Google Scholar] [CrossRef]
- Enkegaard, A.; Brødsgaard, H.F.; Hansen, D.L. Macrolophus caliginosus: Functional response to whiteflies and preference and switching capacity between whiteflies and spider mites. Entomol. Exp. Appl. 2001, 101, 81–88. [Google Scholar] [CrossRef]
- Arditi, R.; Tyutyunov, Y.; Morgulis, A.; Govorukhin, V. Directed movement of predators and the emergence of density-dependence in predator–prey models. Theor. Popul. Biol. 2001, 59, 207–221. [Google Scholar] [CrossRef]
- Fang, Y.; Wu, H.; Wang, J.X.; Dou, W.J.; Zhang, X.M.; Zhang, F.; Xiao, C.; Chen, G.H. Investigation on the species of parasitic natural enemies of Drosophila variegata in Yunnan Province. J. Environ. Insects 2019, 41, 592–598. [Google Scholar]
- Mondal, R.P.; Chandra, G.; Bandyopadhyay, S.; Ghosh, A. Effect of temperature and search area on the functional response of Anisops sardea (Hemiptera: Notonectidae) against Anopheles stephensi in laboratory bioassay. Acta Trop. 2017, 166, 262–267. [Google Scholar] [CrossRef]
- Jin, L.H. Functional response and control potential of Orius sauteri (Hemiptera: Anthocoridae) on tea thrips (Dendrothrips minowai Priesner). Insects 2021, 12, 1132. [Google Scholar]
- Latifian, M.; Soleymannejadian, E.; Ghazavy, M.; Mosadegh, M.S.; Rad, B. Effect of the fungus, Beauveria bassiana (Balsamo) (Asc., Hypocreales) on the functional response and host preference of the parasitoid Cephalonomia tarsalis (Ashmead) (Hym., Bethylidae) in larval population of the sawtoothed beetle Oryzaephilus surinamensis L. (Col., Silvanidae). J. Entomol. Res. 2011, 3, 251–264. [Google Scholar]
- Solomon, M.E. The natural control of animal populations. J. Anim. Ecol. 1949, 18, 1–35. [Google Scholar] [CrossRef]
- Modesto, D.P.P.; Juan, R.G.G.; Hernandez-Suarez, E.M.; Cabello, T. Effect of temperature on life history and parasitization behavior of Trichogramma achaeae Nagaraja and Nagarkatti (Hym.: Trichogrammatidae). Insects 2020, 11, 482. [Google Scholar]
- Bayoumy, M. Functional response of the aphelinid parasitoid, Aphytis diaspidis: Effect of host scale species, Diaspidiotus perniciosus and Hemiberlesia lataniae. Acta Phytopathol. Entomol. Hung. 2011, 46, 101–113. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Zhou, X.G.; Xiao, Q.; Tang, P.; Chen, X.X. The potential of Parapanteles hyposidrae and Protapanteles immunis (Hymenoptera: Braconidae) as biocontrol agents for the tea grey geometrid Ectropis grisescens (Lepidoptera). Insects 2022, 13, 937. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Zhang, F.; Yang, F.X.; Xiao, C.; Zhang, X.M.; Chen, G.H. Study on biological characteristics of Leptopolina japonica. J. Environ. Entomol. 2021, 43, 191–198. [Google Scholar]
- Mills, N.J.; Lacan, I. Ratio dependence in the functional response of insect parasitoids: Evidence from Trichogramma minutum foraging for eggs in small host patches. Ecol. Entomol. 2004, 29, 208–216. [Google Scholar] [CrossRef]
- Li, S.R.; Wu, W.J. Effect of performance on functional response of the parasitoid Spalangiaendius (Hymenoptera: Pharmacolidae) on the pubaeo of the mellonfly, Bactroceraa curbitae (Diptera: Tephritidae). J. Biosaf. 2017, 26, 289–292. [Google Scholar]
- Chen, J.Y.; Chen, T.Y.; Fu, Y.G.; Zhang, F.P.; Han, D.Y.; Niu, L.M. Functional response of Encarsia guadeloupae Viggiani to Aleurodicus dispersus Russell. Chin. J. Biol. Control 2013, 29, 175–180. [Google Scholar]
- Skovgrd, H.; Nachman, G. Temperature-Dependent Functional Response of Spalangia cameroni (Hymenoptera: Pteromalidae), a Parasitoid of Stomoxys calcitrans (Diptera: Muscidae). Environ. Entomol. 2015, 44, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Vasseur, D.A.; Mccann, K.S. A mechanistic approach for modeling temperature-dependent consumer-resource dynamics. Am. Nat. 2005, 166, 184–198. [Google Scholar] [CrossRef]
- Wang, J.Y.; Wan, N.F.; Fan, N.N.; Jiang, J.X. Functional response of Microplitis pallidipes parasitizing Spodoptera frugiperda. Chin. J. Appl. Entomol. 2020, 57, 1319–1325. [Google Scholar]
- Shi, M.; Tang, P.; Wang, Z.Z.; Huang, J.H.; Chen, X.X. Parasitic wasps in China and their application in biological control of pests. Chin. J. Appl. Entomol. 2020, 57, 491–548. [Google Scholar]
- Grether, G.F.; Losin, N.; Anderson, C.N.; Okamoto, K. The role of interspecific interference competition in character displacement and the evolution of competitor recognition. Biol. Rev. 2009, 84, 617–635. [Google Scholar] [CrossRef] [PubMed]
- Van Veen, F.J.F.; van Holland, P.D.; Godfray, H.C.J. Stable coexistence in insect communities due to density-and trait-mediated indirect effects. Ecology 2005, 86, 3182–3189. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).