

Functional Characterization of Pheromone Receptors in the Beet Webworm, Loxostege sticticalis (Lepidoptera: Pyralidae)
 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. RNA Extraction and cDNA Synthesis
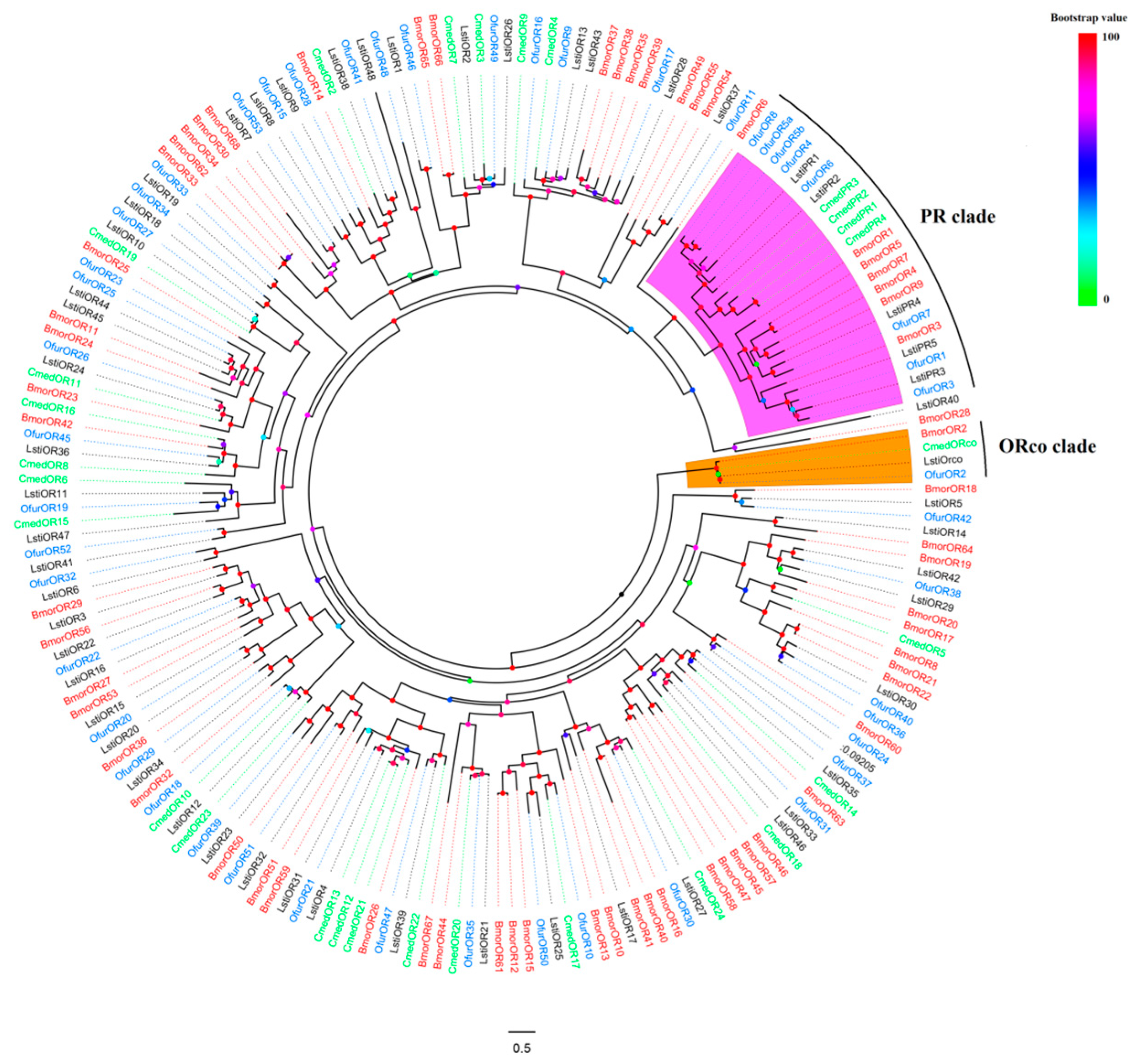
2.3. Phylogenetic Analysis
2.4. Cloning Full-Length Sequence of LstiPRs
2.5. Pheromone Components
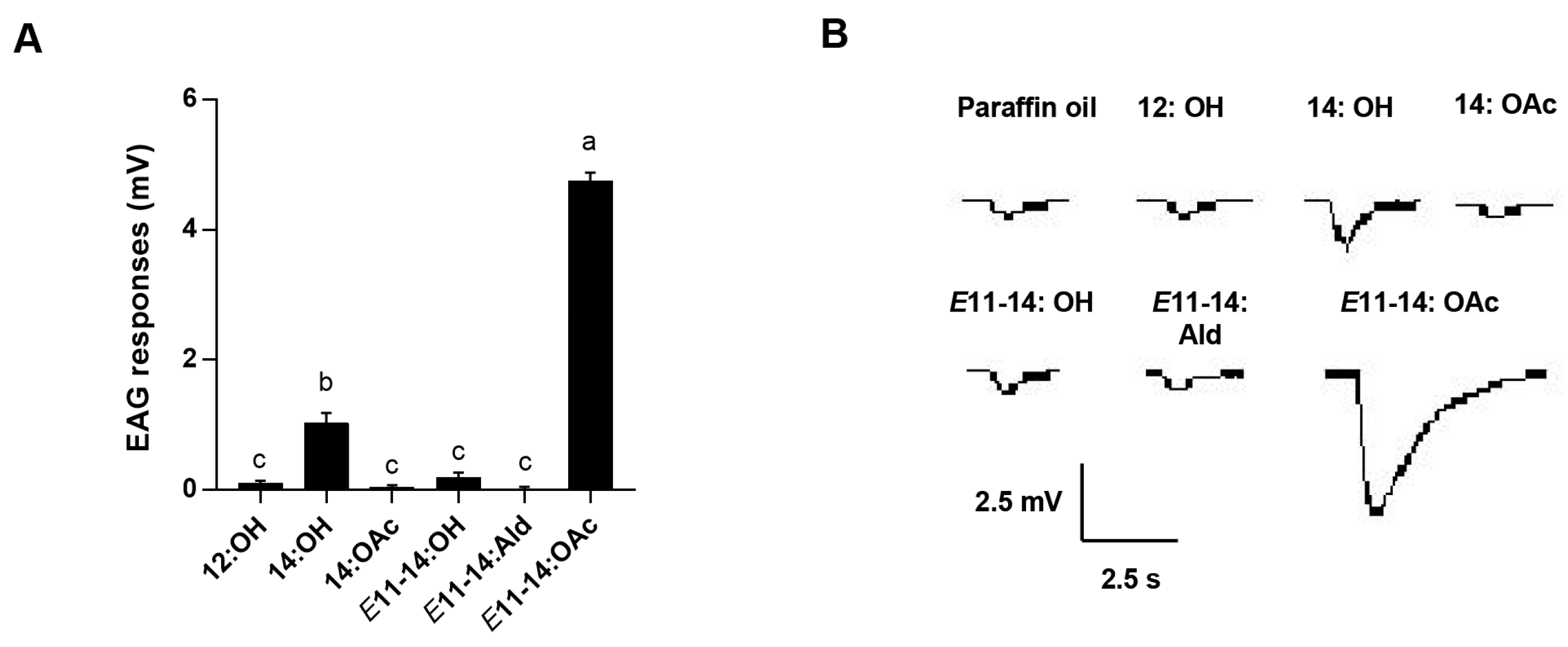
2.6. Electroantennography Responses
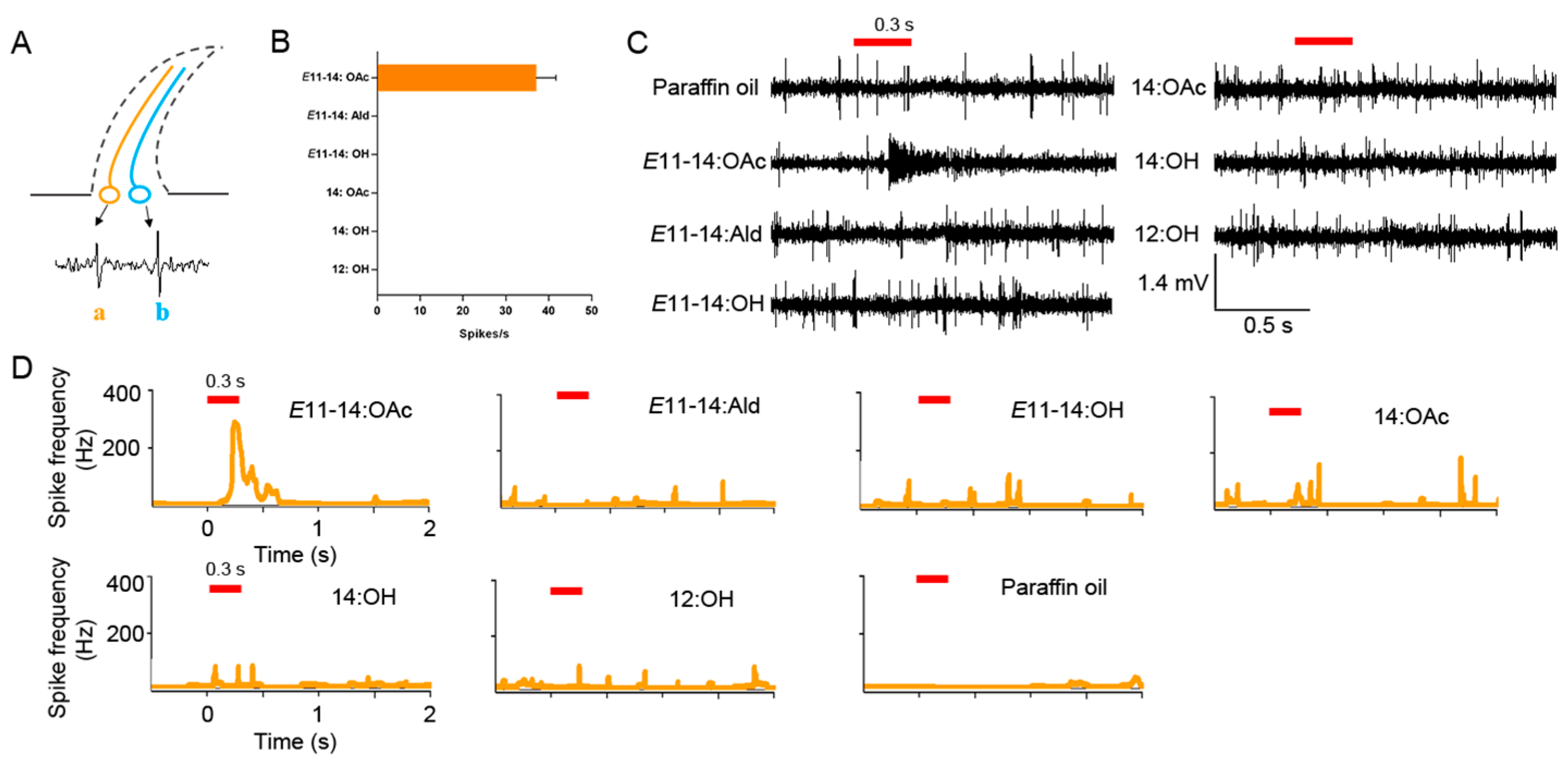
2.7. Single Sensillum Recordings
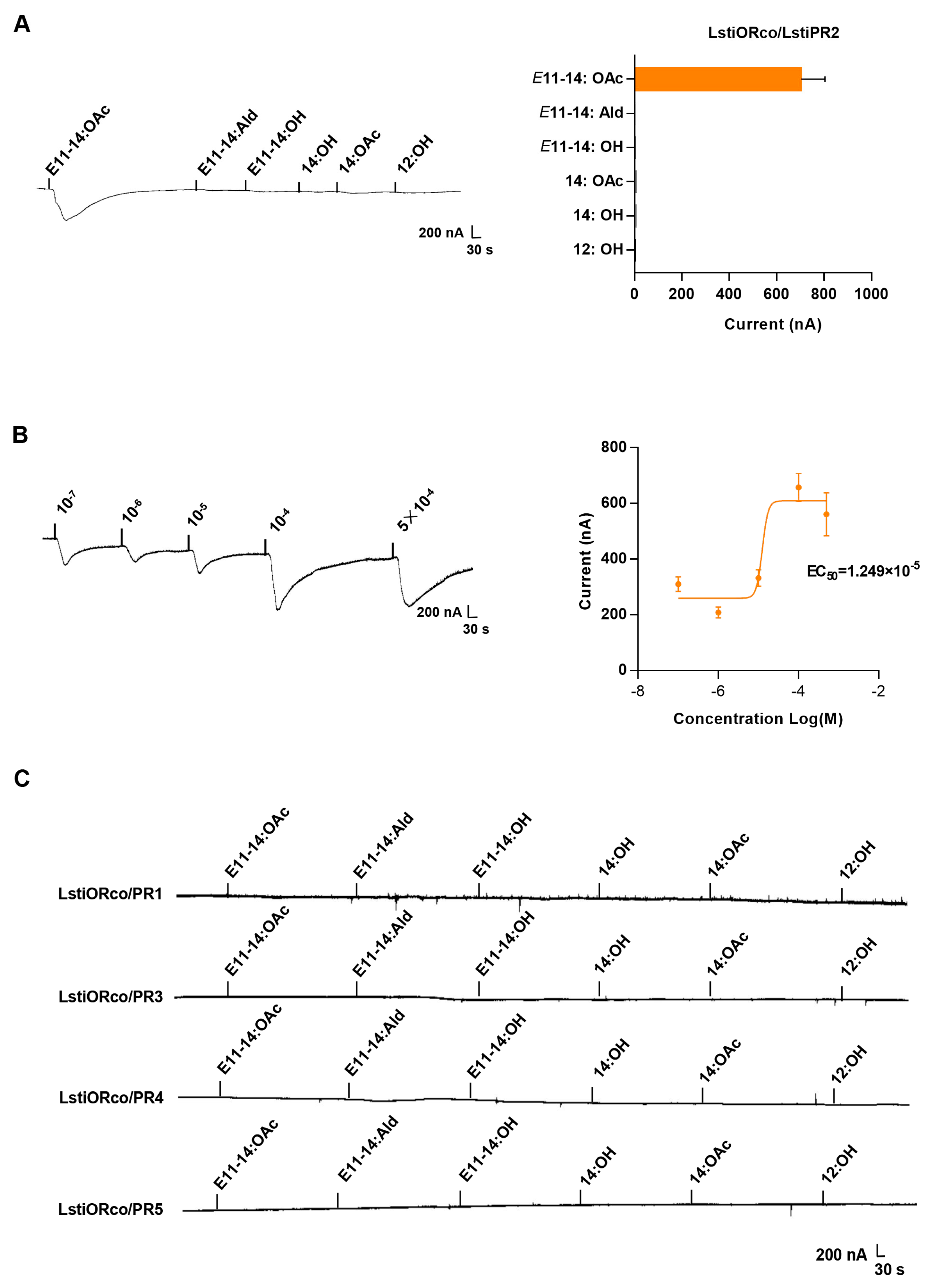
2.8. Xenopus Oocyte System and Two-Voltage Clamp Recordings
2.9. Data Analysis
3. Results
3.1. Gene Cloning and Phylogenetic Analysis
3.2. Electroantennogram Assays of Pheromone Compounds in L. sticticalis Males
3.3. The Responses of the Sensilla Trichodea ORNs of L. sticticalis Males to Pheromones
3.4. Functional Reconstitution of LstiORco and LstiPRx in Heterologous Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Auer, T.O.; Khallaf, M.A.; Silbering, A.F.; Zappia, G.; Ellis, K.; Álvarez-Ocaña, R.; Arguello, J.R.; Hansson, B.S.; Jefferis, G.S.X.E.; Caron, S.J.C.; et al. Olfactory receptor and circuit evolution promote host specialization. Nature 2020, 579, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Crespo, J.G. A review of chemosensation and related behavior in aquatic insects. J. Insect Sci. 2011, 11, 62. [Google Scholar] [CrossRef]
- Walgenbach, J.F.; Schoof, S.C.; Bosch, D.; Escudero-Colomar, L.-A.; Lingren, B.; Krawczyk, G. Comparison of sex pheromone and kairomone-enhanced pheromone lures for monitoring oriental fruit moth (Lepidoptera: Tortricidae) in mating disruption and non-disruption tree fruit orchards. Environ. Entomol. 2021, 50, 1063–1074. [Google Scholar] [CrossRef]
- Zanolli, P.; Annoscia, D.; Zanni, V.; Nazzi, F.; Pavan, F. Behavioural evidence and chemical identification of a female sex pheromone in Anagrus atomus (Hymenoptera: Mymaridae). J. Chem. Ecol. 2021, 47, 534–543. [Google Scholar] [CrossRef] [PubMed]
- Koutroumpa, F.A.; Jacquin-Joly, E. Sex in the night: Fatty acid-derived sex pheromones and corresponding membrane pheromone receptors in insects. Biochimie 2014, 107, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Butenandt, V.A.; Beckmann, R.; Stamm, D.; Hecker, E. Über den sexual-lockstoff des seidenspinners Bombyx mori. reindarstellung und konstitution. Z. Naturforschg 1959, 14, 283–284. [Google Scholar]
- Fleischer, J.; Rausch, A.; Dietze, K.; Erler, S.; Cassau, S.; Krieger, J. A small number of male-biased candidate pheromone receptors are expressed in large subsets of the olfactory sensory neurons in the antennae of drones from the European honey bee Apis mellifera. Insect Sci. 2022, 29, 749–766. [Google Scholar] [CrossRef]
- Grant, G.G.; Katovich, S.A.; Hall, D.J.; Lombardo, D.A.; Lessor, K.N.S.A. Sex pheromone identification and trapping of Dioryctria resinosella (Lepidoptera, Pyralidae). Environ. Entomol. 1993, 22, 154–161. [Google Scholar] [CrossRef]
- Lee, S.C.; Lee, J.W.; Lee, D.H.; Huh, M.J.; Nam, I.; Park, J.H.; Jung, M.; Park, I.K. Identification of sex pheromone components of Korean Dioryctria abietella (Lepidoptera: Pyralidae) population and synergism of pheromone and pine cone volatile blends. J. Econ. Entomol. 2022, 115, 178–186. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, S.; Guo, M.B.; Hou, X.Q.; Yang, B.; Wang, G.R. Optimization of a pheromone lure by analyzing the peripheral coding of sex pheromones of Spodoptera frugiperda in China. Pest Manag. Sci. 2022, 78, 2995–3004. [Google Scholar] [CrossRef]
- Yuvaraj, J.K.; Jordan, M.D.; Zhang, D.-D.; Andersson, M.N.; Löfstedt, C.; Newcomb, R.D.; Corcoran, J.A. Sex pheromone receptors of the light brown apple moth, Epiphyas postvittana, support a second major pheromone receptor clade within the Lepidoptera. Insect Biochem. Mol. Biol. 2022, 141, 103708. [Google Scholar] [CrossRef]
- Zhang, J.; Yan, S.; Liu, Y.; Jacquin-Joly, E.; Dong, S.; Wang, G. Identification and functional characterization of sex pheromone receptors in the common cutworm (Spodoptera litura). Chem. Senses 2015, 40, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Bawin, T.; Collard, F.; De Backer, L.; Yarou, B.B.; Compère, P.; Francis, F.; Verheggen, F.J. Structure and distribution of the sensilla on the antennae of Tuta absoluta (Lepidoptera: Gelechiidae). Micron 2017, 96, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Zacharuk, R.Y.; Shields, V.D. Sensilla of immature insects. Annu. Rev. Enotomol. 1991, 36, 331–354. [Google Scholar] [CrossRef]
- da Silva, K.B.; da Silva, C.B.; Júnior, K.A.L.R.; de Freitas, J.M.D.; de Freitas, J.D.; Chia, G.S.; Tinôco, R.S.; da Costa, J.G.; Goulart, H.F.; Santana, A.E.G. Morphology and distribution of antennal sensilla of Automeris liberia (Lepidoptera: Saturniidae). Micron 2019, 123, 102682. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.A.; Ignell, R.; Carlson, J.R. Chemosensory coding by neurons in the coeloconic sensilla of the Drosophila antenna. J. Neurosci. 2005, 25, 8359–8367. [Google Scholar] [CrossRef]
- Cloonan, K.; Rizzato, A.R.; Ferguson, L.; Hillier, N.K. Detection of heliothine sex pheromone components in the Australian budworm moth, Helicoverpa punctigera: Electrophysiology, neuroanatomy, and behavior. J. Comp. Physiol. A 2020, 206, 939–950. [Google Scholar] [CrossRef]
- Kohl, J.; Huoviala, P.; Jefferis, G.S. Pheromone processing in Drosophila. Curr. Opin. Neurobiol. 2015, 34, 149–157. [Google Scholar] [CrossRef]
- Liu, H.; Liu, L.; Cheng, P.; Huang, X.; Gong, M. An odorant receptor from Anopheles sinensis in China is sensitive to oviposition attractants. Malar. J. 2018, 17, 348. [Google Scholar] [CrossRef]
- Riesgo-Escovar, J.R.; Piekos, W.B.; Carlson, J.R. The Drosophila antenna: Ultrastructural and physiological studies in wild-type and lozenge mutants. J. Comp. Physiol. A 1997, 180, 151–160. [Google Scholar] [CrossRef]
- Wang, G.; Vásquez, G.M.; Schal, C.; Zwiebel, L.J.; Gould, F. Functional characterization of pheromone receptors in the tobacco budworm Heliothis virescens. Insect Mol. Biol. 2011, 20, 125–133. [Google Scholar] [CrossRef]
- Schneider, D. Insect antennae. Annu. Rev. Entomol. 1964, 9, 103–122. [Google Scholar] [CrossRef]
- Terrado, M.; Pinnelli, G.R.; Sanes, J.; Plettner, E. Binding interactions, structure-activity relationships and blend effects in pheromone and host olfactory detection of herbivorous Lepidoptera. In Olfactory Concepts of Insect Control-Alternative to Insecticides, 2nd ed.; Picimbon, J.F., Ed.; Springer Nature Switzerland AG: Cham, Switzerland, 2019; Volume 2, pp. 265–310. [Google Scholar]
- Leal, W.S. Odorant reception in insects: Roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef]
- Larsson, M.C.; Domingos, A.I.; Jones, W.D.; Chiappe, M.E.; Amrein, H.; Vosshall, L.B. Or83b encodes a broadly expressed odorant receptor essential for Drosophila olfaction. Neuron 2004, 43, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Krieger, J.; Grosse-Wilde, E.; Gohl, T.; Dewer, Y.M.; Raming, K.; Breer, H. Genes encoding candidate pheromone receptors in a moth (Heliothis virescens). Proc. Natl. Acad. Sci. USA 2004, 101, 11845–11850. [Google Scholar] [CrossRef]
- Nakagawa, T.; Sakurai, T.; Nishioka, T.; Touhara, K. Insect sex-pheromone signals mediated by specific combinations of olfactory receptors. Science 2005, 307, 1638–1642. [Google Scholar] [CrossRef]
- Liu, X.L.; Sun, S.J.; Khuhro, S.A.; Elzaki, M.E.A.; Yan, Q.; Dong, S.L. Functional characterization of pheromone receptors in the moth Athetis dissimilis (Lepidoptera: Noctuidae). Pestic. Biochem. Physiol. 2019, 158, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.N.; Du, L.X.; Xu, J.W.; Wang, B.; Zhang, X.Q.; Yan, Q.; Wang, G. Functional characterization of four sex pheromone receptors in the newly discovered maize pest Athetis lepigone. J. Insect Physiol. 2018, 113, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Cao, S.; Zhang, Z.; Kong, X.; Liu, F.; Wang, G.; Zhang, S. Evolution of sex pheromone receptors in Dendrolimus punctatus Walker (lepidoptera: Lasiocampidae) is divergent from other moth species. Insect Biochem. Mol. Biol. 2020, 122, 103375. [Google Scholar] [CrossRef]
- Fleischer, J.; Pregitzer, P.; Breer, H.; Krieger, J. Access to the odor world: Olfactory receptors and their role for signal transduction in insects. Cell Mol. Life Sci. 2018, 75, 485–508. [Google Scholar]
- Fleischer, J.; Krieger, J. Insect pheromone receptors-key elements in sensing intraspecific chemical signals. Front. Cell. Neurosci. 2018, 12, 425. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.; Lv, M.; Mao, N.; Wang, C.; Cheng, Y.; Zhang, L.; Jiang, X.; Luo, L. Molecular characterization of a lysozyme gene and its altered expression profile in crowded beet webworm (Loxostege sticticalis). PLoS ONE 2016, 11, e0161384. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.S.; Li, K.B.; Zhang, S.; Cao, Y.Z.; Yin, J. Identification of candidate chemosensory genes by transcriptome analysis in Loxostege sticticalis Linnaeus. PLoS ONE 2017, 12, e0174036. [Google Scholar] [CrossRef] [PubMed]
- Wen, M.; Li, E.; Chen, Q.; Kang, H.; Zhang, S.; Li, K.; Wang, Y.; Jiao, Y.; Ren, B. A herbivore-induced plant volatile of the host plant acts as a collective foraging signal to the larvae of the meadow moth, Loxostege sticticalis (Lepidoptera: Pyralidae). J. Insect. Physiol. 2019, 118, 103941. [Google Scholar] [CrossRef] [PubMed]
- Rizvi, S.A.H.; George, J.; Reddy, G.V.P.; Zeng, X.; Guerrero, A. Latest developments in insect sex pheromone research and its application in agricultural pest management. Insects 2021, 12, 484. [Google Scholar] [CrossRef]
- Sakurai, T.; Namiki, S.; Kanzaki, R. Molecular and neural mechanisms of sex pheromone reception and processing in the silkmoth Bombyx Mori. Front. Physiol. 2014, 5, 125. [Google Scholar] [CrossRef]
- Ando, T.; Inomata, S.; Yamamoto, M. Lepidopteran sex pheromones. Top Curr. Chem. 2004, 239, 51–96. [Google Scholar]
- Liu, A.P.; Cao, Y.X.; Xu, L.B.; Gao, S.J.; Gao, X.L.; Fan, G.M. The preliminary screening of synthetic pheromone of Loxostege sticticaiis. Chin. J. Appl. Enotomol. 2011, 48, 790–795. [Google Scholar]
- Shakhmaev, R.N.; Ishbaeva, A.U.; Shayakhmetova, I.S. Stereoselective synthesis of 11(E)-tetradecen-1-yl acetate- sex pheromone of sod webworm (Loxostege sticticalis). Russ. J. Gen. Chem. 2009, 79, 1171–1174. [Google Scholar] [CrossRef]
- Croset, V.; Rytz, R.; Cummins, S.F.; Budd, A.; Brawand, D.; Kaessmann, H.; Gibson, T.J.; Benton, R. Ancient protostome origin of chemosensory ionotropic glutamate receptors and the evolution of insect taste and olfaction. PLoS Genet. 2010, 6, e1001064. [Google Scholar] [CrossRef]
- Zeng, F.-F.; Zhao, Z.-F.; Yan, M.-J.; Zhou, W.; Zhang, Z.; Zhang, A.; Lü, Z.-X.; Wang, M.-Q. Identification and comparative expression profiles of chemoreception genes revealed from major chemoreception organs of the rice leaf folder, Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). PLoS ONE 2015, 10, e0144267. [Google Scholar] [CrossRef]
- Yang, B.; Ozaki, K.; Ishikawa, Y.; Matsuo, T. Identification of candidate odorant receptors in Asian corn borer Ostrinia furnacalis. PLoS ONE 2015, 10, e0121261. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Syed, Z.; Leal, W.S. Electrophysiological measurements from a moth olfactory system. J. Vis. Exp. 2011, 49, e2489. [Google Scholar]
- Liu, W.; Jiang, X.C.; Cao, S.; Yang, B.; Wang, G.R. Functional studies of sex pheromone receptors in Asian corn borer Ostrinia furnacalis. Front. Physiol. 2018, 9, 591. [Google Scholar] [CrossRef]
- Wanner, K.W.; Nichols, A.S.; Allen, J.E.; Bunger, P.L.; Garczynski, S.F.; Linn, C.E.; Robertson, H.M.; Luetje, C.W. Sex pheromone receptor specificity in the European corn borer moth, Ostrinia nubilalis. PLoS ONE 2010, 5, e8685. [Google Scholar] [CrossRef]
- Vogel, H.; Heidel, A.J.; Heckel, D.G.; Groot, A.T. Transcriptome analysis of the sex pheromone gland of the noctuid moth Heliothis virescens. BMC Genom. 2010, 11, 29. [Google Scholar] [CrossRef]
- Renou, M.; Anton, S. Insect olfactory communication in a complex and changing world. Curr. Opin. Insect Sci. 2020, 42, 1–7. [Google Scholar] [CrossRef]
- Leal, W.S. Pheromone Reception. In The Chemistry of Pheromones and Other Semiochemicals, 2nd ed.; Schulz, S., Ed.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 341–360. [Google Scholar]
- Benton, R.; Sachse, S.; Michnick, S.W.; Vosshall, L.B. Atypical membrane topology and heteromeric function of Drosophila odorant receptors in vivo. PLoS Biol. 2006, 4, e20. [Google Scholar] [CrossRef]
- Jiang, N.J.; Mo, B.T.; Guo, H.; Yang, J.; Tang, R.; Wang, C.Z. Revisiting the sex pheromone of the fall armyworm Spodoptera frugiperda, a new invasive pest in South China. Insect Sci. 2022, 29, 865–878. [Google Scholar] [CrossRef]
- Wang, C.; Li, Y.H.; Wang, L.; Yang, B.; Wang, G.R. Development of a new sex attractant via the peripheral coding of pheromones in Mythimna loreyi. J. Agric. Food Chem. 2023, 71, 2795–2803. [Google Scholar] [CrossRef]
- Ansebo, L.; Ignell, R.; Löfqvist, J.; Hansson, B.S. Responses to sex pheromone and plant odours by olfactory receptor neurons housed in sensilla auricillica of the codling moth, Cydia pomonella (Lepidoptera: Tortricidae). J. Insect Physiol. 2005, 51, 1066–1074. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Liu, Y.; Wang, B.; Wang, G.R. A single point mutation causes one-way alteration of pheromone receptor function in two Heliothis species. IScience 2021, 24, 102981. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.T.; Liu, Y.; Yang, T.; Pelosi, P.; Dong, S.L.; Wang, G.R. Pheromone binding proteins enhance the sensitivity of olfactory receptors to sex pheromones in Chilo suppressalis. Sci. Rep. 2015, 5, 13090. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, J.M.; Gonzalez, F.; Cattaneo, A.M.; Montagné, N.; Walker, W.B.; Bengtsson, M.; Anfora, G.; Ignell, R.; Jacquin-Joly, E.; Witzgall, P. A predicted sex pheromone receptor of codling moth Cydia pomonella detects the plant volatile pear ester. Front. Ecol. Evol. 2014, 2, 33. [Google Scholar] [CrossRef]
- Reyes-Garcia, L.; Cuevas, Y.; Ballesteros, C.; Curkovic, T.; Löfstedt, C.; Bergmann, J. A 4-component sex pheromone of the Chilean fruit leaf roller Proeulia auraria (Lepidoptera: Tortricidae). Cien. Inv. Agr. 2014, 41, 9–10. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward | Reverse |
|---|---|---|
| LstiORco | atcactagtgggcccGCCACCTTCAAGATGATGACCAAAGTGAAA (ApaI) | ctagtcagtcgcggccgcTCATCTACTTCAGTTGCACCAACA (NotI) |
| LstiPR1 | atcactagtgggcccGCCACCTGGTTATTTCGGTATGAACTCTCTG (ApaI) | ctagtcagtcgcggccgcTTAATTTAAAGTGTTTGTAAGAATGCATA (NotI) |
| LstiPR2 | atcactagtgggcccGCCACCAAATCCCTCAAAATGAAGAATAAATC (ApaI) | ctagtcagtcgcggccgcCACTATTCTCCCATCGTTTGCA (NotI) |
| LstiPR3 | atcactagtgggcccGAATCTACGAAAATGTTTAAAATATGCT (ApaI) | ctagtcagtcgcggccgcGCGATGTCAATGTTCACTACTTCC (NotI) |
| LstiPR4 | atcactagtgggcccGCCACCTTTATGCATAAACTCAGAATGTTTTTAA (ApaI) | ctagtcagtcgcggccgcGTTTAATCTTTAGTTGCGAAAGTTTG (NotI) |
| LstiPR5 | atcactagtgggcccGCCACCTTCCGCGATGTAAATTACCG (ApaI) | ctagtcagtcgcggccgcAGTCTCGCTCTGAGTACCGAGAA (NotI) |
| Chemicals | CAS Number | Purity (%) |
|---|---|---|
| (E)-11-tetradecenyl acetate (E11-14:OAc) | 33189-72-9 | >90 |
| (E)-11-tetradecenal (E11-14:Ald) | 35746-21-5 | >90 |
| (E)-11-tetradecen-1-ol (E11-14:OH) | 35153-53-8 | >95 |
| 1-tetradecanol (14:OH) | 112-72-1 | >97 |
| tetradecyl acetate (14:OAc) | 638-59-5 | >97 |
| 1-dodecanol (12:OH) | 112-53-8 | >99 |
| Gene Name | Identity (%) | ||||
|---|---|---|---|---|---|
| LstiPR1 | LstiPR2 | LstiPR3 | LstiPR4 | LstiPR5 | |
| LstiPR1 | 100.00 | ||||
| LstiPR2 | 60.80 | 100.00 | |||
| LstiPR3 | 33.75 | 33.90 | 100.00 | ||
| LstiPR4 | 33.20 | 30.52 | 35.96 | 100.00 | |
| LstiPR5 | 30.01 | 29.91 | 37.10 | 53.87 | 100.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Han, H.-B.; Li, Y.-Y.; Xu, L.-B.; Hao, L.-F.; Wang, H.; Wang, W.-H.; Gao, S.-J.; Lin, K.-J. Functional Characterization of Pheromone Receptors in the Beet Webworm, Loxostege sticticalis (Lepidoptera: Pyralidae). Insects 2023, 14, 584. https://doi.org/10.3390/insects14070584
Zhang Y, Han H-B, Li Y-Y, Xu L-B, Hao L-F, Wang H, Wang W-H, Gao S-J, Lin K-J. Functional Characterization of Pheromone Receptors in the Beet Webworm, Loxostege sticticalis (Lepidoptera: Pyralidae). Insects. 2023; 14(7):584. https://doi.org/10.3390/insects14070584
Chicago/Turabian StyleZhang, Yu, Hai-Bin Han, Yan-Yan Li, Lin-Bo Xu, Li-Fen Hao, Hui Wang, Wen-He Wang, Shu-Jing Gao, and Ke-Jian Lin. 2023. "Functional Characterization of Pheromone Receptors in the Beet Webworm, Loxostege sticticalis (Lepidoptera: Pyralidae)" Insects 14, no. 7: 584. https://doi.org/10.3390/insects14070584
APA StyleZhang, Y., Han, H.-B., Li, Y.-Y., Xu, L.-B., Hao, L.-F., Wang, H., Wang, W.-H., Gao, S.-J., & Lin, K.-J. (2023). Functional Characterization of Pheromone Receptors in the Beet Webworm, Loxostege sticticalis (Lepidoptera: Pyralidae). Insects, 14(7), 584. https://doi.org/10.3390/insects14070584