Diamondback Moth Larvae Trigger Host Plant Volatiles that Lure Its Adult Females for Oviposition

 ,
, 
 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plants and Insects
2.2. Behavioral Response Treatments
2.3. Olfactory Responses to Volatiles
2.4. Headspace Collection and Volatiles Analysis
2.5. Adult Female Response to Synthetic HIPV Sources
2.6. Statistical Analysis
3. Results
3.1. Quantities of Isothiocyanate Detected from the Headspace of Multiple Damaged Barbarea vulgaris Plants
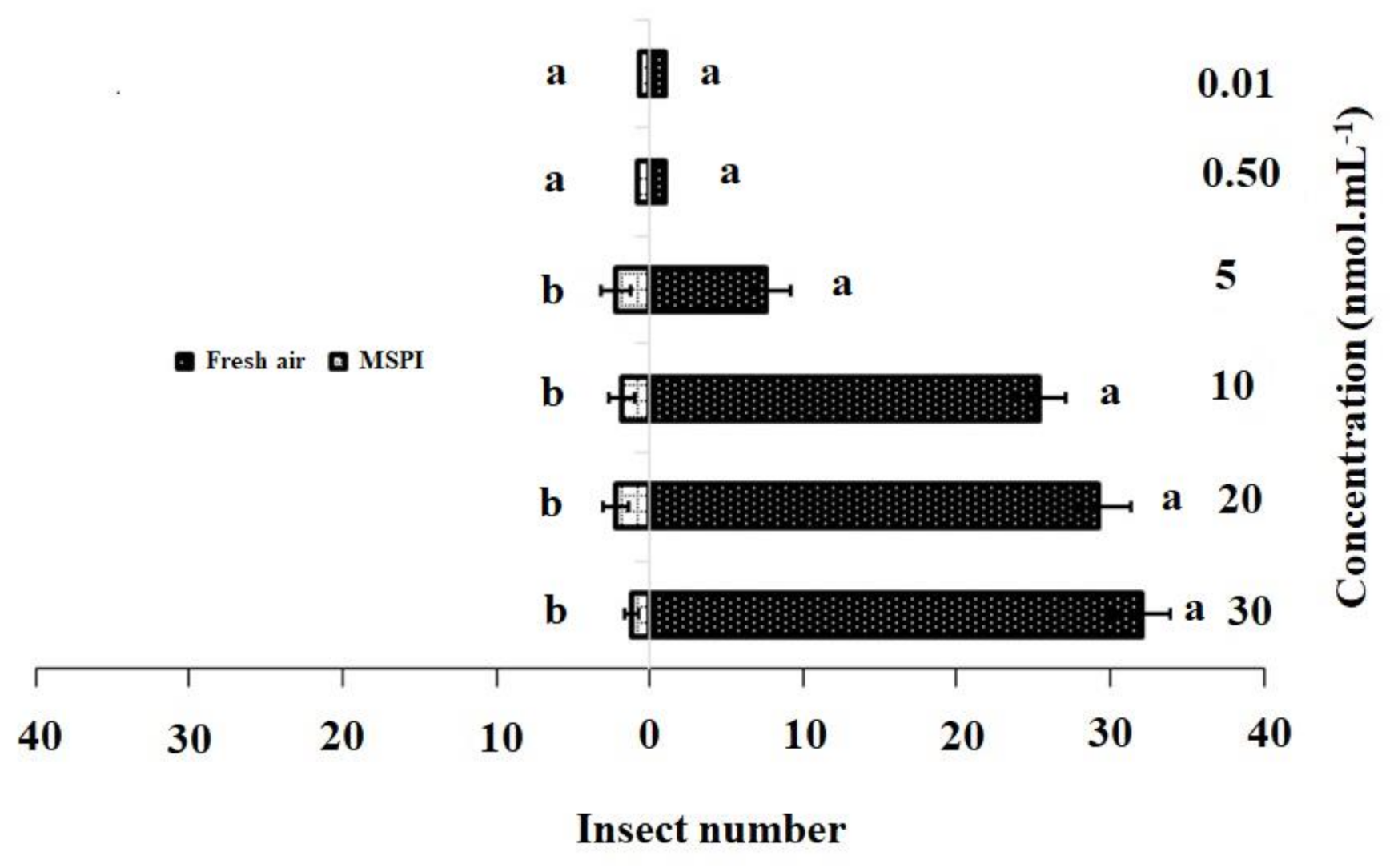
3.2. Y-Tube Olfactometer Bioassays
3.3. Adult Female Responses to Synthetic Compounds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rowen, E.; Kaplan, I. Eco-evolutionary factors drive induced plant volatiles: A meta-analysis. New Phytol. 2016, 210. [Google Scholar] [CrossRef] [PubMed]
- Dicke, M. Induced responses to herbivory by R. Karban and I.T. Baldwin. Trends Ecol. Evol. 1998, 13, 83. [Google Scholar] [CrossRef]
- Quintana-Rodriguez, E.; Morales-Vargas, A.T.; Molina-Torres, J.; Adame-Alvarez, R.M.; Acosta-Gallegos, J.A.; Heil, M. Plant volatiles cause direct, induced and associational resistance in common bean to the fungal pathogen Colletotrichum lindemuthianum. J. Ecol. 2015, 103, 250–260. [Google Scholar] [CrossRef]
- Shinoda, T.; Nagao, T.; Nakayama, M.; Serizawa, H.; Koshioka, M.; Okabe, H.; Kawai, A. Identification of a triterpenoid saponin from a crucifer, Barbarea vulgaris, as a feeding deterrent to the diamondback moth, Plutella xylostella. J. Chem. Ecol. 2002, 28, 587–599. [Google Scholar] [CrossRef]
- Shelton, A.M.; Nault, B.A. Dead-end trap cropping: A technique to improve management of the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Crop. Protion 2004, 23, 497–503. [Google Scholar] [CrossRef]
- Badenes-Perez, F.R. Simultaneous Use of Barbarea vulgaris R. Br. (Brassicaceae) as a Trap Crop for Insect Pest Management and a Salad Vegetable. Acta Hortic. 2013, 979, 737–742. [Google Scholar] [CrossRef]
- Badenes-Perez, F.R.; Reichelt, M.; Gershenzon, J.; Heckel, D.G. Using plant chemistry and insect preference to study the potential of Barbarea (Brassicaceae) as a dead-end trap crop for diamondback moth (Lepidoptera: Plutellidae). Phytochemistry 2014, 98, 137–144. [Google Scholar] [CrossRef]
- Bruin, J.; Sabelis, M.W.; Dicke, M. Do plants tap SOS signals from their infested neighbours? Trends Ecol. Evol. 1995, 10, 167–170. [Google Scholar] [CrossRef]
- Meiners, T.; Wackers, F.; Lewis, W.J. The effect of molecular structure on olfactory discrimination by the parasitoid Microplitis croceipes. Chem. Senses 2002, 27, 811–816. [Google Scholar] [CrossRef]
- Furlong, M.J.; Wright, D.J.; Dosdall, L.M. Diamondback moth ecology and management: Problems, progress, and prospects. Annu. Rev. Entomol. 2013, 58, 517–541. [Google Scholar] [CrossRef]
- Heckel, D.G.; Gahan, L.J.; Liu, Y.B.; Tabashnik, B.E. Genetic mapping of resistance to Bacillus thuringiensis toxins in diamondback moth using biphasic linkage analysis. Proc. Natl. Acad. Sci. USA 1999, 96, 8373–8377. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Huang, F.; Ghimire, M.N.; Leonard, B.R.; Siegfried, B.D.; Rangasamy, M.; Yang, Y.; Wu, Y.; Gahan, L.J.; Heckel, D.G.; et al. Efficacy of genetically modified Bt toxins against insects with different genetic mechanisms of resistance. Nat. Biotechnol. 2011, 29, 1128–1131. [Google Scholar] [CrossRef] [PubMed]
- Cusumano, A.; Weldegergis, B.T.; Colazza, S.; Dicke, M.; Fatouros, N.E. Attraction of egg-killing parasitoids toward induced plant volatiles in a multi-herbivore context. Oecologia 2015, 179, 163–174. [Google Scholar] [CrossRef]
- Spathe, A.; Reinecke, A.; Haverkamp, A.; Hansson, B.S.; Knaden, M. Host plant odors represent immiscible information entities—Blend composition and concentration matter in hawkmoths. PLoS ONE 2013, 8, e77135. [Google Scholar] [CrossRef] [PubMed]
- Allmann, S.; Spathe, A.; Bisch-Knaden, S.; Kallenbach, M.; Reinecke, A.; Sachse, S.; Baldwin, I.T.; Hansson, B.S. Feeding-induced rearrangement of green leaf volatiles reduces moth oviposition. eLife 2013, 2, e00421. [Google Scholar] [CrossRef]
- Poelman, E.H.; Zheng, S.J.; Zhang, Z.; Heemskerk, N.M.; Cortesero, A.M.; Dicke, M. Parasitoid-specific induction of plant responses to parasitized herbivores affects colonization by subsequent herbivores. Proc. Natl. Acad. Sci. USA 2011, 108, 19647–19652. [Google Scholar] [CrossRef]
- Dannon, E.A.; Tamo, M.; Van Huis, A.; Dicke, M. Effects of volatiles from Maruca vitrata larvae and caterpillar-infested flowers of their host plant Vigna unguiculata on the foraging behavior of the parasitoid Apanteles taragamae. J. Chem. Ecol. 2010, 36, 1083–1091. [Google Scholar] [CrossRef]
- Turlings, T.C.; Fritzsche, M.E. Attraction of parasitic wasps by caterpillar-damaged plants. In Proceedings of the Novartis Foundation Symposium on the Neuronal and Cognitive Effects of Oestrogens, London, UK, 7–9 September 1999; Volume 223, pp. 21–31. [Google Scholar]
- Mithen, R.; Raybould, A.; Giamoustaris, A. Divergent selection for secondary metabolites between wild populations of Brassica oleracea and its implications for plant-herbivore interactions. Heredity 1995, 75, 472. [Google Scholar] [CrossRef]
- Dimock, M.B.; Renwick, J.A.; Radke, C.D.; Sachdev-Gupta, K. Chemical constituents of an unacceptable crucifer, Erysimum cheiranthoides, deter feeding by Pieris rapae. J. Chem. Ecol. 1991, 17, 525–533. [Google Scholar] [CrossRef]
- Renwick, J.A.; Haribal, M.; Gouinguene, S.; Stadler, E. Isothiocyanates stimulating oviposition by the diamondback moth, Plutella xylostella. J. Chem. Ecol. 2006, 32, 755–766. [Google Scholar] [CrossRef]
- You, M.; Yue, Z.; He, W.; Yang, X.; Yang, G.; Xie, M.; Zhan, D.; Baxter, S.W.; Vasseur, L.; Gurr, G.M. A heterozygous moth genome provides insights into herbivory and detoxification. Nat. Genet. 2013, 45, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Lin, S.; Akutse, K.S.; Hussain, M.; Wang, L. Diaphorina citri induces huanglongbing-infected citrus plant volatiles to repel and reduce the performance of Propylaea japonica. Front. Plant Sci. 2016, 7, 1969. [Google Scholar] [CrossRef] [PubMed]
- Pineda, A.N.A.; Soler, R.; Weldegergis, B.T.; Shimwela, M.M.; Van Loon, J.J.; Dicke, M. Non-pathogenic rhizobacteria interfere with the attraction of parasitoids to aphid-induced plant volatiles via jasmonic acid signaling. Plant Cell Environ. 2013, 36, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Hountondji, F.C.; Sabelis, M.W.; Hanna, R.; Janssen, A. Herbivore-induced plant volatiles trigger sporulation in entomopathogenic fungi: The case of Neozygites tanajoae infecting the cassava green mite. J. Chem. Ecol. 2005, 31, 1003–1021. [Google Scholar] [CrossRef]
- Lucas-Barbosa, D. Integrating Studies on Plant-Pollinator and Plant-Herbivore Interactions. Trends Plant Sci. 2016, 21, 125–133. [Google Scholar] [CrossRef]
- Seidl-Adams, I.; Richter, A.; Boomer, K.B.; Yoshinaga, N.; Degenhardt, J.; Tumlinson, J.H. Emission of herbivore elicitor-induced sesquiterpenes is regulated by stomatal aperture in maize (Zea mays) seedlings. Plant Cell Environ. 2015, 38, 23–34. [Google Scholar] [CrossRef]
- Bruinsma, M.; Posthumus, M.A.; Mumm, R.; Mueller, M.J.; van Loon, J.J.; Dicke, M. Jasmonic acid-induced volatiles of Brassica oleracea attract parasitoids: Effects of time and dose, and comparison with induction by herbivores. J. Exp. Bot. 2009, 60, 2575–2587. [Google Scholar] [CrossRef]
- Coleman, R.A.; Ramchunder, S.J.; Davis, K.M.; Moody, A.J.; Foggo, A. Herbivore-induced infochemicals influence foraging behaviour in two intertidal predators. Oecologia 2007, 151, 454–463. [Google Scholar] [CrossRef]
- Arimura, G.; Kost, C.; Boland, W. Herbivore-induced, indirect plant defences. Biochim. Biophys. Acta 2005, 1734, 91–111. [Google Scholar] [CrossRef]
- Dicke, M. Specificity of herbivore-induced plant defences. In Proceedings of the Novartis Foundation Symposium on the Neuronal and Cognitive Effects of Oestrogens, London, UK, 7–9 September 1999; Volume 223, pp. 43–54. [Google Scholar]
- Sachdev-Gupta, K.; Renwick, J.A.; Radke, C.D. Isolation and identification of oviposition deterrents to cabbage butterfly, Pieris rapae, from Erysimum cheiranthoides. J. Chem. Ecol. 1991, 17, 247. [Google Scholar] [CrossRef]
- Renwick, J.A.; Radke, C.D.; Sachdev-Gupta, K. Chemical constituents of Erysimum cheiranthoides deterring oviposition by the cabbage butterfly, Pieris rapae. J. Chem. Ecol. 1989, 15, 2161–2169. [Google Scholar] [CrossRef]
- Renwick, J.A.; Radke, C.D. Chemical stimulants and deterrents regulating acceptance or rejection of crucifers by cabbage butterflies. J. Chem. Ecol. 1987, 13, 1771–1776. [Google Scholar] [CrossRef]
- Kuzina, V.; Ekstrom, C.T.; Andersen, S.B.; Nielsen, J.K.; Olsen, C.E.; Bak, S. Identification of defense compounds in Barbarea vulgaris against the herbivore Phyllotreta nemorum by an ecometabolomic approach. Plant Physiol. 2009, 151, 1977–1990. [Google Scholar] [CrossRef]
- Agerbirk, N.; Olsen, C.E.; Nielsen, J.K. Seasonal variation in leaf glucosinolates and insect resistance in two types of Barbarea vulgaris ssp. arcuata. Phytochemistry 2001, 58, 91–100. [Google Scholar] [CrossRef]
- Senatore, F.; D’Agostino, M.; Dini, I. Flavonoid glycosides of Barbarea vulgaris L. (Brassicaceae). J. Agric. Food Chem. 2000, 48, 2659–2662. [Google Scholar] [CrossRef] [PubMed]
- Strauss, S.Y.; Agrawal, A.A. The ecology and evolution of plant tolerance to herbivory. Trends Ecol. Evol. 1999, 14, 179–185. [Google Scholar] [CrossRef]
- Van Leur, H.; Vet, L.E.; van der Putten, W.H.; van Dam, N.M. Barbarea vulgaris glucosinolate phenotypes differentially affect performance and preference of two different species of lepidopteran herbivores. J. Chem. Ecol. 2008, 34, 121–131. [Google Scholar] [CrossRef]
- Van Leur, H.; Raaijmakers, C.E.; van Dam, N.M. A heritable glucosinolate polymorphism within natural populations of Barbarea vulgaris. Phytochemistry 2006, 67, 1214–1223. [Google Scholar] [CrossRef] [PubMed]
- Conti, E.; Salerno, G.; Leombruni, B.; Frati, F.; Bin, F. Short-range allelochemicals from a plant-herbivore association: A singular case of oviposition-induced synomone for an egg parasitoid. J. Exp. Biol. 2010, 213, 3911–3919. [Google Scholar] [CrossRef]
- Hossain, M.S.; Ye, W.; Hossain, M.A.; Okuma, E.; Uraji, M.; Nakamura, Y.; Mori, I.C.; Murata, Y. Glucosinolate degradation products, isothiocyanates, nitriles, and thiocyanates, induce stomatal closure accompanied by peroxidase-mediated reactive oxygen species production in Arabidopsis thaliana. Biosci. Biotechnol. Biochem. 2013, 77, 977–983. [Google Scholar] [CrossRef]
- Khokon, M.A.; Jahan, M.S.; Rahman, T.; Hossain, M.A.; Muroyama, D.; Minami, I.; Munemasa, S.; Mori, I.C.; Nakamura, Y.; Murata, Y. Allyl isothiocyanate (AITC) induces stomatal closure in Arabidopsis. Plant Cell Environ. 2011, 34, 1900–1906. [Google Scholar] [CrossRef] [PubMed]
- De Vos, M.; Kriksunov, K.L.; Jander, G. Indole-3-acetonitrile production from indole glucosinolates deters oviposition by Pieris rapae. Plant Physiol. 2008, 146, 916–926. [Google Scholar] [CrossRef] [PubMed]
- Barillari, J.; Iori, R.; Rollin, P.; Hennion, F. Glucosinolates in the subantarctic crucifer Kerguelen cabbage (Pringlea antiscorbutica). J. Nat. Prod. 2005, 68, 234–236. [Google Scholar] [CrossRef] [PubMed]
- Ahman, I. Toxicities of host secondary compounds to eggs of the Brassica specialist Dasineura brassicae. J. Chem. Ecol. 1986, 12, 1481–1488. [Google Scholar] [CrossRef]
- Tollsten, L.; Bergström, G. Headspace volatiles of whole plants and macerated plant parts of Brassica and Sinapis. Phytochemistry 1988, 27, 2073–2077. [Google Scholar] [CrossRef]
- Bukovinszky, T.; Gols, R.; Posthumus, M.A.; Vet, L.E.; Van Lenteren, J.C. Variation in plant volatiles and attraction of the parasitoid Diadegma semiclausum (Hellen). J. Chem. Ecol. 2005, 31, 461–480. [Google Scholar] [CrossRef]
- Kergunteuil, A.; Dugravot, S.; Mortreuil, A.; Le Ralec, A.; Cortesero, A.M. Selecting volatiles to protect brassicaceous crops against the cabbage root fly, Delia radicum. Entomol. Exp. Appl. 2012, 144, 69–77. [Google Scholar] [CrossRef]
- Marazzi, C.; Patrian, B.; Stadler, E. Secondary metabolites of the leaf surface affected by sulphur fertilisation and perceived by the diamondback moth. Chemoecology 2004, 14, 81–86. [Google Scholar] [CrossRef]
- Lu, J.H.; Liu, S.S.; Shelton, A.M. Laboratory evaluations of a wild crucifer Barbarea vulgaris as a management tool for the diamondback moth Plutella xylostella (Lepidoptera: Plutellidae). Bull. Entomol. Res. 2004, 94, 509–516. [Google Scholar] [CrossRef]
- Badenes-Perez, F.R.; Shelton, A.M.; Nault, B.A. Evaluating trap crops for diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). J. Econ. Entomol. 2004, 97, 1365–1372. [Google Scholar] [CrossRef]
- Muniappan, R.; Cruz, J.; Bamba, J. Trap crops for diamondback moth and other crucifer pests in Guam. In Proceedings of the Fourth International Workshop—Management of Diamondback Moth and Other Crucifer Pests, Melbourne, Australia, 26–29 November 2001; pp. 141–146. [Google Scholar]
- Badenes-Perez, F.R.; Reichelt, M.; Heckel, D.G. Can sulfur fertilisation improve the effectiveness of trap crops for diamondback moth, Plutella xylostella (L.) (Lepidoptera: Plutellidae)? Pest Manag. Sci. 2010, 66, 832–838. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.K.; Nagao, T.; Okabe, H.; Shinoda, T. Resistance in the plant, Barbarea vulgaris, and counter-adaptations in flea beetles mediated by saponins. J. Chem. Ecol. 2010, 36, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Badenes-Perez, F.R.; Shelton, A.M.; Nault, B.A. Using yellow rocket as a trap crop for diamondback moth (Lepidoptera: Plutellidae). J. Econ. Entomol. 2005, 98, 884–890. [Google Scholar] [CrossRef]
- Hussain, M.; Debnath, B.; Qasim, M.; Bamisile, B.S.; Islam, W.; Hameed, M.S.; Wang, L.; Qiu, D. Role of saponins in plant defense against specialist herbivores. Molecules 2019, 24, 2067. [Google Scholar] [CrossRef] [PubMed]
- Badenes-Perez, F.R.; Nault, B.A.; Shelton, A.M. Manipulating the attractiveness and suitability of hosts for diamondback moth (Lepidoptera: Plutellidae). J. Econ. Entomol. 2005, 98, 836–844. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment * | Major Active Compounds (ng) | |
|---|---|---|
| 3-Methylsulfinylpropyl Isothiocyanate | 4-Methylsulfinyl-3-Butenyl Isothiocyanate | |
| HB | 2.09 ± 0.16 b | 20.13 ± 2.4 a |
| FLB | 0.12 ± 0.03 c | 1.56 ± 0.04 c |
| TLB | 3.51 ± 0.28 a | 3.55 ± 0.55 c |
| MB | 2.18 ± 0.31 b | 12.01 ± 2.47 b |
| Main Compounds | Peak Area × 10−6 pA·s (μL/mL) | |||
|---|---|---|---|---|
| HB * | FLB | TLB | MB | |
| hexahydrofarnesyl acetone | 4.18 | 5.38 | 10.21 | 3.53 |
| n-heptanal | 4.19 | 4.74 | 0.17 | 3.59 |
| α-pinene | 2.34 | 5.07 | 2.92 | 3.16 |
| hexadecanoic acid | 12.20 | 9.97 | 15.62 | 11.28 |
| while phytol | 2.59 | 2.9 | 0.14 | 2.75 |
| (Z)-3-hexenyl acetate | 7.67 | 7.78 | 6.36 | 8.3 |
| sabinene | 2.8 | 2.99 | 2.52 | 3.15 |
| 3-methylsulfinylpropyl isothiocyanate | 26.64 | 1.63 | 7.36 | 24.9 |
| 4-methylsulfinyl-3-butenyl isothiocyanate | 1.68 | 0.65 | 0.45 | 1.88 |
| Treatment * | Mean Moths (%) ± SD ** | 95% Confidence Interval |
|---|---|---|
| HB | 32.00 ± 1.949 a | 26.58–37.41 |
| FLB | 14.80 ± 1.855 d | 9.65–19.95 |
| TLB | 20.40 ± 1.778 c | 15.46–25.33 |
| MB | 26.60 ± 1.931 b | 21.28–31.91 |
| Main Compounds | Correlation Coefficients between Compounds and Activity of Volatile Blends | |
|---|---|---|
| Pearson Correlation | Sig. (2-Tailed) | |
| hexahydrofarnesyl acetone | −0.455 | 0.55 |
| n-heptanal | 0.122 | 0.88 |
| α-pinene | −0.861 | 0.14 |
| hexadecanoic acid | −0.665 | 0.34 |
| while phytol | −0.181 | 0.82 |
| (Z)-3-hexenyl acetate | −0.272 | 0.73 |
| sabinene | 0.052 | 0.95 |
| 3-methylsulfinylpropyl isothiocyanate | 0.962 * | 0.04 |
| 4-methylsulfinyl-3-butenyl isothiocyanate | 0.825 * | 0.18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussain, M.; Gao, J.; Bano, S.; Wang, L.; Lin, Y.; Arthurs, S.; Qasim, M.; Mao, R. Diamondback Moth Larvae Trigger Host Plant Volatiles that Lure Its Adult Females for Oviposition. Insects 2020, 11, 725. https://doi.org/10.3390/insects11110725
Hussain M, Gao J, Bano S, Wang L, Lin Y, Arthurs S, Qasim M, Mao R. Diamondback Moth Larvae Trigger Host Plant Volatiles that Lure Its Adult Females for Oviposition. Insects. 2020; 11(11):725. https://doi.org/10.3390/insects11110725
Chicago/Turabian StyleHussain, Mubasher, Jing Gao, Summyya Bano, Liande Wang, Yongwen Lin, Steve Arthurs, Muhammad Qasim, and Runqian Mao. 2020. "Diamondback Moth Larvae Trigger Host Plant Volatiles that Lure Its Adult Females for Oviposition" Insects 11, no. 11: 725. https://doi.org/10.3390/insects11110725
APA StyleHussain, M., Gao, J., Bano, S., Wang, L., Lin, Y., Arthurs, S., Qasim, M., & Mao, R. (2020). Diamondback Moth Larvae Trigger Host Plant Volatiles that Lure Its Adult Females for Oviposition. Insects, 11(11), 725. https://doi.org/10.3390/insects11110725