
Role of Tidal Volume on Ventilator-Induced Lung Injury Under Heterogeneous Immunological Capabilities: A Mathematical Model Study

Abstract
1. Introduction
2. Methods
2.1. Model Description
2.2. Lung Immune Response System
2.3. Tidal Volume Damage System
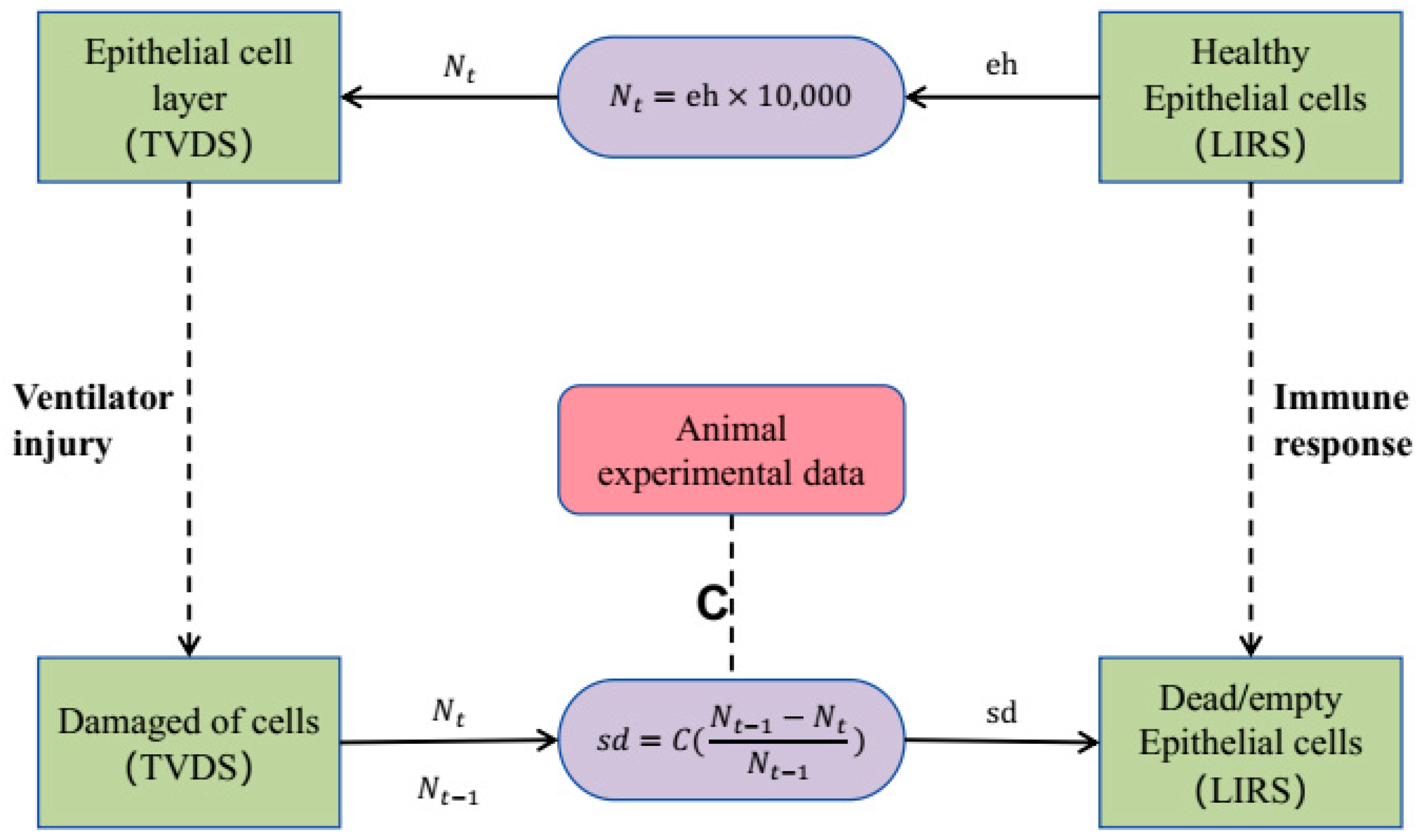
2.4. Coupling Boundary
2.5. Model Parameterization
3. Results
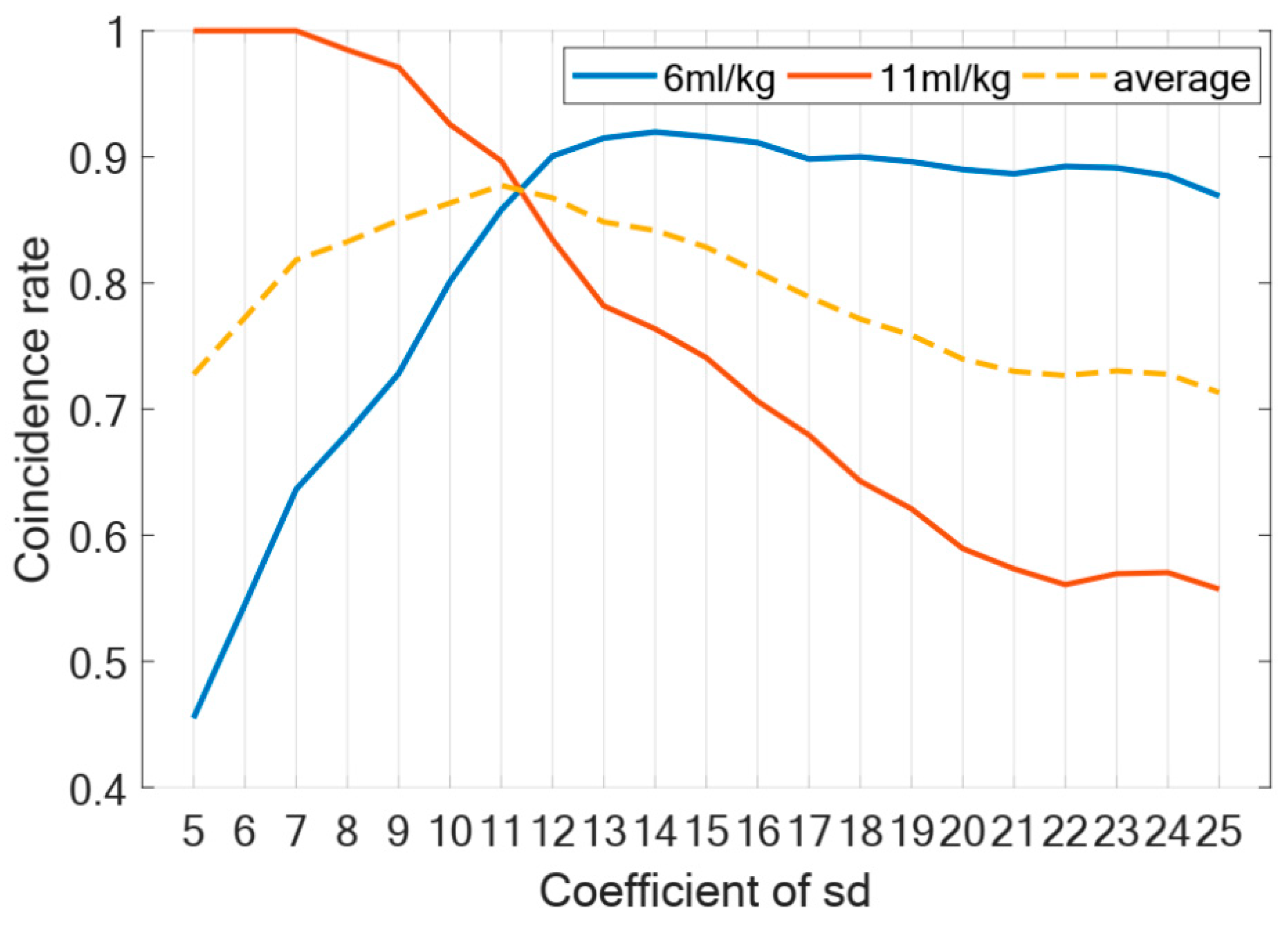
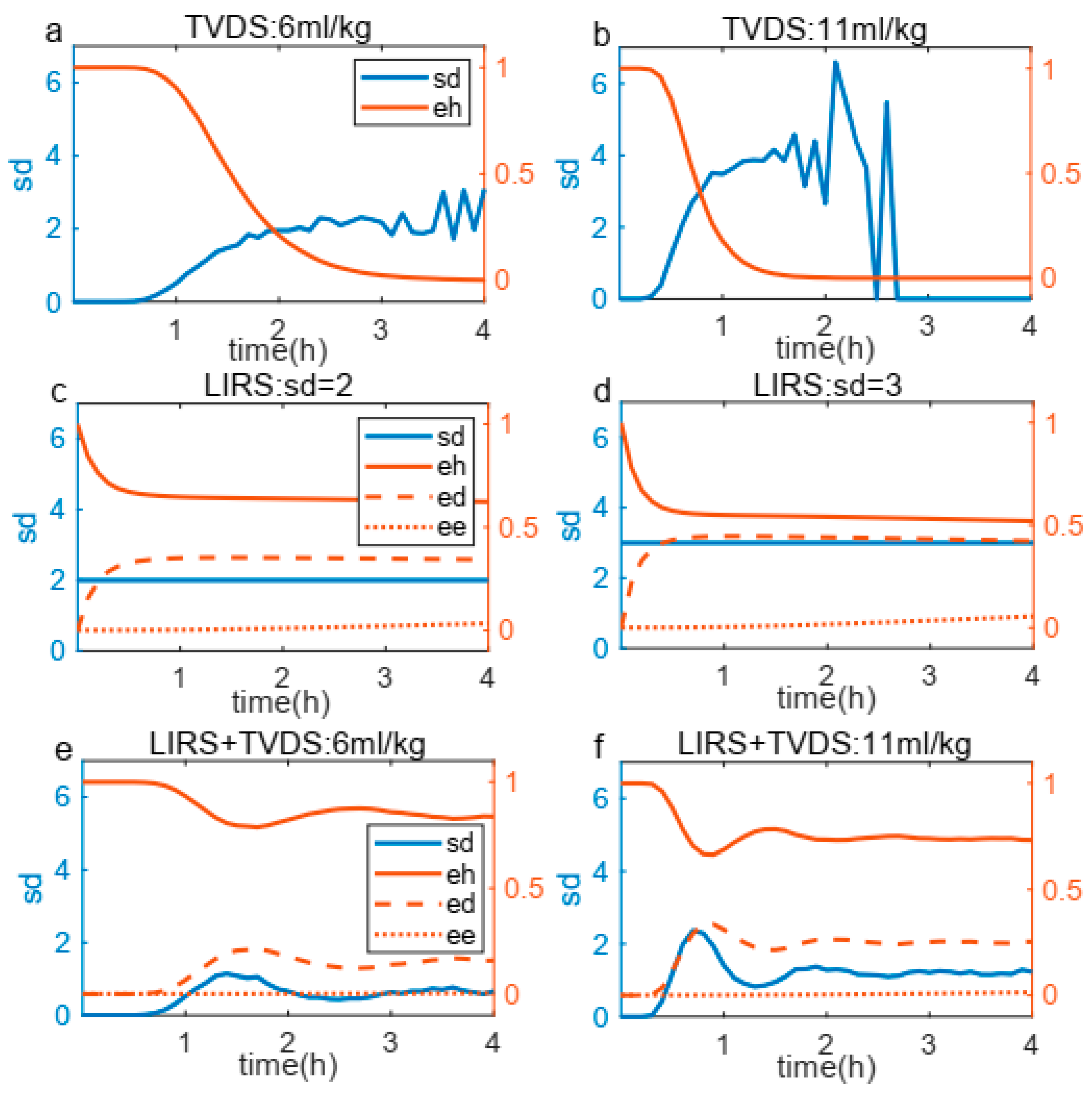
3.1. Validity of the Coupling Boundary
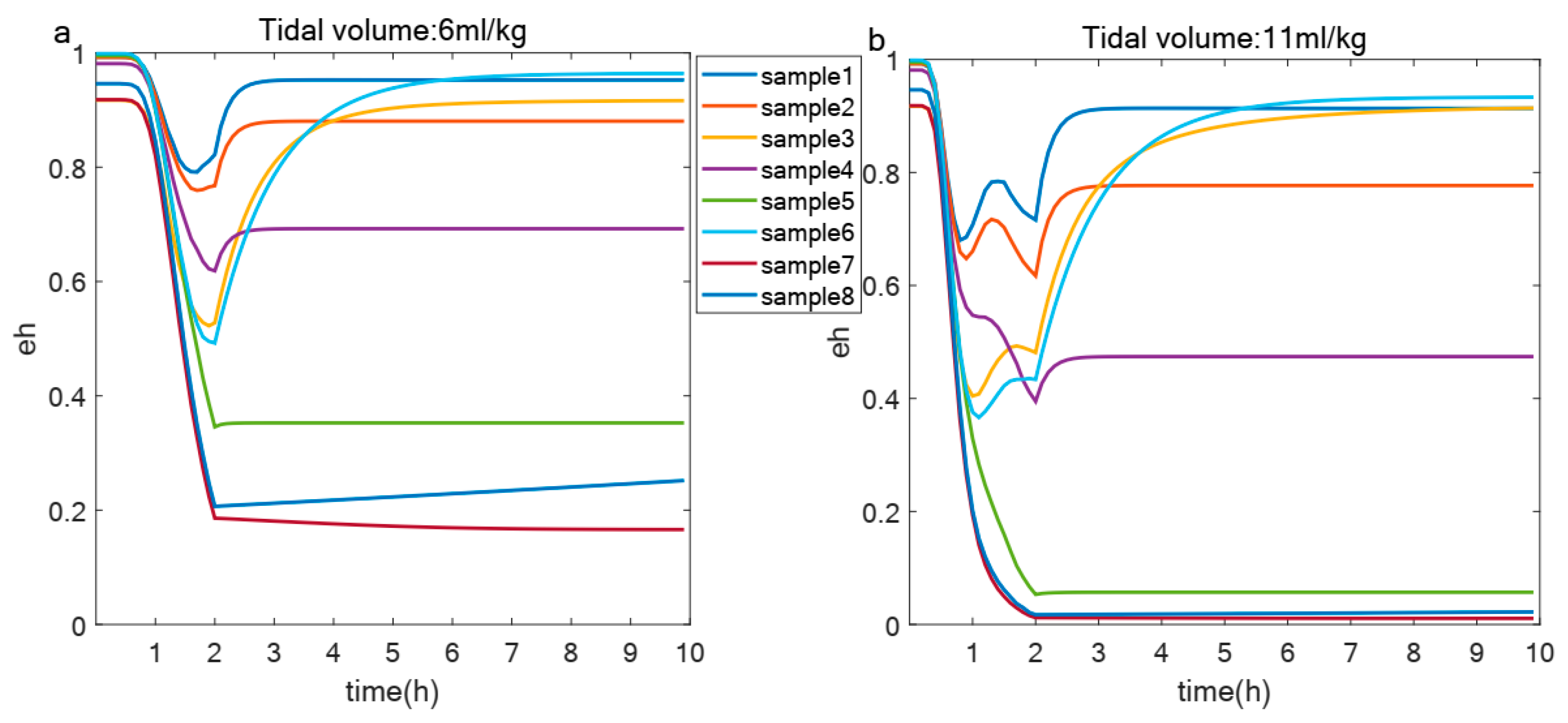
3.2. Effect of Immunological Capacity on the Degree of VILI
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Description |
|---|---|
| Relative effectiveness of a at inhibiting M0 differentiation to M1 | |
| Baseline decay of damaged cells | |
| Baseline self-resolving repair of epithelial cells | |
| Baseline repair of damaged cells | |
| Decay rate of pro-inflammatory mediators in bloodstream | |
| Decay rate of anti-inflammatory mediators in bloodstream | |
| Decay rate of naive macrophages in bloodstream | |
| Decay rate of M1 in bloodstream | |
| Decay rate of M2 in bloodstream | |
| Decay rate of pro-inflammatory mediators in lung | |
| Decay rate of anti-inflammatory mediators in lung | |
| Decay rate of naive macrophages in lung | |
| Decay rate of M1 in lung | |
| Decay rate of M2 in lung | |
| Decay rate of repair mediators in lung | |
| Decay rate of unactivated neutrophils in bloodstream | |
| Decay rate of activated neutrophils in bloodstream | |
| Source rate of naive macrophages in bloodstream | |
| Source rate of unactivated neutrophils in bloodstream | |
| Rate of self-resolving repair mediated by p | |
| Rate of repair of damaged cells by R | |
| Rate of phagocytosis of damaged cells by M1 | |
| Rate of collateral damage to epithelial cells by macrophages and neutrophils | |
| Rate of phagocytosis of damaged cells by Neutrophils in lung | |
| Source rate of background pro-inflammatory mediators in bloodstream | |
| Source rate of background anti-inflammatory mediators in bloodstream | |
| Regulates effectiveness of macrophages and neutrophils to damage epithelial cells | |
| Regulates effectiveness of M2 in bloodstream recruitment by R | |
| Regulates effectiveness of M2 in bloodstream recruitment by a | |
| Regulates effectiveness of repair of damaged cells by R |
References
- Ramcharran, H.; Bates, J.H.T.; Satalin, J.; Blair, S.; Andrews, P.L.; Gaver, D.P.; Gatto, L.A.; Wang, G.; Ghosh, A.J.; Robedee, B.; et al. Protective ventilation in a pig model of acute lung injury: Timing is as important as pressure. J. Appl. Physiol. 2022, 133, 1093–1105. [Google Scholar] [CrossRef] [PubMed]
- Minucci, S.B.; Heise, R.L.; Reynolds, A.M. Review of Mathematical Modeling of the Inflammatory Response in Lung Infections and Injuries. Front. Appl. Math. Stat. 2020, 6, 36. [Google Scholar] [CrossRef]
- Kneyber, M.C.J.; Zhang, H.; Slutsky, A.S. Ventilator-induced Lung Injury. Similarity and Differences between Children and Adults. Am. J. Respir. Crit. Care Med. 2014, 190, 258–265. [Google Scholar] [CrossRef]
- Smith, B.J.; Grant, K.A.; Bates, J.H.T. Linking the Development of Ventilator-Induced Injury to Mechanical Function in the Lung. Ann. Biomed. Eng. 2012, 41, 527–536. [Google Scholar] [CrossRef]
- Fan, E.; Del Sorbo, L.; Goligher, E.C.; Hodgson, C.L.; Munshi, L.; Walkey, A.J.; Adhikari, N.K.J.; Amato, M.B.P.; Branson, R.; Brower, R.G.; et al. An Official American Thoracic Society/European Society of Intensive Care Medicine/Society of Critical Care Medicine Clinical Practice Guideline: Mechanical Ventilation in Adult Patients with Acute Respiratory Distress Syndrome. Am. J. Respir. Crit. Care Med. 2017, 195, 1253–1263. [Google Scholar] [CrossRef]
- Marino, S.; Hogue, I.B.; Ray, C.J.; Kirschner, D.E. A methodology for performing global uncertainty and sensitivity analysis in systems biology. J. Theor. Biol. 2008, 254, 178–196. [Google Scholar] [CrossRef]
- Gary, N.; David, B.; Joydeep, S.; Brian, K.; Cordelia, Z. A two-compartment mathematical model of endotoxin-induced inflammatory and physiologic alterations in swine. Crit. Care Med. 2012, 40, 1052–1063. [Google Scholar]
- Cantone, M.; Santos, G.; Pia Wentker Lai, X.; Vera, J. Multiplicity of Mathematical Modeling Strategies to Search for Molecular and Cellular Insights into Bacteria Lung Infection. Front. Physiol. 2017, 8, 645. [Google Scholar]
- Eftimie, R.; Gillard, J.J.; Cantrell, D.A. Mathematical Models for Immunology: Current State of the Art and Future Research Directions. Bull. Math. Biol. 2016, 78, 2091–2134. [Google Scholar] [CrossRef]
- Minucci, S.; Heise, R.L.; Valentine, M.S.; Kamga Gninzeko, F.J.; Reynolds, A.M. Mathematical modeling of ventilator-induced lung inflammation. J. Theor. Biol. 2021, 526, 110738. [Google Scholar] [CrossRef]
- Bates, J.H.T.; Nieman, G.F.; Kollisch-Singule, M.; Gaver, D.P. Ventilator-Induced Lung Injury as a Dynamic Balance Between Epithelial Cell Damage and Recovery. Ann. Biomed. Eng. 2023, 51, 1052–1062. [Google Scholar] [CrossRef] [PubMed]
- Crosby, L.M.; Waters, C.M. Epithelial repair mechanisms in the lung. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2010, 298, L715–L731. [Google Scholar] [CrossRef] [PubMed]
- Gardner, A.; Borthwick, L.A.; Fisher, A.J. Lung epithelial wound healing in health and disease. Expert Rev. Respir. Med. 2014, 4, 647–660. [Google Scholar] [CrossRef]
- Beers, M.F.; Morrisey, E.E. The three R’s of lung health and disease: Repair, remodeling, and regeneration. J. Clin. Investig. 2011, 121, 2065–2073. [Google Scholar] [CrossRef]
- Herold, S.; Mayer, K.; Lohmeyer, J. Acute Lung Injury: How Macrophages Orchestrate Resolution of Inflammation and Tissue Repair. Front. Immunol. 2011, 2, 65. [Google Scholar] [CrossRef]
- Aggarwal, N.R.; King, L.S.; D’Alessio, F.R. Diverse macrophage populations mediate acute lung inflammation and resolution. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2014, 306, L709–L725. [Google Scholar] [CrossRef]
- Jacob, A.-M.; Gaver, D.P. Atelectrauma disrupts pulmonary epithelial barrier integrity and alters the distribution of tight junction proteins ZO-1 and claudin 4. J. Appl. Physiol. 2012, 113, 1377–1387. [Google Scholar] [CrossRef]
- Bates, J.H.T.; Irvin, C.G. Time dependence of recruitment and derecruitment in the lung: A theoretical model. J. Appl. Physiol. 2002, 93, 705–713. [Google Scholar] [CrossRef]
- Han, B. Ventilator-induced Lung Injury: Role of Protein-Protein Interaction in Mechanosensation. Proc. Am. Thorac. Soc. 2005, 2, 181–187. [Google Scholar] [CrossRef]
- Novak, C.; Ballinger, M.N.; Ghadiali, S. Mechanobiology of Pulmonary Diseases: A Review of Engineering Tools to Understand Lung Mechanotransduction. J. Biomech. Eng. 2021, 143, 110801. [Google Scholar] [CrossRef]
- Pelosi, P.; Rocco, P.R. Effects of mechanical ventilation on the extracellular matrix. Intensive Care Med. 2008, 34, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, E.; Yao, J.; Aymond, A.; Chrisey, D.B.; Nieman, G.F.; Bates, J.H.T.; Gaver, D.P. Electric Cell-Substrate Impedance Sensing (ECIS) as a Platform for Evaluating Barrier-Function Susceptibility and Damage from Pulmonary Atelectrauma. Biosensors 2022, 12, 390. [Google Scholar] [CrossRef]
- Rosenberger, P.; Hamlington, K.L.; Bates, J.H.T.; Roy, G.S.; Julianelle, A.J.; Charlebois, C.; Suki, B.; Smith, B.J. Alveolar leak develops by a rich-get-richer process in ventilator-induced lung injury. PLoS ONE 2018, 13, e0193934. [Google Scholar]
- Santos, R.S.; Maia, L.D.; Oliveira, M.V.; Santos, C.L.; Moraes, L.; Pinto, E.F.; Samary, C.D.; Machado, J.A.; Carvalho, A.C.; Fernandes, M.V.D.; et al. Biologic Impact of Mechanical Power at High and Low Tidal Volumes in Experimental Mild Acute Respiratory Distress Syndrome. Anesthesiology 2018, 128, 1193–1206. [Google Scholar] [CrossRef]
- Wang, T.; Chai, Z.; Wang, L.; Liu, B.; Zhao, J.; Ren, J.; Yang, B.; Wei, X.; Jiang, L.; Liu, F. IL-9 blockade attenuates inflammation in a murine model of mechanical ventilation-induced lung injury by inhibiting the NLRP3 inflammasome pathway. Inflammopharmacology 2022, 30, 1395–1406. [Google Scholar] [CrossRef]
- Rezoagli, E.; Laffey, J.G.; Bellani, G. Monitoring Lung Injury Severity and Ventilation Intensity during Mechanical Ventilation. Semin. Respir. Crit. Care Med. 2022, 43, 346–368. [Google Scholar] [CrossRef]




| TVDS | LIRS | Description |
|---|---|---|
| pb | Pro-inflammatory mediators in bloodstream | |
| ab | Anti-inflammatory mediators in bloodstream | |
| M0b | Naive macrophages in bloodstream | |
| M1b | M1 pro-inflammatory macrophages in bloodstream | |
| M2b | M2 anti-inflammatory macrophages in bloodstream | |
| N0b | Unactivated neutrophils in bloodstream | |
| Na | Activated neutrophils in bloodstream | |
| p | Pro-inflammatory mediators in lung | |
| a | Anti-inflammatory mediators in lung | |
| M0 | Naive macrophages in lung | |
| M1 | M1 pro-inflammatory macrophages in lung | |
| M2 | M2 anti-inflammatory macrophages in lung | |
| N | Neutrophils in lung | |
| AN | Apoptotic neutrophils in lung | |
| R | Repair mediators in lung | |
| eh | eh | Healthy epithelial cells |
| ed | ed | Damaged epithelial cells |
| ee | Dead epithelial cells/empty space | |
| pbreak | Probability of intercellular attachments breakage | |
| nbreak | Number of broken intercellular attachments | |
| n | Number of intercellular attachments | |
| k | Conform pbreak to a nonlinear expression constrained to lie between 0 and 1 | |
| Nt | The total number of epithelial cells at the moment t | |
| Nt−1 | The total number of epithelial cells at the moment | |
| sd | sd | Damage rate from ventilator |
| VILI Severity Score | eh Values at 2 h |
|---|---|
| Score 1 | |
| Score 2 | |
| Score 3 | |
| Score 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Y.; Liu, Y.; Lu, F.; Fang, L.; Ning, G.; Pan, Q. Role of Tidal Volume on Ventilator-Induced Lung Injury Under Heterogeneous Immunological Capabilities: A Mathematical Model Study. Life 2025, 15, 835. https://doi.org/10.3390/life15060835
Yu Y, Liu Y, Lu F, Fang L, Ning G, Pan Q. Role of Tidal Volume on Ventilator-Induced Lung Injury Under Heterogeneous Immunological Capabilities: A Mathematical Model Study. Life. 2025; 15(6):835. https://doi.org/10.3390/life15060835
Chicago/Turabian StyleYu, Yao, Yuxi Liu, Fei Lu, Luping Fang, Gangmin Ning, and Qing Pan. 2025. "Role of Tidal Volume on Ventilator-Induced Lung Injury Under Heterogeneous Immunological Capabilities: A Mathematical Model Study" Life 15, no. 6: 835. https://doi.org/10.3390/life15060835
APA StyleYu, Y., Liu, Y., Lu, F., Fang, L., Ning, G., & Pan, Q. (2025). Role of Tidal Volume on Ventilator-Induced Lung Injury Under Heterogeneous Immunological Capabilities: A Mathematical Model Study. Life, 15(6), 835. https://doi.org/10.3390/life15060835