


Integrative Application of Foliar Yeast Extract and Gibberellic Acid Improves Morpho-Physiological Responses and Nutrient Uptake of Solidago virgaurea Plant in Alkaline Soil
 , , ,
, , ,  ,
,  and
and 
Abstract
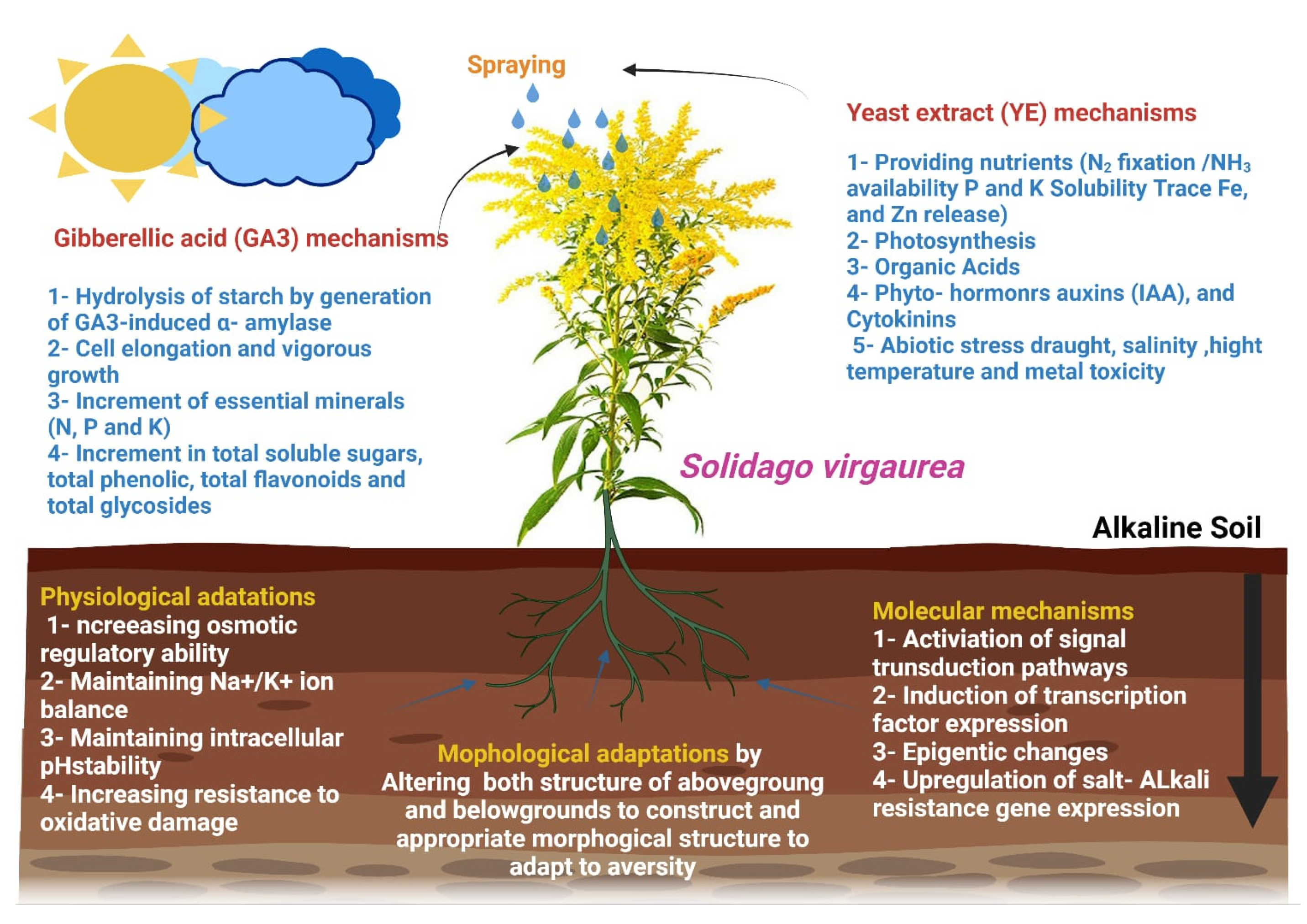
:1. Introduction
2. Materials and Methods
2.1. Experimental Layout and Soil Analysis
2.2. Plant Material
2.3. Treatments and Experimental Design
2.4. Agricultural Practices
2.5. Recording Data
2.5.1. Morphological Measurements
2.5.2. Physiological Determinations
2.5.3. Macronutrients Determinations
2.5.4. Biochemical Constituents
2.6. Statistical Analysis
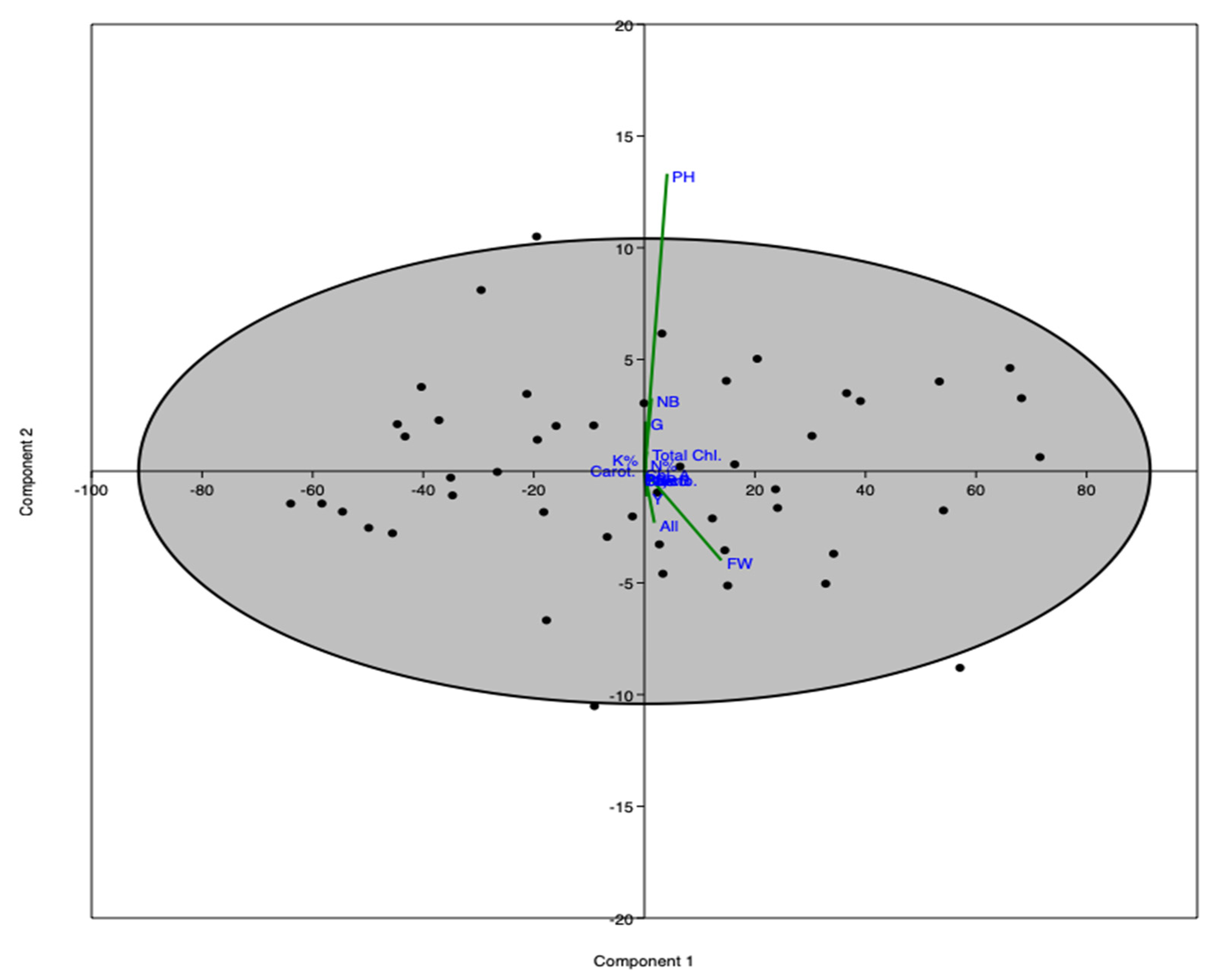
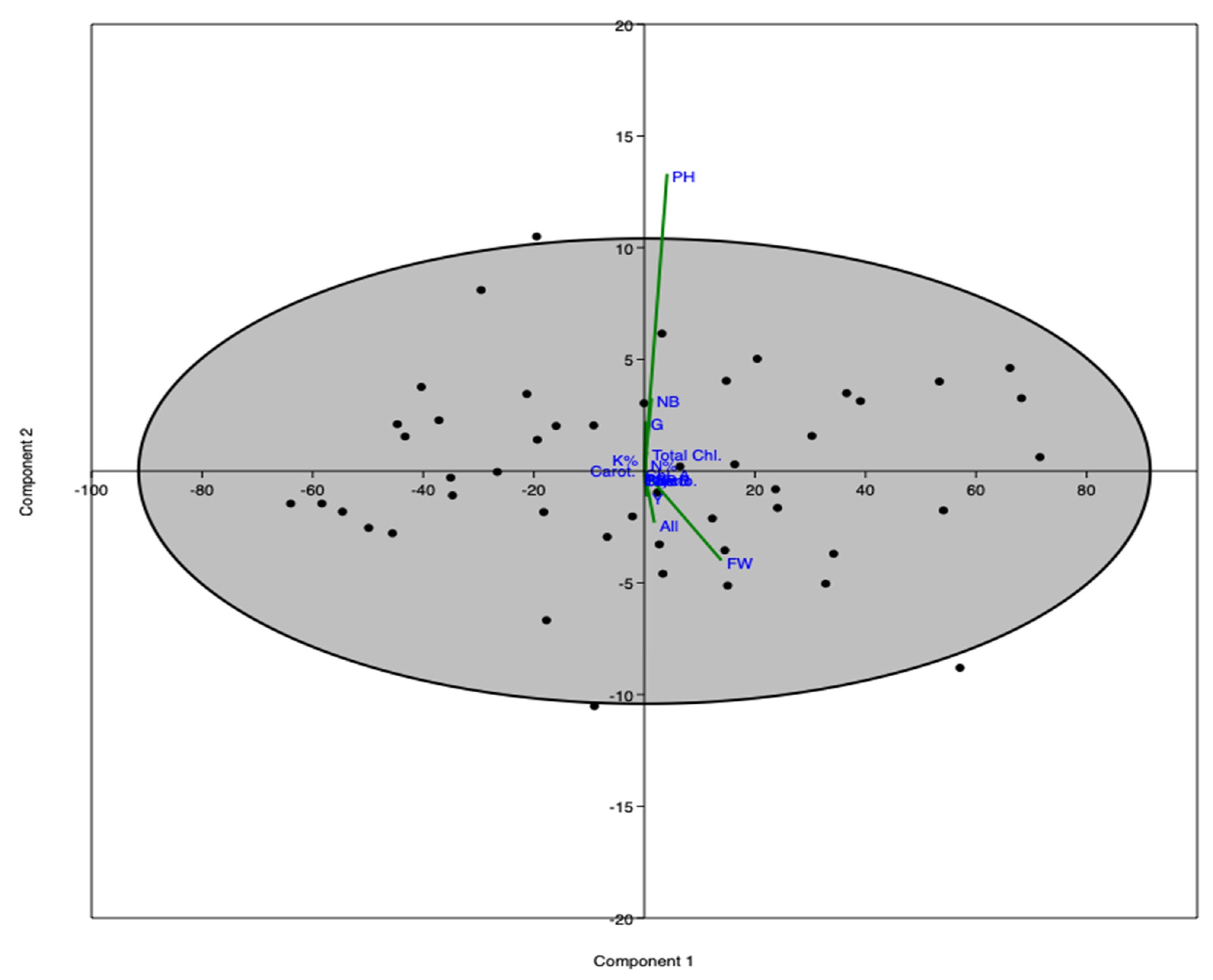
3. Results
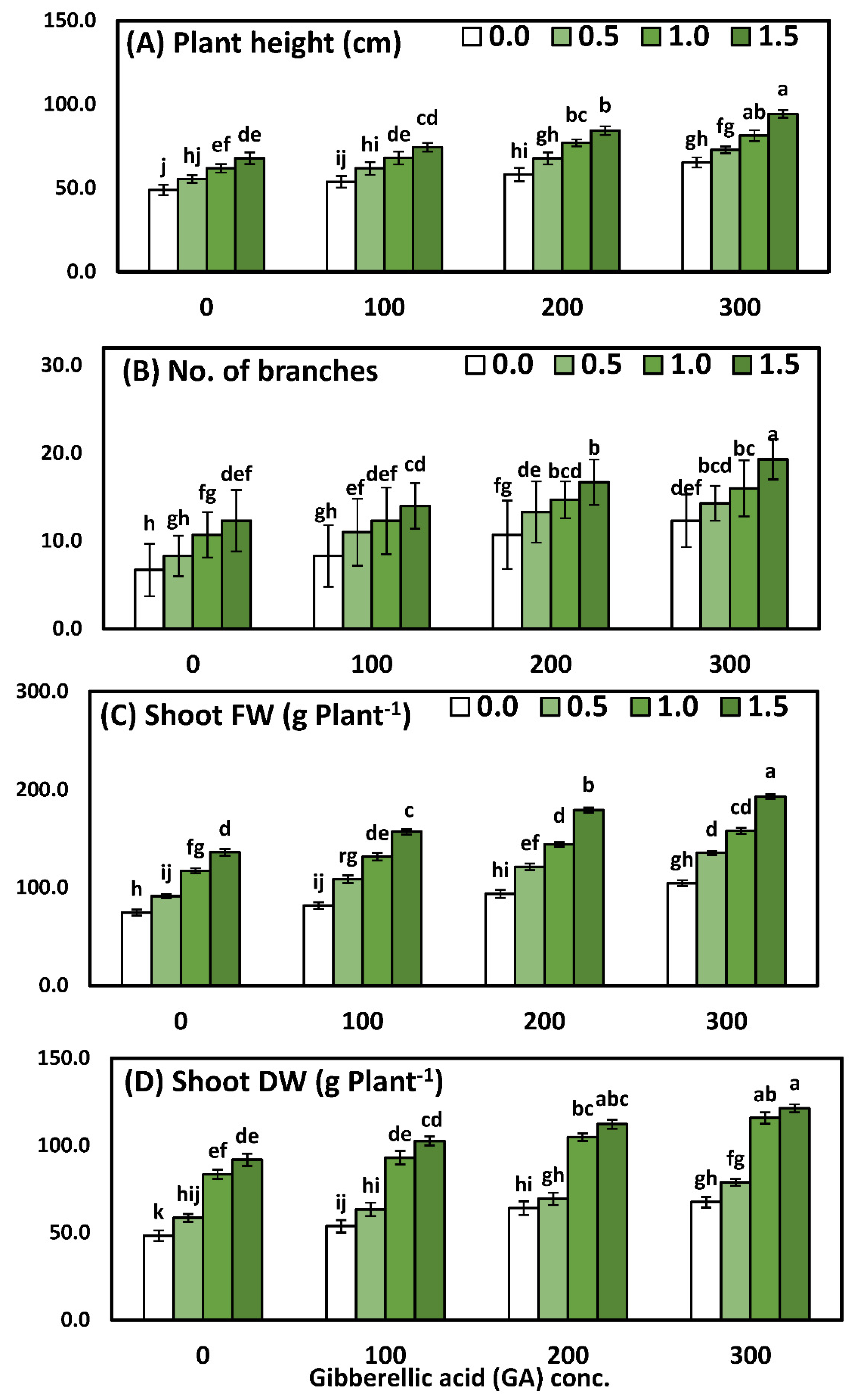
3.1. Effects of YE and GA3 Spraying on the Morphological Measurements of the Solidago Virgaurea Plant, as Well as Their Interactions
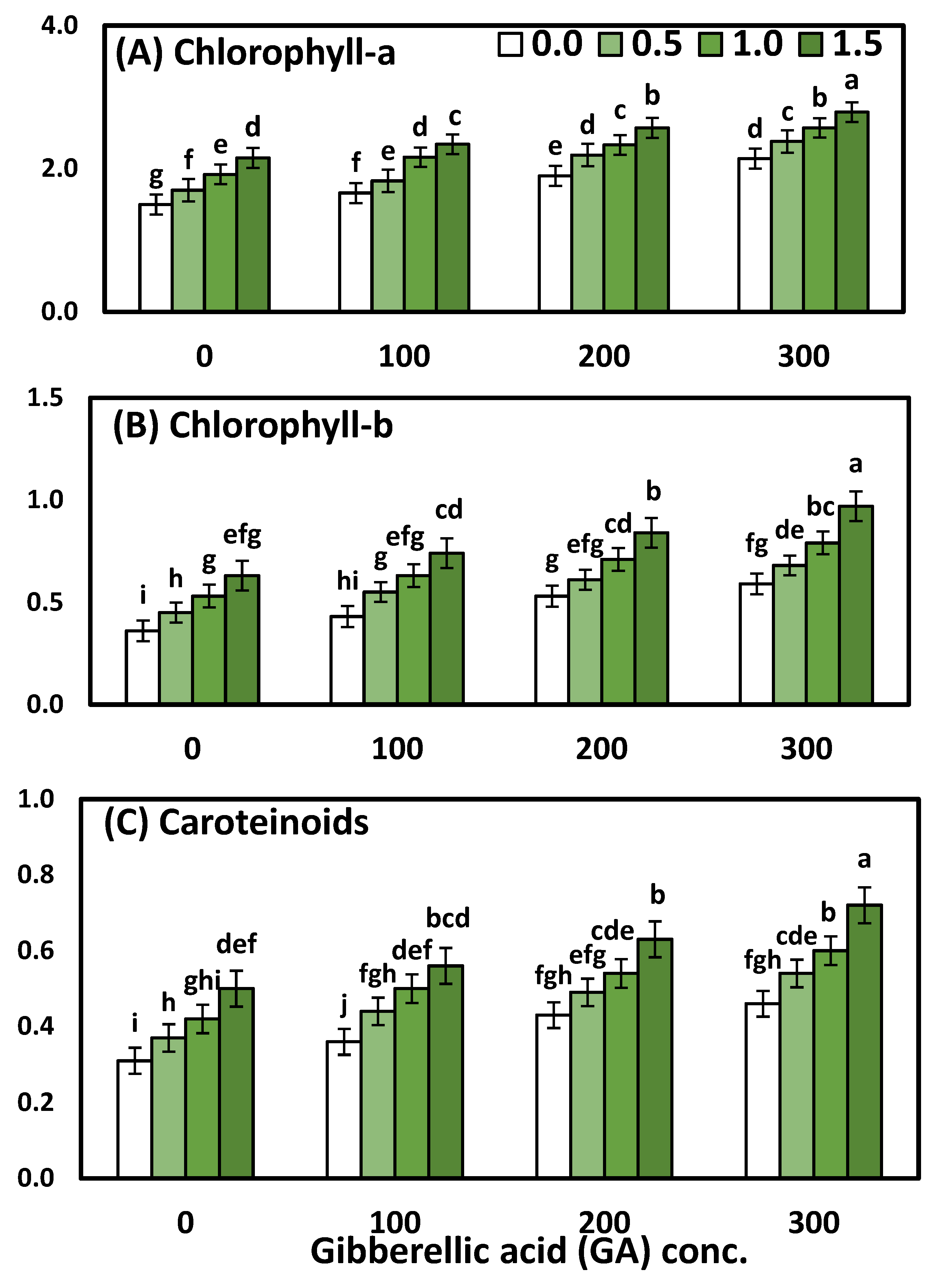
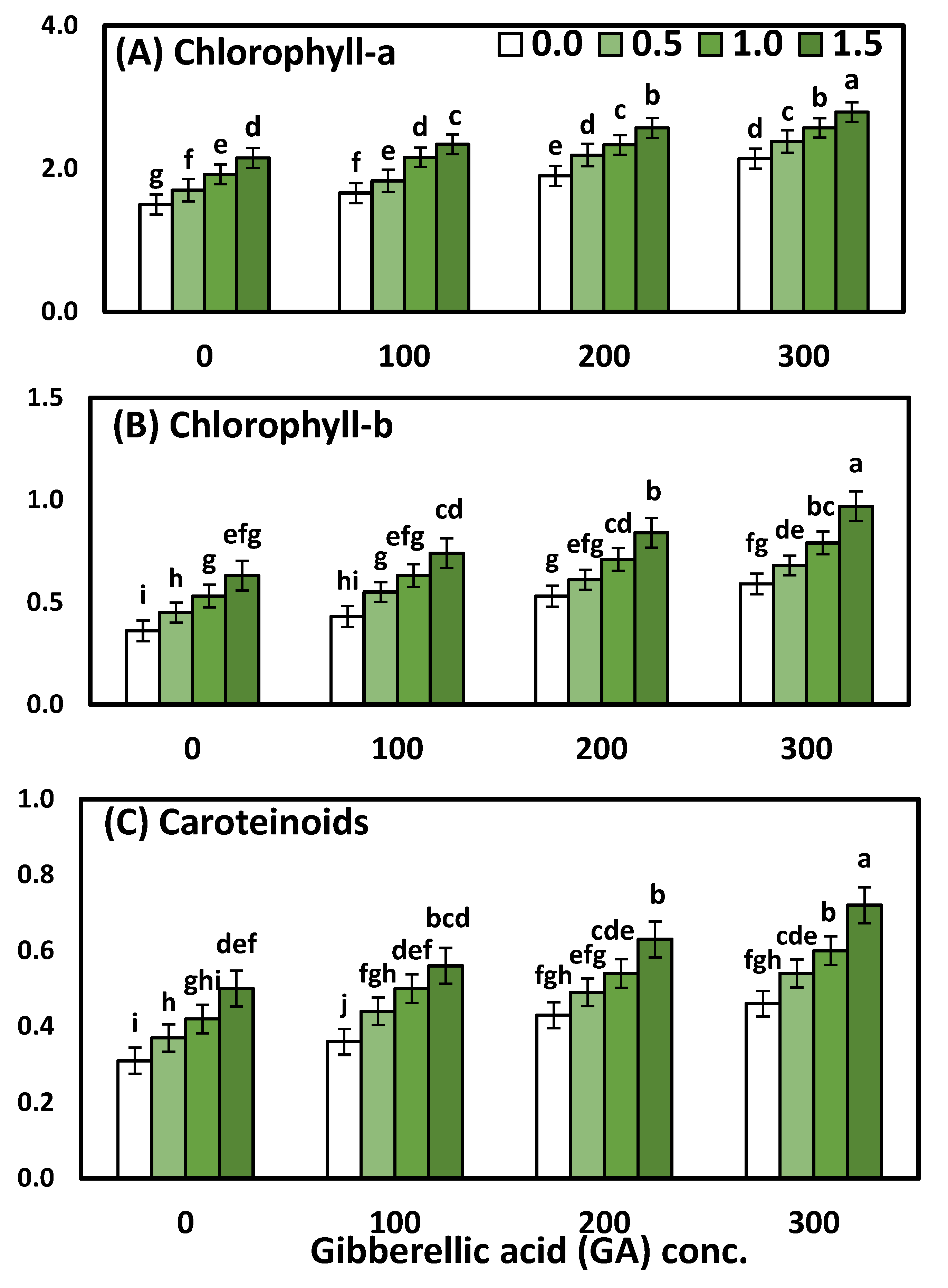
3.2. Effects of YE and GA3 Spraying on the Physiological Measurements of the Solidago Virgaurea Plant, as Well as Their Interactions
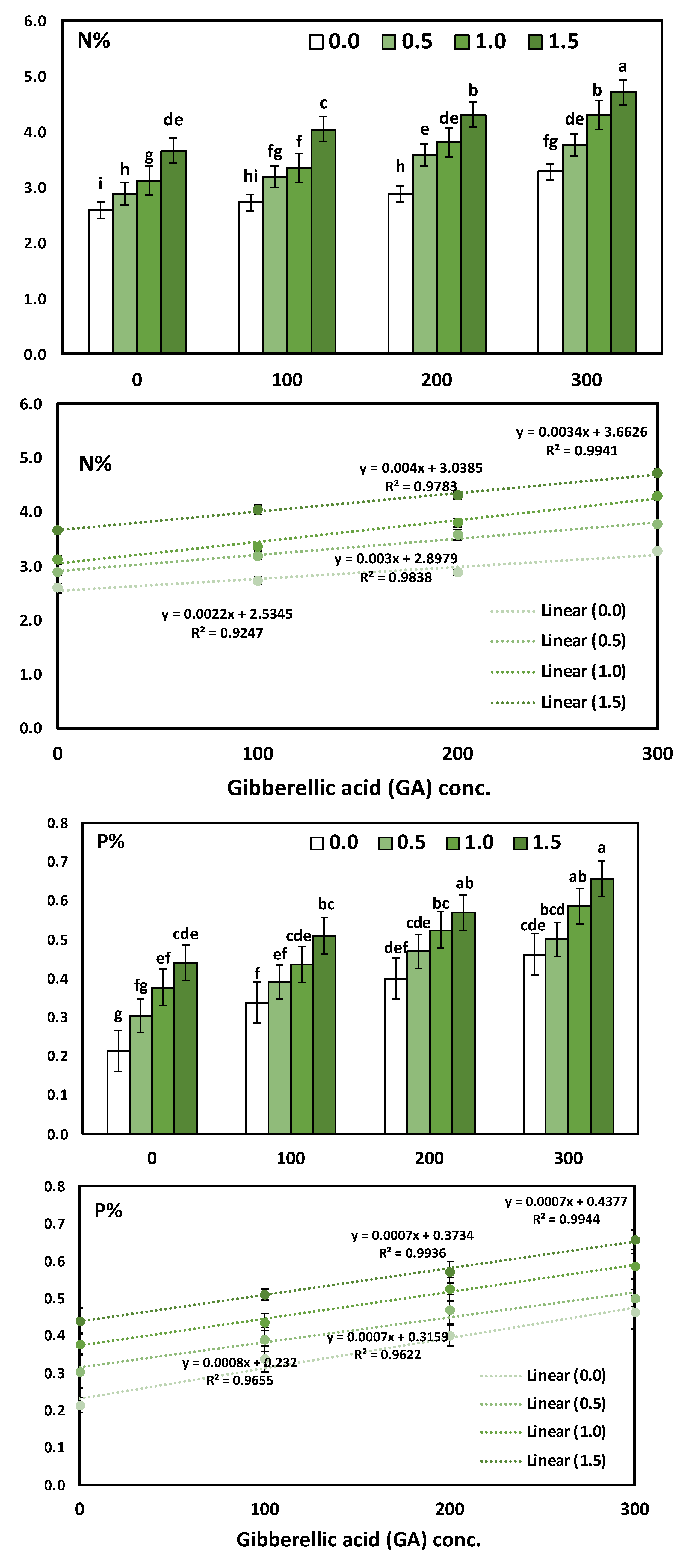
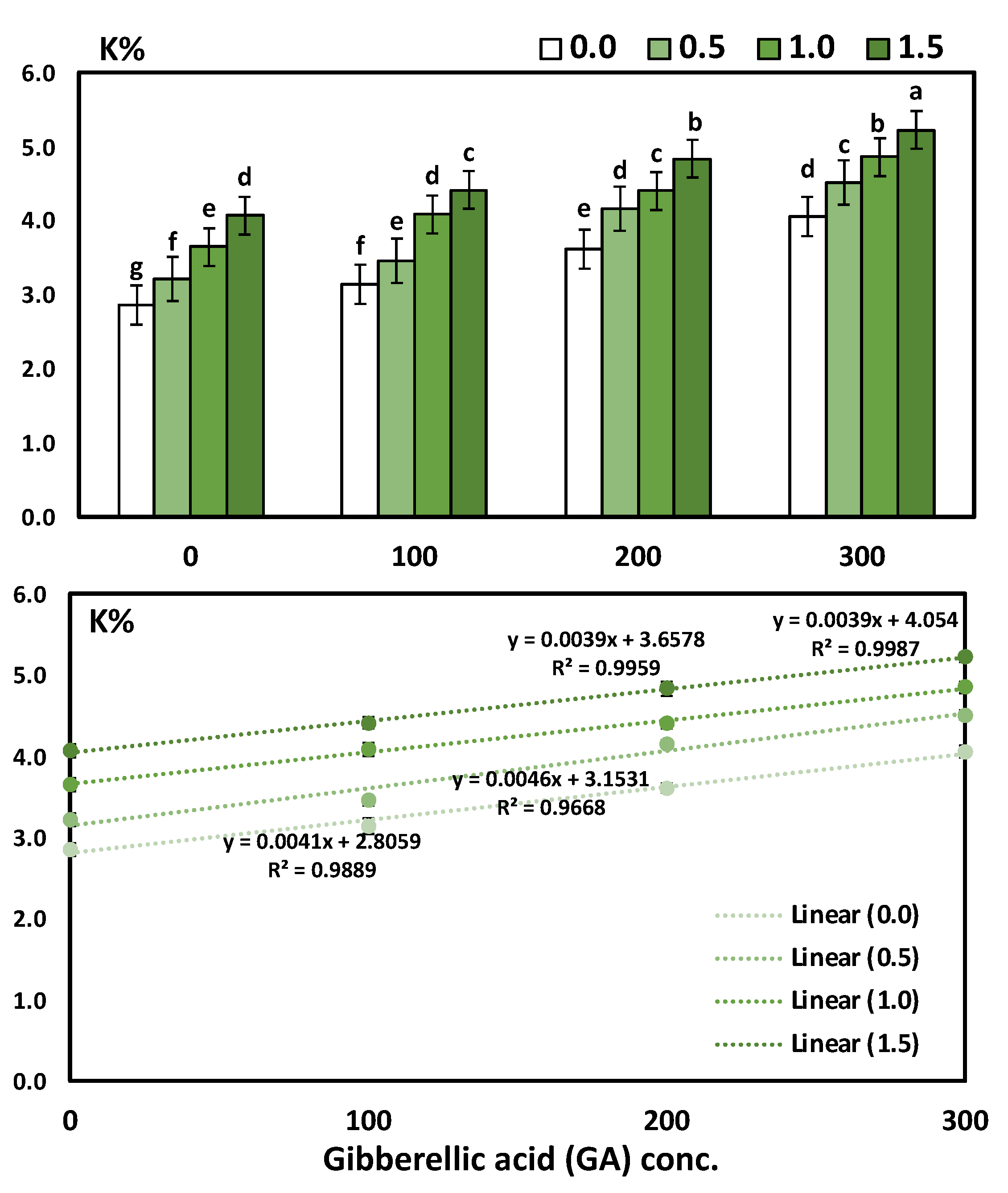
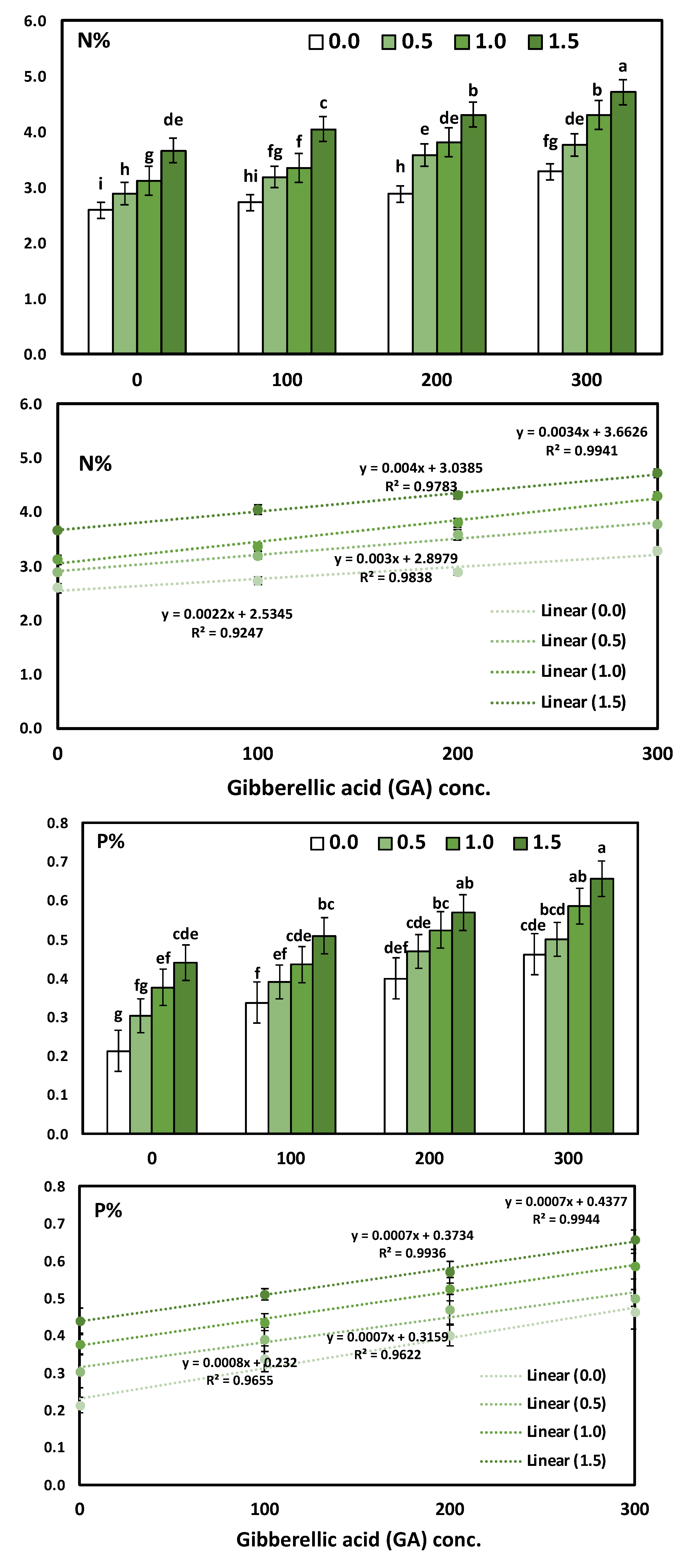
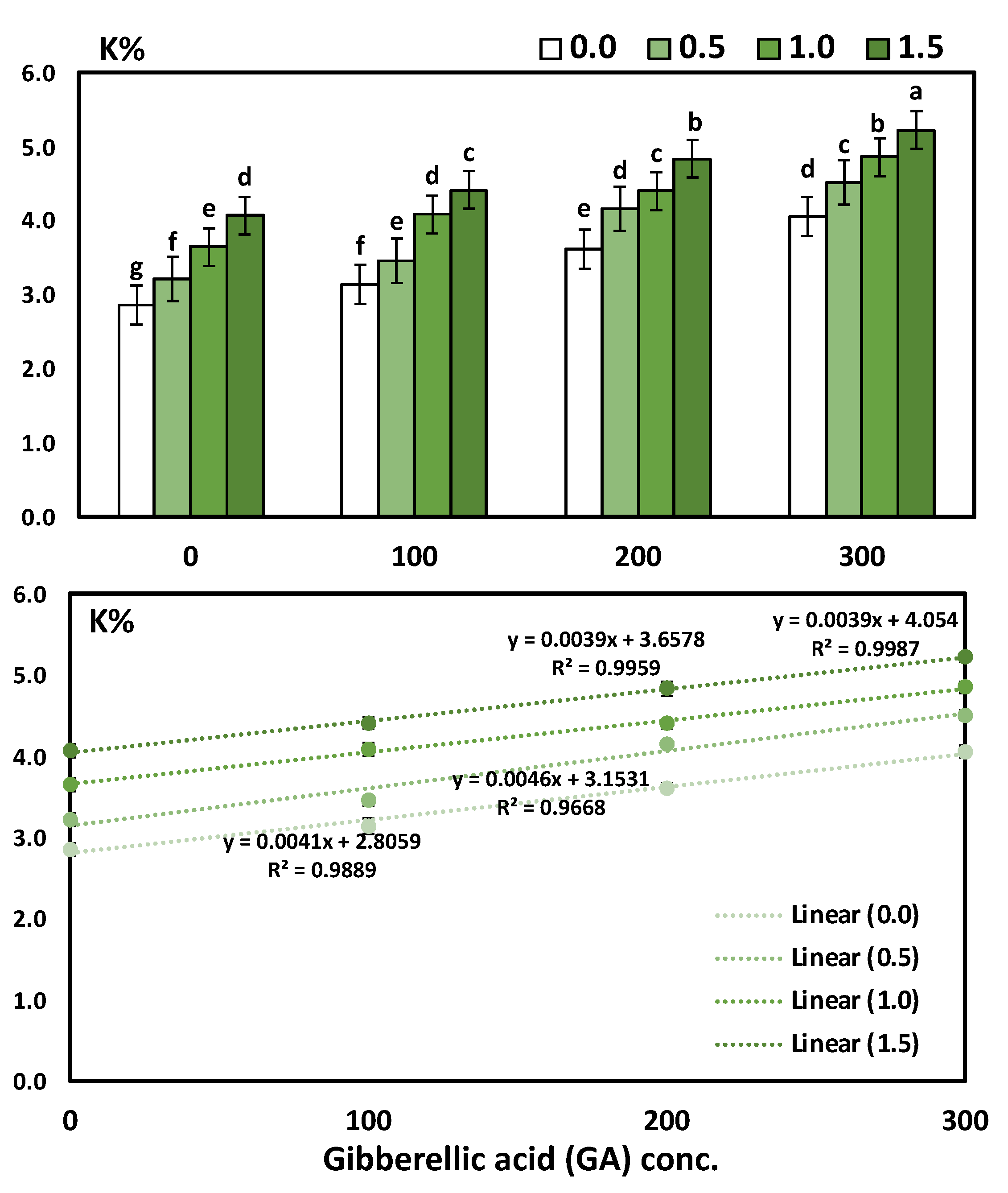
3.3. Effects of YE and GA3 Spraying on the Macronutrient’s Measurements of the Solidago Virgaurea Plant, as Well as Their Interactions
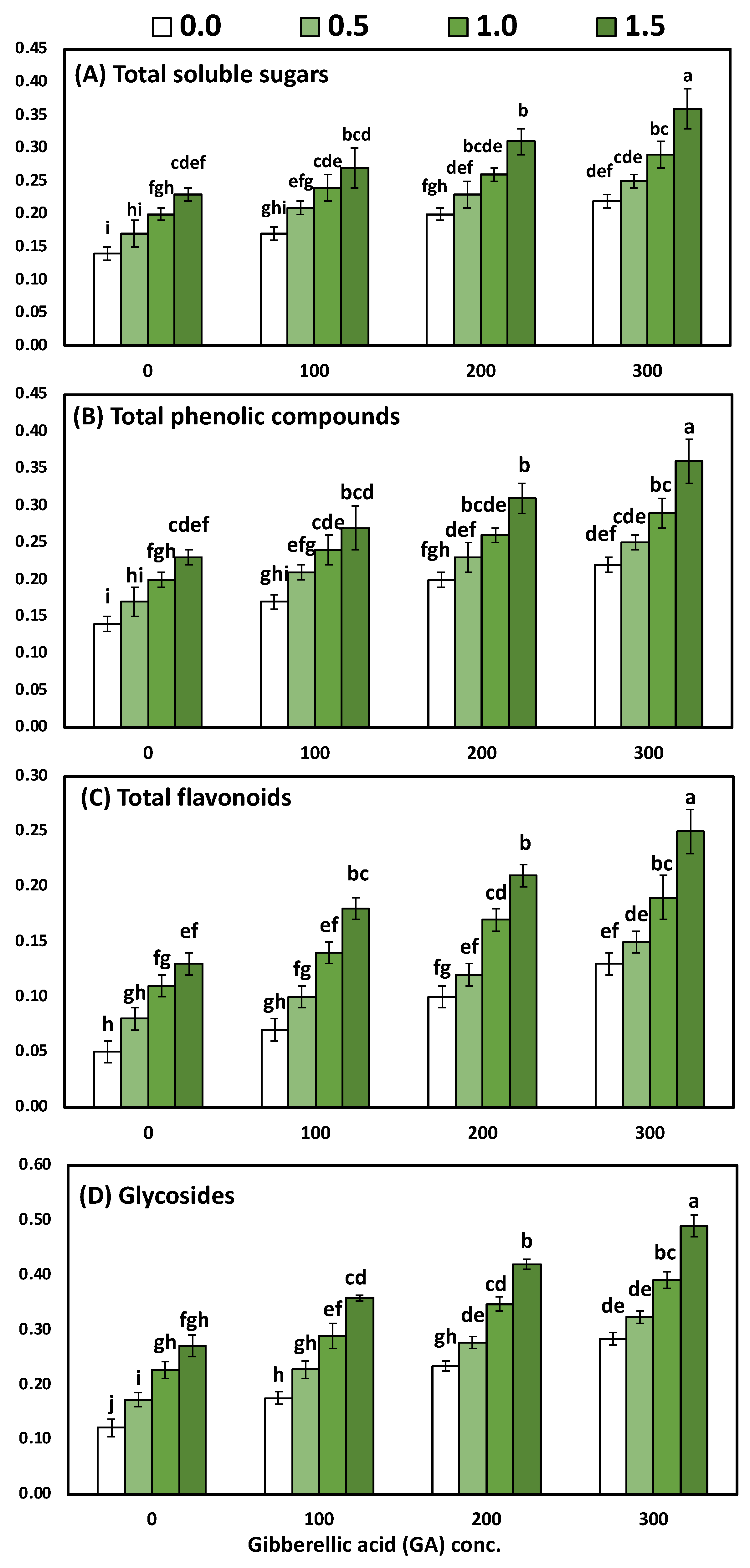
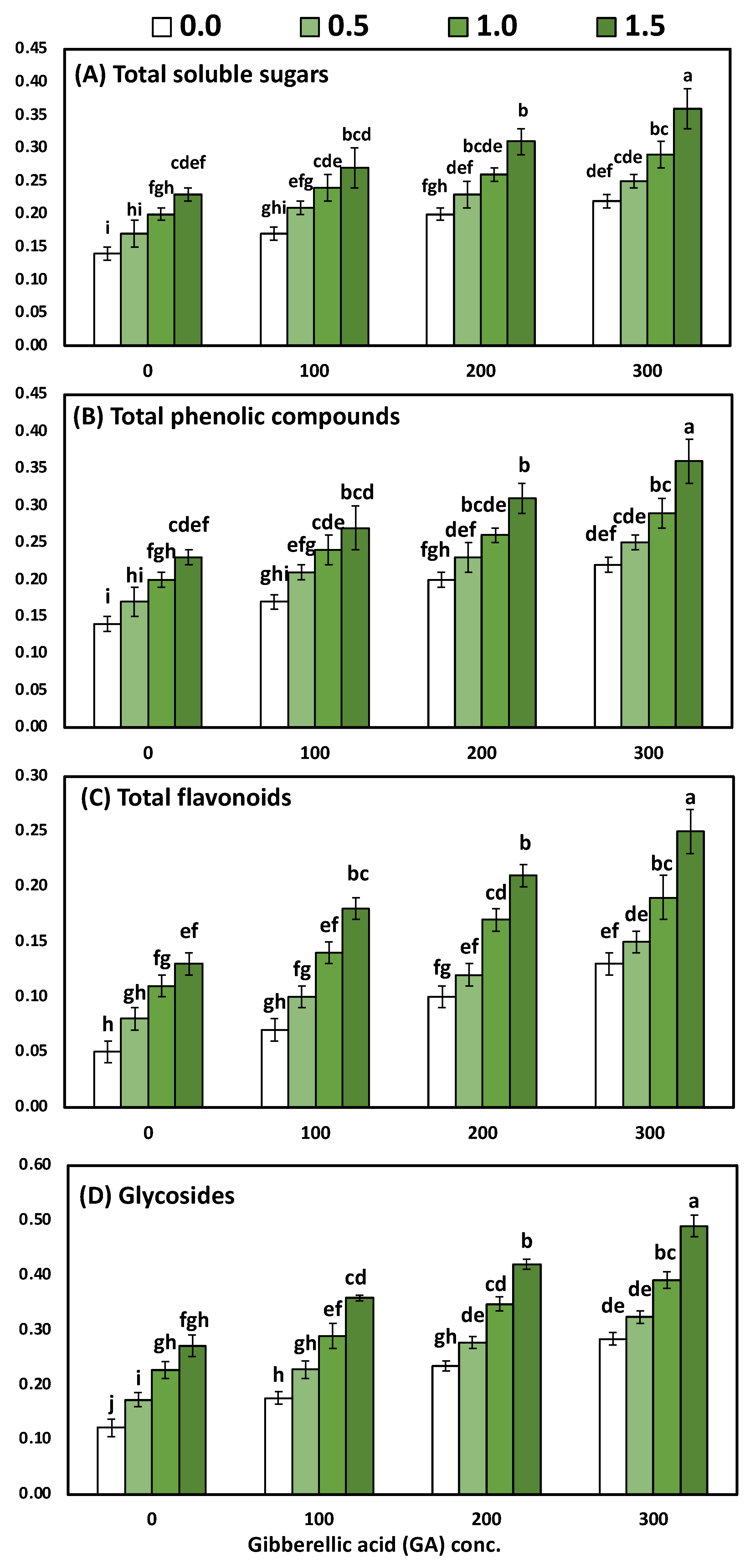
3.4. Effects of YE and GA3 Spraying on the Total Soluble Sugars, Total Phenolic, Total Flavonoids and Total Glycosides Measurements of the Solidago Virgaurea Plant, as Well as Their Interactions
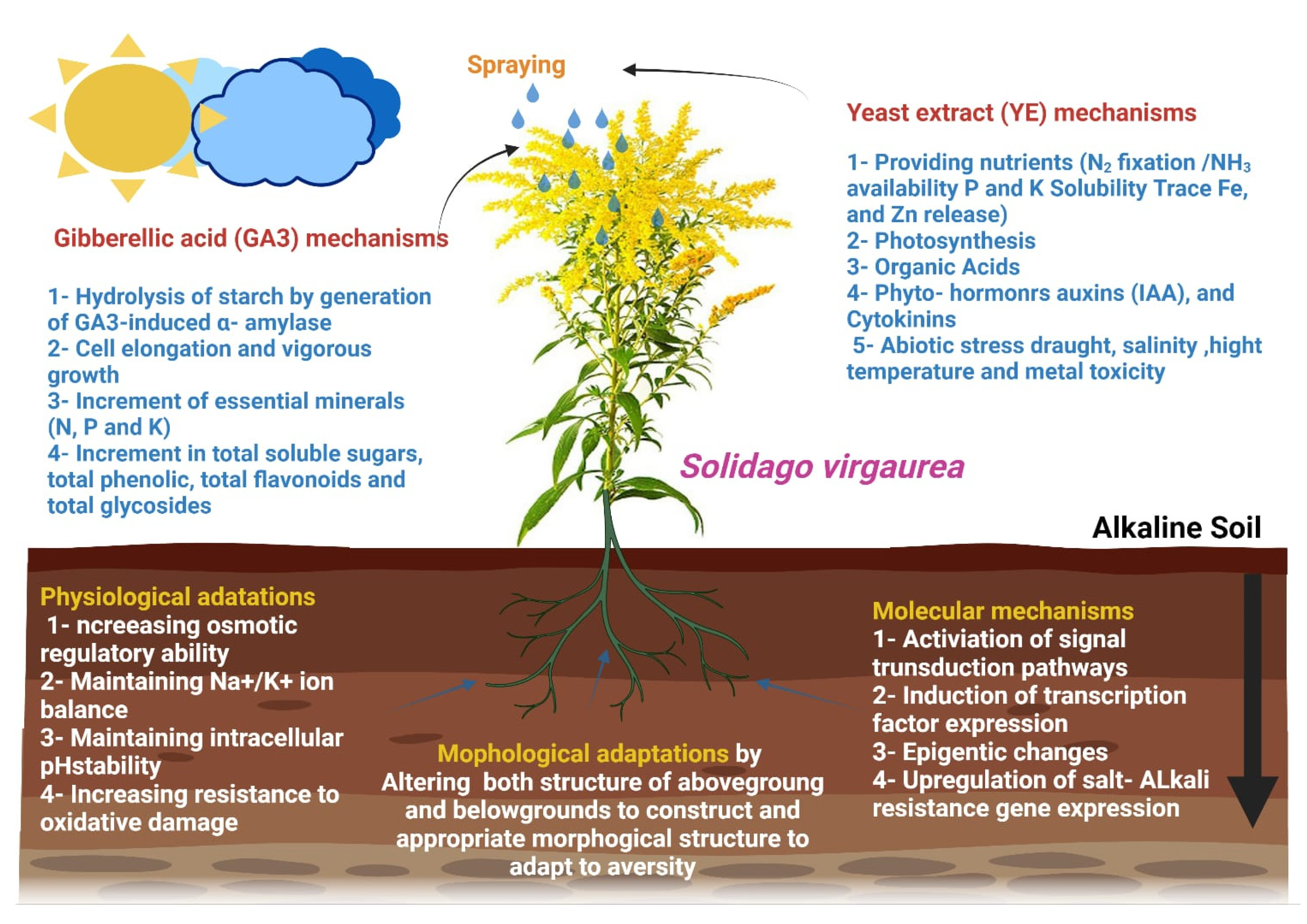
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Kervatkar, N.G.; Raut, V.U.; Chopde, N.; Thakre, S. Effect of spacing and GA3 on growth, flowering and yield of golden rod (Solidago canadensis L.). Pharma Innov. J. 2021, 10, 759–762. [Google Scholar] [CrossRef]
- Toiu, A.; Vlase, L.; Vodnar, D.C.; Gheldiu, A.M.; Oniga, I. Solidago graminifolia L. Salisb. (Asteraceae) as a Valuable Source of Bioactive Polyphenols. HPLC Profile, In Vitro Antioxidant and Antimicrobial Potential. Molecules 2019, 24, 2666. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, D.; Ślusarczyk, S.; Domaradzki, K.; Dry’s, A.; Matkowski, A. Comparison of Polyphenol Profile and Antimutagenic and Antioxidant Activities in Two Species Used as Source of Solidaginis herba—Goldenrod. Chem. Biodivers. 2018, 15, e1800023. [Google Scholar] [CrossRef] [PubMed]
- Motaal, A.A.; Ezzat, S.M.; Tadros, M.G.; El-Askary, H.I. In vivo anti-inflammatory activity of caffeoylquinic acid derivatives from Solidago virgaurea in rats. Pharm. Biol. 2016, 54, 2864–2870. [Google Scholar] [CrossRef]
- Ismail, M.A.; Amin, M.A.; Eid, A.M.; Hassan, S.E.-D.; Mahgoub, H.A.; Lashin, I.; Abdelwahab, A.T.; Azab, E.; Gobouri, A.A.; Elkelish, A. Comparative Study between Exogenously Applied Plant Growth Hormones versus Metabolites of Microbial Endophytes as Plant Growth-Promoting for Phaseolus vulgaris L. Cells 2021, 10, 1059. [Google Scholar] [CrossRef]
- Cai, T.; Caola, I.; Tessarolo, F.; Piccoli, F.; D’Elia, C.; Caciagli, P.; Nollo, G.; Malossini, G.; Nesi, G.; Mazzoli, S. Solidago, orthosiphon, birch and cranberry extracts can decrease microbial colonization and biofilm development in indwelling urinary catheter: A microbiologic and ultrastructural pilot study. World J. Urol. 2014, 32, 1007–1014. [Google Scholar] [CrossRef]
- Gross, S.C.; Goodarzi, G.; Waters, M.; Bandyopadhyay, S.; Pai, S.; Watanabe, K. Antineoplastic activity of Solidago virgaurea on prostate tumor cells in an SCID mouse model. Nutr. Cancer. 2002, 43, 76–81. [Google Scholar] [CrossRef]
- Vuolo, M.M.; Lima, V.S.; Junior, M.R.M. Chapter 2—Phenolic Compounds: Structure, Classification, and Antioxidant Power. In Bioactive Compounds; Rubi, M., Campos, S., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2019; pp. 33–50. [Google Scholar]
- Melzig, M.; Löser, B.; Bader, G.; Papsdorf, G. European goldenrod as anti-inflammatory drug. Z. Für Phyther. 2000, 21, 67–70. [Google Scholar]
- Meyer, B.; Schneider, W.; Elstner, E. Antioxidative properties of alcoholic extracts from Fraxinus excelsior, Populus tremula and Solidago virgaurea. Drug Res. 1995, 45, 174–176. [Google Scholar]
- Soliman, M.H.; Abdulmajeed, A.M.; Alhaithloul, H.; Alharbi, B.M.; El-Esawi, M.A.; Hasanuzzaman, M.; Elkelish, A. Saponin Biopriming Positively Stimulates Antioxidants Defense, Osmolytes Metabolism and Ionic Status to Confer Salt Stress Tolerance in Soybean. Acta Physiol Plant 2020, 42, 114. [Google Scholar] [CrossRef]
- Soliman, M.; Qari, S.H.; Abu-Elsaoud, A.; El-Esawi, M.; Alhaithloul, H.; Elkelish, A. Rapid Green Synthesis of Silver Nanoparticles from Blue Gum Augment Growth and Performance of Maize, Fenugreek, and Onion by Modulating Plants Cellular Antioxidant Machinery and Genes Expression. Acta Physiol Plant 2020, 42, 1–16. [Google Scholar] [CrossRef]
- Alnusairi, G.S.H.; Mazrou, Y.S.A.; Qari, S.H.; Elkelish, A.A.; Soliman, M.H.; Eweis, M.; Abdelaal, K.; El-Samad, G.A.; Ibrahim, M.F.M.; ElNahhas, N. Exogenous Nitric Oxide Reinforces Photosynthetic Efficiency, Osmolyte, Mineral Uptake, Antioxidant, Expression of Stress-Responsive Genes and Ameliorates the Effects of Salinity Stress in Wheat. Plants 2021, 10, 1693. [Google Scholar] [CrossRef]
- Abd-Alla, M.H.; El-Enany, A.E.; Nafady, N.A.; Khalaf, D.M.; Morsy, F.M. Synergistic interaction of Rhizobium leguminosarum bv. viciae and arbuscular mycorrhizal fungi as a plant growth promoting biofertilizers for faba bean (Vicia faba L.) in alkaline soil. Microbiol. Res. 2014, 169, 49–58. [Google Scholar] [CrossRef]
- Ibrahim, M.F.M.; Abd El-Samad, G.; Ashour, H.; El-Sawy, A.M.; Hikal, M.; Elkelish, A.; El-Gawad, H.A.; El-Yazied, A.A.; Hozzein, W.N.; Farag, R. Regulation of Agronomic Traits, Nutrient Uptake, Osmolytes and Antioxidants of Maize as Influenced by Exogenous Potassium Silicate under Deficit Irrigation and Semiarid Conditions. Agronomy 2020, 10, 1212. [Google Scholar] [CrossRef]
- Shi, D.; Sheng, Y. Effect of various salt–alkaline mixed stress conditions on sunflower seedlings and analysis of their stress factors. Environ. Exp. Bot. 2005, 54, 8–21. [Google Scholar] [CrossRef]
- Slam, M.S.; Akhter, M.M.; El Sabagh, A.; Liu, L.Y.; Nguyen, N.T.; Ueda, A.; Masaoka, Y.; Saneoko, H. Comparative studies on growth and physiological responses to saline and alkaline stresses of Foxtail millet (Setaria italica L.) and Proso millet (Panicum miliaceum L.). Aust. J. Crop. Sci. 2011, 5, 1269–1277. [Google Scholar]
- Al-Abadi, J.S.; Salih, H.M.; Saadoun, H.S. Trace elements and their uses as a spray on all agricultural crops. Gen. Auth. Agric. Ext. Coop. Ext. Bull. 2007, N, 41. [Google Scholar]
- Taha, L.S.; Ibrahim, S.M.M.; Abdel Aziz, N.G. Vegetative growth, chemical composition, and flavonoids content of Azadirachta indica plants as affected by application of yeast natural extract. J. Appl. Pharm. Sci. 2016, 6, 93–97. [Google Scholar] [CrossRef]
- Ghada, M.R.; El-Shawa; Rashwan, E.M.; Abdelaal, K.A. Mitigating salt stress effects by exogenous application of proline and yeast extract on morphophysiological, biochemical and anatomical characters of calendula plants. Sci. J. Flowers Ornam. Plants. 2020, 7, 461–482. [Google Scholar]
- Engy, A.S.; Taha, Z.K. Enhancement of biometrics, anatomical characteristics and fatty acids profile of Sunflower plants by foliar application with yeast extract and salicylic acid. Middle East J. Agric. Res. 2019, 8, 1271–1280. [Google Scholar]
- Khudair, T.Y.; Hajam, A.J. The effect of yeast extract and organic fertilizer algidex spray on the vegetative, root and floral growth of the Chinese carnation. Plant Cell Biotechnol. Mol. Biol. 2021, 22, 165–173. [Google Scholar]
- Hassanain, M.A.; Osman, A.R.; Eman, S.; El-Dein, M.S. Influence of nitrogen and yeast on production of Marigold plants. World Rural Observ. 2018, 10, 37–46. [Google Scholar]
- Taha, R.S.; Selemian, M.F.; Alhammad, B.A.; Alkahtani, J.; Alwahibi, M.S.; Mahdi, A.H.A. Activated yeast extract enhances growth, anatomical structure, and productivity of Lupinus termis L. Plants under Actual Salinity Conditions. Agronomy 2021, 11, 74. [Google Scholar] [CrossRef]
- Singh, J.; Nigam, R.; Singh, R.; Kumar, A.; Kumar, A. Effect of gibberellic acid and cycocel on growth, flowering and yield of chrysanthemum (Dendranthema grandiflora Ramat.) cv. Birbal Sahni. J. Pharmacogn. Phytochem. 2018, 7, 2753–2758. [Google Scholar]
- Roy, R.; Nasiruddin, K.M. Effect of different level of GA3 on growth and yield of cabbage. Int. J. Environ. Sci. Nat. Resour. 2011, 4, 79–82. [Google Scholar]
- Sure, S.; Arooie, H.; Azizi, M. Influence of plant growth regulators (PGRs) and planting method on growth and yield in oil pumpkin (Cucurbita pepo var. styriaca). Not. Sci. Biol. 2012, 4, 101–107. [Google Scholar] [CrossRef]
- Baghele, P.; Raut, V.U.; Chopda, N.; Goramnagar, H.B. Response of golden rod (Solidago canadensis L.) to planting time and GA3. J. Pharmacogn. Phytochem. 2020, 9, 565–568. [Google Scholar]
- Narute, T.T.; Parulekar, Y.R.; Narute, T.K. Effect of plant growth regulators on yield and yield attributing character of Marigold cv. Calcutta Marigold under Konkan conditions. Int. J. Curr. Microbiol. App. Sci. 2020, 9, 3998–4005. [Google Scholar] [CrossRef]
- Klute, A. Methods of Soil Analysis, Part 1: Physical and Mineralogical Methods, 2nd ed.; American Society of Agronomy: Madison, WI, USA, 1986. [Google Scholar]
- Page, A.L.; Miller, R.H.; Kenney, D.R. Methods of Soil Analysis: Chemical and Microbiological Properties, Part 2; American Society of Agronomy: Madison, WI, USA, 1982; pp. 39–41. [Google Scholar]
- Abou El-Yazied, A.; Mady, M.A. Effect of boron and yeast extract foliar application on growth pod setting and both green pod and seed yield of broad bean (Cicia faba L). J. App. Sci. Res. 2012, 8, 1240–1251. [Google Scholar]
- Moran, R.; Porath, D. Chlorophyll determination in intact tissues using n,n-dimethylformamide. Plant Physiol. 1980, 65, 478–479. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophyll a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Rady, M.M. A novel organo-mineral fertilizer can mitigate salinity stress effects for tomato production on reclaimed saline soil. S. Afr. J. Bot. 2012, 81, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.L. Soil Chemical Analysis; Prentice Hall, Inc.: Hoboken, NJ, USA, 1973; p. 448. [Google Scholar]
- Irigoyen, J.J.; Emerich, D.W.; Sanchez-Diaz, M. Water stress induced changes in the concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Plant Physiol. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic–phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Shi, J.I.Y.; Zou, X.B.; Zhao, J.W.; Mel, H.; Wang, K.L.; Wang, X.; Chen, H. Determination of total flavonoids content in fresh Ginkgo biloba leaf with different colors using near infrared spectroscopy. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2012, 94, 271–276. [Google Scholar] [CrossRef]
- Zhao, L.; Liu, W.; Xiong, S.; Tang, J.; Lou, Z.; Xie, M.; Xia, B.; Lin, L.; Liao, D. Determination of total flavonoids contents and antioxidant activity of Ginkgo biloba leaf by near-infrared reflectance method. Int. J. Anal. Chem. 2018, 2018, 8195784. [Google Scholar] [CrossRef]
- Solich, P.; Sedliakova, V.; Karlicek, R. Spectrophotometric determination of cardiac glycosides by flow-injection analysis. Anal. Chim. Acta. 1992, 269, 199–203. [Google Scholar] [CrossRef]
- Tofighi, Z.; Ghazi Saeidi, N.; Hadjiakhoondi, A.; Yassa, N. Determination of cardiac glycosides and total phenols in different generations of Securigera securidaca suspension culture. Res. J. Pharmacogn. 2016, 3, 25–31. [Google Scholar]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 8th ed.; Affiliated East-West Press: Delhi, India; Iowa State University Press: Ames, IA, USA, 1994. [Google Scholar]
- Yang, J.Y.; Zheng, W.; Tian, Y.; Wu, Y.; Zhou, D.W. Effects of various mixed salt-alkaline stresses on growth, photosynthesis, and photosynthetic pigment concentrations of Medicago ruthenica seedlings. Photosynthetica 2011, 49, 275–284. [Google Scholar] [CrossRef]
- Abbas, S.M. The influence of biostimulants on the growth and on the biochemical composition of Vicia faba cv. Giza 3 beans. Rom. Biotechnol. Lett. 2013, 18, 8061–8068. [Google Scholar]
- Sarhan, A.M.Z.; Habib, A.M.; Mahmoud, A.N.; Noor El-Deen, T.M. Effect of nano, bio, chemical fertilization and leaves extract of moringa plant on flowering and chemical constituents of gladiolus plant. Egypt. J. Chem. 2022, 65, 221–230. [Google Scholar] [CrossRef]
- Dawood, M.G.; El-Lethy, S.R.; Sadak, M.S. Role of methanol and yeast in improving growth, yield, nutritive value and antioxidants of Soybean. World Appl. Sci. J. 2013, 26, 6–14. [Google Scholar]
- Warring, P.E.; Phillips, I.D.G. The Control of Growth and Differentiation in Plants; Pergamon Press Ltd.: Oxford, UK, 1973. [Google Scholar]
- Shafeek, M.R.; Helmy, Y.I.; Shalaby, M.A.; Omer, N.M. Response of onion plants to foliar application of sources and levels of some amino acids under sandy soil conditions. J. Appl. Sci. Res. 2012, 8, 5521–5527. [Google Scholar]
- Olaiya, C.O. Presowing bioregulator seed treatments increase the seedling growth and yield of tomato (Lycopersicon esculentum). J. Plant Growth Regul. 2010, 29, 349–356. [Google Scholar] [CrossRef]
- Alavi Mehryan, S.M.; Zare, N.; Masumiasl, A.; Sheikhzadeh, P.; Asghari, R. Effect of Salicylic Acid and Yeast Extract Elicitors on the Expression of HMGR and GPPS Genes Involved in Biosynthesis of Terpenes in Medicinal Plant Ferulago angulata under Cell Suspension Culture Condition. Plant Genet. Res. 2020, 7, 63–76. [Google Scholar] [CrossRef]
- Al-Juthery, H.W.A.; Ali, E.A.H.M.; Al-Ubori, R.N.; NAl-Shami, Q.M.; AL-Taey, D.K.A. Role of foliar application of nano npk, micro fertilizers and yeast extract on growth and yield of wheat. Int. J. Agricult. Stat. Sci. 2020, 6, 1295–1300. [Google Scholar]
- Abraham, F.; Bhatt, A.; Keng, C.L.; Indrayanto, G.; Sulaiman, S.F. Effect of yeast extract and chitosan on shoot proliferation, morphology and antioxidant activity of Curcuma mangga in vitro plantlets. Afr. J. Biotechnol. 2011, 10, 7787–7795. [Google Scholar]
- Abdallah, M.M.S.; El Habbasha, S.F.; El Sebai, T. Comparison of yeast extract and Nicotinaminde foliar applications effect on quinoa plants grown under sandy soil condition. Int. J. Pharmtech Res. 2016, 9, 24–32. [Google Scholar]
- Nassar, R.M.A.; Shanan, N.T.; Reda, F.M. Active yeast extract counteracts the on harmful effects of salinity stress the growth of leucaena plant. Sci. Hortic. 2016, 201, 61–67. [Google Scholar] [CrossRef]
- Dawood, M.G.; Sadak, M.S.; Abdallah, M.S.; Bakry, A.B.; Darwish, M.O. Influence of biofertilizers on growth and some biochemical aspects of flax cultivars grown under sandy soil conditions. Bull. Natil. Res. Cent. 2019, 43, 81. [Google Scholar] [CrossRef]
- Pal, P.; Yadav, K.; Kumar, K.; Singh, N. effect of gibberellic acid and potassium foliar sprays on productivity and physiological and biochemical parameters of Parthenocarpic cucumber cv. ‘seven star f1’. J. Hortic. Res. 2016, 24, 93–100. [Google Scholar] [CrossRef]
- Sajid, M.; Amin, N.; Ahmad, H.; Khan, K. Effect of gibberellic acid on enhancing flowering time in Chrysanthemum morifolium. Pak. J. Bot. 2016, 48, 477–483. [Google Scholar]
- Tiwari, H.; Kumar, M.; Naresh, R.K.; Singh, M.K.; Malik, S.; Singh, S.P.; Chaudhary, V. Effect of organic and inorganic fertilizers with foliar application of gibberellic acid on productivity, profitability and soil health of marigold (Tagetes erecta l.) cv. pusa narangi gainda. Int. J. Agricult. Stat. Sci. 2018, 14, 575–585. [Google Scholar]
- Janowska, B.; Andrzejak, R. Effect of gibberellic acid spraying and soaking of rhizomes on the growth and flowering of calla lily (Zantedeschia spreng). Acta. Agrobot. 2010, 63, 155–160. [Google Scholar] [CrossRef]
- Ibrahim, S.M.M.; Taha, L.S.; Farahat, M.M. Vegetative growth and chemical constituents of croton plants as affected by foliar application of benzyl adenine and gibberellic acid. J. Am. Sci. 2010, 6, 126–130. [Google Scholar]
- Ali, M.; Abbasi, B.H. Thidiazuron-induced changes in biomass parameters, total phenolic content, and antioxidant activity in callus cultures of Artemisia absinthium L. Appl. Biochem. Biotechnol. 2014, 172, 2363–2376. [Google Scholar] [CrossRef]
- Sarrou, E.; Chatzopoulou, P.; Dimassi-Theriou, K.; Therios, I.; Koularmani, A. Effect of melatonin, salicylic acid and gibberellic acid on leaf essential oil and other secondary metabolites of bitter orange young seedlings. J. Essent. Oil Res. 2015, 27, 487–496. [Google Scholar] [CrossRef]
- Zarrin, N.; Shoor, M.; Tehranifar, A.; Karimian, Z. Effect of day length and gibberrelic acid on some morphological and biochemical characteristics of chrysanthemum. Ir. J. Hortic. Sci. 2021, 51, 979–993. [Google Scholar]
- Hassan, A.A.; Abd El-Azeim, M.M. Gibberellic acid usage relieving salinity adverse effects on growth, flowering and bulb production of tuberose plants (Polianthes tuberosa L.). J. Plant Prod. Mansoura Univ. 2019, 10, 1051–1058. [Google Scholar] [CrossRef]
- Wilde, S.A.; Corey, R.B.; Lyer, J.G.; Voigt, G.K. Soil and Plant Analysis for Tree Culture, 3rd ed.; Oxford IBM Publishing: New Delhi, India, 1985; pp. 93–106. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Value | |
|---|---|---|
| 2018 | 2019 | |
| Particle size distribution | ||
| Coarse sand % | 7.40 | 7.44 |
| Fine sand % | 41.61 | 41.80 |
| Silt % | 18.17 | 18.23 |
| Clay % | 32.82 | 32.80 |
| Texture class | Sandy clay loam | |
| PH | 7.82 | 7.81 |
| ECe (dS/m) | 3.42 | 3.40 |
| Soluble cations (meq 100/g soil) | ||
| Ca2+ | 7.92 | 7.94 |
| Mg2+ | 6.80 | 6.90 |
| Na+ | 15.27 | 15.33 |
| K+ | 0.48 | 0.45 |
| Soluble anions (meq /100 g soil) | ||
| Cl− | 15.98 | 15.95 |
| HCO3− | 0.86 | 0.85 |
| SO42− | 17.27 | 17.25 |
| CO32− | 0.00 | 0.00 |
| Nutrients available | ||
| N% | 0.18 | 0.20 |
| P% | 0.22 | 0.24 |
| K% | 0.32 | 0.30 |
| Amino Acid % | Vitamins (mg/100 g DW) | Minerals | |||
|---|---|---|---|---|---|
| Alanine | 1.69 | Vit.B1 | 23.33 | Nitrogen | 6.88% |
| Arginine | 1.49 | Vit.B2 | 21.04 | Phosphorus | 0.66% |
| Aspartic acid | 2.32 | Vit.B6 | 20.67 | potassium | 0.95% |
| Cystine | 0.63 | Vit.B12 | 19.17 | Magnesium | 0.19% |
| Glutamic acid | 3.76 | Thiamine | 23.21 | Calcium | 0.17% |
| Glycine | 1.45 | Riboflavin | 27.29 | Sulphur | 0.48% |
| Histidine | 0.71 | Inositol | 20.43 | Iron | 107 ppm |
| Isoleucine | 0.85 | Biotin | 20.04 | Zinc | 77 ppm |
| Leucine | 1.91 | Nicotinic acid | 73.92 | Copper | 5 ppm |
| Lysine | 1.13 | Pantothenic acid | 38.43 | Manganese | 13 ppm |
| Phenylalanine | 1.18 | P aminobenzoic acid | 29.49 | Growth regulators (ppm) | |
| Proline | 1.29 | Folic acid | 26.22 | Adenine | 31 |
| Serine | 1.98 | Pyridoxine | 22.09 | Betaines | 56 |
| Threonine | 1.54 | Others % | |||
| Tryptophan | 0.25 | Crude Protein | 43.00 | ||
| Tyrosine | 0.99 | Crude Fat | 2.20 | ||
| Valine | 1.4 | Carbohydrates | 33.21 | ||
| Methionine | 0.4 | Crude Fibre | 7.20 | ||
| Ash | 3.80 | ||||
| Treatments | Plant Height (cm) | No. of Branches | Weight of Shoot System (g Plant−1) | |
|---|---|---|---|---|
| FW | DW | |||
| YE (g/L) | ||||
| 0.0 (Control) | 56.5 ± 2.9 d | 9.5 ± 0.8 d | 88.7 ± 4.1 d | 58.4 ± 3.9 d |
| 0.5 | 64.3 ± 3.4 c | 11.8 ± 0.8 c | 114.3 ± 4.3 c | 67.5 ± 4.0 c |
| 1.0 | 72.0 ± 3.0 b | 13.4 ± 0.8 b | 137.9 ± 4.1 b | 99.2 ± 4.4 b |
| 1.5 | 80.2 ± 2.6 a | 15.6 ± 0.8 a | 166.6 ± 4.4 a | 107.0 ± 3.8 a |
| ANOVA-1-way | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** |
| GA (ppm) | ||||
| 0.0 (Control) | 58.4 ± 3.4 d | 9.5 ± 0.9 d | 104.9 ± 4.3 d | 70.5 ± 4.4 d |
| 100 | 64.4 ± 2.9 c | 11.4 ± 0.7 c | 119.8 ± 3.9 c | 78.2 ± 3.4 c |
| 200 | 71.8 ± 2.9 b | 13.8 ± 0.8 b | 134.7 ± 4.1 b | 87.6 ± 4.2 b |
| 300 | 78.4 ± 2.8 a | 15.5 ± 0.8 a | 148.0 ± 4.5 a | 95.9 ± 4.1 a |
| ANOVA-1-way | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** |
| Two-way analysis of variance | ||||
| Corr. model | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** |
| YE | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** |
| GA | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** |
| Treatments | Chl. a | Chl. b | Total Carotenoids |
|---|---|---|---|
| mg mm−2 | |||
| YE (g/L) | |||
| 0.0 (Control) | 1.80 ± 0.04 d | 0.48 ± 0.02 d | 0.39 ± 0.02 d |
| 0.5 | 2.02 ± 0.04 c | 0.57 ± 0.02 c | 0.46 ± 0.02 c |
| 1.0 | 2.25 ± 0.03 b | 0.67 ± 0.02 b | 0.52 ± 0.02 b |
| 1.5 | 2.46 ± 0.03 a | 0.80 ± 0.03 a | 0.60 ± 0.03 a |
| ANOVA-1-way | <0.001 *** | <0.001 *** | <0.001 *** |
| GA (ppm) | |||
| 0.0 (Control) | 1.82 ± 0.04 d | 0.49 ± 0.02 d | 0.40 ± 0.02 d |
| 100 | 2.00 ± 0.03 c | 0.59 ± 0.03 c | 0.46 ± 0.03 c |
| 200 | 2.25 ± 0.04 b | 0.67 ± 0.02 b | 0.52 ± 0.02 b |
| 300 | 2.47 ± 0.04 a | 0.76 ± 0.02 a | 0.58 ± 0.02 a |
| ANOVA-1-way | <0.001 *** | <0.001 *** | <0.001 *** |
| Two-way analysis of variance | |||
| Corr. model | <0.001 *** | <0.001 *** | <0.001 *** |
| YE | <0.001 *** | <0.001 *** | <0.001 *** |
| GA | <0.001 *** | <0.001 *** | <0.001 *** |
| Treatments | N | P | K |
|---|---|---|---|
| % | |||
| YE (g/L) | * | * | * |
| 0.0 (Control) | 2.87 ± 0.06 d | 0.35 ± 0.03 d | 3.42 ± 0.08 d |
| 0.5 | 3.35 ± 0.07 c | 0.42 ± 0.03 c | 3.84 ± 0.08 c |
| 1.0 | 3.64 ± 0.07 b | 0.48 ± 0.03 b | 4.25 ± 0.06 b |
| 1.5 | 4.18 ± 0.07 a | 0.54 ± 0.03 a | 4.64 ± 0.06 a |
| ANOVA-1-way | <0.001 *** | <0.001 *** | <0.001 *** |
| 0.0 (Control) | 3.06 ± 0.07 d | 0.33 ± 0.03 d | 3.45 ± 0.07 d |
| 100 | 3.32 ± 0.06 c | 0.42 ± 0.04 c | 3.78 ± 0.06 c |
| 200 | 3.64 ± 0.08 b | 0.49 ± 0.03 b | 4.25 ± 0.07 b |
| 300 | 4.01 ± 0.07 a | 0.55 ± 0.03 a | 4.66 ± 0.07 a |
| ANOVA-1-way | <0.001 *** | <0.001 *** | <0.001 *** |
| Two-way analysis of variance | |||
| Corr. model | <0.001 *** | <0.001 *** | <0.001 *** |
| YE | <0.001 *** | <0.001 *** | <0.001 *** |
| GA | <0.001 *** | <0.001 *** | <0.001 *** |
| Treatments | Total Soluble Sugars | Total Phenolic | Total Flavonoids | Total Glycosides |
|---|---|---|---|---|
| mg/g DW | ||||
| YE (g/L) | ||||
| 0.0 (Control) | 0.18 ± 0.01 d | 0.21 ± 0.01 d | 0.09 ± 0.01 d | 0.20 ± 0.01 d |
| 0.5 | 0.21 ± 0.02 c | 0.25 ± 0.01 c | 0.11 ± 0.01 c | 0.25 ± 0.01 c |
| 1.0 | 0.25 ± 0.01 b | 0.27 ± 0.02 b | 0.15 ± 0.01 b | 0.31 ± 0.02 b |
| 1.5 | 0.29 ± 0.02 a | 0.31 ± 0.02 a | 0.20 ± 0.01 a | 0.38 ± 0.01 a |
| ANOVA-1-way | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** |
| GA (ppm) | * | * | * | * |
| 0.0 (Control) | 0.19 ± 0.01 d | 0.22 ± 0.01 d | 0.09 ± 0.01 d | 0.20 ± 0.02 d |
| 100 | 0.22 ± 0.02 c | 0.24 ± 0.01 c | 0.12 ± 0.01 c | 0.26 ± 0.01 c |
| 200 | 0.25 ± 0.02 b | 0.27 ± 0.01 b | 0.15 ± 0.01 b | 0.32 ± 0.01 b |
| 300 | 0.28 ± 0.02 a | 0.30 ± 0.01 a | 0.18 ± 0.01 a | 0.37 ± 0.01 a |
| ANOVA-1-way | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** |
| Two-way analysis of variance | ||||
| Corr. model | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** |
| YE | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** |
| GA | <0.001 *** | <0.001 *** | <0.001 *** | <0.001 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Youssef, S.M.; Abdella, E.M.M.; Al-Elwany, O.A.; Alshallash, K.S.; Alharbi, K.; Ibrahim, M.T.S.; Tawfik, M.M.; Abu-Elsaoud, A.M.; Elkelish, A. Integrative Application of Foliar Yeast Extract and Gibberellic Acid Improves Morpho-Physiological Responses and Nutrient Uptake of Solidago virgaurea Plant in Alkaline Soil. Life 2022, 12, 1405. https://doi.org/10.3390/life12091405
Youssef SM, Abdella EMM, Al-Elwany OA, Alshallash KS, Alharbi K, Ibrahim MTS, Tawfik MM, Abu-Elsaoud AM, Elkelish A. Integrative Application of Foliar Yeast Extract and Gibberellic Acid Improves Morpho-Physiological Responses and Nutrient Uptake of Solidago virgaurea Plant in Alkaline Soil. Life. 2022; 12(9):1405. https://doi.org/10.3390/life12091405
Chicago/Turabian StyleYoussef, Samah M., Ebtsam M. M. Abdella, Omar A. Al-Elwany, Khalid S. Alshallash, Khadiga Alharbi, Mariam T. S. Ibrahim, Moataz M. Tawfik, Abdelghafar M. Abu-Elsaoud, and Amr Elkelish. 2022. "Integrative Application of Foliar Yeast Extract and Gibberellic Acid Improves Morpho-Physiological Responses and Nutrient Uptake of Solidago virgaurea Plant in Alkaline Soil" Life 12, no. 9: 1405. https://doi.org/10.3390/life12091405
APA StyleYoussef, S. M., Abdella, E. M. M., Al-Elwany, O. A., Alshallash, K. S., Alharbi, K., Ibrahim, M. T. S., Tawfik, M. M., Abu-Elsaoud, A. M., & Elkelish, A. (2022). Integrative Application of Foliar Yeast Extract and Gibberellic Acid Improves Morpho-Physiological Responses and Nutrient Uptake of Solidago virgaurea Plant in Alkaline Soil. Life, 12(9), 1405. https://doi.org/10.3390/life12091405