

Survival and Neurogenesis-Promoting Effects of the Co-Overexpression of BCLXL and BDNF Genes on Wharton’s Jelly-Derived Mesenchymal Stem Cells

 ,
,  and
and 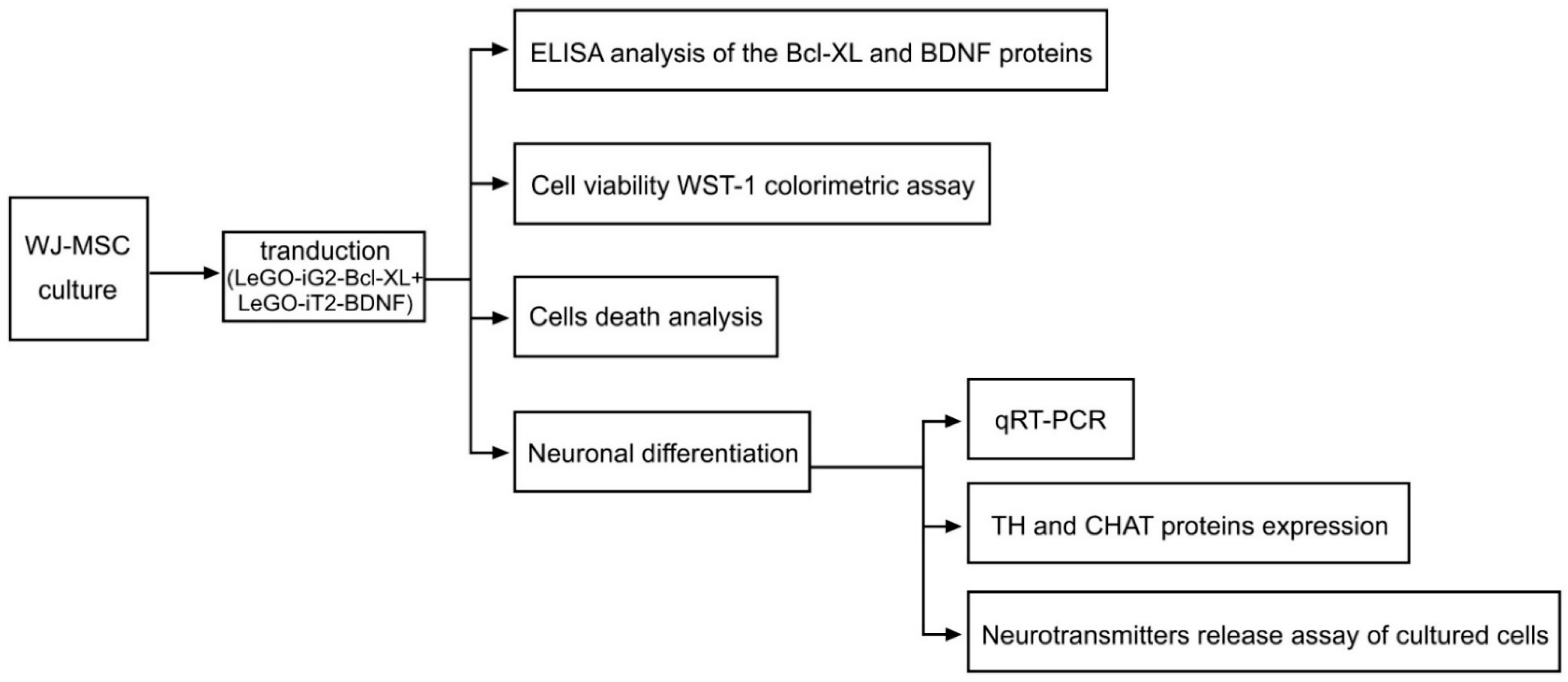{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
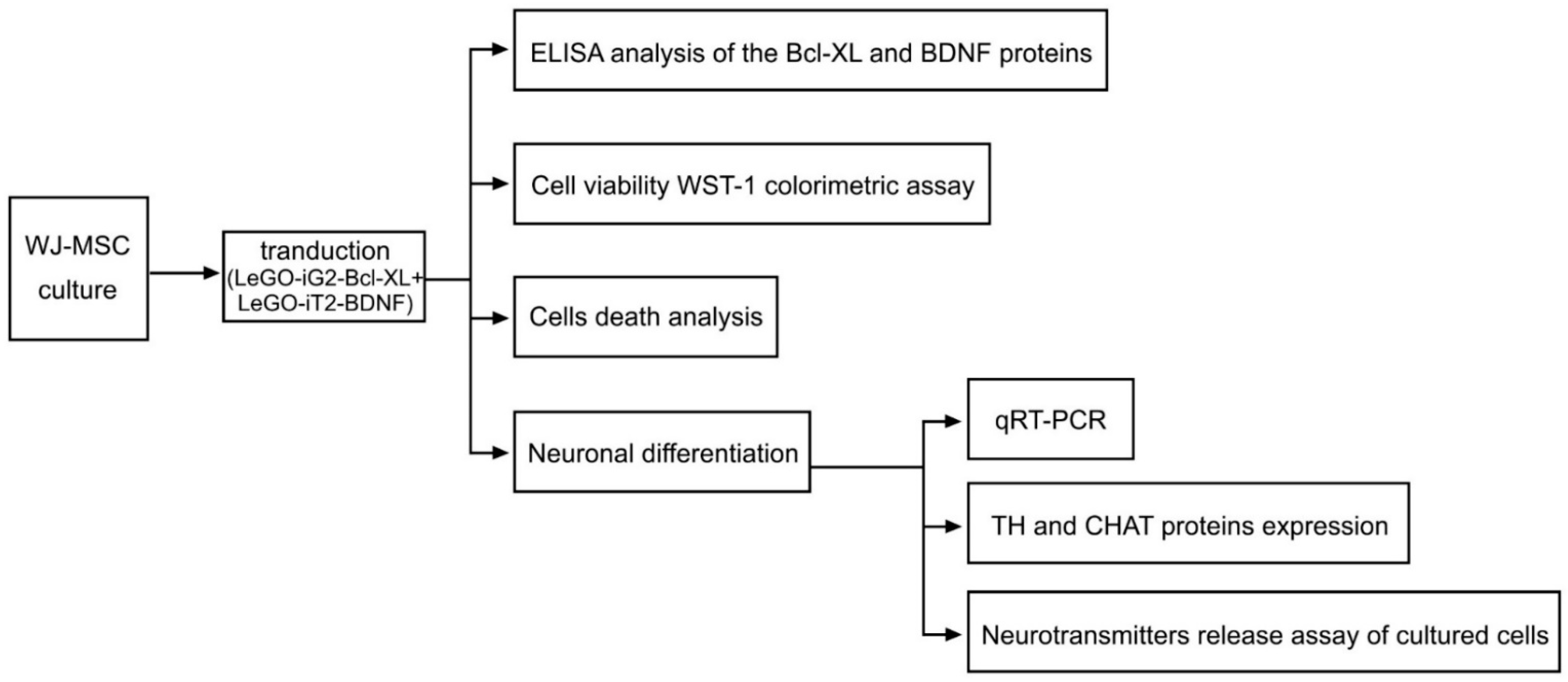
2. Materials and Methods
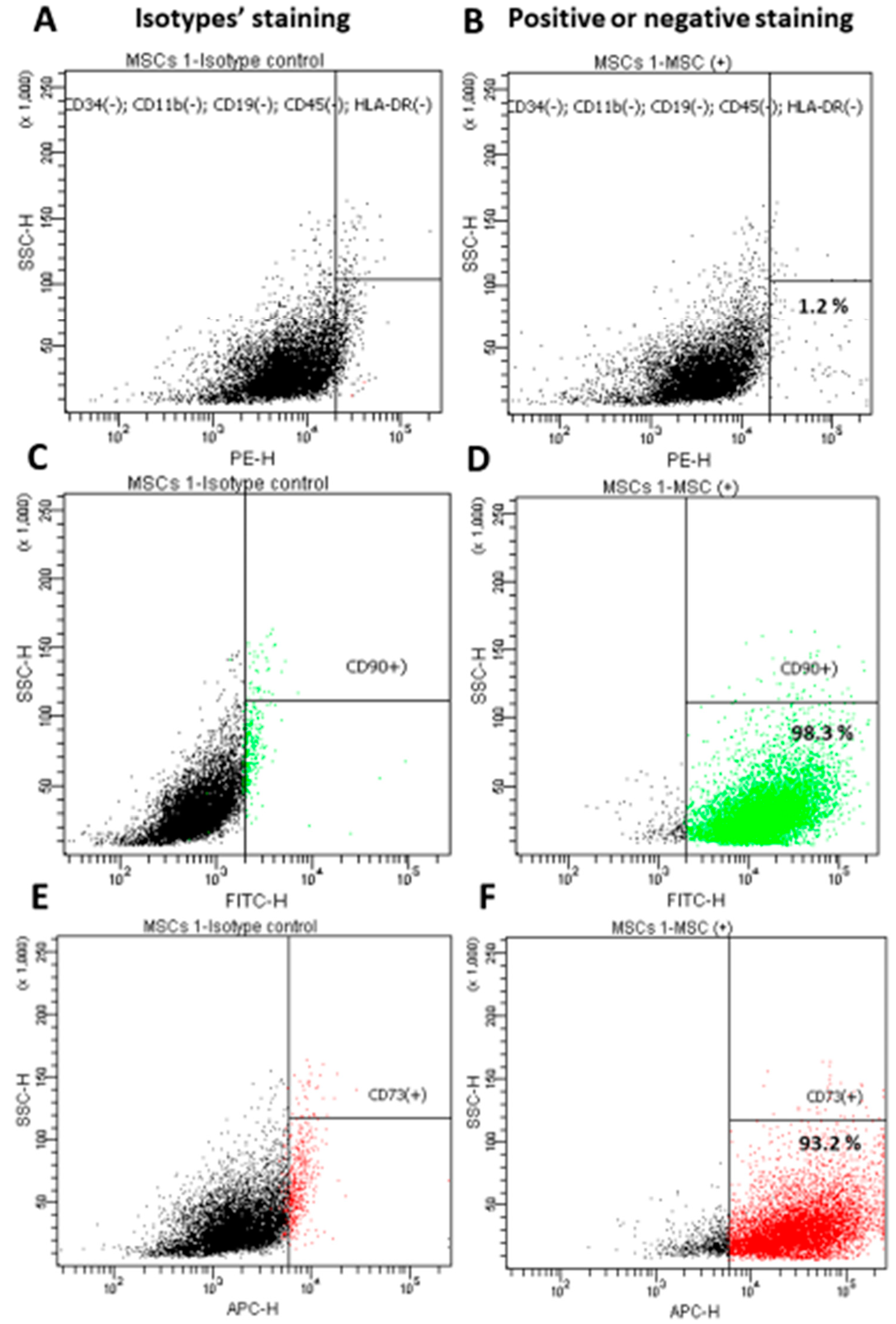
2.1. Isolating, Culturing and Characterizing the WJ-MSC
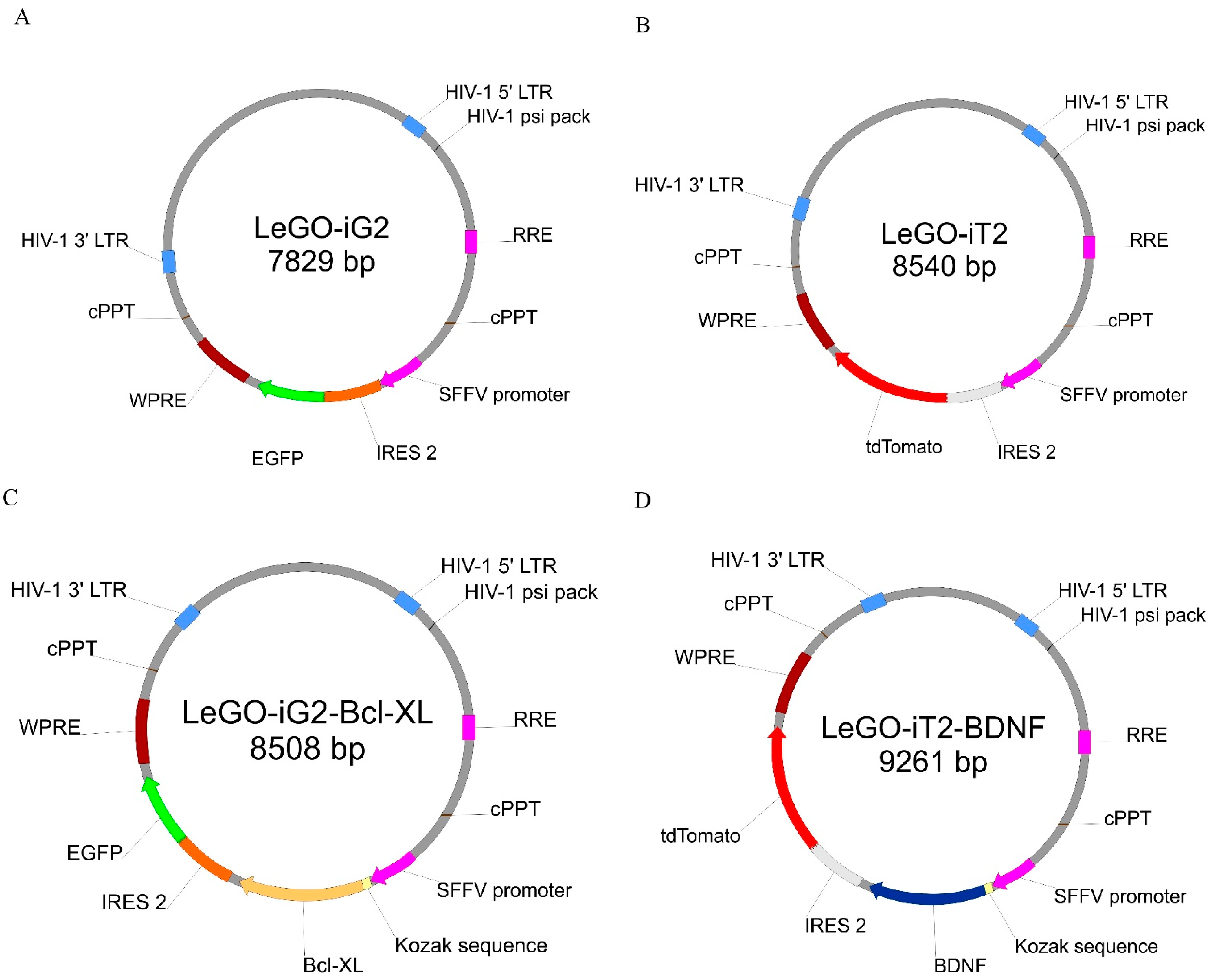
2.2. Lentiviral Vectors
2.3. WJ-MSC Transduction, Protein Extraction
2.4. ELISA Analysis of the Bcl-XL and BDNF Proteins
2.5. Cell Viability WST-1 Colorimetric Assay
2.6. Cell Death Analysis
2.7. Neuronal Differentiation
2.8. qRT-PCR
2.9. Evaluating the TH and CHAT Expression of the Proteins
2.10. Neurotransmitters Release Assay of Cultured Cells
2.11. Statistical Analysis
3. Results
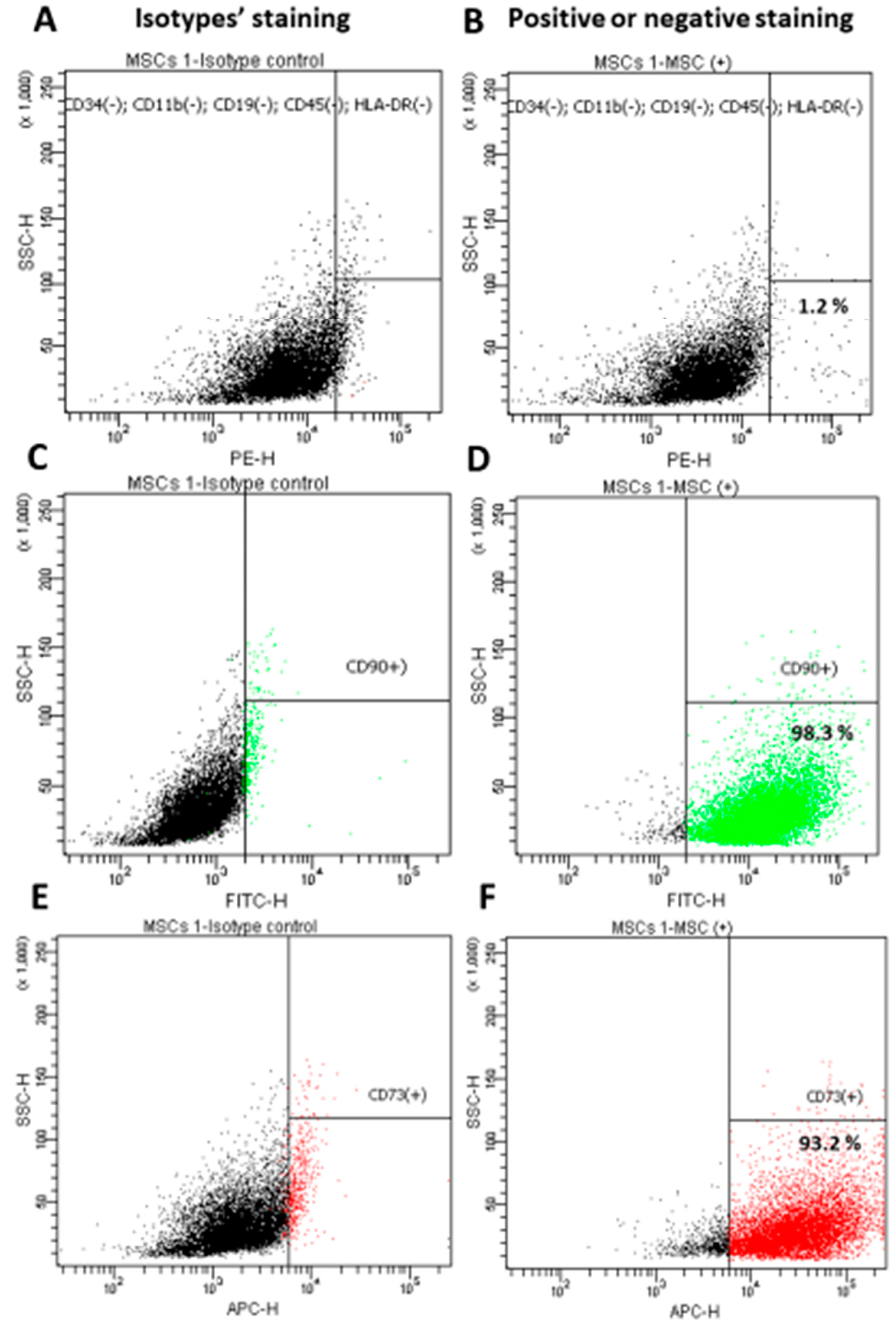
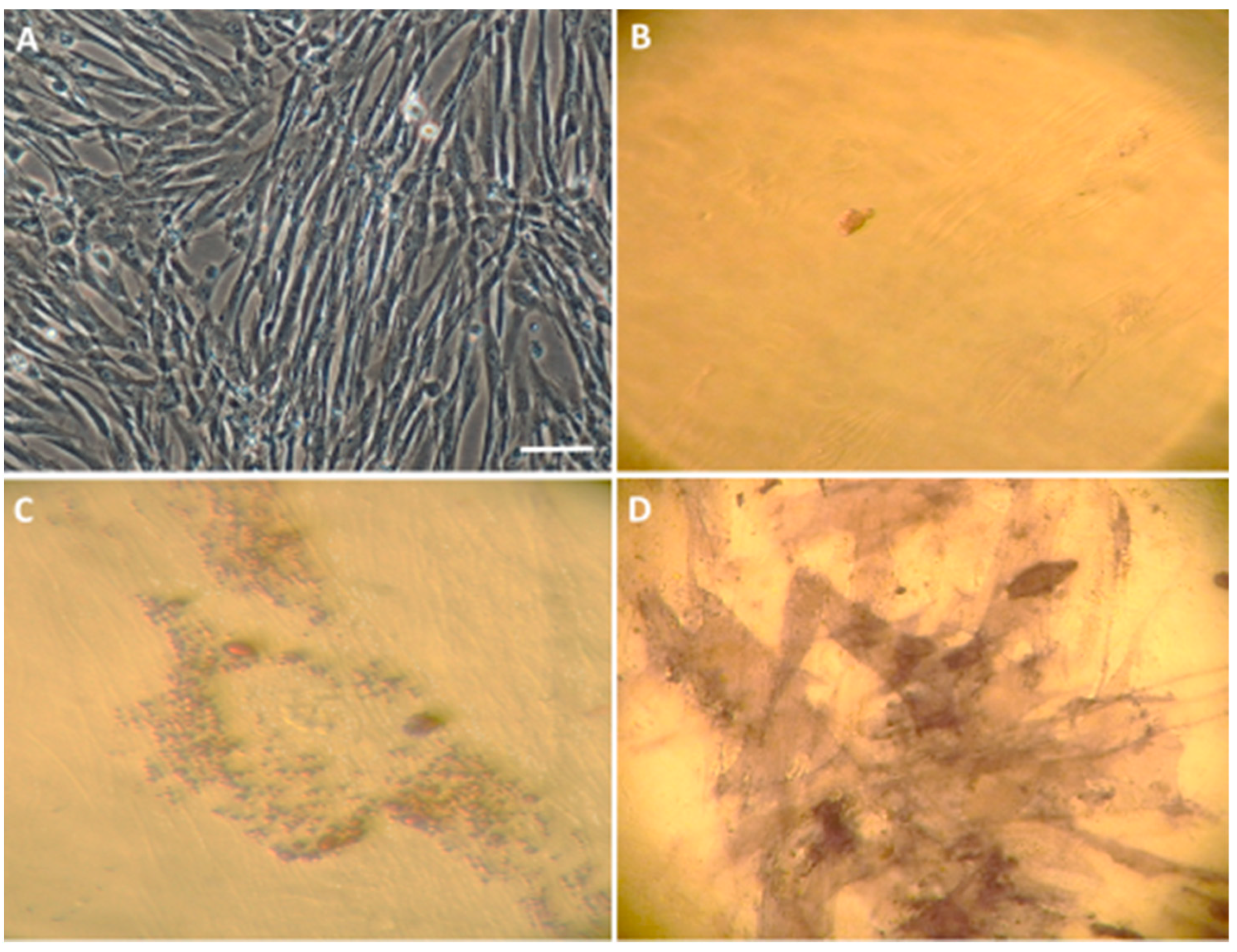
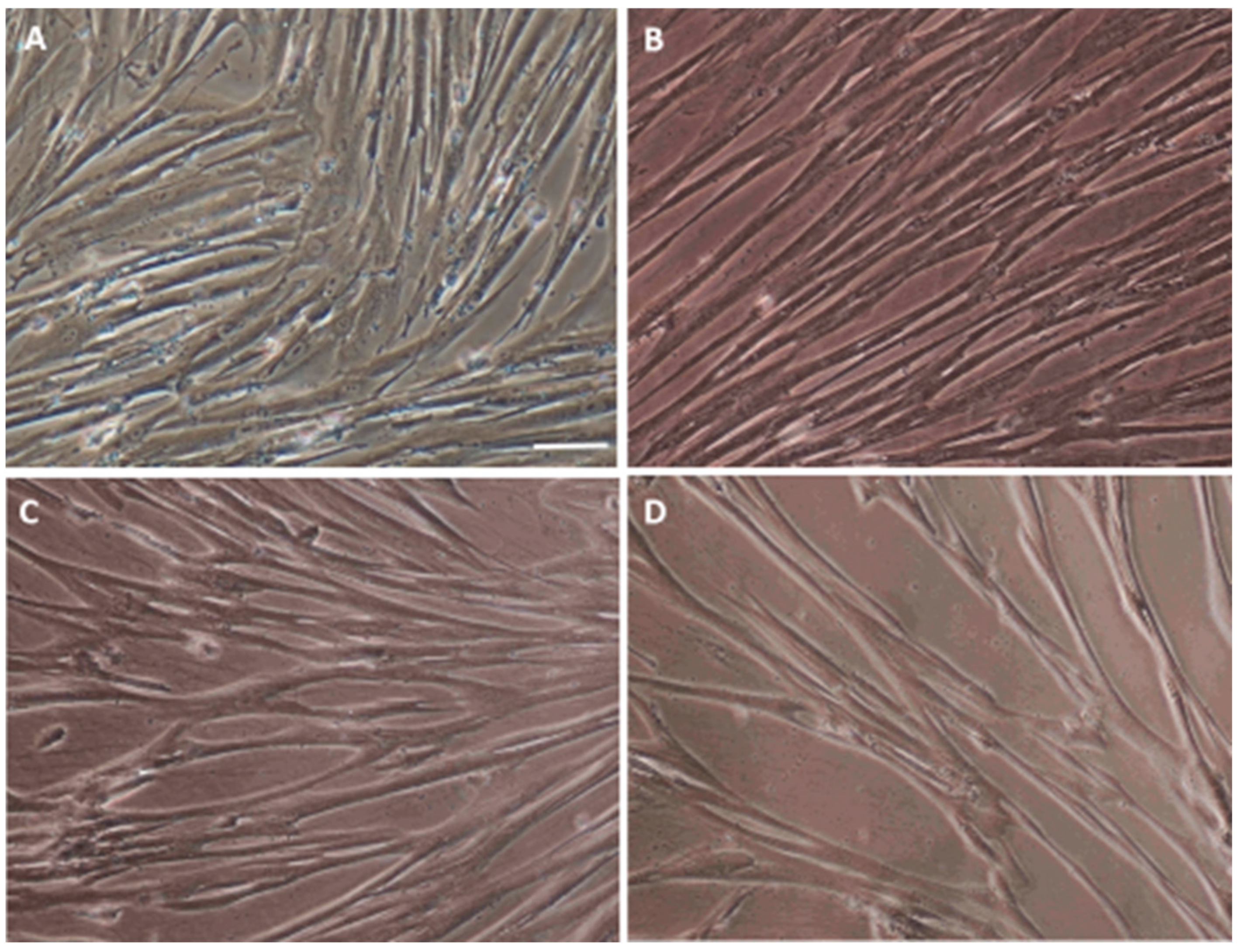
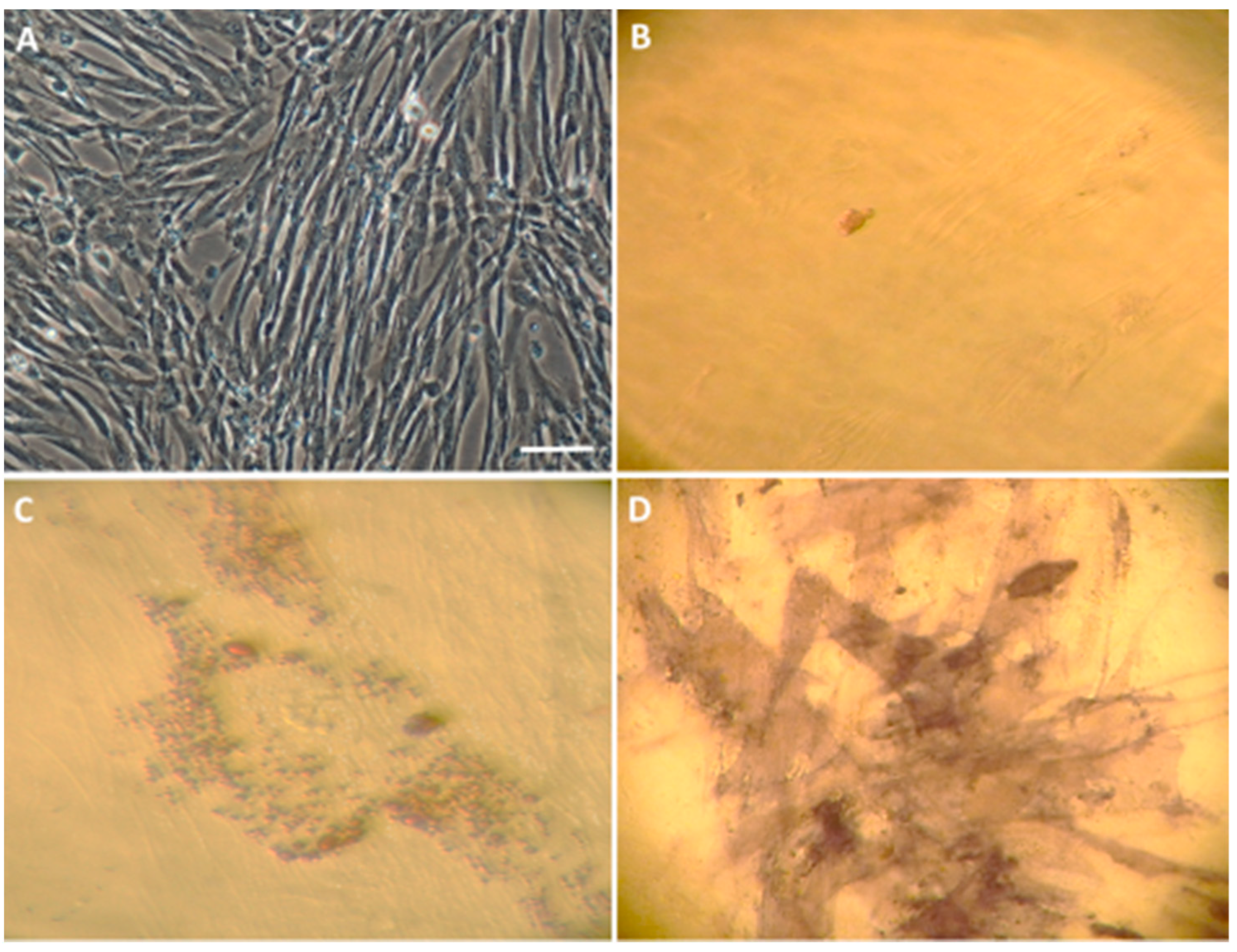
3.1. Characterization of the Isolated WJ-MSC
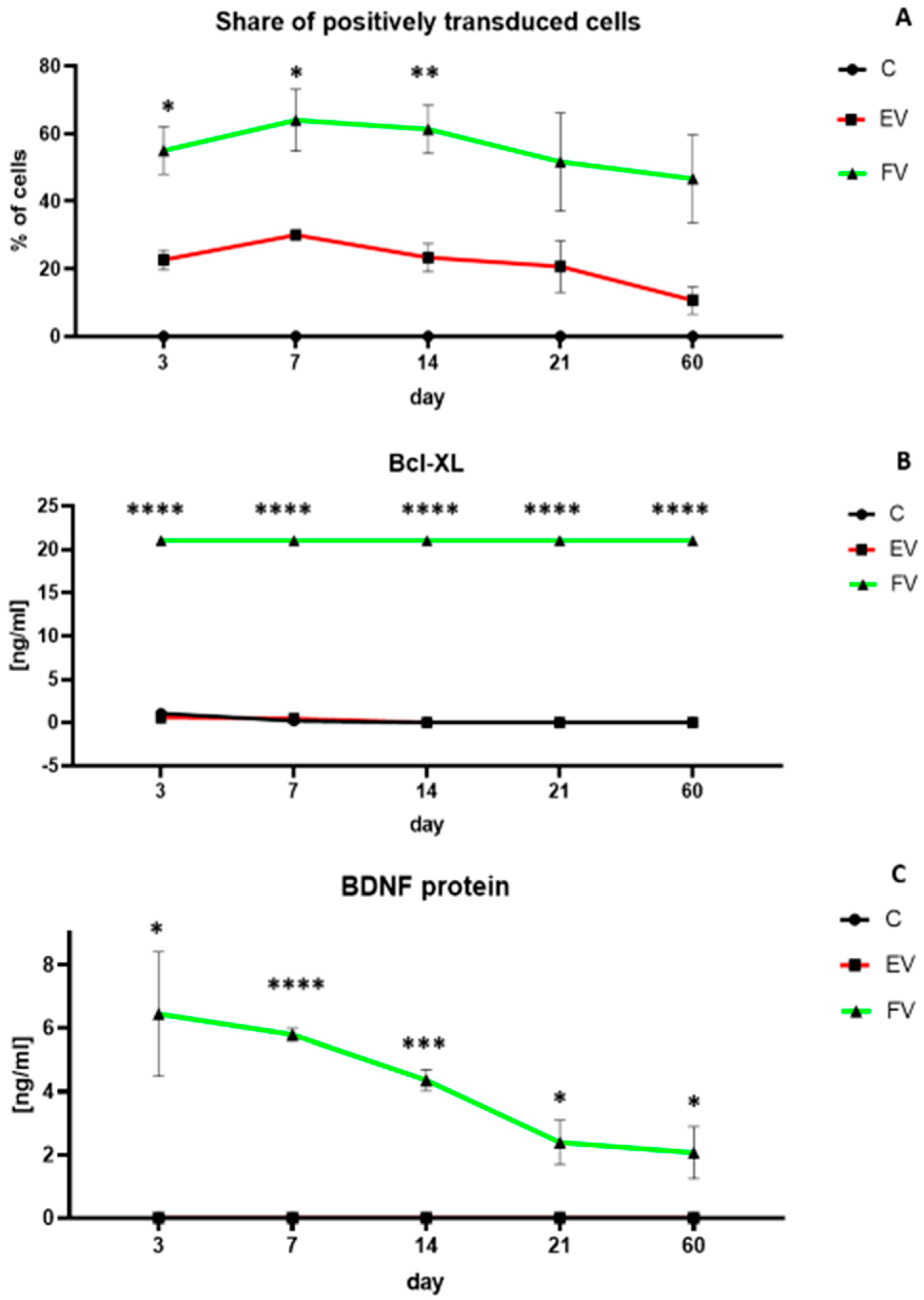
3.2. Efficiency of WJ-MSC Transduction and Overexpressed Proteins Level
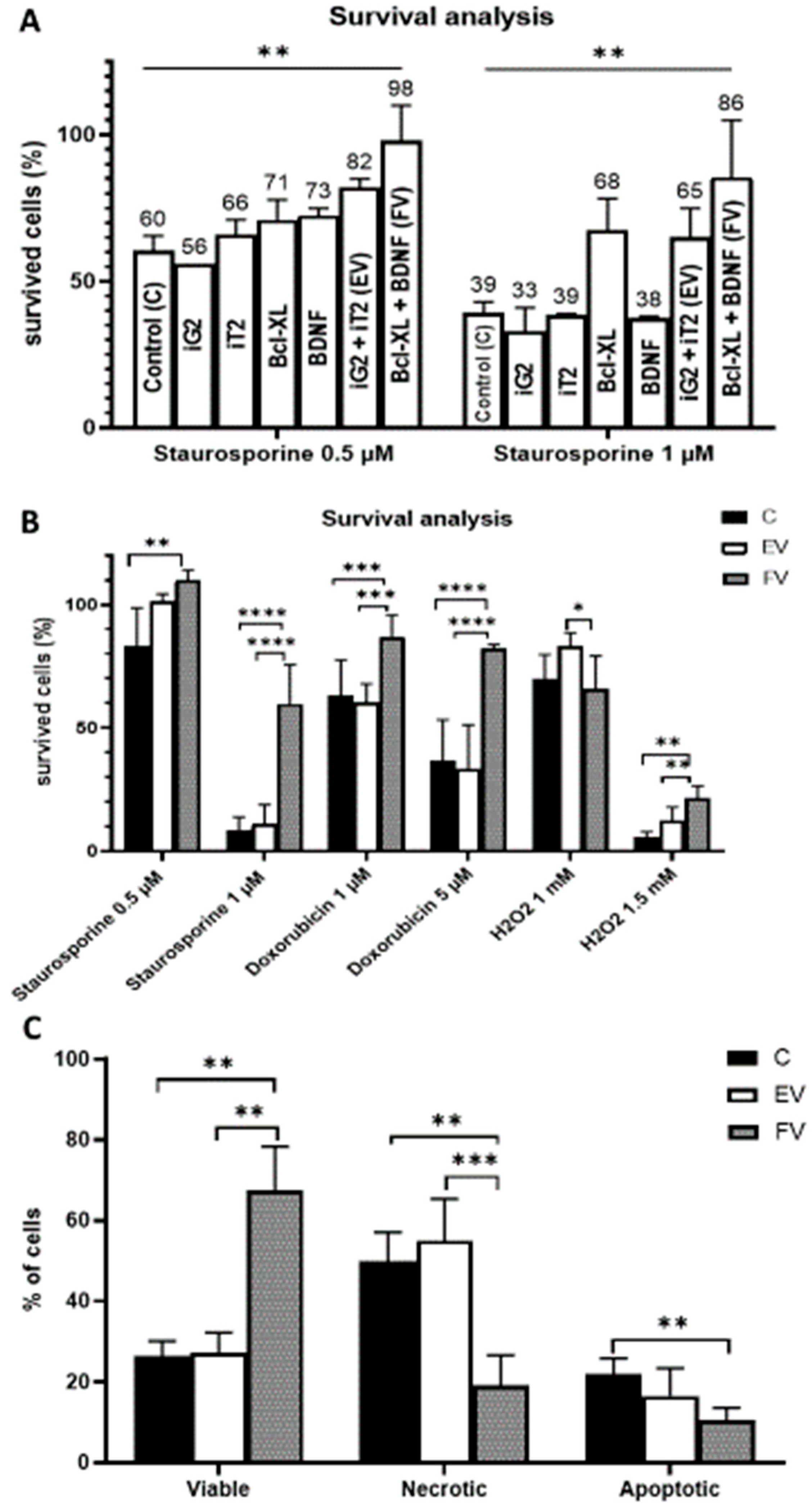
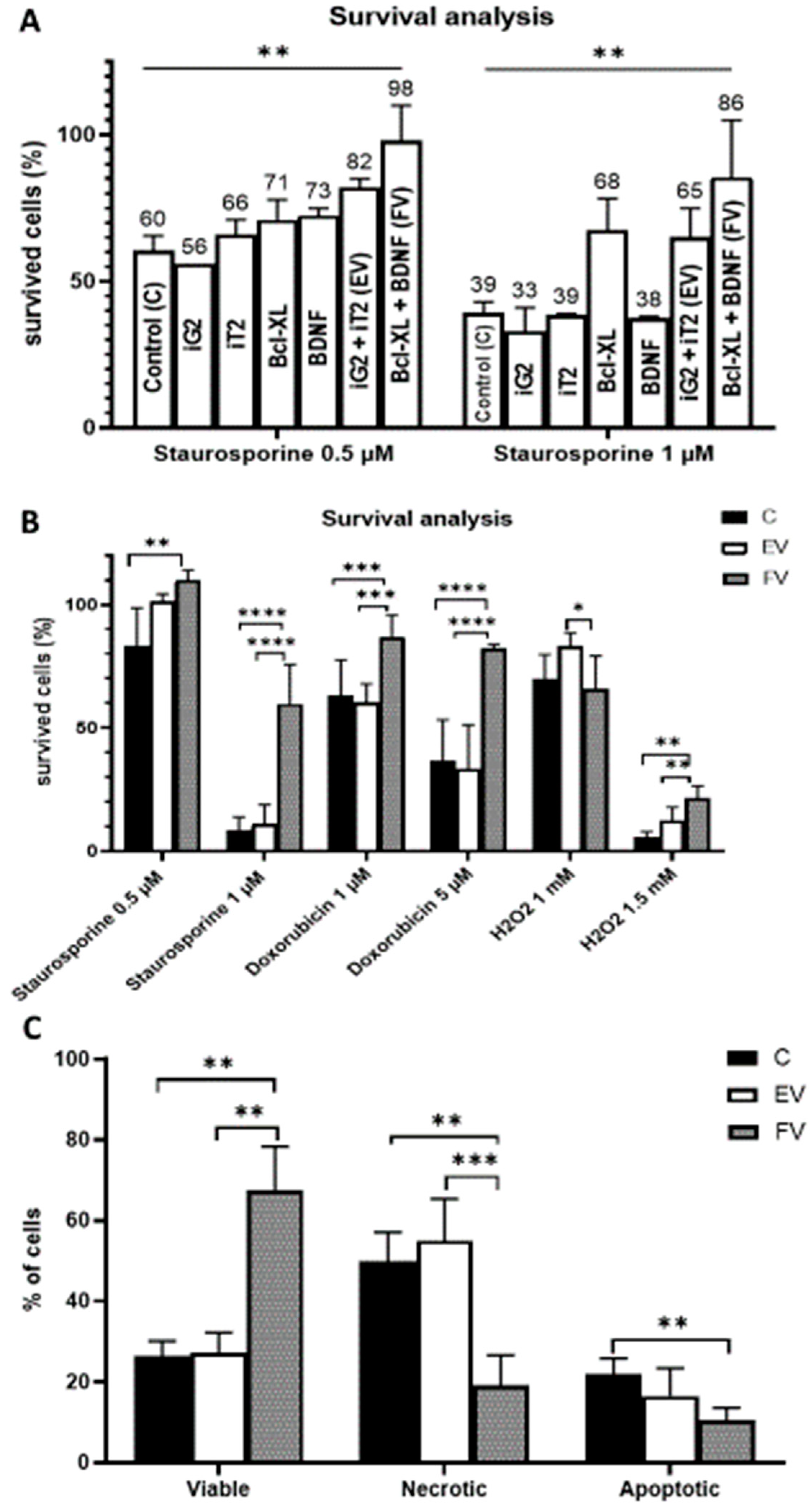
3.3. Cell Viability
3.4. Cell Death Analysis
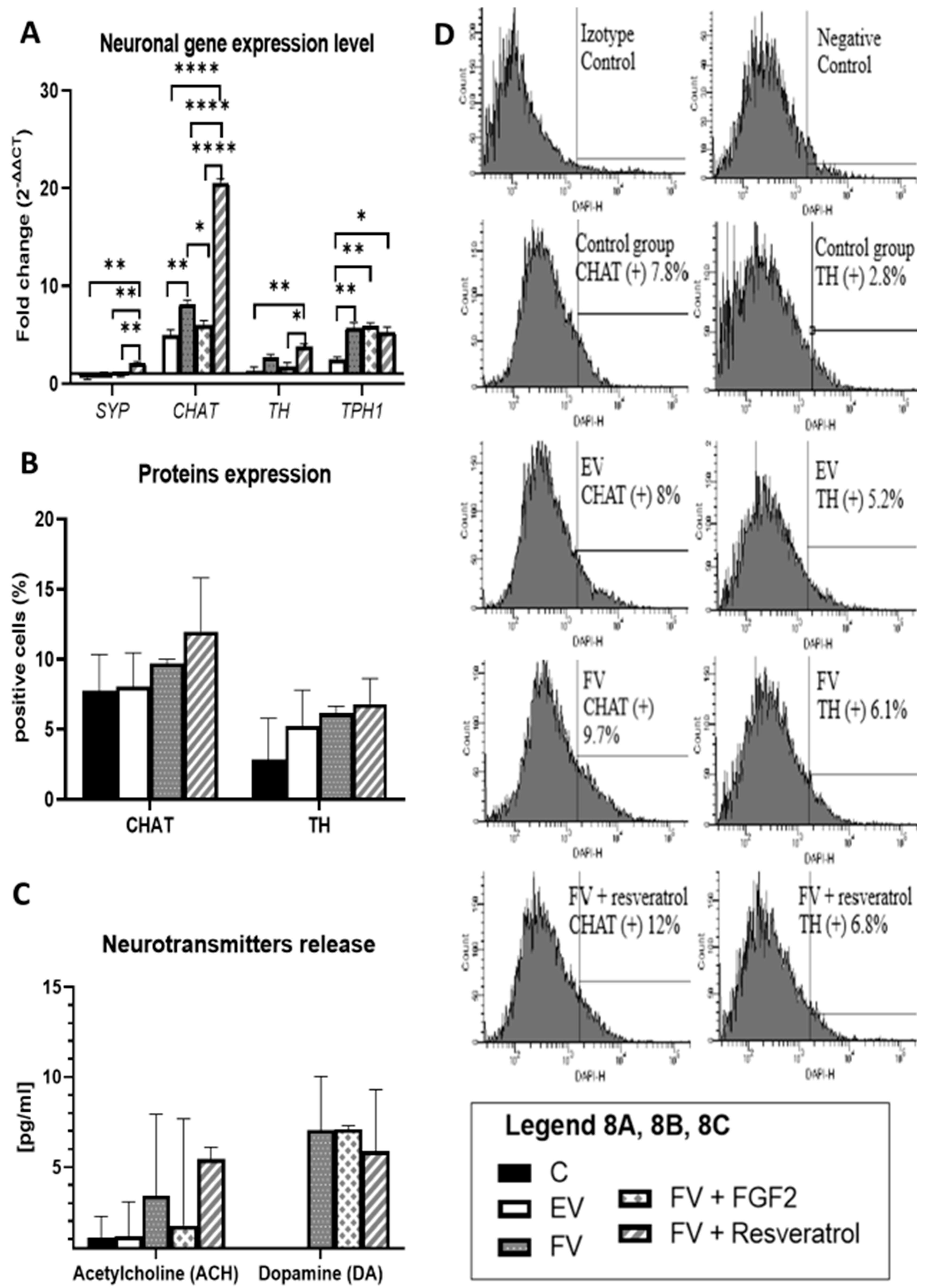
3.5. Neuronal Gene Expression Level
3.6. TH and CHAT Protein Expression
3.7. Neurotransmitter Release
4. Discussion
5. Conclusions
- The overexpressed BCLXL and BDNF genes increase the survival rate of the cells that have been transduced under toxic conditions, which increases their chance of survival after a stroke transplant procedure.
- The overexpressed BCLXL and BDNF genes guide the transduced cells into the neuronal differentiation pathway. As a result, nerve cell progenitors can be obtained. These cells can therefore be a universal material that can be used in studies of various disease models.
- The cells that are obtained as a result of the process produce neurotransmitters, which might suggest that after the cells have been transplanted into the brain, and are then subjected to the additional impact of the brain’s microenvironment, they might have a valuable impact on further therapeutic measures.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Somoza, R.; Juri, C.; Baes, M.; Wyneken, U.; Rubio, F.J. ntranigral transplantation of epigenetically induced BDNF-secreting human mesenchymal stem cells: Implications for cell-based therapies in Parkinson’s disease. Biol. Blood Marrow Transpl. 2010, 6, 1530–1540. [Google Scholar] [CrossRef] [PubMed]
- Karussis, D.; Karageorgiou, C.; Vaknin-Dembinsky, A.; Gowda-Kurkalli, B.; Gomori, J.M.; Kassis, I.; Bulte, J.W.M.; Petrou, P.; Ben-Hur, T.; Abramsky, O.; et al. Safety and immunological effects of mesenchymal stem cell transplantation in patients with multiple sclerosis and amyotrophic lateral sclerosis. Arch. Neurol. 2010, 67, 1187–1194. [Google Scholar] [CrossRef]
- Borkowska-Kuczkowska, A.; Sługocka, D.; Świątkowska-Flis, B.; Boruczkowski, D. The use of mesenchymal stem cells for the treatment of progressive retinal diseases: A review. Regen. Med. 2019, 14, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Borkowska, P.; Zielińska, A.; Paul-Samojedny, M.; Stojko, R.; Kowalski, J. Evaluation of reference genes for quantitative real-time PCR in Wharton’s Jelly-derived mesenchymal stem cells after lentiviral transduction and differentiation. Mol. Biol. Rep. 2020, 47, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.S.; Kang, K.K.; Oh, S.K.; Sung, S.E.; Kim, K.S.; Kwon, Y.S.; Yun, S. Isolation and Characterization of Feline Wharton’s Jelly-Derived Mesenchymal Stem Cells. Vet Sci. 2021, 8, 24. [Google Scholar] [CrossRef]
- Baghaei, K.; Hashemi, S.M.; Tokhanbigli, S.; Rad, A.; Assadzadeh-Aghdaei, H.; Sharifian, A.; Zali, M.R. Isolation, differentiation, and characterization of mesenchymal stem cells from human bone marrow. Gastroenterol. Hepatol. Bed Bench 2017, 10, 208–213. [Google Scholar]
- Alstrup, T.; Eijken, M.; Bohn, A.B.; Møller, B.; Damsgaard, T.E. Isolation of Adipose Tissue-Derived Stem Cells: Enzymatic Digestion in Combination with Mechanical Distortion to Increase Adipose Tissue-Derived Stem Cell Yield from Human Aspirated Fat. Curr. Protoc. Stem Cell Biol. 2019, 48, e68. [Google Scholar] [CrossRef]
- Bieback, K.; Klüter, H. Mesenchymal stromal cells from umbilical cord blood. Curr. Stem Cell Res. Ther. 2007, 2, 310–323. [Google Scholar] [CrossRef]
- Bianco, P.; Riminucci, M.; Gronthos, S.; Robey, P.G. Bone marrow stromal stem cells: Nature, biology, and potential applications. Stem Cells 2001, 19, 180–192. [Google Scholar] [CrossRef]
- Greco, S.J.; Zhou, C.; Ye, J.H.; Rameshwar, P. An interdisciplinary approach and characterization of neuronal cells transdifferentiated from human mesenchymal stem cells. Stem Cells Dev. 2007, 16, 811–826. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Araki, H.; Kato, J.; Nakamura, K.; Kawano, Y.; Kobune, M.; Sato, T.; Miyanishi, K.; Takayama, T.; Takahashi, M.; et al. Human mesenchymal stem cells xenografted directly to rat liver are differentiated into human hepatocytes without fusion. Blood 2005, 106, 756–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, S.A.; Roobrouck, V.D.; Verfaillie, C.M.; Van Gool, S.W. Immunological characteristics of human mesenchymal 10 Stem Cells International stem cells and multipotent adult progenitor cells. Immunol. Cell Biol. 2013, 91, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Trzaska, K.A.; Rameshwar, P. Dopaminergic neuronal differentiation protocol for human mesenchymal stem cells. Methods Mol. Biol. 2011, 698, 295–303. [Google Scholar] [PubMed]
- Singh, M.; Kakkar, A.; Sharma, R.; Kharbanda, O.P.; Monga, N.; Kumar, M.; Chowdhary, S.; Airan, B.; Mohanty, S. Synergistic Effect of BDNF and FGF2 in Efficient Generation of Functional Dopaminergic Neurons from human Mesenchymal Stem Cells. Sci. Rep. 2017, 4, 10378. [Google Scholar] [CrossRef]
- Nandy, S.B.; Mohanty, S.; Singh, M.; Behari, M.; Airan, B. Fibroblast Growth Factor-2 alone as an efficient inducer for differentiation of human bone marrow mesenchymal stem cells into dopaminergic neurons. J. Biomed. Sci. 2014, 24, 83. [Google Scholar] [CrossRef] [PubMed]
- Mendivil-Perez, M.; Velez-Pardo, C.; Jimenez-Del-Rio, M. Direct transdifferentiation of human Wharton’s jelly mesenchymal stromal cells into cholinergic-like neurons. J. Neurosci. Methods 2019, 15, 126–138. [Google Scholar] [CrossRef]
- Castilho, R.F.; Hansson, O.; Brundin, P. Improving the survival of grafted embryonic dopamine neurons in rodent models of Parkinson’s disease. Prog. Brain Res. 2000, 127, 203–231. [Google Scholar]
- Kirkeby, A.; Nolbrant, S.; Tiklova, K.; Heuer, A.; Kee, N.; Cardoso, T.; Ottosson, D.R.; Lelos, M.J.; Rifes, P.; Dunnett, S.B.; et al. Predictive markers guide differentiation to improve graft outcome in clinical translation of hESC-based therapy for Parkinson’s disease. Cell Stem Cell 2017, 20, 135–148. [Google Scholar] [CrossRef]
- Niclis, J.C.; Gantner, C.W.; Hunt, C.P.J.; Kauhausen, J.A.; Durnall, J.C.; Haynes, J.M.; Pouton, C.W.; Parish, C.L.; Thompson, L.H. A PITX3-EGFP reporter line reveals connectivity of dopamine and non-dopamine neuronal subtypes in grafts generated from human embryonic stem cells. Stem Cell Rep. 2017, 9, 868–882. [Google Scholar] [CrossRef]
- Björklund, A.; Parmar, M. Neuronal Replacement as a Tool for Basal Ganglia Circuitry Repair: 40 Years in Perspective. Front. Cell. Neurosci. 2020, 14, 146. [Google Scholar] [CrossRef]
- Chan, E.W.L.; Yeo, E.T.Y.; Wong, K.W.L.; See, M.L.; Wong, K.Y.; Yap, J.K.Y.; Gan, S.Y. Piper sarmentosum Roxb. Attenuates Beta Amyloid (Aβ)-Induced Neurotoxicity via the Inhibition of Amyloidogenesis and Tau Hyperphosphorylation in SH-SY5Y Cells. Curr. Alzheimer Res. 2021, 18, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Woo, M.S.; Park, J.S.; Choi, I.Y.; Kimf, W.K.; Kim, H.S. Inhibition of MMP-3 or -9 suppresses lipopolysaccharide-induced expression of proinflammatory cytokines and iNOS in microglia. J. Neurochem. 2008, 106, 770–780. [Google Scholar] [CrossRef] [PubMed]
- Dziedzic, A.; Saluk-Bijak, J.; Miller, E.; Niemcewicz, M.; Bijak, M. The Impact of SARS-CoV-2 Infection on the Development of Neurodegeneration in Multiple Sclerosis. Int. J. Mol. Sci. 2021, 22, 1804. [Google Scholar] [CrossRef] [PubMed]
- Yuste, V.J.; Sánchez-López, I.; Solé, C.; Encinas, M.; Bayascas, J.; Boix, J.; Cornella, J. The prevention of the staurosporine-induced apoptosis by Bcl-X(L), but not by Bcl-2 or caspase inhibitors, allows the extensive differentiation of human neuroblastoma cells. J. Neurochem. 2002, 80, 126–139. [Google Scholar] [CrossRef]
- Oliver, L.; Hue, E.; Rossignol, J.; Bougras, G.; Hulin, P.; Naveilhan, P.; Heymann, D.; Lescaudron, L.; Valette, F. Distinct roles of Bcl-2 and Bcl-Xl in the apoptosis of human bone marrow mesenchymal stem cells during differentiation. PLoS ONE 2011, 12, e19820. [Google Scholar] [CrossRef]
- Nibuya, M.; Morinobu, S.; Duman, R.S. Regulation of BDNF and trkB mRNA in rat brain by chronic electroconvulsive seizure and antidepressant drug treatments. J. Neurosci. 1995, 15, 7539–7547. [Google Scholar] [CrossRef]
- Lu, B.; Nagappan, G.; Guan, X.; Nathan, P.J.; Wren, P. BDNF-based synaptic repair as a disease-modifying strategy for neurodegenerative diseases. Nat. Rev. Neurosci. 2013, 14, 401–416. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Xu, J.C.; Cho, G.S.; Kwon, C.; Dawson, T.; Dawson, V. Neurons Derived from Human Induced Pluripotent Stem Cells Integrate into Rat Brain Circuits and Maintain Both Excitatory and Inhibitory Synaptic Activities. eNeuro 2019, 22, 6. [Google Scholar] [CrossRef]
- Kopach, O. Monitoring maturation of neural stem cell grafts within a host microenvironment. World J. Stem Cells 2019, 26, 982–989. [Google Scholar] [CrossRef]
- Kopach, O.; Pivneva, T. Cell-based therapies for neural replacement strategies in stroke-related neurodegeneration: Neurophysiological insights into stem progenitor cell neurogenesis within a host environment. Neural Regen. Res. 2018, 13, 1350–1351. [Google Scholar] [CrossRef]
- Sun, D.; Gugliotta, M.; Rolfe, A.; Reid, W.; McQuiston, A.R.; Hu, W.; Young, H. Sustained Survival and Maturation of Adult Neural Stem/Progenitor Cells after Transplantation into the Injured Brain. J. Neurotrauma 2011, 28, 961–972. [Google Scholar] [CrossRef] [PubMed]
- Simão, D.; Silva, M.M.; Terrasso, A.P.; Arez, F.; Sousa, M.F.Q.; Mehrjadi, N.Z.; Šarić, T. Recapitulation of Human Neural Microenvironment Signatures in iPSC-Derived NPC 3D Differentiation. Stem Cell Rep. 2018, 14, 552–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borkowska, P.; Zielinska, A.; Paul-Samojedny, M.; Stojko, R.; Kowalski, J. Synergistic Effect of the Long-Term Overexpression of Bcl-2 and BDNF Lentiviral in Cell Protecting against Death and Generating TH Positive and CHAT Positive Cells from MSC. Int. J. Mol. Sci. 2021, 22, 7086. [Google Scholar] [CrossRef] [PubMed]
- Weber, K.; Bartsch, U.; Stocking, C.; Fehse, B. A multicolor panel of novel lentiviral “gene ontology” (LeGO) vectors for functional gene analysis. Mol. Ther. 2008, 16, 698–706. [Google Scholar] [CrossRef]
- Martín, F.; Tristán-Manzano, M.; Maldonado-Pérez, N.; Sánchez-Hernández, S.; Benabdellah, K.; Cobo, M. Stable Genetic Modification of Mesenchymal Stromal Cells Using Lentiviral Vectors. Methods Mol. Biol. 2019, 1937, 267–280. [Google Scholar]
- Mendes-Pinheiro, B.; Anjo, S.I.; Manadas, B.; Da Silva, J.D.; Marote, A.; Behie, L.A.; Teixeira, F.G.; Salgado, A.J. Bone Marrow Mesenchymal Stem Cells’ Secretome Exerts Neuroprotective Effects in a Parkinson’s Disease Rat Model. Front. Bioeng. Biotechnol. 2019, 1, 294. [Google Scholar] [CrossRef]
- Nitzsche, F.; Müller, C.; Lukomska, B.; Jolkkonen, J.; Deten, A.; Boltze, J. MSC Adhesion Cascade-Insights into Homing and Transendothelial Migration. Stem Cells 2017, 35, 1446–1460. [Google Scholar] [CrossRef]
- Wennersten, A.; Meier, X.; Holmin, S.; Wahlberg, L.; Matheisen, T. Proliferation, migration, and differentiation of human neural stem/progenitor cells after transplantation into a rat model of traumatic brain injury. J. Neurosurg. 2004, 100, 88–96. [Google Scholar] [CrossRef]
- Einstein, O.; Karussis, D.; Grigoriadis, N.; Mizchari-Kol, R.; Reinhartz, E.; Abramsky, O.; Ben-Hur, T. Intraventricular transplantation of neural precursor cell spheres attenuates acute experimental allergic encephalomyelitis. Mol. Cell Neurosci. 2003, 24, 1074–1082. [Google Scholar] [CrossRef]
- Shahror, R.A.; Linares, G.R.; Wang, Y.; Hsueh, S.C.; Wu, J.; Chuang, D.M.; Chiang, Y.H.; Chen, K.Y. Transplantation of Mesenchymal Stem Cells Overexpressing Fibroblast Growth Factor 21 Facilitates Cognitive Recovery and Enhances Neurogenesis in a Mouse Model of Traumatic Brain Injury. J. Neurotrauma 2020, 37, 14–26. [Google Scholar] [CrossRef]
- Scheper, V.; Schwieger, J.; Hamm, A.; Lenarz, T. BDNF-overexpressing human mesenchymal stem cells mediate increased neuronal protection in vitro. J. Neurosci. Res. 2019, 97, 1414–1429. [Google Scholar] [CrossRef] [PubMed]
- Lejkowska, R.; Kawa, M.P.; Pius-Sadowska, E.; Rogińska, D.; Łuczkowska, K.; Machaliński, B.; Machalińska, A. Preclinical Evaluation of Long-Term Neuroprotective Effects of BDNF-Engineered Mesenchymal Stromal Cells as Intravitreal Therapy for Chronic Retinal Degeneration in Rd6 Mutant Mice. Int. J. Mol. Sci. 2019, 20, 777. [Google Scholar] [CrossRef] [PubMed]
- Fiebig, A.A.; Zhu, W.; Hollerbach, C.; Leber, B.; Andrews, D.W. Bcl-XL is qualitatively different from and ten times more effective than Bcl-2 when expressed in a breast cancer cell line. BMC Cancer 2006, 23, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, J.B.; Li, X.; Zhong, S.M.; Liu, J.D.; Chen, C.B.; Wu, X.Y. Knockdown of long noncoding antisense RNA brain-derived neurotrophic factor attenuates hypoxia/reoxygenation-induced nerve cell apoptosis through the BDNF-TrkB-PI3K/Akt signaling pathway. Neuroreport 2017, 28, 910–916. [Google Scholar] [CrossRef]
- Chen, A.; Xiong, L.J.; Tong, Y.; Mao, M. The neuroprotective roles of BDNF in hypoxic ischemic brain injury. Biomed. Rep. 2013, 1, 167–176. [Google Scholar] [CrossRef]
- Linares, G.R.; Leng, Y.; Maric, D.; Chuang, D.M. Overexpression of fibroblast growth factor-21 (FGF-21) protects mesenchymal stem cells against caspase-dependent apoptosis induced by oxidative stress and inflammation. Cell Biol. Int. 2020, 44, 2163–2169. [Google Scholar] [CrossRef]
- Simenc, J.; Lipnik-Stangelj, M. Staurosporine induces different cell death forms in cultured rat astrocytes. Radiol. Oncol. 2012, 46, 312–320. [Google Scholar] [CrossRef]
- Belmokhtar, C.A.; Hillion, J.; Ségal-Bendirdjian, E. Staurosporine induces apoptosis through both caspase-dependent and caspase-independent mechanisms. Oncogene 2001, 20, 3354–3362. [Google Scholar] [CrossRef]
- Dunai, Z.A.; Imre, G.; Barna, G.; Korcsmáros, T.; Peták, I.; Bauer, P.; Mihalik, R. Staurosporine induces necroptotic cell death under caspase-compromised conditions in U937 cells. PLoS ONE 2012, 7, e41945. [Google Scholar] [CrossRef]
- Christidi, E.; Brunham, L.R. Regulated cell death pathways in doxorubicin-induced cardiotoxicity. Cell Death Dis. 2021, 12, 339. [Google Scholar] [CrossRef]
- Reichert, C.O.; de Freitas, F.A.; Sampaio-Silva, J.; Rokita-Rosa, L.; De Lima Barros, P.; Levy, D.; Bydlowski, S.P. Ferroptosis Mechanisms Involved in Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 8765. [Google Scholar] [CrossRef] [PubMed]
- Uhl, L.; Gerstel, A.; Chabalier, M.; Dukan, S. Hydrogen peroxide induced cell death: One or two modes of action? Heliyon 2015, 1, e00049. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Guo, C.; Kong, J. Oxidative stress in neurodegenerative diseases. Neural Regen. Res. 2012, 15, 376–385. [Google Scholar]
- Koh, J.Y.; Wie, M.B.; Gwag, B.J.; Sensi, S.; Canzoniero, L.; Demaro, J.; Csernansky, C.; Choi, D. Staurosporine-induced neuronal apoptosis. Exp. Neurol. 1995, 135, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Wiesner, D.A.; Dawson, G. Staurosporine induces programmed cell death in embryonic neurons and activation of the ceramide pathway. J. Neurochem. 1996, 66, 1418–1425. [Google Scholar] [CrossRef] [PubMed]
- MacManus, J.P.; Rasquinha, I.; Black, M.A.; Laferrière, N.; Monette, R.; Walker, T.; Morley, P. Glutamate-treated rat cortical neuronal cultures die in a way different from the classical apoptosis induced by staurosporine. Exp. Cell Res. 1997, 233, 310–320. [Google Scholar] [CrossRef]
- Gaidin, S.G.; Turovskaya, M.V.; Gavrish, M.S.; Babaev, A.A.; Mal’tseva, V.N.; Blinova, E.V.; Turovsky, E.A. The selective BDNF overexpression in neurons protects neuroglial networks against OGD and glutamate-induced excitotoxicity. Int. J. Neurosci. 2020, 130, 363–383. [Google Scholar] [CrossRef]
- Mitroshina, E.V.; Mishchenko, T.A.; Usenko, A.V.; Epifanova, E.A.; Yarkov, R.S. AAV-Syn-BDNF-EGFP Virus Construct Exerts Neuroprotective Action on the Hippocampal Neural Network during Hypoxia In Vitro. Int. J. Mol. Sci. 2018, 19, 2295. [Google Scholar] [CrossRef]
- Turovskaya, M.V.; Gaidin, S.G.; Vedunova, M.V.; Babaev, A.A.; Turovsky, E.A. BDNF Overexpression Enhances the Preconditioning Effect of Brief Episodes of Hypoxia, Promoting Survival of GABAergic Neurons. Neurosci. Bull. 2020, 36, 733–760. [Google Scholar] [CrossRef]
- Godin, J.D.; Poizat, G.; Hickey, M.A.; Maschat, F. Mutant huntingtin-impaired degradation of β-catenin causes neurotoxicity in Huntington’s disease. EMBO J. 2010, 29, 2433–2445. [Google Scholar] [CrossRef] [PubMed]
- Browne, S.E.; Beal, M.F. Oxidative Damage in Huntington’s Disease Pathogenesis. Antioxid. Redox Signal. 2006, 8, 11–12. [Google Scholar] [CrossRef]
- Salama, M.; Arias-Carrión, O. Natural toxins implicated in the development of Parkinson’s disease. Ther. Adv. Neurol. Disord. 2011, 4, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.B.; McDowell, K.A.; Siebert, A.A. Environmental neurotoxin-induced progressive model of parkinsonism in rats. Ann. Neurol. 2010, 68, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.W.; Koh, H.C.; Chang, M.Y.; Roh, E.; Choi, C.Y.; Oh, Y.; Son, H.; Lee, Y.S.; Studer, L.; Lee, S.H. Enhanced in vitro midbrain dopamine neuron differentiation, dopaminergic function, neurite outgrowth, and 1-methyl-4-phenylpyridium resistance in mouse embryonic stem cells overexpressing Bcl-XL. J. Neurosci. 2004, 24, 843–852. [Google Scholar] [CrossRef] [Green Version]
- Liste, I.; Garcia-Garcia, E.; Martinez-Serrano, A. The generation of dopaminergic neurons by human neural stem cells is enhanced by Bcl-XL, both in vitro and in vivo. J. Neurosci. 2004, 24, 10786–10795. [Google Scholar] [CrossRef]
- Liste, I.; García-García, E.; Bueno, C.; Martínez-Serrano, A. Bcl-XL modulates the differentiation of immortalized human neural stem cells. Cell Death Differ. 2007, 14, 1880–1892. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.Y.; Saarma, M.; Arumäe, U. Death receptors and caspases but not mitochondria are activated in the GDNF- or BDNF-deprived dopaminergic neurons. J. Neurosci. 2008, 23, 7467–7475. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Wang, X.; Liu, L.; Chen, H.; Wang, N. Ginkgolide-Platinum(II) Complex GPt(II) Exhibits Therapeutic Effect on Depression in Mice via Upregulation of DA and 5-HT Neurotransmitters. Med. Sci. Monit. 2020, 2, e922052. [Google Scholar] [CrossRef] [PubMed]
- Corpuz, H.M.; Arimura, M.; Chawalitpong, S.; Miyazaki, K.; Sawaguchi, M.; Nakamura, S.; Katayama, S. Oral Administration of Okara Soybean By-Product Attenuates Cognitive Impairment in a Mouse Model of Accelerated Aging. Nutrients 2019, 11, 2939. [Google Scholar] [CrossRef]
- Piotrowicz, Z.; Chalimoniuk, M.; Płoszczyca, K.; Czuba, M.; Langfort, J. Exercise-Induced Elevated BDNF Level Does Not Prevent Cognitive Impairment Due to Acute Exposure to Moderate Hypoxia in Well-Trained Athletes. Int. J. Mol. Sci. 2020, 21, 5569. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borkowska, P.; Morys, J.; Zielinska, A.; Sadlocha, M.; Kowalski, J. Survival and Neurogenesis-Promoting Effects of the Co-Overexpression of BCLXL and BDNF Genes on Wharton’s Jelly-Derived Mesenchymal Stem Cells. Life 2022, 12, 1406. https://doi.org/10.3390/life12091406
Borkowska P, Morys J, Zielinska A, Sadlocha M, Kowalski J. Survival and Neurogenesis-Promoting Effects of the Co-Overexpression of BCLXL and BDNF Genes on Wharton’s Jelly-Derived Mesenchymal Stem Cells. Life. 2022; 12(9):1406. https://doi.org/10.3390/life12091406
Chicago/Turabian StyleBorkowska, Paulina, Julia Morys, Aleksandra Zielinska, Marcin Sadlocha, and Jan Kowalski. 2022. "Survival and Neurogenesis-Promoting Effects of the Co-Overexpression of BCLXL and BDNF Genes on Wharton’s Jelly-Derived Mesenchymal Stem Cells" Life 12, no. 9: 1406. https://doi.org/10.3390/life12091406
APA StyleBorkowska, P., Morys, J., Zielinska, A., Sadlocha, M., & Kowalski, J. (2022). Survival and Neurogenesis-Promoting Effects of the Co-Overexpression of BCLXL and BDNF Genes on Wharton’s Jelly-Derived Mesenchymal Stem Cells. Life, 12(9), 1406. https://doi.org/10.3390/life12091406