Colloidal Filterable Bacteria Enhance Ammonia Nitrogen Enrichment in River Colloids under Different Turbidity Conditions: Bacterial Diversity, Assembly Mechanism, and Nitrogen Transformation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Bulk Water and Colloids
2.2. Physicochemical Characterization of Bulk Water and Colloids
2.3. RNA Extraction and Pyrosequencing
2.4. Statistical Analysis
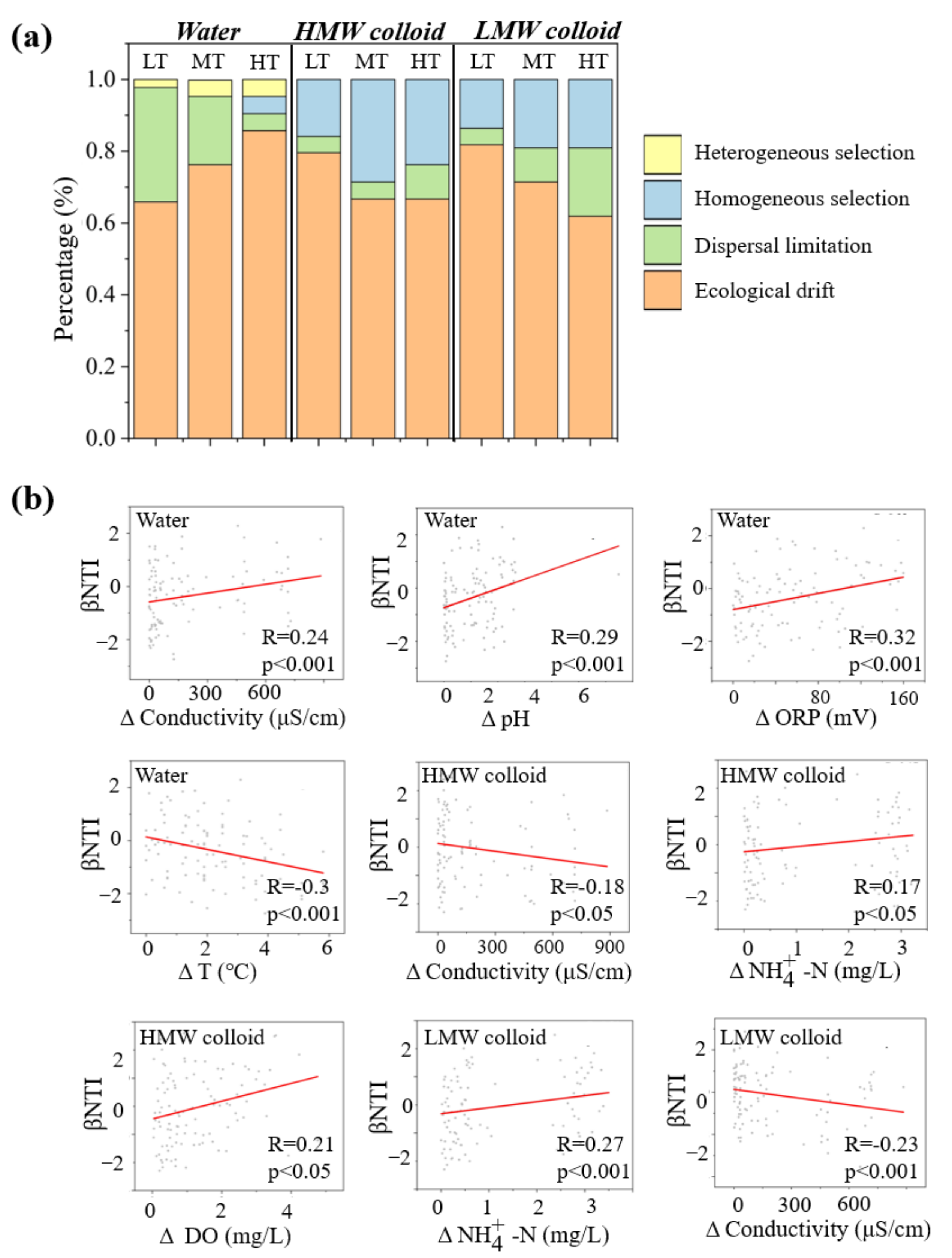
2.5. Quantifying the Community Assembly Process
3. Results
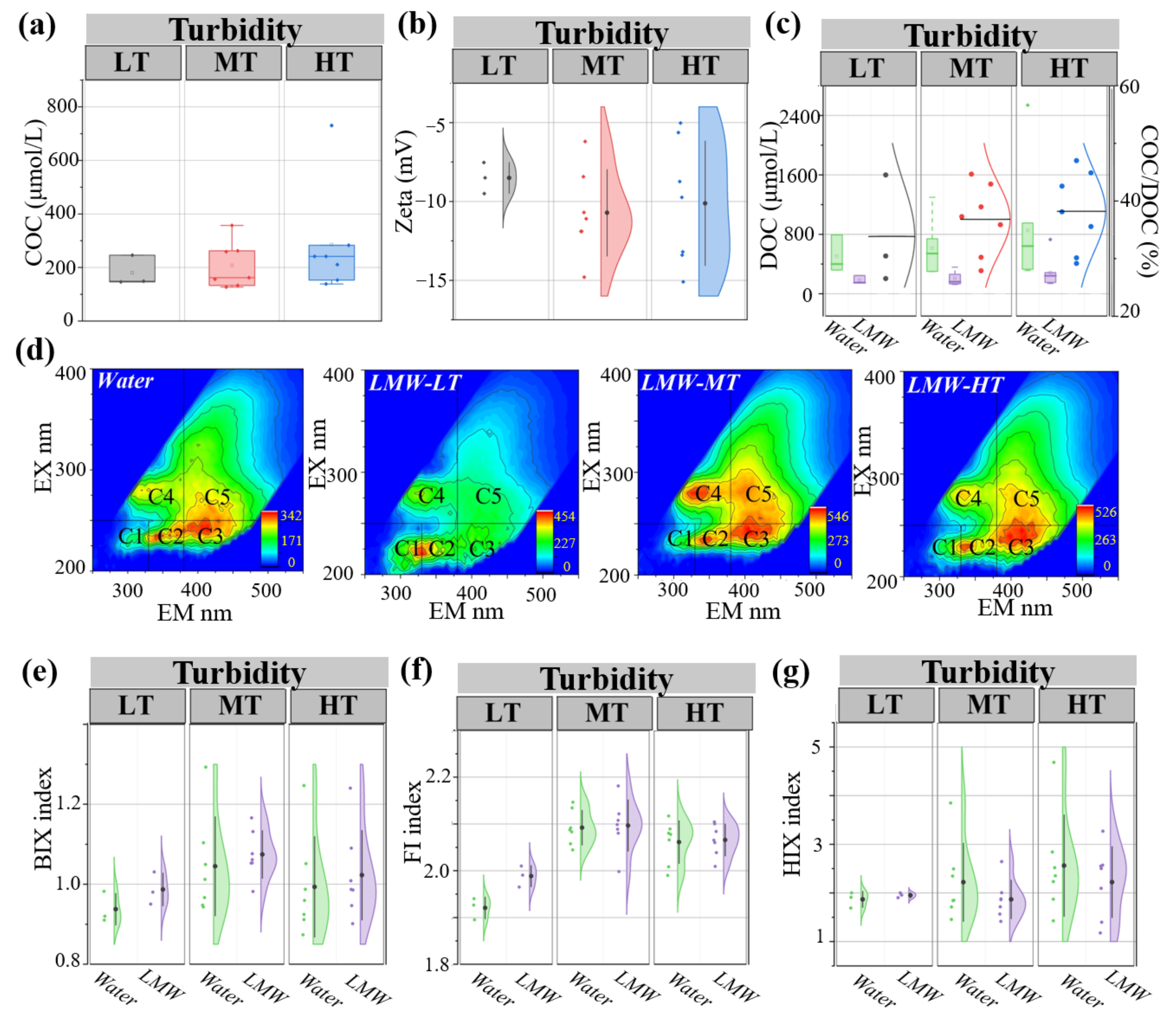
3.1. Occurrence Characteristics of LMW Colloids under Different Turbidity Conditions
3.2. Composition and Function Characteristics of Active Bacterial Communities Colonized in Colloids
3.3. Characteristics of Nitrogen Transformation Function
4. Discussion
4.1. Occurrence Characteristics of LMW Colloids under Different Turbidity Conditions
4.2. Characteristics of Filterable Bacteria in Colloids
4.3. Enrichment of Ammonia Nitrogen in Colloids
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, L.Q.; Xu, Y.J.; Li, S.Y. Riverine dissolved organic matter (DOM) as affected by urbanization gradient. Environ. Res. 2022, 212, 9. [Google Scholar] [CrossRef] [PubMed]
- Gobler, C.J. Climate Change and Harmful Algal Blooms: Insights and perspective. Harmful Algae 2020, 91, 4. [Google Scholar] [CrossRef] [PubMed]
- Karathanasis, A.D.; Johnson, D.M.C. Stability and transportability of biosolid colloids through undisturbed soil monoliths. Geoderma 2006, 130, 334–345. [Google Scholar] [CrossRef]
- Sun, W.L.; Zhou, K. Adsorption of three selected endocrine disrupting chemicals by aquatic colloids and sediments in single and binary systems. J. Soils Sediments 2015, 15, 456–466. [Google Scholar] [CrossRef]
- Pham, M.K.; Garnier, J.M. Distribution of trace elements associated with dissolved compounds (<0.45 μm–1 nm) in freshwater using coupled (frontal cascade) ultrafiltration and chromatographic separations. Environ. Sci. Technol. 1998, 32, 440–449. [Google Scholar] [CrossRef]
- Xu, H.C.; Ji, L.; Kong, M.; Jiang, H.L.; Chen, J. Molecular weight-dependent adsorption fractionation of natural organic matter on ferrihydrite colloids in aquatic environment. Chem. Eng. J. 2019, 363, 356–364. [Google Scholar] [CrossRef]
- Gebka, K.; Beldowska, M.; Saniewska, D.; Korejwo, E.; Saniewski, M. Meteorological phenomenon as a key factor controlling variability of labile particulate mercury in rivers and its inflow into coastal zone of the sea. Environ. Res. 2020, 184, 13. [Google Scholar] [CrossRef]
- Zhang, W.L.; Shi, M.; Wang, L.Q.; Li, Y.; Wang, H.L.; Niu, L.H.; Zhang, H.J.; Wang, L.F. New insights into nitrogen removal potential in urban river by revealing the importance of microbial community succession on suspended particulate matter. Environ. Res. 2022, 204, 13. [Google Scholar] [CrossRef]
- Tang, X.Q.; Li, R.; Han, D.; Scholz, M. Response of Eutrophication Development to Variations in Nutrients and Hydrological Regime: A Case Study in the Changjiang River (Yangtze) Basin. Water 2020, 12, 19. [Google Scholar] [CrossRef]
- Zhang, W.L.; Zhou, P.C.; Pan, S.Y.; Li, Y.; Lin, L.; Niu, L.H.; Wang, L.F.; Zhang, H.J. The role of microbial communities on primary producers in aquatic ecosystems: Implications in turbidity stress resistance. Environ. Res. 2022, 215, 9. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.C.; Xu, M.W.; Li, Y.N.; Liu, X.; Guo, L.D.; Jiang, H.L. Characterization, origin and aggregation behavior of colloids in eutrophic shallow lake. Water Res. 2018, 142, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Gudmundsson, L.; Boulange, J.; Do, H.X.; Gosling, S.N.; Grillakis, M.G.; Koutroulis, A.G.; Leonard, M.; Liu, J.G.; Schmied, H.M.; Papadimitriou, L.; et al. Globally observed trends in mean and extreme river flow attributed to climate change. Science 2021, 371, 1159–1162. [Google Scholar] [CrossRef]
- Dethier, E.N.; Renshaw, C.E.; Magilligan, F.J. Rapid changes to global river suspended sediment flux by humans. Science 2022, 376, 1447–1452. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.K. A review on the adsorption of heavy metals by clay minerals, with special focus on the past decade. Chem. Eng. J. 2017, 308, 438–462. [Google Scholar] [CrossRef]
- Liu, R.X.; Lead, J.R.; Baker, A. Fluorescence characterization of cross flow ultrafiltration derived freshwater colloidal and dissolved organic matter. Chemosphere 2007, 68, 1304–1311. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.A.; Santschi, P.H.; Leppard, G.G.; West, M.M. Characterization of organic-rich colloids from surface and ground waters at the actinide-contaminated Rocky Flats Environmental Technology Site (RFETS), Colorado, USA. Colloid Surf. A-Physicochem. Eng. Asp. 2004, 244, 105–111. [Google Scholar] [CrossRef]
- Wilding, A.; Liu, R.X.; Zhou, J.L. Dynamic behaviour of river colloidal and dissolved organic matter through cross-flow ultrafiltration system. J. Colloid Interface Sci. 2005, 287, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.D.; Santschi, P.H. Composition and cycling of colloids in marine environments. Rev. Geophys. 1997, 35, 17–40. [Google Scholar] [CrossRef]
- Xu, H.C.; Houghton, E.M.; Houghton, C.J.; Guo, L.D. Variations in size and composition of colloidal organic matter in a negative freshwater estuary. Sci. Total Environ. 2018, 615, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Sun, X.; Luo, Z.; Chi, Q.; Zhu, G. Analysis of colloidal organic carbon content and its influencing factors in the waters of Taihu Lake in spring. China Environ. Sci. 2006, 26, 166–170. [Google Scholar]
- Klun, K.; Sket, P.; Falnoga, I.; Faganeli, J. Variation in Colloidal Organic Matter Composition and Aggregation in Coastal Waters (Gulf of Trieste, Northern Adriatic Sea). Geomicrobiol. J. 2015, 32, 609–615. [Google Scholar] [CrossRef]
- Stolpe, B.; Guo, L.D.; Shiller, A.M.; Hassellöv, M. Size and composition of colloidal organic matter and trace elements in the Mississippi River, Pearl River and the northern Gulf of Mexico, as characterized by flow field-flow fractionation. Mar. Chem. 2010, 118, 119–128. [Google Scholar] [CrossRef]
- Judy, J.D.; Kirby, J.K.; Farrell, M.; McLaughlin, M.J.; Wilkinson, S.N.; Bartley, R.; Bertsch, P.M. Colloidal nitrogen is an important and highly-mobile form of nitrogen discharging into the Great Barrier Reef lagoon. Sci. Rep. 2018, 8, 11. [Google Scholar] [CrossRef] [PubMed]
- Baken, S.; Moens, C.; van der Grift, B.; Smolders, E. Phosphate binding by natural iron-rich colloids in streams. Water Res. 2016, 98, 326–333. [Google Scholar] [CrossRef]
- Yang, B.; Lin, H.; Bartlett, S.L.; Houghton, E.M.; Robertson, D.M.; Guo, L.D. Partitioning and transformation of organic and inorganic phosphorus among dissolved, colloidal and particulate phases in a hypereutrophic freshwater estuary. Water Res. 2021, 196, 14. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.J.; Zhang, Q.; Lu, T.T.; Chen, J.Y.; Wei, Q.Q.; Chen, W.F.; Zhou, Y.M.; Qi, Z.C. Colloid-mediated transport of tetracycline in saturated porous media: Comparison between ferrihydrite and montmorillonite. J. Environ. Manag. 2021, 299, 10. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Li, Y. Effects of colloids on ammonia nitrogen release under different ion conditions in natural sediments of Lake Taihu, China. Environ. Sci. Pollut. Res. 2022, 29, 41455–41466. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.C.; Guo, L.D. Molecular size-dependent abundance and composition of dissolved organic matter in river, lake and sea waters. Water Res. 2017, 117, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.L.; Wang, H.L.; Li, Y.; Lin, L.; Hui, C.Z.; Gao, Y.; Niu, L.H.; Zhang, H.J.; Wang, L.F.; Wang, P.F.; et al. Bend-induced sediment redistribution regulates deterministic processes and stimulates microbial nitrogen removal in coarse sediment regions of river. Water Res. 2020, 170, 10. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, L.; Thamdrup, B.; Trimmer, M. Coupled nitrification and N2 gas production as a cryptic process in oxic riverbeds. Nat. Commun. 2021, 12, 8. [Google Scholar] [CrossRef]
- Guéguen, C.; Belin, C.; Dominik, J. Organic colloid separation in contrasting aquatic environments with tangential flow filtration. Water Res. 2002, 36, 1677–1684. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhao, L.; Gao, L.; Guo, L.D. Seasonal Variations in Molecular Size of Chromophoric Dissolved Organic Matter from Tile Lower Changjiang (Yangtze) River. J. Geophys. Res.-Biogeosci. 2021, 126, 17. [Google Scholar] [CrossRef]
- Belzile, C.; Guo, L.D. Optical properties of low molecular weight and colloidal organic matter: Application of the ultrafiltration permeation model to DOM absorption and fluorescence. Mar. Chem. 2006, 98, 183–196. [Google Scholar] [CrossRef]
- Ren, H.M.; Liu, H.J.; Qu, J.H.; Berg, M.; Qi, W.X.; Xu, W. The influence of colloids on the geochemical behavior of metals in polluted water using as an example Yongdingxin River, Tianjin, China. Chemosphere 2010, 78, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.H.; Buesseler, K.O.; Ripple, P.; Andrews, J.; Belastock, R.A.; Gustafsson, O.; Moran, S.B. Evaluation of two cross-flow ultrafiltration membranes for isolating marine organic colloids. Mar. Chem. 1998, 62, 117–136. [Google Scholar] [CrossRef]
- Stutter, M.I.; Langan, S.J.; Lumsdon, D.G.; Clark, L.M. Multi-element signatures of stream sediments and sources under moderate to low flow conditions. Appl. Geochem. 2009, 24, 800–809. [Google Scholar] [CrossRef]
- Baker, A. Fluorescence excitation-emission matrix characterization of some sewage-impacted rivers. Environ. Sci. Technol. 2001, 35, 948–953. [Google Scholar] [CrossRef]
- Baker, A.; Inverarity, R.; Charlton, M.; Richmond, S. Detecting river pollution using fluorescence spectrophotometry: Case studies from the Ouseburn, NE England. Environ. Pollut. 2003, 124, 57–70. [Google Scholar] [CrossRef] [PubMed]
- McKnight, D.M.; Boyer, E.W.; Westerhoff, P.K.; Doran, P.T.; Kulbe, T.; Andersen, D.T. Spectrofluorometric characterization of dissolved organic matter for indication of precursor organic material and aromaticity. Limnol. Oceanogr. 2001, 46, 38–48. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. Isme J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed]
- Richa, K.; Balestra, C.; Piredda, R.; Benes, V.; Borra, M.; Passarelli, A.; Margiotta, F.; Saggiomo, M.; Biffali, E.; Sanges, R.; et al. Distribution, Community Composition, and Potential Metabolic Activity of Bacterioplankton in an Urbanized Mediterranean Sea Coastal Zone. Appl. Environ. Microbiol. 2017, 83, 17. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Stegen, J.C.; Lin, X.J.; Fredrickson, J.K.; Konopka, A.E. Estimating and mapping ecological processes influencing microbial community assembly. Front. Microbiol. 2015, 6, 15. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Westerhoff, P.; Leenheer, J.A.; Booksh, K. Fluorescence excitation—Emission matrix regional integration to quantify spectra for dissolved organic matter. Environ. Sci. Technol. 2003, 37, 5701–5710. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.J.; Cao, B.D.; Wang, D.S.; Ma, T.; Xia, H.; Yu, D.H. Influence of wastewater sludge treatment using combined peroxyacetic acid oxidation and inorganic coagulants re-flocculation on characteristics of extracellular polymeric substances (EPS). Water Res. 2016, 88, 728–739. [Google Scholar] [CrossRef] [PubMed]
- Ifon, B.E.; Adyari, B.; Hou, L.Y.; Zhang, L.P.; Liao, X.; Peter, P.O.; Rashid, A.; Yu, C.P.; Hu, A.Y. Insight into variation and controlling factors of dissolved organic matter between urban rivers undergoing different anthropogenic influences. J. Environ. Manage. 2023, 326, 11. [Google Scholar] [CrossRef] [PubMed]
- Salve, P.R.; Lohkare, H.; Gobre, T.; Bodhe, G.; Krupadam, R.J.; Ramteke, D.S.; Wate, S.R. Characterization of Chromophoric Dissolved Organic Matter (CDOM) in Rainwater Using Fluorescence Spectrophotometry. Bull. Environ. Contam. Toxicol. 2012, 88, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.N.; Grybos, M.; Rabiet, M.; Deluchat, V. How do colloid separation and sediment storage methods affect water-mobilizable colloids and phosphorus? An insight into dam reservoir sediment. Colloid Surf. A-Physicochem. Eng. Asp. 2020, 606, 15. [Google Scholar] [CrossRef]
- Liu, J.T.; Ge, X.; Ye, X.X.; Wang, G.Z.; Zhang, H.M.; Zhou, H.J.; Zhang, Y.X.; Zhao, H.J. 3D graphene/delta-MnO2 aerogels for highly efficient and reversible removal of heavy metal ions. J. Mater. Chem. A 2016, 4, 1970–1979. [Google Scholar] [CrossRef]
- Soong, J.L.; Calderon, F.J.; Betzen, J.; Cotrufo, M.F. Quantification and FTIR characterization of dissolved organic carbon and total dissolved nitrogen leached from litter: A comparison of methods across litter types. Plant Soil 2014, 385, 125–137. [Google Scholar] [CrossRef]
- Zhao, Q.; Xie, F.X.; Zhang, F.F.; Zhou, K.; Sun, H.B.; Zhao, Y.J.; Yang, Q. Analysis of bacterial community functional diversity in late-stage shrimp (Litopenaeus vannamei) ponds using Biolog EcoPlates and PICRUSt2. Aquaculture 2022, 546, 15. [Google Scholar] [CrossRef]
- Niu, L.H.; Xie, X.D.; Li, Y.; Hu, Q.; Wang, C.; Zhang, W.L.; Zhang, H.J.; Wang, L.F. Effects of nitrogen on the longitudinal and vertical patterns of the composition and potential function of bacterial and archaeal communities in the tidal mudflats. Sci. Total Environ. 2022, 806, 13. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, C.; Zhang, W.L.; Lin, L.; Wang, L.F.; Niu, L.H.; Zhang, H.J.; Wang, P.F.; Wang, C. Response of bacterial community in composition and function to the various DOM at river confluences in the urban area. Water Res. 2020, 169, 11. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.T. Colloidal organic carbon in Huanghe, Changjiang and Qiantang River. Chin. Sci. Bull. 1998, 43, 915–917. [Google Scholar]
- Lee, B.J.; Hur, J.; Toorman, E.A. Seasonal Variation in Flocculation Potential of River Water: Roles of the Organic Matter Pool. Water 2017, 9, 14. [Google Scholar] [CrossRef]
- Jaffe, R.; Boyer, J.N.; Lu, X.; Maie, N.; Yang, C.; Scully, N.M.; Mock, S. Source characterization of dissolved organic matter in a subtropical mangrove-dominated estuary by fluorescence analysis. Mar. Chem. 2004, 84, 195–210. [Google Scholar] [CrossRef]
- Li, Q.; Li, H.; Chen, j.; Li, Y. Effect of natural colloid on ammonia nitrogen transport in different acidic and basic environment. Hydrogeol. Eng. Geol. 2014, 41, 131–137. [Google Scholar]
- Bo, W.; Hai-ming, L.I.; Xiao-yu, J.I.A.; Jin-lan, W.U. Effect of Natural Colloid on Ammonia Nitrogen Transport in Water-bearing Media. Enuivon. Sci. Technol. 2009, 32, 18–21. [Google Scholar]
- Lu, J.B.; Liu, H.J.; Zhao, X.; Jefferson, W.; Cheng, F.; Qu, J.H. Phosphate removal from water using freshly formed Fe-Mn binary oxide: Adsorption behaviors and mechanisms. Colloid Surf. A-Physicochem. Eng. Asp. 2014, 455, 11–18. [Google Scholar] [CrossRef]
- Gu, B.H.; Mehlhorn, T.L.; Liang, L.Y.; McCarthy, J.F. Competitive adsorption, displacement, and transport of organic matter on iron oxide: 1. Competitive adsorption. Geochim. Cosmochim. Acta 1996, 60, 1943–1950. [Google Scholar] [CrossRef]
- Gu, B.; Schmitt, J.; Chen, Z.; Liang, L.; McCarthy, J.F. Adsorption and desorption of natural organic matter on iron oxide: Mechanisms and models. Environ. Sci. Technol. 1994, 28, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Hornák, K.; Kasalicky, V.; Simek, K.; Grossart, H.P. Strain-specific consumption and transformation of alga-derived dissolved organic matter by members of the Limnohabitans-C and Polynucleobacter-B clusters of Betaproteobacteria. Environ. Microbiol. 2017, 19, 4519–4535. [Google Scholar] [CrossRef] [PubMed]
- Kasalicky, V.; Jezbera, J.; Hahn, M.W.; Simek, K. The Diversity of the Limnohabitans Genus, an Important Group of Freshwater Bacterioplankton, by Characterization of 35 Isolated Strains. PLoS ONE 2013, 8, e58209. [Google Scholar] [CrossRef] [PubMed]
- Kolton, M.; Erlacher, A.; Berg, G.; Cytryn, E. The Flavobacterium Genus in the Plant Holobiont: Ecological, Physiological, and Applicative Insights. In Microbial Models: From Environmental to Industrial Sustainability; Castro-Sowinski, S., Ed.; Springer: Singapore, 2016; pp. 189–207. [Google Scholar]
- Hubeny, J.; Korzeniewska, E.; Buta-Hubeny, M.; Zielinski, W.; Rolbiecki, D.; Harnisz, M. Characterization of carbapenem resistance in environmental samples and Acinetobacter spp. isolates from wastewater and river water in Poland. Sci. Total Environ. 2022, 822, 153437. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.M.; Liu, X.H.; Zhao, S.N.; Cui, B.S.; Bai, J.H.; Li, Z.J. Influence of the natural colloids on the multi-phase distributions of antibiotics in the surface water from the largest lake in North China. Sci. Total Environ. 2017, 578, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, M.F.; Zheng, K.K.; Wan, C.L.; Li, J. Promising carbon utilization for nitrogen recovery in low strength wastewater treatment: Ammonia nitrogen assimilation, protein production and microbial community structure. Sci. Total Environ. 2020, 710, 11. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.P.; Zhai, T.R.; Zhang, L.J.; Zhao, T.T.; Xing, Z.L.; Liu, H. Domestication and pilot-scale culture of mixed bacteria HY-1 capable of heterotrophic nitrification-aerobic denitrification. Bioresour. Technol. 2023, 384, 9. [Google Scholar] [CrossRef]
- Wang, X.; Wang, P.F.; Wang, C.; Chen, J.; Miao, L.Z.; Yuan, Q.S.; Liu, S.; Feng, T. Do bacterioplankton respond equally to different river regulations? A quantitative study in the single-dammed Yarlung Tsangpo River and the cascade-dammed Lancang River. Environ. Res. 2020, 191, 11. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.F.; Holmfeldt, K.; Hubalek, V.; Lundin, D.; Åström, M.; Bertilsson, S.; Dopson, M. Microbial metagenomes from three aquifers in the Fennoscandian shield terrestrial deep biosphere reveal metabolic partitioning among populations. ISME J. 2016, 10, 1192–1203. [Google Scholar] [CrossRef]
- Luef, B.; Frischkorn, K.R.; Wrighton, K.C.; Holman, H.Y.N.; Birarda, G.; Thomas, B.C.; Singh, A.; Williams, K.H.; Siegerist, C.E.; Tringe, S.G.; et al. Diverse uncultivated ultra-small bacterial cells in groundwater. Nat. Commun. 2015, 6, 6372. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.Y.; Li, Y.; Sun, X.X.; Adams, J.M.; Wang, L.F.; Zhang, H.J.; Chu, H.Y. More Robust Co-Occurrence Patterns and Stronger Dispersal Limitations of Bacterial Communities in Wet than Dry Seasons of Riparian Wetlands. mSystems 2023, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Fan, T.; Wang, S.; Yu, X.; Lu, A.; Wang, X.; Zhou, W.; Yuan, H.; Zhang, L. Seasonal changes driving shifts in microbial community assembly and species coexistence in an urban river. Sci. Total Environ. 2023, 905, 167027. [Google Scholar] [CrossRef]
- Dini-Andreote, F.; Stegen, J.C.; van Elsas, J.D.; Salles, J.F. Disentangling mechanisms that mediate the balance between stochastic and deterministic processes in microbial succession. Proc. Natl. Acad. Sci. USA 2015, 112, E1326–E1332. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.H.; Zhang, S.Q.; Meng, F.G. Large-sized planktonic bioaggregates possess high biofilm formation potentials: Bacterial succession and assembly in the biofilm metacommunity. Water Res. 2020, 170, 10. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Huang, W.; Yang, J.; Ran, Y.; Chen, D.; Yang, Y.; Wu, C.; Zhan, Y. Occurrence of colloid-bound endocrine-disrupting chemicals in the Pearl River, China. China Environ. Sci. 2015, 35, 617–623. [Google Scholar]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, L.; Chen, R.; Li, Y.; Chen, Y.; Zhang, W.; Zhang, H.; Wang, L. Colloidal Filterable Bacteria Enhance Ammonia Nitrogen Enrichment in River Colloids under Different Turbidity Conditions: Bacterial Diversity, Assembly Mechanism, and Nitrogen Transformation. Water 2024, 16, 1024. https://doi.org/10.3390/w16071024
Niu L, Chen R, Li Y, Chen Y, Zhang W, Zhang H, Wang L. Colloidal Filterable Bacteria Enhance Ammonia Nitrogen Enrichment in River Colloids under Different Turbidity Conditions: Bacterial Diversity, Assembly Mechanism, and Nitrogen Transformation. Water. 2024; 16(7):1024. https://doi.org/10.3390/w16071024
Chicago/Turabian StyleNiu, Lihua, Rui Chen, Yi Li, Yamei Chen, Wenlong Zhang, Huanjun Zhang, and Longfei Wang. 2024. "Colloidal Filterable Bacteria Enhance Ammonia Nitrogen Enrichment in River Colloids under Different Turbidity Conditions: Bacterial Diversity, Assembly Mechanism, and Nitrogen Transformation" Water 16, no. 7: 1024. https://doi.org/10.3390/w16071024
APA StyleNiu, L., Chen, R., Li, Y., Chen, Y., Zhang, W., Zhang, H., & Wang, L. (2024). Colloidal Filterable Bacteria Enhance Ammonia Nitrogen Enrichment in River Colloids under Different Turbidity Conditions: Bacterial Diversity, Assembly Mechanism, and Nitrogen Transformation. Water, 16(7), 1024. https://doi.org/10.3390/w16071024